1. Introduction
Carrot (
Daucus carota L.), from the family
Apiaceae, is a winter vegetable crop with high demand [
1]. It is a rich source of carotenoids, anthocyanins, dietary fiber, essential vitamins, and minerals [
2]. Root color and flavor are the most important qualitative factors in varietal selection [
3,
4]. The origin of this crop is considered to be Central Asia and is now produced globally with medicinal and nutritional values [
5,
1]. The worldwide production of carrot is increasing, which is estimated to be about 27.39 million tonnes per year, in an area of 9, 90 thousand hectares [
6]. In Pakistan, carrot ranks third among winter vegetables and was cultivated in an area of 14.32 thousand hectares, with an annual production of 241.91 thousand tonnes, during the years 2017–2018 [
7].
The anticipated demand for vegetable seeds in Pakistan stands at 5070 MT. However, merely 82 MT is accessible domestically, failing to meet the growers’ need for seeds [
8]. Due to the production of low-quality carrot seeds produced at a small level in Pakistan, a large quantity of the seeds available in the market is imported from other countries, due to which farmers have to pay high costs for the seeds [
9,
10]. Thus, to improve the production of carrot at the local level, the accessibility of quality seeds is crucial [
11]. The yield and quality of carrot seeds can be improved by adopting improved practices, from seed planting to harvesting and postharvest treatment of seeds [
10].
Umbel position (primary, secondary, or tertiary umbels) and size also influence the production of good-quality carrot seeds, e.g., vigor and germination [
12,
13]. Primary and secondary umbel seeds are of higher quality than tertiary umbel seeds [
14]. Seed quality differs among various umbel orders because the seed-setting process begins with primary umbels, followed by the flowering of secondary umbels, while tertiary umbels are still emerging. Consequently, seeds in tertiary umbels mature much later, coinciding with the shattering of seeds from primary umbels [
15,
16,
17], particularly in hot subtropical environments [
18]. Carrot seed quality can be improved by altering the umbel order, i.e., the suppression of tertiary umbels [
13]. Recently, the exogenous application of auxins has been reported not only to reduce tertiary umbels in carrot but also to improve the quality of seeds produced on auxin-treated plants [
18].
Sustainable agriculture is fundamentally important because it offers the potential to meet future agricultural needs of fast-growing populations, which is beyond the capabilities of traditional agriculture [
19]. The utilization of plant-growth-promoting bacteria, either rhizosphere bacteria or endophytes, is gaining popularity in various parts of the world [
20,
21]. Plant-growth-promoting rhizobacteria (PGPR) are beneficial bacteria that colonize plant roots [
22] and various kinds of bacteria, such as
Pseudomonas,
Azospirillum,
Azotobacter,
Klebsiella,
Enterobacter [
23,
24],
Alcaligenes,
Arthrobacter,
Burkholderia,
Bacillus [
25], and
Serratia [
26], lead to increased plant growth via different mechanisms like phosphate solubilization [
27], biological nitrogen fixation (BNF), siderophore production [
28,
29], and the production of phytohormones such as gibberellins, cytokinins and indole-3-acetic acid (IAA) [
30]. PGPR can also modulate phytohormone levels in plant tissues [
31].
Auxins play an important role in the growth and developmental processes of plants, such as cell division and enlargement, apical dominance control, flowering, fruiting, photosynthetic capacity, and the effective translocation of assimilates [
32,
33]. Tryptophan is a remarkable amino acid commonly found in root exudates. It is the main precursor molecule for the biosynthesis of indole-3-acetic acid in bacteria and higher plants [
34]. Indole-3-acetic acid (IAA) is one of the most extensively studied and abundant types of auxins in plants. The production of IAA is affected by variations in different rhizobacteria species and strains, culture conditions, growth stages, and substrate availability [
35]. The endogenous hormonal level of the plants can be altered with the exogenous application of phytohormones or their precursors [
36].
It has been established that the biosynthesis of IAA by plant-associated bacteria with L-tryptophan (L-TRP) substrate improves the growth and yield of various crops [
37,
38,
39,
40]. Bacterial phytohormone production by rhizobacteria using L-tryptophan (L-TRP) as a stimulant from root exudates is responsible for increasing auxin production, which enhances root hair density and improves seed germination and plant growth [
41]. Similarly, Khan et al. [
42] stated that the endophyte
B.
subtilis LK14 improved shoot and root biomass and chlorophyll (a and b) contents in tomato by producing the highest amount of IAA. Previously, Noor et al. [
18] reported that the exogenous application of synthetic auxins can potentially alter the umbel order in carrot via apical dominance control. However, the role of microbial auxin in controlling apical dominance has not been studied with regard to seed production in carrot.
