
Current Acquaintance on Agronomic Biofortification to Modulate the Yield and Functional Value of Vegetable Crops: A Review

 ,
, 
 ,
,  , ,
, ,  ,
, 

Abstract
1. Introduction
2. Agronomic Biofortification
3. Direct and Indirect Effects of the Most Common Microelements Used in Vegetables Biofortification Programs
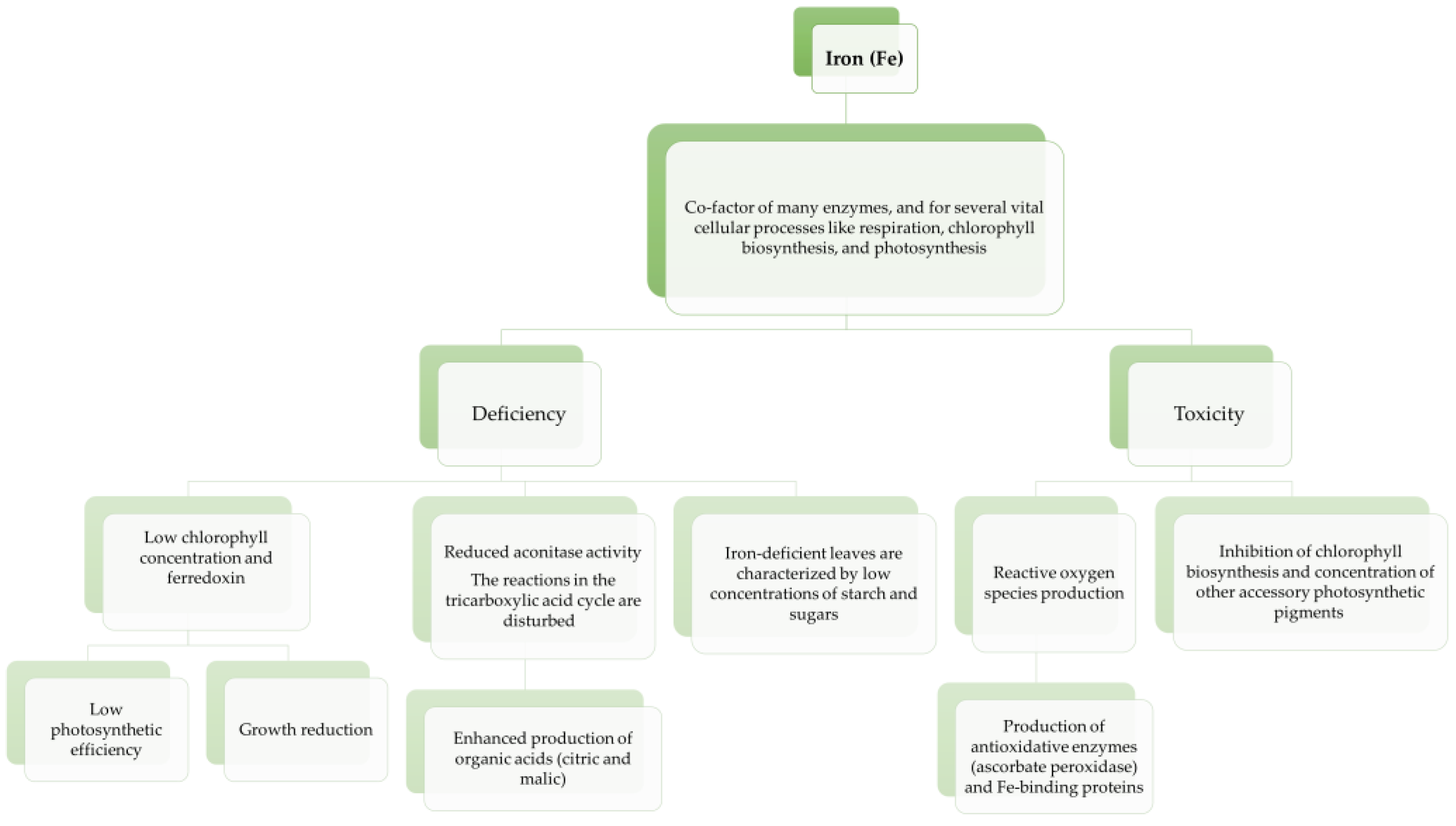
3.1. Iron (Fe)
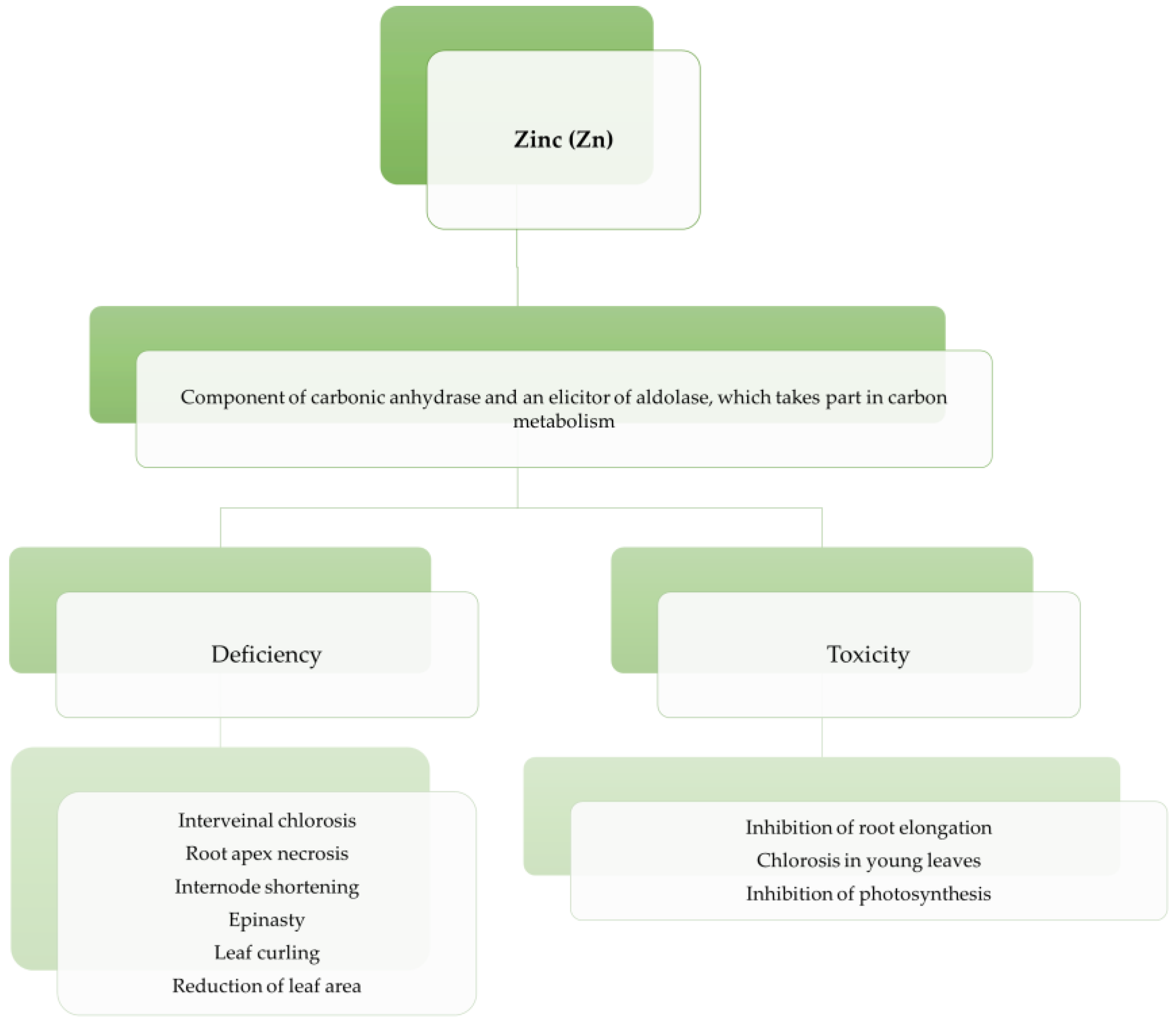
3.2. Zinc (Zn)
3.3. Iodine (I)
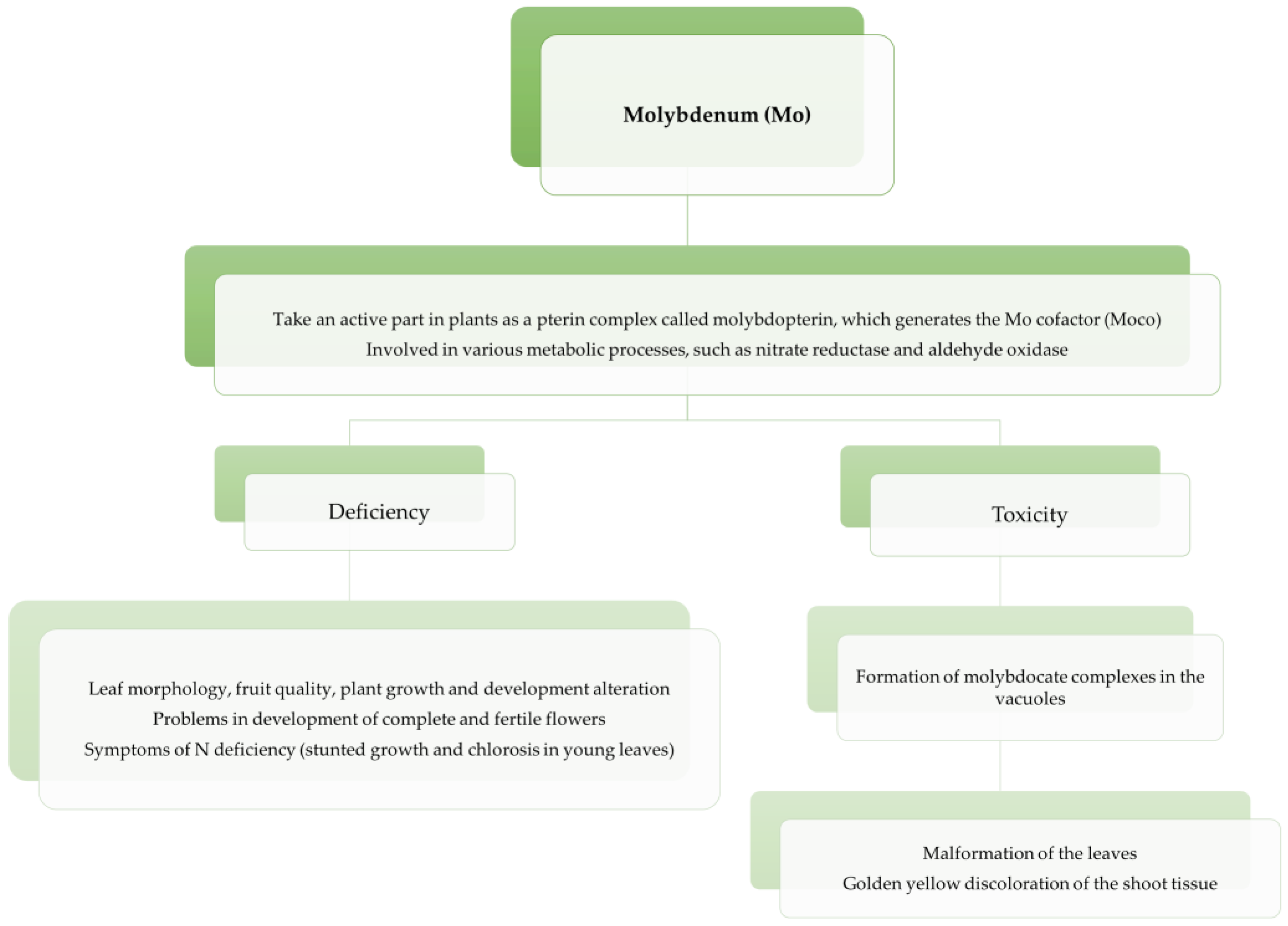
3.4. Molybdenum (Mo)
3.5. Selenium (Se)
3.6. Silicon (Si)
3.7. Joint Biofortification of Vegetables
3.8. Microgreens Biofortification
4. Effects of Biofortification on Human Health
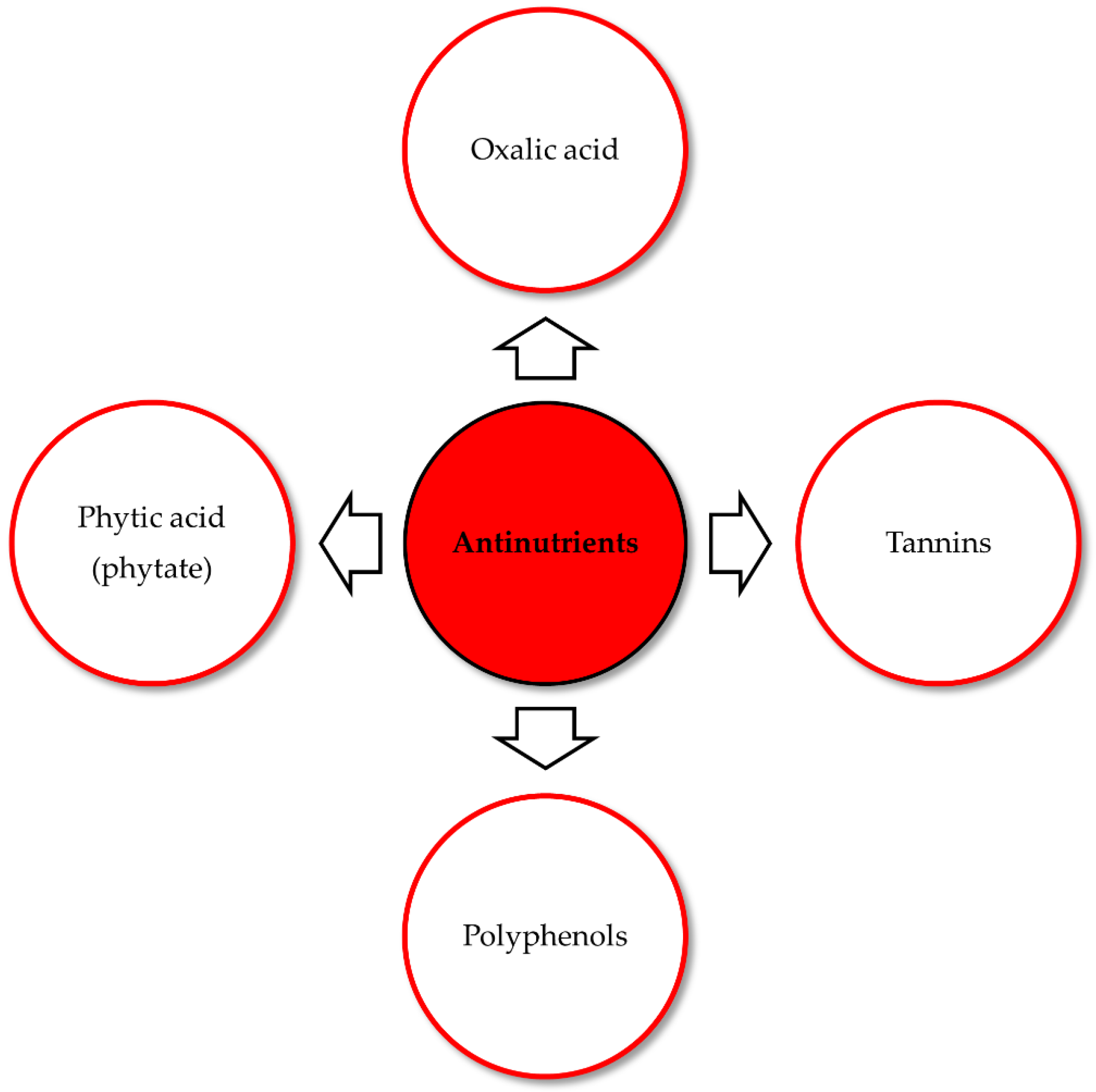
5. Regulating Causes: Antinutrients and Promoters
6. Conclusions and Future Perspectives
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- De Pascale, S.; Rouphael, Y. Chemical eustress and biofortification: Targeted nutrient solution management for enhancing quality in hydroponically grown vegetables. In Proceedings of the III International Symposium on Soilless Culture and Hydroponics: Innovation and Advanced Technology for Circular Horticulture 1321, Lemesos, Cyprus, 19–22 March 2021; pp. 179–184. [Google Scholar]
- Zandalinas, S.I.; Fritschi, F.B.; Mittler, R. Signal transduction networks during stress combination. J. Exp. Bot. 2020, 71, 1734–1741. [Google Scholar] [CrossRef]
- Inglese, P.; Sabatino, L. The Fresh Produce Supply Chain and Its Changes after the COVID-19. In COVID-19 and Communities; Springer: Berlin/Heidelberg, Germany, 2022; pp. 193–197. [Google Scholar]
- Vinoth, A.; Ravindhran, R. Biofortification in millets: A sustainable approach for nutritional security. Front. Plant Sci. 2017, 8, 29. [Google Scholar] [CrossRef]
- Pinstrup-Andersen, P. Agricultural research and policy for better health and nutrition in developing countries: A food systems approach. Agric. Econ. 2007, 37, 187–198. [Google Scholar] [CrossRef]
- Bouis, H. Reducing Mineral and Vitamin Deficiencies through Biofortification: Progress under HarvestPlus. In Hidden Hunger: Strategies to Improve Nutrition Quality; Karger Publishers: Basel, Switzerland, 2018; Volume 118, pp. 112–122. [Google Scholar]
- Jangir, C.K.; Kumar, S.; Lakhran, H.; Meena, R. Towards mitigating malnutrition in pulses through biofortification. Trends Biosci. 2017, 10, 2999–3002. [Google Scholar]
- Baldassano, S.; Di Gaudio, F.; Sabatino, L.; Caldarella, R.; De Pasquale, C.; Di Rosa, L.; Nuzzo, D.; Picone, P.; Vasto, S. Biofortification: Effect of Iodine fortified food in the healthy population, double-arm nutritional study. Front. Nutr. 2022, 9, 871638. [Google Scholar] [CrossRef]
