3.1. Marketable Yield and Its Components
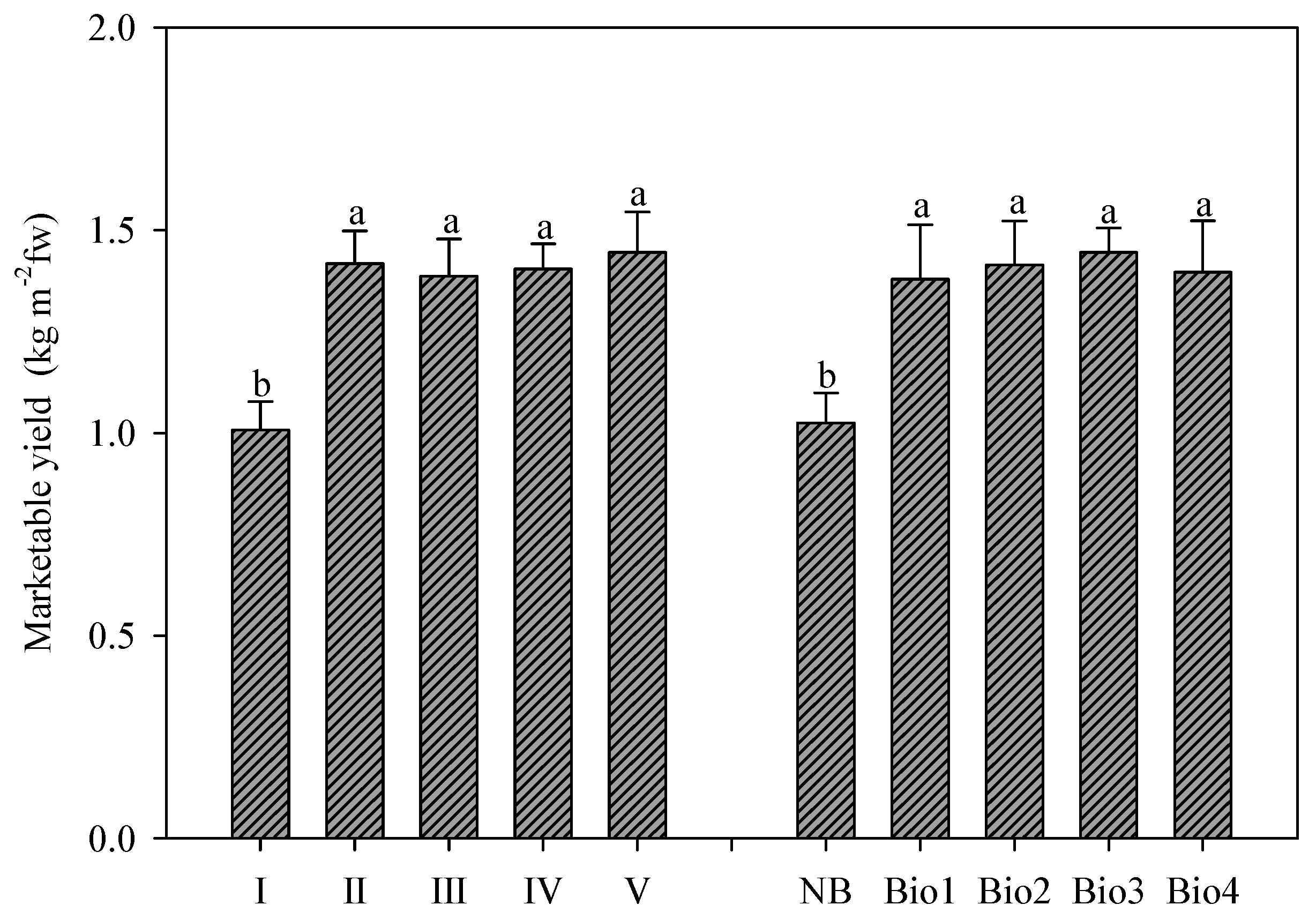
The marketable yield of rocket was significantly affected by the number of harvests (
Figure 2). Notably, after the second harvest, no significant differences were recorded, since only the first harvest was significantly lower than the successive ones (1.00 vs. 1.41 kg m
−2, mean value of II–V harvests) (
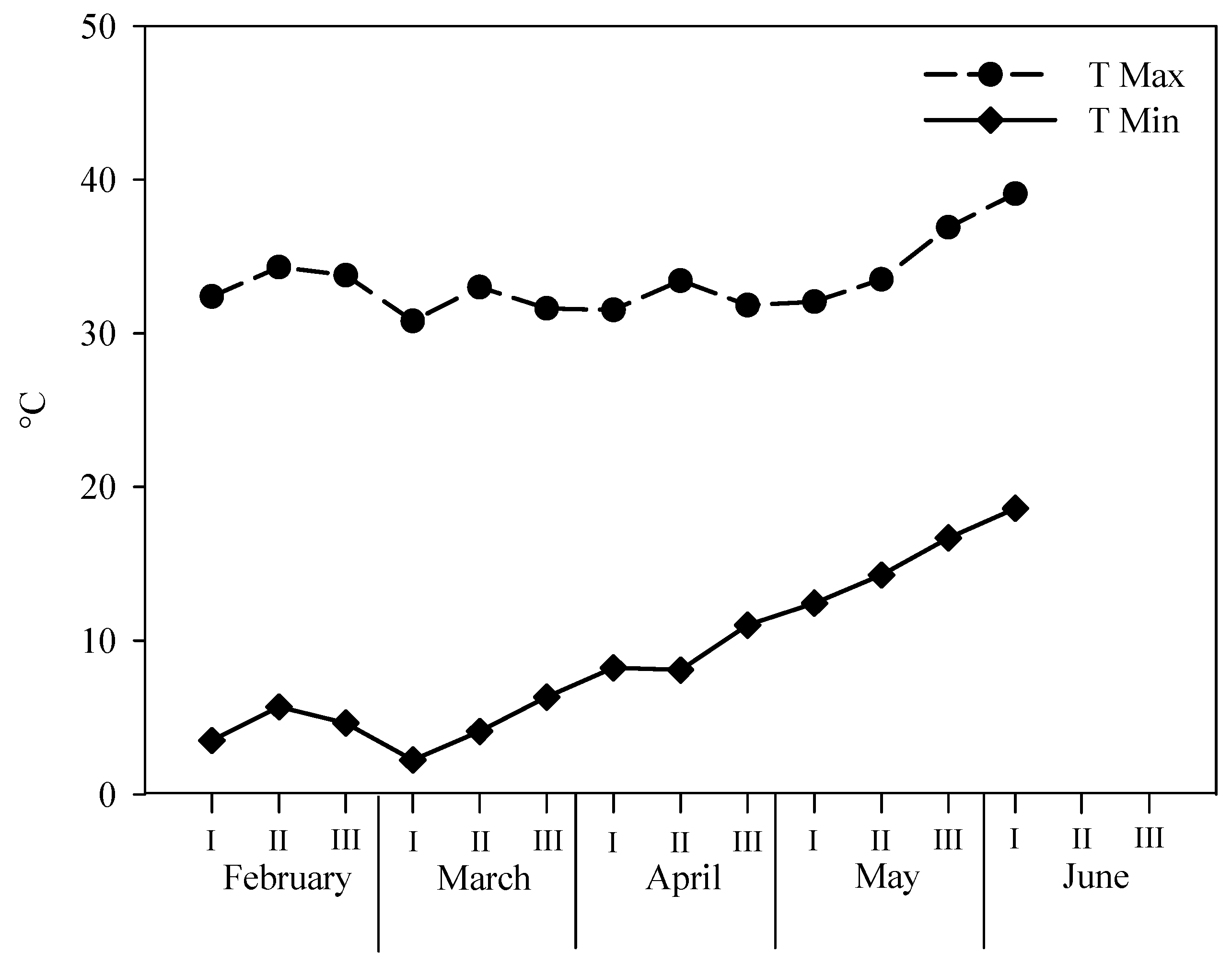
Figure 1). Our results indicate that rocket plants could be subjected to multiple harvests considering the short growth cycle of the plant and the short time interval between harvests, thus allowing prolonged growing periods and increased total fresh yields. The low fresh yield recorded in the first cutting of our study could be attributed to the low temperatures that prevailed in this period (beginning of February to mid-March) which resulted in a slow growth rate, as indicated by the long growth cycle of the first cutting (49 days compared to 17–22 for the rest of the harvests). The increasing temperatures throughout the rest of the growing period, especially the increase in Tmin, resulted in faster plant growth and higher fresh biomass production. In the study of Petropoulos et al. [
21], it was also suggested that the fresh yields of separate cuttings may significantly differ from each other, while the growing period (autumn–winter and winter–spring) may also affect the fresh yield of the different cuttings due to different temperature and light (light intensity and photoperiod) regimes. Furthermore, Abdalla [
34], who also applied multiple harvest in rocket plants grown in a floating system during the January–March growing period, suggested that five cuttings were possible. However, when compared to the practice in which plants were replaced with new ones after harvest, the authors did not record any difference in the total fresh yield between the two cropping systems despite the different number of growth cycles implemented (three growth cycles in the single harvest system and five growth cycles in the multiple harvest system) [
34]. In contrast to our study, Corrado et al. [
23] and Carillo et al. [
35] suggested that the first harvest yield was higher than the second one for basil (
Ocimum basilicum) and lettuce (