Primarily, the objective of this study was to assess the effectiveness of bacterial strains in changing umbel order to enhance the production of high-quality carrot seeds. While numerous researchers have explored carrot root crops extensively, there has been no prior research published on the manipulation of umbel order through microbial application in carrot seed crops. More specifically, the present study is designed to evaluate the potential of plant-growth-promoting bacteria and L-tryptophan (an auxin precursor) for the growth enhancement and quality seed production of carrot under natural field conditions.
3. Results
Plant height showed significant differences for foliar treatments during both years (
Table 1). Among foliar treatments, maximum plant height (127.2 and 118.2 cm) was recorded for MN54+L-TRP during both years, i.e., a 30% increment in plant height. The minimum height of the plant (94.4 and 93.8 cm) was observed for PsJN, but it was not significantly different from PsJN+L-TRP for Year I (
Table 1). Adventitious root weight was significantly affected by the foliar treatments. The highest weight of adventitious root (1.56 and 1.82 g) on the 30th day after steckling plantation was recorded for MN34+L-TRP during both years. Moreover, the weight of adventitious roots was 42% higher than that of untreated plants (
Table 1). The maximum adventitious root weight (1.96 and 2.50 g) on the 45th day after steckling plantation was observed in plants treated with MN34+L-TRP during Years I and II, respectively, with 45% more root weight recorded for MN34+L-TRP than control (