- Baldassano, S.; Polizzi, M.R.; Sabatino, L.; Caldarella, R.; Macaluso, A.; Alongi, A.; Caldara, G.F.; Ferrantelli, V.; Vasto, S. A New potential dietary approach to supply micronutrients to physically active people through consumption of biofortified vegetables. Nutrients 2022, 14, 2971. [Google Scholar] [CrossRef]
- La Bella, S.; Consentino, B.B.; Rouphael, Y.; Ntatsi, G.; De Pasquale, C.; Iapichino, G.; Sabatino, L. Impact of Ecklonia maxima seaweed extract and mo foliar treatments on biofortification, spinach yield, quality and nue. Plants 2021, 10, 1139. [Google Scholar] [CrossRef] [PubMed]
- Sabatino, L.; Consentino, B.B.; Rouphael, Y.; De Pasquale, C.; Iapichino, G.; D’Anna, F.; La Bella, S. Protein hydrolysates and mo-biofortification interactively modulate plant performance and quality of ‘canasta’ lettuce grown in a protected environment. Agronomy 2021, 11, 1023. [Google Scholar] [CrossRef]
- Sabatino, L.; Di Gaudio, F.; Consentino, B.B.; Rouphael, Y.; El-Nakhel, C.; La Bella, S.; Vasto, S.; Mauro, R.P.; D’Anna, F.; Iapichino, G. Iodine biofortification counters micronutrient deficiency and improve functional quality of open field grown curly endive. Horticulturae 2021, 7, 58. [Google Scholar] [CrossRef]
- Pérez-Massot, E.; Banakar, R.; Gómez-Galera, S.; Zorrilla-López, U.; Sanahuja, G.; Arjó, G.; Miralpeix, B.; Vamvaka, E.; Farré, G.; Rivera, S.M. The contribution of transgenic plants to better health through improved nutrition: Opportunities and constraints. Genes Nutr. 2013, 8, 29–41. [Google Scholar] [CrossRef] [PubMed]
- Kyriacou, M.C.; Rouphael, Y. Towards a new definition of quality for fresh fruits and vegetables. Sci. Hortic. 2018, 234, 463–469. [Google Scholar] [CrossRef]
- Buturi, C.V.; Mauro, R.P.; Fogliano, V.; Leonardi, C.; Giuffrida, F. Mineral biofortification of vegetables as a tool to improve human diet. Foods 2021, 10, 223. [Google Scholar] [CrossRef] [PubMed]
- Marschner, H. Marschner’s Mineral Nutrition of Higher Plants, 3rd ed.; Academic Press: London, UK, 2012. [Google Scholar]
- Cakmak, I. Enrichment of cereal grains with zinc: Agronomic or genetic biofortification? Plant Soil 2008, 302, 1–17. [Google Scholar] [CrossRef]
- Chilimba, A.D.C.; Young, S.D.; Black, C.R.; Meacham, M.C.; Lammel, J.; Broadley, M.R. Agronomic biofortification of maize with selenium (Se) in Malawi. Field Crops Res. 2012, 125, 118–128. [Google Scholar] [CrossRef]