Lactuca sativa) plants, respectively, while Di Mola et al. [
36] reported similar results for wild rocket plants cultivated between April and June. This contradiction could be partly attributed to different growing periods and consequently to different air temperatures throughout the experiments, as well as to differences in the growth cycle duration and harvesting stage between the species [
15,
37]. The effect of growing conditions on fresh yield was also noted in the study of Di Mola et al. [
38], who performed a similar experiment with six successive harvests in wild rocket plants during the autumn–spring period (November to May) and suggested higher yields at the later harvests conducted in April and May, regardless of the biostimulant treatment.
On the other hand, the application of biostimulants had a profound effect on the fresh yield, since all the tested products elicited an increase compared to the untreated control (+37.5% on average), but no significant differences were observed among the various biostimulants implemented on rocket plants (
Figure 2). The positive effects of biostimulants on the growth and yield of rocket plants have been previously reported in the literature, which revealed that biostimulatory products of varied composition had a beneficial effect. For example, Candido et al. [
24] suggested that the use of an azoxystrobin-based biostimulant may increase the marketable yield of wild rocket plants (
Diplotaxis tenuifolia L. DC) after the first crop cycle, while positive effects on yield were also recorded for biostimulants that contained protein hydrolysates obtained from legumes, tropical extracts,
Trichoderma harzianum,
Moringa oleifera extracts and Azoxystrobin [
14,
39,
40,
41,
42,
43]. Additionally, Di Mola et al. [
38] suggested a significant increase in fresh yield in wild rocket plants treated with a biostimulant that contained seaweed and alfalfa extracts and amino acids, especially for harvests performed in late spring. Apart from the biostimulant product composition, the application dose may also have an effect on plant growth and yield, suggesting that further research is needed in order to fine-tune this practice for commercial application [
34].
The results regarding the dry matter (DM) content are presented in