Table 1).
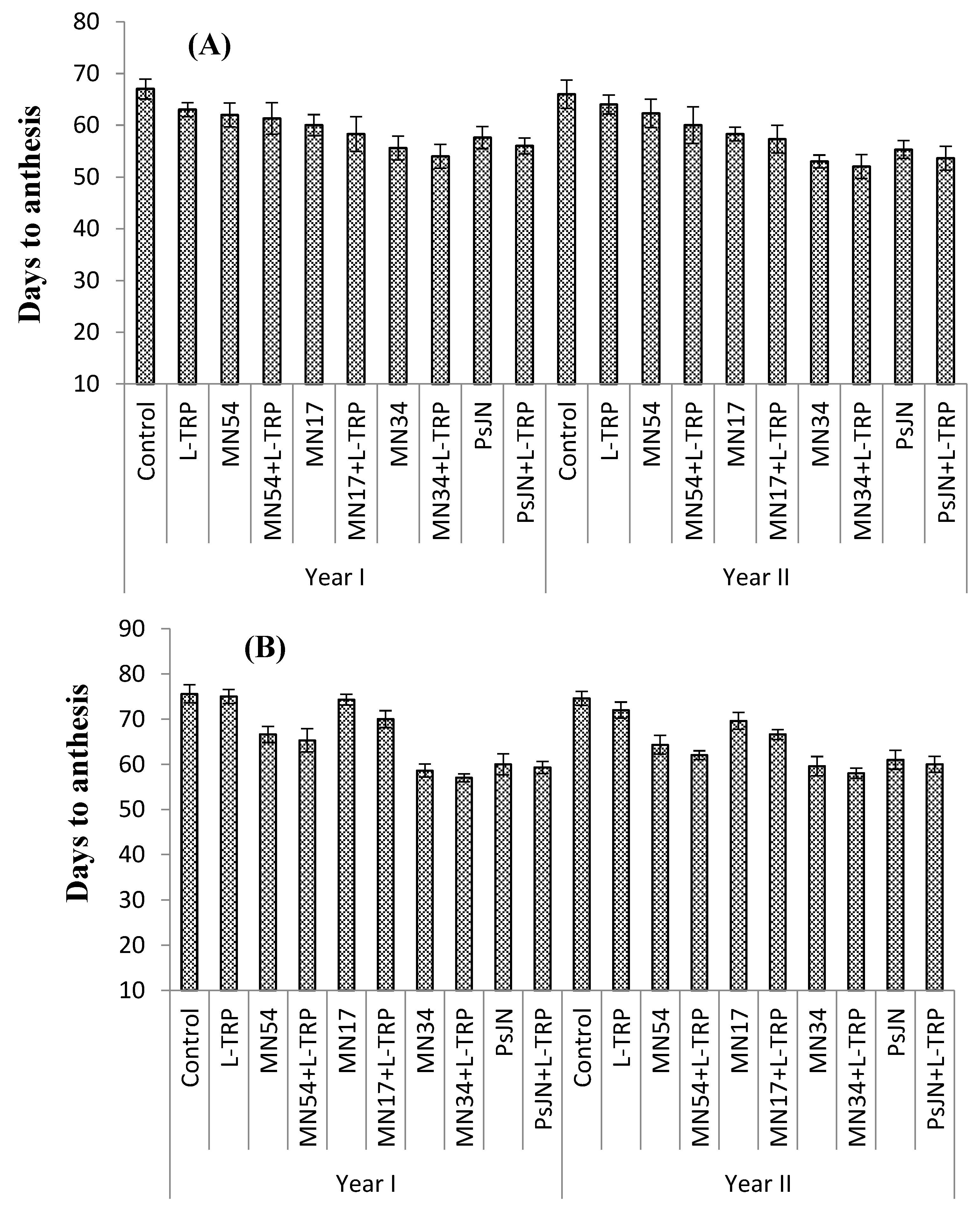
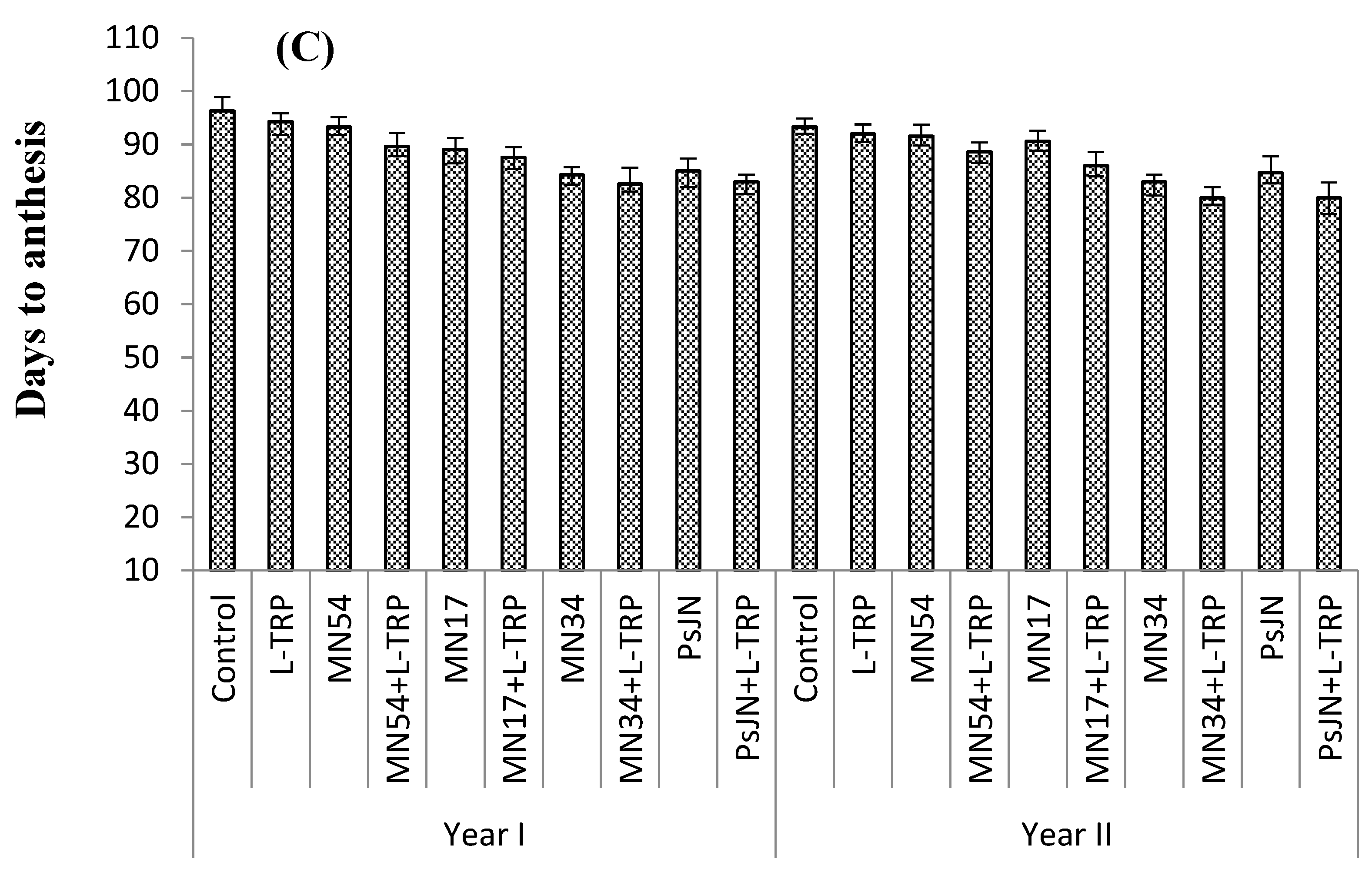
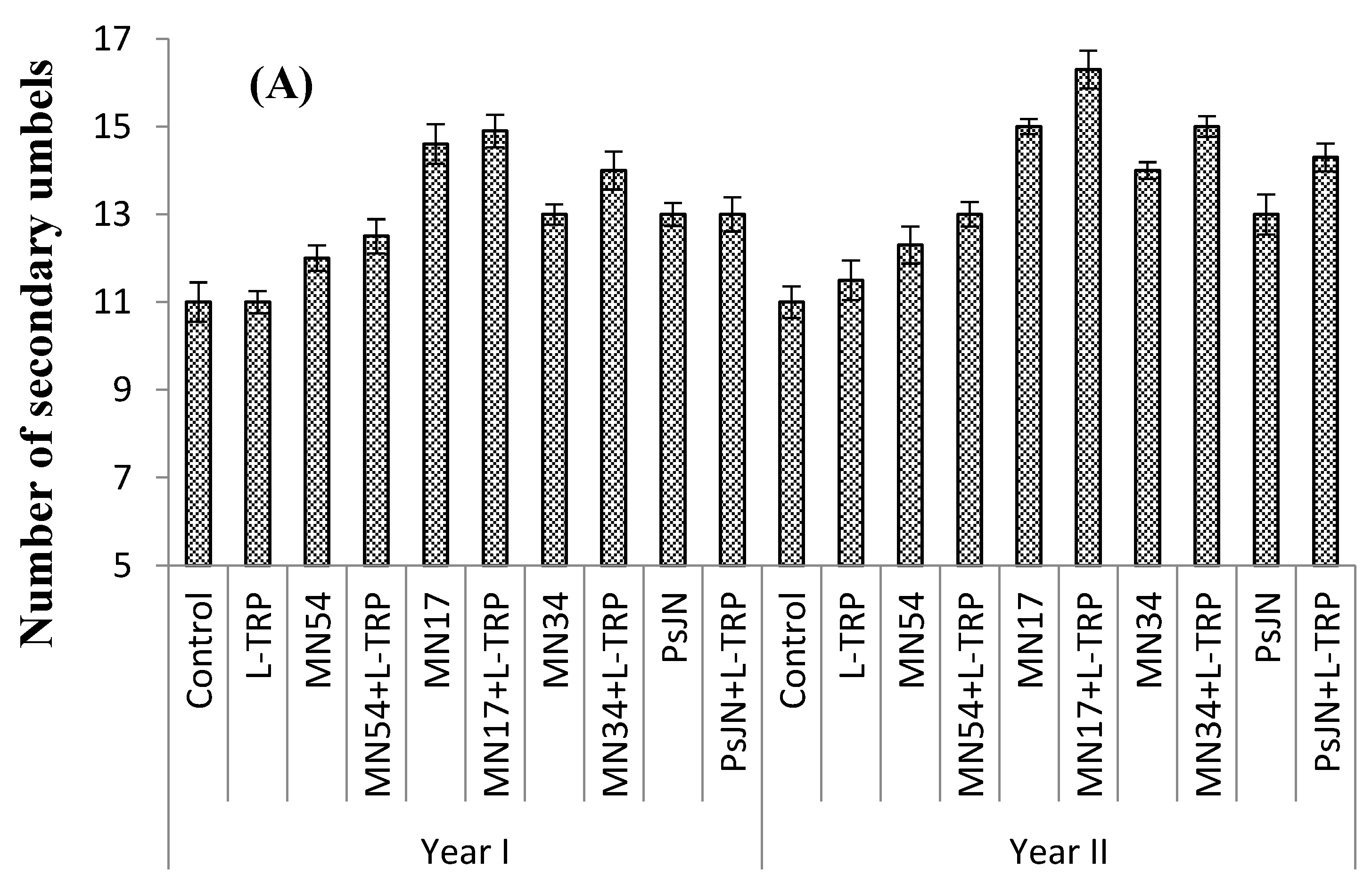
Foliar treatments had a positive effect on number of days taken to anthesis in all types of umbels (
Figure 1A–C). MN34 and L-tryptophan induced earliest flowering in primary (54.1 and 52.0 days), secondary (57.0 and 58.6 days), and tertiary umbels (82.6 and 80.0 days), which took 20%, 23%, and 14% less time than the control for all umbel orders, respectively, during both years. The maximum days taken to anthesis in primary (67.0 and 66.0 days), secondary (75.6 and 74.6 days), and tertiary umbels (96.3 and 93.3 days) were observed for the control (untreated) plant (
Figure 1).