- Prasad, R.; Shivay, Y.S.; Kumar, D. Agronomic biofortification of cereal grains with iron and zinc. Adv. Agron. 2019, 125, 55–91. [Google Scholar]
- de Valenca, A.W.; Bake, A.; Brouwer, I.D.; Giller, K.E. Agronomic biofortification of crops to fight hidden hunger in sub-Saharan Africa. Glob. Food Sec. 2017, 12, 8–14. [Google Scholar] [CrossRef]
- Jaiswal, D.K.; Krishna, R.; Chouhan, G.K.; de Araujo Pereira, A.P.; Ade, A.B.; Prakash, S.; Verma, S.K.; Prasad, R.; Yadav, J.; Verma, J.P. Bio-fortification of minerals in crops: Current scenario and future prospects for sustainable agriculture and human health. Plant Growth Regul. 2022, 98, 5–22. [Google Scholar] [CrossRef]
- Stangoulis, J.C.R.; Knez, M. Biofortification of major crop plants with iron and zinc—Achievements and future directions. Plant Soil 2022, 474, 57–76. [Google Scholar] [CrossRef]
- Carvalho, S.M.; Vasconcelos, M.W. Producing more with less: Strategies and novel technologies for plant-based food biofortification. Food Res. Int. 2013, 54, 961–971. [Google Scholar] [CrossRef]
- White, P.J.; Broadley, M.R. Biofortification of crops with seven mineral elements often lacking in human diets–iron, zinc, copper, calcium, magnesium, selenium and iodine. New Phytol. 2009, 182, 49–84. [Google Scholar] [CrossRef]
- Li, R.; Li, D.-W.; Liu, H.-P.; Hong, C.-L.; Song, M.-Y.; Dai, Z.-X.; Liu, J.-W.; Zhou, J.; Weng, H.-X. Enhancing iodine content and fruit quality of pepper (Capsicum annuum L.) through biofortification. Sci. Hortic. 2017, 214, 165–173. [Google Scholar] [CrossRef]
- Li, R.; Liu, H.P.; Hong, C.L.; Dai, Z.X.; Liu, J.W.; Zhou, J.; Hu, C.Q.; Weng, H.X. Iodide and iodate effects on the growth and fruit quality of strawberry. J. Sci. Food Agric. 2017, 97, 230–235. [Google Scholar] [CrossRef] [PubMed]
- Wiesner-Reinhold, M.; Schreiner, M.; Baldermann, S.; Schwarz, D.; Hanschen, F.S.; Kipp, A.P.; Rowan, D.D.; Bentley-Hewitt, K.L.; McKenzie, M.J. Mechanisms of selenium enrichment and measurement in brassicaceous vegetables, and their application to human health. Front. Plant Sci. 2017, 8, 1365. [Google Scholar] [CrossRef] [PubMed]
- Montesano, F.F.; D’Imperio, M.; Parente, A.; Cardinali, A.; Renna, M.; Serio, F. Green bean biofortification for Si through soilless cultivation: Plant response and Si bioaccessibility in pods. Sci. Rep. 2016, 6, 31662. [Google Scholar] [CrossRef]
- White, P.J.; Brown, P. Plant nutrition for sustainable development and global health. Ann. Bot. 2010, 105, 1073–1080. [Google Scholar] [CrossRef]