Table 5. After the third harvest, the leaf dry matter showed lower values than the first two harvests did, with the lowest value of 8.1% being registered for the third harvest. The second harvest was characterized by the highest DM value of 10.9% and was significantly higher than all the other harvests (
Table 5). Moreover, the leaf DM was not significantly different among the different biostimulant treatments, except for the case of Bio4 which exhibited significantly lower DM compared to the untreated control where the highest overall value was recorded (
Table 5). The differences in the DM content of leaves between different harvestings has been also reported in leafy vegetables such as basil [
23], lettuce [
35] and rocket [
42]. According to the literature, the application of biostimulants may have a varied effect on leaf dry matter content depending on the biostimulatory product. For example, Di Mola et al. [
38] suggested that the application of a biostimulant that contained alfalfa and seaweed extracts and amino acids resulted in a decrease in the DM of rocket leaves compared to that of the untreated plants, regardless of the nitrogen application rate, while they also recorded significant differences between successive harvests. In another study, the application of legume-derived protein hydrolysates and/or tropical plant extracts on baby rocket leaves increased the amount of water per cm
2 of leaf area and consequently decreased the DM content [
13,
36]. In contrast, Giordano et al. [
42] suggested that the effect of biostimulants on DM content of perennial wall rocket leaves depends on the harvesting date when successive harvests are implemented and that biostimulants may increase DM content after the first harvesting. Similar results were reported by Caruso et al. [
40], who also suggested a positive effect of biostimulants on the DM content of perennial wall rocket, while they also detected a significant effect of the growing period on the same parameter. Therefore, it should be noted that such inconsistencies in literature reports could be associated not only with the varied composition of biostimulants but also with differences in the growing conditions, since according to Tuncay et al. [
44], dry matter content in rocket leaves may be significantly affected by the growing month.