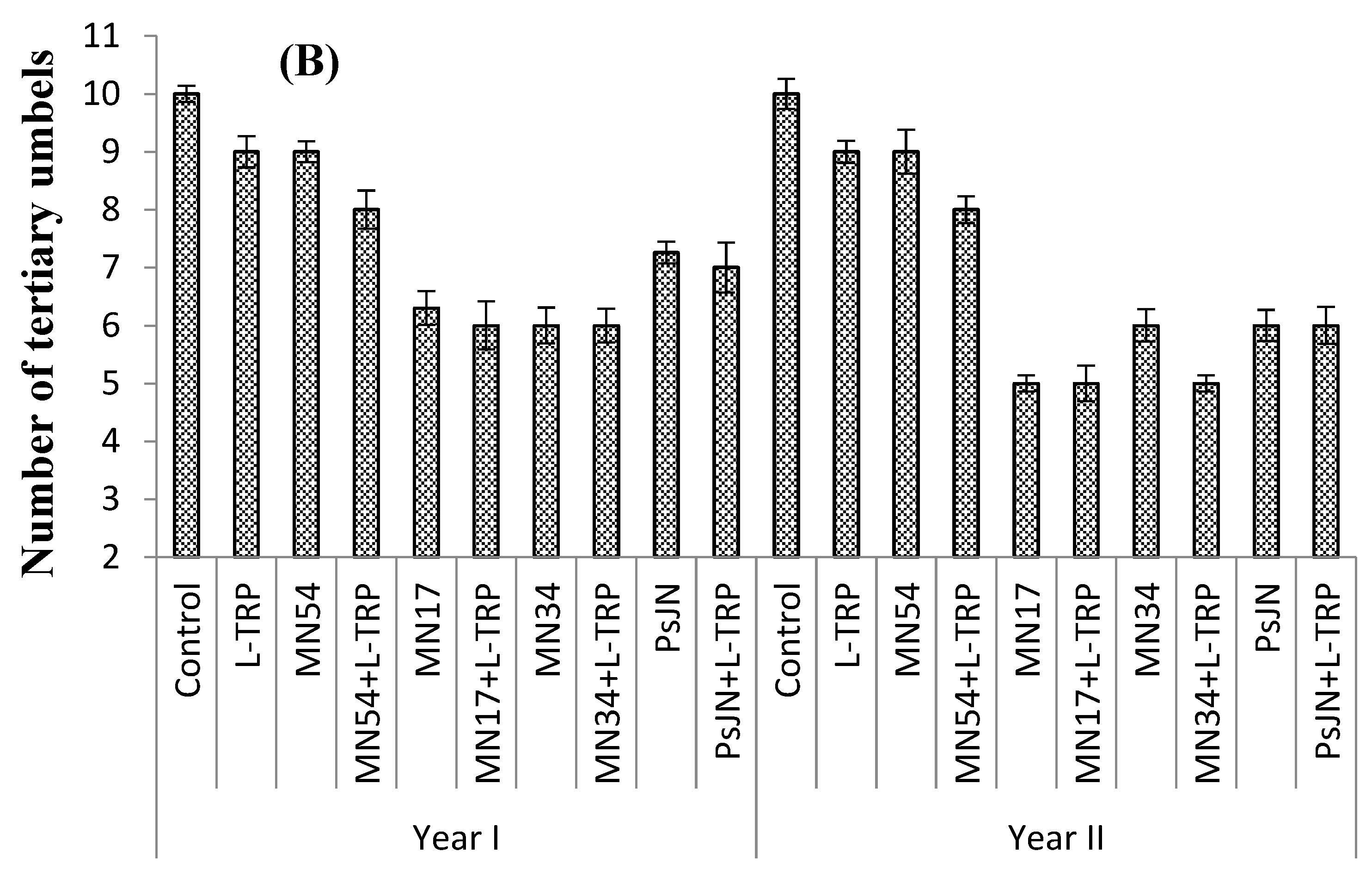
The maximum number of secondary umbels per plant (15.0 and 16.0 umbels) was documented with MN17+L-TRP, during Years I and II, respectively, with 40% more secondary umbels per plant recorded in samples treated with MN17+L-TRP, compared with the control (untreated) plants. However, the minimum number of secondary umbels per plant (11 umbels) was found in control during both years (
Figure 2A). The least number of tertiary umbels per plant (6.00 and 5.00) was recorded for MN17+L-TRP and MN34+L-TRP, with 45% fewer tertiary umbels per plant recorded compared with the untreated plants during Years I and II. The maximum number of tertiary umbels per plant (10.0 umbels) was recorded for control (untreated) during both years but was statistically similar to L-TRP and MN54 during both years (
Figure 2B).
Seed weights per umbel differed significantly among the foliar treatments and were significantly higher with L-TRP (
Table 2). PsJN and L-tryptophan produced higher seed weight from primary (13.7 g and 12.8 g), secondary (56.0 g and 55.6 g), and tertiary umbels (13.1 and 12.7 g), during Years I and II, with 44%, 41%, and 42% higher seed weight recorded in all umbel orders, i.e., primary, secondary, and tertiary umbels, respectively, compared with respective control plants (
Table 2). The effects of foliar treatments on seed quality were more variable. A higher value of 1000-seed weight was observed in primary (5.9 and 5.7 g), secondary (5.3 and 5.2 g), and tertiary umbels (2.74 and 2.51 g) for plants sprayed with PsJN and L-TRP, indicating 42%, 34%, and 35% increases in the umbel orders, respectively, compared with the control (untreated) plants during both years (
Table 2).