- Niu, J.; Liu, C.; Huang, M.; Liu, K.; Yan, D. Effects of foliar fertilization: A review of current status and future perspectives. J. Soil Sci. Plant Nutr. 2021, 21, 104–118. [Google Scholar] [CrossRef]
- Mao, H.; Wang, J.; Wang, Z.; Zan, Y.; Lyons, G.; Zou, C. Using agronomic biofortification to boost zinc, selenium, and iodine concentrations of food crops grown on the loess plateau in China. J. Soil Sci. Plant Nutr. 2014, 14, 459–470. [Google Scholar] [CrossRef]
- Bouis, H.E.; Saltzman, A. Improving nutrition through biofortification: A review of evidence from HarvestPlus, 2003 through 2016. Glob. Food Secur. 2017, 12, 49–58. [Google Scholar] [CrossRef]
- Lemanceau, P.; Bauer, P.; Kraemer, S.; Briat, J.-F. Iron dynamics in the rhizosphere as a case study for analyzing interactions between soils, plants and microbes. Plant Soil 2009, 321, 513–535. [Google Scholar] [CrossRef]
- Kobayashi, T.; Nozoye, T.; Nishizawa, N.K. Iron transport and its regulation in plants. Free Radic. Biol. Med. 2019, 133, 11–20. [Google Scholar] [CrossRef]
- Tsukamoto, T.; Nakanishi, H.; Uchida, H.; Watanabe, S.; Matsuhashi, S.; Mori, S.; Nishizawa, N.K. 52Fe translocation in barley as monitored by a positron-emitting tracer imaging system (PETIS): Evidence for the direct translocation of Fe from roots to young leaves via phloem. Plant Cell Physiol. 2009, 50, 48–57. [Google Scholar] [CrossRef]
- Zhang, C.; Römheld, V.; Marschner, H. Distribution pattern of root-supplied 59iron in iron-sufficient and iron-deficient bean plants. J. Plant Nutr. 1995, 18, 2049–2058. [Google Scholar] [CrossRef]
- Zhang, X.; Zhang, D.; Sun, W.; Wang, T. The adaptive mechanism of plants to iron deficiency via iron uptake, transport, and homeostasis. Int. J. Mol. Sci. 2019, 20, 2424. [Google Scholar] [CrossRef]
- Ranieri, A.; Castagna, A.; Baldan, B.; Soldatini, G.F. Iron deficiency differently affects peroxidase isoforms in sunflower. J. Exp. Bot. 2001, 52, 25–35. [Google Scholar] [CrossRef] [PubMed]
- Sijmons, P.C.; Kolattukudy, P.; Bienfait, H.F. Iron deficiency decreases suberization in bean roots through a decrease in suberin-specific peroxidase activity. Plant Physiol. 1985, 78, 115–120. [Google Scholar] [CrossRef] [PubMed]
- Jin, C.W.; You, G.Y.; He, Y.F.; Tang, C.; Wu, P.; Zheng, S.J. Iron deficiency-induced secretion of phenolics facilitates the reutilization of root apoplastic iron in red clover. Plant Physiol. 2007, 144, 278–285. [Google Scholar] [CrossRef]
- Nahim-Granados, S.; Oller, I.; Malato, S.; Pérez, J.S.; Polo-Lopez, M. Commercial fertilizer as effective iron chelate (Fe3+-EDDHA) for wastewater disinfection under natural sunlight for reusing in irrigation. Appl. Catal. B 2019, 253, 286–292. [Google Scholar] [CrossRef]