The number of leaves m
−2 showed a parabolic trend; thus, it increased until the fourth harvest and then a rapid decrease was recorded. The latter was significantly higher than the first two and the fifth harvests, while no significant differences were recorded from the third harvest (
Table 5). This finding indicates that growing conditions may affect the number of leaves, since the increasing temperatures and the improvement in lighting conditions after the first harvests were associated with a higher number of leaves until the fourth harvest. It is interesting to point out that the lower number of leaves m
−2 in the fifth harvest was counterweighed by the higher average leaf weight, thus achieving a yield equal to that of the other harvests (see
Table 5 and
Figure 2). This difference in leaf morphology at the last harvest (less and larger leaves) compared to the preceding ones could be also attributed to the high temperatures and long photoperiods which may induce flowering and the formation of larger and longer leaves [
45]. This finding is in agreement with the results of Corrado et al. [
23], who also reported that the higher number of leaves was not associated with the highest fresh leaf weight and specific leaf area in basil plants that were harvested twice. Additionally, Petropoulos et al. [
21] suggested that the number of leaves in
Cichorium spinosum plants grown in the winter–spring period may increase after the first harvest and then show a decrease in subsequent harvests, while the same trends were observed in three Genovese basil cultivars [
20] and lettuce plants [
35,
46]. However, the authors associated the higher number of leaves with higher yields, which was not the case in our study, probably due to the different species tested. On the other hand, the different biostimulants elicited a 27.8% mean increase in leaf number compared to the NB plants, but only Bio1 and Bio3 were significantly different from the control (NB) treatment (
Table 5). As for the average leaf weight (ALW), it was only influenced by the successive harvests, with biostimulants showing no significant effects compared to the control treatment (
Table 5). In particular, the average leaf weight was higher in the second and fifth harvest than in the first and fourth harvests, whereas no significant differences were recorded between the second and third harvest. According to the literature reports, the application of biostimulants is associated with an increased number of leaves without any significant effects on total yield or the average leaf weight [
38,
39]
3.2. Color Parameters, SPAD Index, and Mineral Content of Rocket Leaves
Differences in the color parameters of rocket leaves in this experiment were only dictated by successive harvests and not by the different biostimulant applicationa (
Table 6). The lightness (L*) of rocket leaves showed a mean value of 42.1, where the significant lowest value was recorded in the second harvest, and the highest ones were recorded at the last two harvests and at the first one. The green intensity was lower (higher a* parameter values) at the second and third harvest (
Table 6), being not significantly different from the values recorded at the first and fourth harvest, whereas the lowest overall value was recorded at the last harvest. Similarly, the b* parameter reached the highest value (yellowish color) at the fifth harvest, although it was not significantly different from that at the fourth harvest. Finally, the hue parameter showed the lowest value at the last harvest, being not significantly different from that of the fourth harvest, whereas Chroma was the highest at the last harvest, but not significantly different from that at the first and fourth harvest. (
Table 6). Our results indicate that for most of the color parameters (except for lightness), the last harvest was clearly discriminated from the rest of the harvest, showing either the highest or the lowest overall values. Di Mola et al. [
38] also reported a significant effect of the harvesting date on the green intensity (a*) of wall rocket leaves, whereas lightness (L*) and b* parameters were not affected. Moreover, Ciriello et al. [
47] recorded significant differences between two harvestings for most of the color parameters (except for b* and Chroma values) in Genovese basil genotypes. In contrast, Bantis et al. [
15], who studied the effect of multiple harvests in spinach and rocket plants, did not observe significant differences in the color parameters of both species when compared to the control plants of which a single harvest was carried out. These contradictions could be attributed to the different growing conditions throughout the experiment and especially to the increasing temperatures.