Germination percentage was highest in the primary (98.6% and 99.6%), secondary (95.3% and 97.0%), and tertiary umbels (66.2% and 68.0%) seeds of plants treated with PsJN+L-TRP, and it increased up to 16%, 15% and 44%, respectively, for seeds in all umbel orders, compared with the seeds obtained from untreated plants during both years (Year I and II, respectively) (
Table 3). Higher vigor index was observed in the primary (1112.9 and 1150.0), secondary (1085.8 and 1111.0), and tertiary umbels (480.8 and 493.7) of plants treated with PsJN+L-TRP than untreated plants, with 44%, 43%, and 46% higher vigor indices than the seeds obtained from untreated plants for all umbel orders, i.e., primary, secondary and tertiary umbels, respectively, during both years (
Table 3).
The lowest conductivity of leachates of harvested seeds, after 24 h of soaking seeds in deionized water, was recorded in the seeds from the primary (427.6 and 372.6 μS/cm), secondary (486.3 and 470.0 μS/cm), and tertiary umbels (1080.3 and 948.7 μS/cm) in response to foliar application of PsJN+L-TRP, i.e., 34.5%, 29.8% and 35.5% lower leachates, respectively, than in the seeds from the corresponding umbels of untreated plants during both years (
Table 4). Malondialdehyde contents of the seeds harvested from the primary (1.16 and 1.22 µmols/g Fw), secondary (1.20 and 1.15 µmols/g Fw), and tertiary umbels (3.59 and 3.43 µmols/g Fw) were lowest for plants treated with PsJN+L-TRP, i.e., 36.1%, 40.6%, and 30.1% lower than seeds obtained from all umbel orders of control plants, respectively during both years (
Table 4).
The activity of antioxidant enzymes in the harvested seeds of all types of umbels was also positively affected by the foliar application of plant-growth-promoting bacterial strains. The superoxide dismutase activity of seedlings raised from the primary (431.8 and 453.8 U kg
−1 protein), secondary (420.4 and 435.1 U kg
−1 protein), and tertiary umbels (254.3 and 266.8 U kg
−1 protein) was highest in plants treated with PsJN+L-TRP, increasing up to 46.5%, 44.1%, and 41.5%, respectively, compared with untreated (control) plants (
Table 5). A higher value of peroxidase activity of seedlings raised from the primary (1193.1 and 1256.6 U kg
−1 protein), secondary (1142 and 1085.7 U kg
−1 protein), and tertiary umbels (600.1 and 615.9 U kg
−1 protein) was observed for plants treated with PsJN+L-TRP, while it was found to be minimum for control, i.e., the plants treated with PsJN+L-TRP exhibited 43.5%, 41.5% and 44% higher peroxidase activity than the untreated (control) plants in all umbel orders, respectively (
Table 5).
The highest catalase activity of seedlings raised from the primary (601.3 and 616.3 U kg
−1 protein), secondary (588.1 and 600.8 U kg
−1 protein), and tertiary umbels (377.7 and 365.1 U kg
−1 protein) was recorded for plants treated with PsJN+L-TRP, with 49%, 47.9%, and 46% increase in catalase activity compared with untreated (control) plants (
Table 6). The total antioxidant content in the seedlings of the primary (99.5% and 99% inhibition) secondary (96.1% and 93.9% inhibition), and tertiary umbels (72.9% and 63.6% inhibition) were recorded highest for plants treated with MN34+L-TRP, increasing up to 32.4%, 40.7%, and 47.7%, respectively, compared with untreated (control) plants in all umbel orders. The least value for total antioxidant content was recorded for the control (
Table 6).
The total phenolic content in the seedlings of primary (295.5 and 300.7 mg GAE 100 g
−1), secondary (264.5 and 274.2 mg GAE 100 g
−1), and tertiary (189.0 and 202.9 mg GAE 100 g
−1) umbels of plants treated with PsJN+L-TRP were increased up to 42.3%, 35.3%, and 45.5%, respectively, compared with the control (
Table 7).