- Briat, J.-F.; Duc, C.; Ravet, K.; Gaymard, F. Ferritins and iron storage in plants. Biochim. Biophys. Acta Gen. Subj. 2010, 1800, 806–814. [Google Scholar] [CrossRef] [PubMed]
- Fourcroy, P.; Vansuyt, G.; Kushnir, S.; Inzé, D.; Briat, J.-F. Iron-regulated expression of a cytosolic ascorbate peroxidase encoded by the APX1 gene in Arabidopsis seedlings. Plant Physiol. 2004, 134, 605–613. [Google Scholar] [CrossRef]
- Giordano, M.; El-Nakhel, C.; Pannico, A.; Kyriacou, M.C.; Stazi, S.R.; De Pascale, S.; Rouphael, Y. Iron biofortification of red and green pigmented lettuce in closed soilless cultivation impacts crop performance and modulates mineral and bioactive composition. Agronomy 2019, 9, 290. [Google Scholar] [CrossRef]
- Buturi, C.V.; Sabatino, L.; Mauro, R.P.; Navarro-León, E.; Blasco, B.; Leonardi, C.; Giuffrida, F. Iron biofortification of greenhouse soilless lettuce: An effective agronomic tool to improve the dietary mineral intake. Agronomy 2022, 12, 1793. [Google Scholar] [CrossRef]
- Lata-Tenesaca, L.F.; de Mello Prado, R.; Ajila-Celi, G.E.; da Silva, D.L.; Junior, J.S.P.; Mattiuz, B.H. Iron biofortification in quinoa: Effect of iron application methods on nutritional quality, anti-nutrient composition, and grain productivity. Food Chem. 2023, 404, 134573. [Google Scholar] [CrossRef] [PubMed]
- Adiloğlu, S. Relation of chelated iron (EDDHA-Fe) applications with iron accumulation and some plant nutrient elements in basil (Ocimum Basilicum L.). Pol. J. Environ. Stud. 2021, 30, 34713479. [Google Scholar] [CrossRef]
- Tsonev, T.; Cebola Lidon, F.J. Zinc in plants-an overview. Emi. J. Food Agric. 2012, 24, 322. [Google Scholar]
- Mengel, K.; Kirkby, E.A. Principles of Plant Nutrition, 5th ed.; Springer Science & Business Media: Berlin, Germany, 2012. [Google Scholar]
- Barak, P.; Helmke, P.A. The Chemistry of Zinc. In Zinc in Soils and Plants; Springer: Berlin/Heidelberg, Germany, 1993; pp. 1–13. [Google Scholar]
- Palmgren, M.G.; Clemens, S.; Williams, L.E.; Krämer, U.; Borg, S.; Schjørring, J.K.; Sanders, D. Zinc biofortification of cereals: Problems and solutions. Trends Plant Sci. 2008, 13, 464–473. [Google Scholar] [CrossRef]
- Stanton, C.; Sanders, D.; Krämer, U.; Podar, D. Zinc in plants: Integrating homeostasis and biofortification. Mol. Plant 2022, 15, 65–85. [Google Scholar] [CrossRef]
- Bashir, K.; Takahashi, R.; Nakanishi, H.; Nishizawa, N.K. The road to micronutrient biofortification of rice: Progress and prospects. Front. Plant Sci. 2013, 4, 15. [Google Scholar] [CrossRef]