Regarding the biostimulant application effect, our results are in agreement with those reported by Visconti et al. [
48], who suggested that the application of
Trichoderma did not affect color parameters in rocket leaves, while Consentino et al. [
49] reported that only the green intensity increased (lower a* values) in the leaves of celery plants due to animal- or plant-derived protein hydrolysates. Similarly, Giordano et al. [
42] and Caruso et al. [
39] did not observe significant effects on the color parameters of wall rocket after the application of tropical plant extracts and legume-derived protein hydrolysates. In contrast to our study, Caruso et al. [
40] suggested that the single and/or combined application of biostimulants affected the leaf color of rocket plants, especially the combined application of
Trichoderma and protein hydrolysates which significantly improved all the tested parameters (L*, a*, and b*). Additionally, Di Mola et al. [
13] reported a significant effect of seaweed extracts and protein hydrolysates on the color parameters of baby leaf lettuce. These contradictions in the literature suggest that biostimulant composition, growing conditions and the genotype are important for defining their effects on the visual quality of leafy vegetables, as indicated by color parameters.
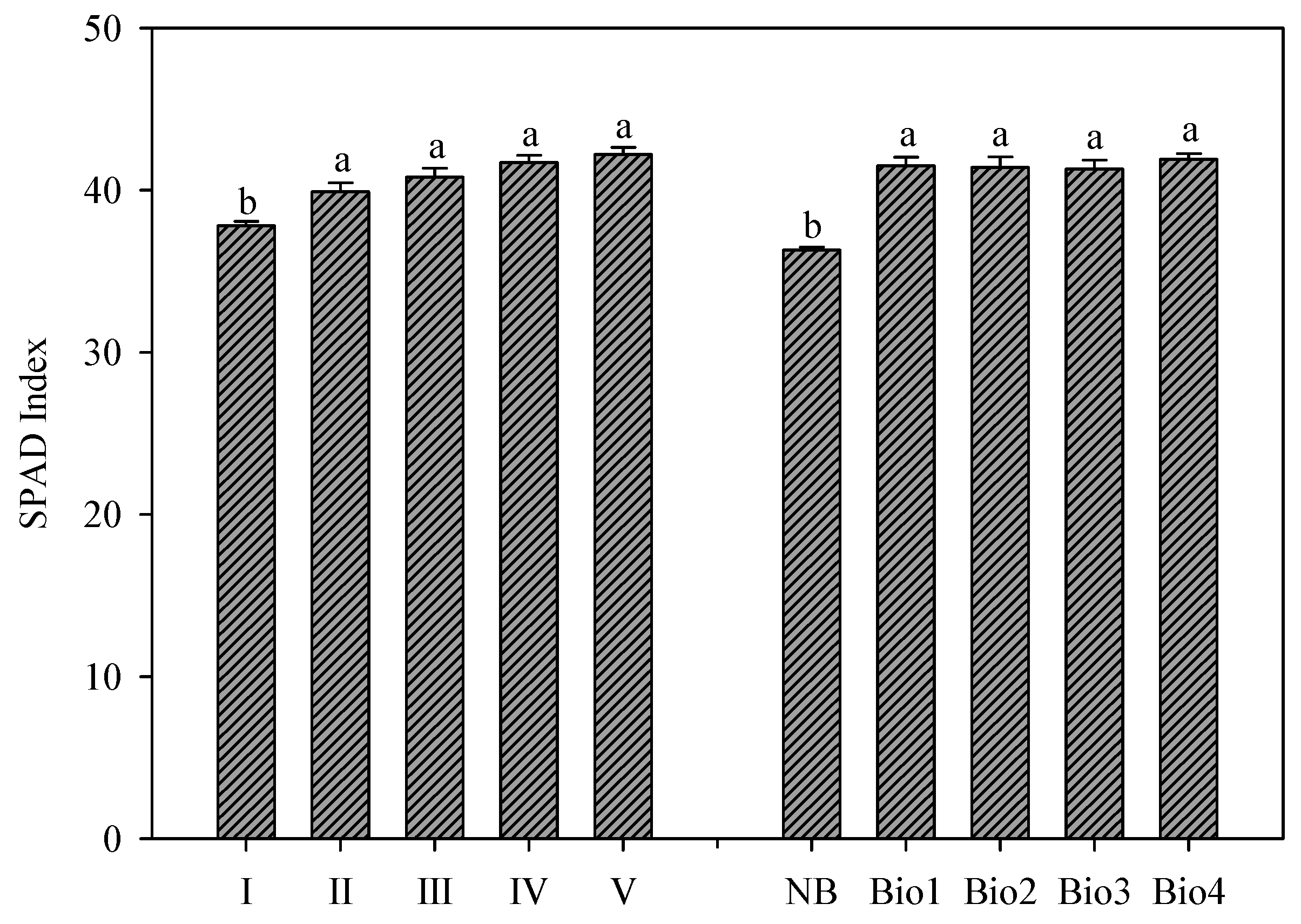
The SPAD index ranged from 37.8 at the first harvest to 42.2 at the last harvest, without significant differences between the successive harvests after the first one (a mean value of 41.2;
Figure 3). Similarly to the marketable yield, the SPAD index was higher in all treated plants with biostimulants with an average increase of 14.4% compared to the NB plants, and no significant difference between the biostimulant treatments (