The bacterial strain colonized the rhizosphere and interior of carrot roots and shoots (
Table 8). When augmented with L-TRP, PsJN resulted in enhanced colonization as compared to sole application in the rhizosphere and plant tissues. During the first year of inoculation treatment, the colonization values were 5.70 × 10
5 CFU g
−1 in rhizosphere soil, 2.5 × 10
5 CFU g
−1 in the root interior, and 5.1 × 10
5 CFU g
−1 in the shoot tissue. Similarly, in the second year, the L-TRP-treated rhizosphere soil CFU g
−1 value was 6.8 × 10
5, whereas in the root interior, it was 3.71 × 10
5, and in the shoot interior, it was 7.8 × 10
5. However, more CFU of the inoculant strain g
−1 dry weight were found from the rhizosphere and the interior of roots and shoots treated with L-TRP.
Data in
Table 9 show that PsJN produced auxin (IAA equivalents) without L-TRP application; however, IAA equivalents significantly increased when the medium was applied with L-TRP. In the second year, PsJN produced maximum IAA equivalents of 12.7 μg mL
−1 when L-TRP was added.
4. Discussion
Plant-growth-promoting bacteria colonize the rhizosphere and interior tissues of host plants and promote the growth and yield of crops through different mechanisms [
53]. In the current study, we found that auxin production by strain
B. phytofirmans PsJN enhances plant growth via aggressive colonization.
Understanding the impact of agricultural technologies on soil microbiome and enzymatic activity is essential for the quantitative and qualitative assessment of agricultural production [
54]. In the present study, it was found that PsJN inoculation improved carrot growth, physiology, root biomass, biochemical activities, and yield traits as compared to the uninoculated control (
Table 1,
Table 2 and
Table 3 and 5–7). Many bacterial mechanisms have been suggested, and hormone production is known as the most important mechanism in regulating plant growth and development [
55]. Among phytohormones, indole-3-acetic acid (IAA) produced by bacteria is a major naturally occurring phytohormone that is also exogenously applied to improve plant growth [
40,
56,
57,
58]. Axillary buds are inhibited by IAA, and shoot apical dominance is induced [
59,
60]. Microorganisms with tryptophan supplementation in culture media increase IAA production; however, in the absence of L-tryptophan (L-TRP), a small number of auxins are produced [
21,
40,
61,
62]. Therefore, in the present study, the impact of PGPB with or without L-TRP on auxin-induced apical dominance and alteration of umbel order was studied in carrot.
The foliar application of plant-growth-promoting bacteria (PGPB) positively influenced various growth traits, yield, and seed-quality attributes. The foliar application of MN54+L-TRP increased the height of carrot seed plants, which is in line with the findings of Hassan and Bano [
63], who reported that the inoculation of
Pseudomonas moraviensis and
Bacillus cereus with tryptophan addition increased the plant height of wheat grown in pots and fields. An increase in plant height with PGPB and L-tryptophan may be responsible for enhanced IAA availability, which induces cell division and cell elongation [
64,
65].
Auxin-producing bacterial strains stimulated root growth in this study, as evidenced by the increased weight of adventitious roots in response to foliar applications of MN34 and PsJN with L-tryptophan. Naveed et al. [
40] also reported that maize seed inoculation with PsJN supplemented with L-TRP notably augmented root biomass compared with the uninoculated control. Moreover, a positive link between IAA production and root elongation in response to PGPR in lentils has also been reported, which reflects our results [
66]. This increase in root growth might also have enhanced nutrient uptake, one of the roles attributed to PGPB [
64,
65]. This supposition is based on a 30% and 27% increase in seed yield from secondary umbels, while a 26% and 22% increase was observed in 1000-seed weight from secondary umbels in response to the foliar application of
Burkholderia phytofirmans PsJN and
Pantoea sp. MN34 along with L-TRP. PsJN has been reported to increase endogenous IAA in maize along with higher nitrogen and phosphorus contents and photosynthetic rate, ultimately resulting in higher biomass [
40]. This increase in seed yield with concomitant improvement in yield can also be explained by the findings of Sun et al. [
67], who envisaged that the overexpression of the auxin response factor 19 gene caused improvement in the seed size and yield of
Arabidopsis thaliana and
Jatropha curcas. Very recently, Iqbal et al. [
68] revealed microbial inoculation-mediated enhancement in the growth and yield of canola through auxin production and nutrient uptake. They further reported that the coinoculation of
Bacillus sp. MN54 and
Piriformospora indica significantly enhanced root growth, most notably through high rhizosphere auxin production, which consequently enhanced biomass and nutrient uptake of canola. Enhanced yield and yield-related components in canola and peanut have also been attributed to increased water and nutrient absorption, followed by improved photosynthesis, leading to more assimilation and thus an increase in seed yield [
69,
70].