- Milner, M.J.; Seamon, J.; Craft, E.; Kochian, L.V. Transport properties of members of the ZIP family in plants and their role in Zn and Mn homeostasis. J. Exp. Bot. 2013, 64, 369–381. [Google Scholar] [CrossRef]
- Tiong, J.; McDonald, G.K.; Genc, Y.; Pedas, P.; Hayes, J.E.; Toubia, J.; Langridge, P.; Huang, C.Y. HvZIP7 mediates zinc accumulation in barley (Hordeum vulgare) at moderately high zinc supply. New Phytol. 2014, 201, 131–143. [Google Scholar] [CrossRef]
- White, P.J.; Whiting, S.N.; Baker, A.J.; Broadley, M.R. Does zinc move apoplastically to the xylem in roots of Thlaspi caerulescens? New Phytol. 2002, 153, 201–207. [Google Scholar] [CrossRef]
- Fernández, V.; Brown, P.H. From plant surface to plant metabolism: The uncertain fate of foliar-applied nutrients. Front. Plant Sci. 2013, 4, 289. [Google Scholar] [CrossRef] [PubMed]
- Mazé, P. Determination of the rarer mineral elements necessary to the development of corn. Comptes Rendus Acad. Sci. 1915, 160, 211–214. [Google Scholar]
- Sommer, A.L.; Lipman, C. Evidence on the indispensable nature of zinc and boron for higher green plants. Plant Physiol. 1926, 1, 231. [Google Scholar] [CrossRef] [PubMed]
- Sharma, S.S.; Dietz, K.-J. The significance of amino acids and amino acid-derived molecules in plant responses and adaptation to heavy metal stress. J. Exp. Bot. 2006, 57, 711–726. [Google Scholar] [CrossRef] [PubMed]
- Broadley, M.R.; White, P.J.; Bryson, R.J.; Meacham, M.C.; Bowen, H.C.; Johnson, S.E.; Hawkesford, M.J.; McGrath, S.P.; Zhao, F.-J.; Breward, N. Biofortification of UK food crops with selenium. Proc. Nutr. Soc. 2006, 65, 169–181. [Google Scholar] [CrossRef]
- Wei, Y.; Shohag, M.J.; Yang, X. Biofortification and bioavailability of rice grain zinc as affected by different forms of foliar zinc fertilization. PLoS ONE 2012, 7, e45428. [Google Scholar] [CrossRef]
- Alloway, B.J. Zinc in Soils and Crop Nutrition, 2nd ed.; International Zinc Association and International Fertilizer Industry Association: Paris, France, 2008. [Google Scholar]
- White, J.G.; Zasoski, R.J. Mapping soil micronutrients. Field Crops Res. 1999, 60, 11–26. [Google Scholar] [CrossRef]
- Hassan, M.U.; Chattha, M.U.; Ullah, A.; Khan, I.; Qadeer, A.; Aamer, M.; Khan, A.U.; Nadeem, F.; Khan, T.A. Agronomic biofortification to improve productivity and grain Zn concentration of bread wheat. Int. J. Agric. Biol. 2019, 21, 615–620. [Google Scholar]