Figure 3). The increasing trends in the SPAD index when successive harvesting is implemented have been also confirmed by Alexopoulos et al. [
50] in wild leafy vegetables. Similarly, several studies have recorded a positive effect of biostimulants on the SPAD index values of rocket and other leafy vegetables compared to those of the untreated ones, while they also associated the increased SPAD index with increased crop yields due to the more efficient function of the photosynthetic apparatus [
12,
13,
38,
39,
42,
51,
52]. In contrast, Formisano et al. [
20] did not record any differences in SPAD values when basil plants were subjected to multiple harvests, while Visconti et al. [
48] did not suggest any effects for
Trichoderma application in lettuce and spinach plants. Considering that literature reports have associated the SPAD index value in leafy vegetables with various agronomic practices (e.g., fertilization, irrigation regime, saline irrigation, etc. [
47,
53,
54], it could be that such inconsistencies in research data could be attributed to differences in growing conditions that may conceal the effects of biostimulant application.
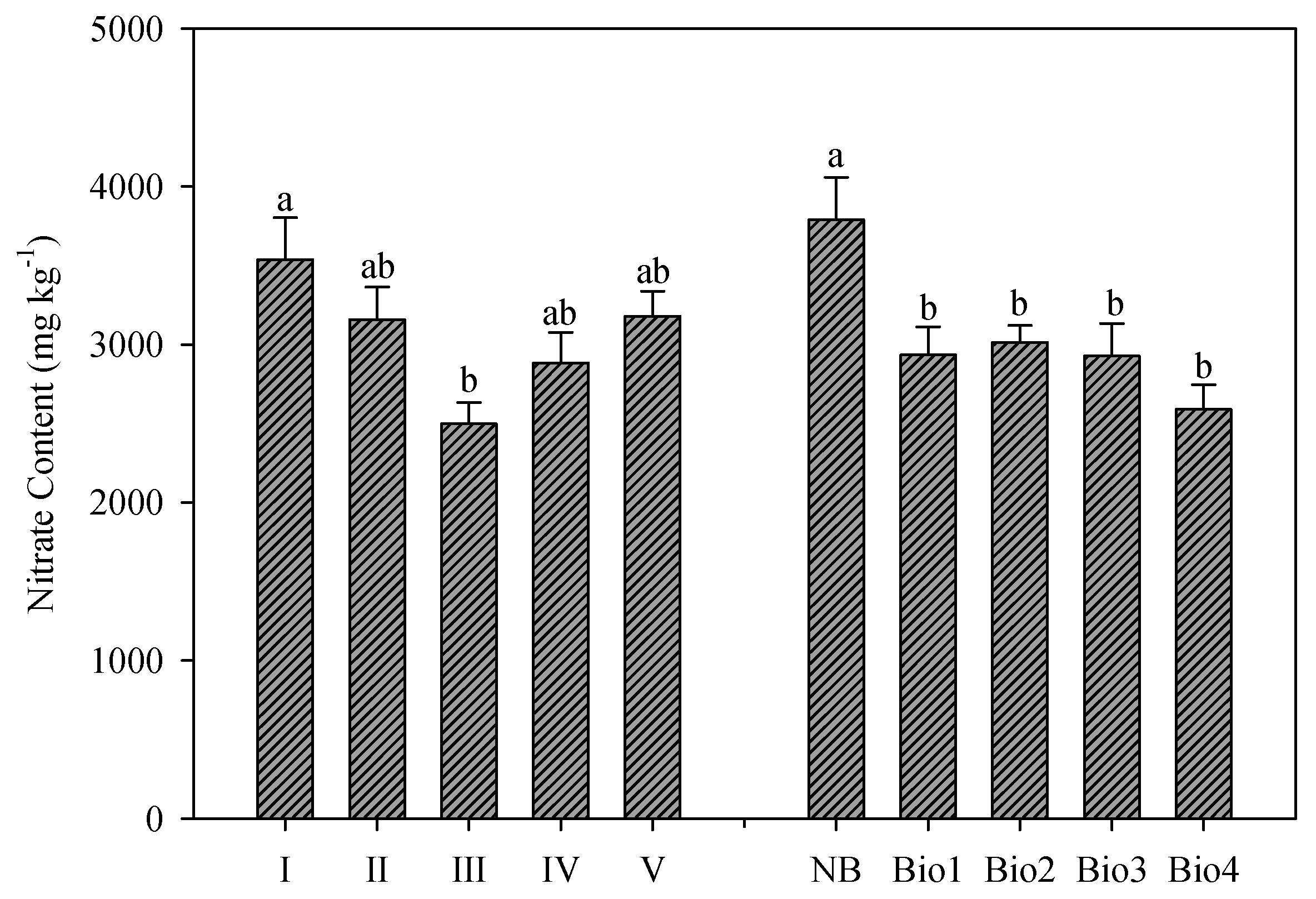
Nitrate content in rocket leaves showed a decreasing trend after the second harvest, with the third harvest recording the lowest overall value and then an increase was observed until the last harvest. The highest nitrate content was measured in the first harvest, being significantly higher only from the third harvest. In any case, nitrate content was 20.80% higher in the first harvest compared to the average values of the successive harvests (2929 vs. 3538 mg kg