Seeds in primary and secondary umbels reach maturity under mild temperatures, whereas during flowering, seed setting, and seed maturation in tertiary umbels, temperatures remain elevated (ranging from 33 to 42.5 °C). Consequently, this has negative impacts on seed yield and quality, including germination and the weight of a thousand seeds. Various researchers have documented this irregular seed maturation across different umbels [
13,
16,
17]. To obtain high-quality seeds, the suppression of tertiary umbels seems to be an option but seems impossible if performed manually. The use of auxins has been suggested in a previous study that resulted in better suppression of tertiary umbels in carrot [
18]. Therefore, in this study, the impact of PGPB application on the suppression of tertiary umbels per plant was evaluated. The foliar application of plant-growth-promoting bacterial strains, i.e.,
Enterobacter sp. MN17 and
Pantoea sp., with and without L-TRP, significantly reduced the number of tertiary umbels compared with control in carrot seed crop; reductions of 41–47% and 48–50% were observed in the tertiary umbel count using MN17 with and without L-TRP, compared with 37–39% and 42–43% reductions using MN34 with and without L-TRP, respectively. The reduction in tertiary umbels by these PGPB may be because of the shoot apical dominance induced by indole-3-acetic acid, and apical auxin might have inhibited tertiary umbels (which appear as side branches on the secondary branches of umbels, possibly due to an increase in the endogenous level of auxin) [
59,
71]. This apical dominance phenomenon in carrot seed crops had also been observed when exogenously applied auxins caused significant suppression of tertiary umbels and an increase in the number of secondary umbels per plant [
18]. Furthermore, the foliar application of MN34+L-TRP and PsJN+L-TRP induced the earliest anthesis in primary, secondary, and tertiary umbels. Reduced time for the onset of flowering was observed in response to the inoculation of strain MN17 in maize cultivars [
21].
The results revealed that the exogenous application of PsJN and MN34 along with L-TRP significantly improved the germination and seedling growth of carrot seeds harvested from primary, secondary, and tertiary umbels. Previously, treatments with plant-growth-promoting bacteria were found to increase germination percentage, seedling vigor, emergence, plant stand, and root and shoot growth in various agronomic and horticultural crops [
21,
72,
73]. IAA-producing rhizobacteria triggered faster germination and induced a higher vigor index, which is an indicator of the state of the health of seedlings and ultimately the productivity of crops [
74]. This high germination and vigor index may be due to the better production and metabolism of auxin by plant-growth-promoting bacteria (PGPB) in the presence of L-TRP, which stimulates cellular division [
75].
The electrical conductivity test measures the amount of electrolyte leakage from seeds during imbibition and thus reflects damage to seed membranes. An increase in conductivity has been found to be correlated with a decrease in seed quality. The leakage of electrolytes from small and physiologically immature seeds is due to the reorganization of membrane components and conformational changes occurring in cell membranes [
76]. The level of lipid peroxidation, expressed as malondialdehyde content, has also been used as an indicator of damage to cell membranes caused by free radicals [
77]. Hence, lower MDA content indicates enhanced antioxidant potential, providing improved tolerance to oxidative stress [
78]. The lowest electrical conductivity and MDA content of seeds from primary, secondary, and tertiary umbels were found in response to the foliar application of selected bacteria strains (PsJN and MN34) with L-TRP, suggesting that the seeds from these treatments were physiologically better than untreated seeds. Moreover, the findings of this study also revealed that the activity of the investigated antioxidant enzymes, the total antioxidant content, and the total phenolic content in seedlings increased in response to all plant-growth-promoting bacteria, particularly with PsJN and L-TRP. Enzymatic activity is the result of accumulated enzyme action as well as proliferating microorganisms [
54]. Previously, researchers [
79,
80] observed enhanced activities of different ROS-scavenging enzymes in PGPB-inoculated tomato and potato plants possibly because of the detoxification of reactive oxygen species by PGPB.
A successful microbial inoculant colonized the external and/or internal part of plant tissues and established a compatible interaction with the host, in addition to persisting in the soil against microorganisms living in the environment through its rhizocompetence traits [
81]. Plant-growth-promoting bacterial inoculants colonize the plant initially, but their persistence over time is not guaranteed. Bacterial invasions in the plant body are difficult, and to improve their efficacy, plant-growth-promoting bacteria were applied three times to ensure their proper invasion in the plant body. Measuring the persistence of microbial inoculants in soil poses technical difficulties, as the inoculant needs to be identified. The tracking and monitoring of the persistence of plant-growth-promoting bacteria released in the environment need to be studied to understand their behavior in soil and determine which factors influence their survival under various conditions. In the present study, the total bacterial count was measured from the rhizosphere using a culture-dependent approach (
Table 9). The growth promotion and quality carrot seed production in the present study were most probably because of the survival of the inoculant strain and its L-TRP-mediated auxin product in the plant rhizosphere. Overall, the MN-34 and PsJN strains along with L-TRP improved growth and quality of carrot seed production.




 ,
, 







{kind=link}
{kind=link}
{kind=link}
{kind=link}