- Mousavi, S.R.; Galavi, M.; Ahmadvand, G. Effect of zinc and manganese foliar application on yield, quality and enrichment on potato (Solanum tuberosum L.). Asian J.Plant Sci. 2007, 6, 1256–1260. [Google Scholar] [CrossRef]
- Eslami, M.; Dehghanzadeh, H.; Jafarzade, M.; Aminian, R. The effect of zinc on yield and yield components of sunflower (Helianthus annuus L.) under drought stress. Sci. J. Crop Sci. 2014, 3, 61–65. [Google Scholar]
- Hajiboland, R.; Amirazad, H. Drought tolerance in Zn-deficient red cabbage (Brassica oleracea L. var. capitata f. rubra) plants. Hortic. Sci. 2010, 37, 88–98. [Google Scholar] [CrossRef]
- Sadoogh, F.; Shariatmadari, H.; Khoshgoftarmanesh, A.; Mosaddeghi, M. Adjusted nutrition of tomato with potassium and zinc in drought stress conditions induced by polyethylene glycol 6000 in hydroponic culture. J. Sci. Technol. Greenh. Cult. 2014, 5, 67–81. [Google Scholar]
- Sago, Y.; Watanabe, N.; Minami, Y. Zinc biofortification of hydroponic baby leaf lettuce grown under artificial lighting with elevated wind speed and root zone temperature. J. Agric. Meteorol. 2018, 74, 173–177. [Google Scholar] [CrossRef]
- D’Imperio, M.; Montesano, F.F.; Serio, F.; Santovito, E.; Parente, A. Mineral Composition and Bioaccessibility in Rocket and Purslane after Zn Biofortification Process. Foods 2022, 11, 484. [Google Scholar] [CrossRef] [PubMed]
- Ciriello, M.; Formisano, L.; Kyriacou, M.; Soteriou, G.A.; Graziani, G.; De Pascale, S.; Rouphael, Y. Zinc biofortification of hydroponically grown basil: Stress physiological responses and impact on antioxidant secondary metabolites of genotypic variants. Front. Plant Sci. 2022, 13, 1049004. [Google Scholar] [CrossRef]
- Chaney, R.A. Zinc Phytotoxicity. In Zinc in Soils and Plants; Springer: Dordrecht, The Netherlands, 1993; pp. 135–150. [Google Scholar]
- Boawn, L.C.; Rasmussen, P. Crop response to excessive zinc fertilization of alkaline soil 1. Agron. J. 1971, 63, 874–876. [Google Scholar] [CrossRef]
- de Oliveira, N.T.; de Rezende, P.M.; de Fatima Piccolo Barcelos, M.; Bruzi, A.T. Zinc biofortification strategies in food-type soybean cultivars. Aust. J. Crop Sci. 2019, 13, 11–16. [Google Scholar] [CrossRef]
- Shivay, Y.S.; Prasad, R.; Pal, M. Effects of source and method of zinc application on yield, zinc biofortification of grain, and Zn uptake and use efficiency in chickpea (Cicer arietinum L.). Commun. Soil Sci. Plant Anal. 2015, 46, 2191–2200. [Google Scholar] [CrossRef]