−1;
Figure 4). The observed trends could be attributed to increasing temperatures, which may increase the uptake and assimilation of nutrients and thus result in decreased nitrate content (up to the third harvest), whereas the increasing nitrate content after the third harvest could be explained by the involvement of nitrates in osmoregulation mechanisms due to the increased water requirements of plants associated with high temperatures [
21]. According to Bonasia et al. [
17], nitrate content fluctuation in wild rocket plants could be expected depending on the genotype, the cultivation system and the growing period, while successive harvesting in basil plants resulted in higher total nitrogen and nitrate content in the second cutting (mid-June) compared to the first one (mid-April) [
23]. Similarly, Ciriello et al. [
47] reported a contrasting effect of successive harvesting on nitrate content in basil leaves depending on the cultivar, thus highlighting the genotypic differences in the nitrogen assimilation of the species, while the harvesting time within the day may also affect the measured nitrate content [
16]. The genotypic effect on nitrate accumulation was also noted by Bantis et al. [
15], who suggested that successive harvesting may increase nitrate content in rocket leaves, while no effects were recorded in the case of spinach. Furthermore, Masclaux-Daubresse et al. [
55] suggested that the formation of new leaves may induce the remobilization of nitrates to expanding leaves and therefore result in increasing nitrate content, as observed in our study after the third harvest where despite the increasing temperatures nitrate accumulation was observed.
As suggested by the manufacturers, all biostimulants limited the nitrate accumulation in rocket leaves. In particular, all the used biostimulants did not exhibit significant differences between them and registered a mean value of 2866 mg kg
−1, which was about 24.3% lower compared to the nitrate content measured in the untreated (NB) plants (
Figure 4). According to Ertani et al. [
56], the use of biostimulants that contain protein hydrolysates may increase nitrogen assimilation and decrease nitrate accumulation, while similar results were reported for caidrine-based biostimulants [
57]. However, contrasting results regarding the effects of biostimulants on nitrate accumulation are also reported in the literature. Therefore, Caruso et al. [
36] noted that the application of
Trichoderma and protein hydrolysates on wall rocket resulted in increased nitrate content compared to the control treatment, while Rouphael et al. [
12] suggested a variable effect depending on the biostimulant composition with seaweed and herbal extracts and vegetal oils inducing nitrate accumulation in spinach plants compared to the untreated plants or plants treated with legume-derived protein hydrolysates. In contrast, El-Nakhel et al. [
4] recorded an increase in the nitrate content of spinach plants for protein hydrolysates obtained from legumes, a difference that could be attributed to the different sowing dates, genotypes tested and growing systems compared the study of Rouphael et al. [
12]. Similar increasing trends were also recorded by Di Mola et al. [
13] in the case of lettuce plants treated with seaweed extracts and protein hydrolysates, as well as by Di Mola et al. [
36] for baby rocket leaves subjected to tropical plant extracts and protein hydrolysates. All of these results highlight the variable response of crops to biostimulant application since they indicate that several other factors are implicated including, the genotype, the growing conditions, the cultivation practices and so forth.
The harvesting time and the application of biostimulants had a significant effect on leaf mineral content (
Table 7). All the mineral elements studied in this experiment (K, Mg, Ca, P, Mg and S) exhibited a concentration decrease in the second and third harvest, while the highest overall value was recorded in the last (fifth) harvest (except for the case of P). In particular, Mg concentration decreased significantly at the third harvest compared with the first harvest, while S concentration decreased significantly at the second and third harvest compared with the first harvest. After the fourth harvest, the content of all the elements increased again and reached similar or higher values than the first harvest values. Moreover, the biostimulant applications elicited an increase in the concentration of all the minerals, except for the case of Ca where the application of biostimulants had no significant effect (
Table 7). The observed increase was significant for K, Mg and P content, only when biostimulants Bio2, Bio3 and Bio4 were applied, whereas Bio1 only significantly increased P concentration compared to the control. In addition, S content was significantly increased only by the Bio2 and Bio4 treatments.
Similarly to our study, Giordano et al. [
42] highlighted the significant single and combinatory effect of biostimulant products on the mineral composition of rocket plants harvested at different dates, although these authors did not study the effect of the harvesting date. On the other hand, Formisano et al. [
20] recorded significant differences in the P and Mg content of basil leaves subjected to successive harvesting, whereas K content remained unaffected. Additionally, Caruso et al. [
39] suggested the significant effect of the growing period and biostimulant application on the mineral composition of rocket plants, with the growing period having an impact on K, S, Mg and Ca content, while biostimulant application increased P and Ca content compared to the control (untreated plants). According to the literature, biostimulant application may induce mineral accumulation in leafy vegetables, as in the case of K and Mg [
12,
49] or P, K, Mg, Ca and S [
51] in spinach and celery leaves, P and Mg in rocket leaves [
40], K, Ca and Mg in perennial wall rocket leaves [
12], and P in lettuce plants [
52]. Rouphael et al. [
12] suggested that such positive effects could be associated with hormonal-like effects, including improved root architecture, which facilitates mineral uptake and translocation, and improved transportation of macronutrients through cell membranes. Moreover, Fiorentino et al. [
41] highlighted the varied response of different species (lettuce and rocket) to the same
Trichoderma-based biostimulants in terms of mineral composition, a finding which indicates crop specificity.




 ,
, 








{kind=link}
{kind=link}
{kind=link}
{kind=link}