- Yilmaz, A.; Ekiz, H.; Torun, B.; Gultekin, I.; Karanlik, S.; Bagci, S.A.; Cakmak, I. Effect of different zinc application methods on grain yield and zinc concentration in wheat cultivars grown on zinc-deficient calcareous soils. J. Plant Nutr. 1997, 20, 461–471. [Google Scholar] [CrossRef]
- Solanki, P.; Laura, J.S. Biofortification of crops using nanoparticles to alleviate plant and human Zn deficiency: A review. Res. J. Life Sci. Bioinform. Pharm. Chem. Sci. 2018, 4, 364–385. [Google Scholar]
- Long, X.X.; Yang, X.E.; Ni, W.Z.; Ye, Z.Q.; He, Z.L.; Calvert, D.V.; Stoffella, J.P. Assessing zinc thresholds for phytotoxicity and potential dietary toxicity in selected vegetable crops. Commun. Soil Sci. Plant Anal. 2003, 34, 1421–1434. [Google Scholar] [CrossRef]
- Gonzali, S.; Kiferle, C.; Perata, P. Iodine biofortification of crops: Agronomic biofortification, metabolic engineering and iodine bioavailability. Curr. Opin. Biotechnol. 2017, 44, 16–26. [Google Scholar] [CrossRef]
- Fuge, R.; Johnson, C.C. Iodine and human health, the role of environmental geochemistry and diet, a review. Appl. Geochem. 2015, 63, 282–302. [Google Scholar] [CrossRef]
- Halka, M.; Smoleń, S.; Czernicka, M.; Klimek-Chodacka, M.; Pitala, J.; Tutaj, K. Iodine biofortification through expression of HMT, SAMT and S3H genes in Solanum lycopersicum L. Plant Physiol. Biochem. 2019, 144, 35–48. [Google Scholar] [CrossRef]
- Ren, Q.; Fan, J.; Zhang, Z.; Zheng, X.; DeLong, G.R. An environmental approach to correcting iodine deficiency: Supplementing iodine in soil by iodination of irrigation water in remote areas. J. Trace Elem. Med. Biol. 2008, 22, 1–8. [Google Scholar] [CrossRef]
- Medrano-Macías, J.; Leija-Martínez, P.; González-Morales, S.; Juárez-Maldonado, A.; Benavides-Mendoza, A. Use of iodine to biofortify and promote growth and stress tolerance in crops. Front. Plant Sci. 2016, 7, 1146. [Google Scholar] [CrossRef]
- Weng, H.-X.; Weng, J.-K.; Yan, A.-L.; Hong, C.-L.; Yong, W.-B.; Qin, Y.-C. Increment of iodine content in vegetable plants by applying iodized fertilizer and the residual characteristics of iodine in soil. Biol. Trace Elem. Res. 2008, 123, 218–228. [Google Scholar] [CrossRef]
- Caffagni, A.; Arru, L.; Meriggi, P.; Milc, J.; Perata, P.; Pecchioni, N. Iodine fortification plant screening process and accumulation in tomato fruits and potato tubers. Commun. Soil Sci. Plant Anal. 2011, 42, 706–718. [Google Scholar] [CrossRef]
- Hong, C.-L.; Weng, H.-X.; Yan, A.-L.; Islam, E.-U. The fate of exogenous iodine in pot soil cultivated with vegetables. Environ. Geochem. Health 2009, 31, 99–108. [Google Scholar] [CrossRef]
- Kiferle, C.; Gonzali, S.; Holwerda, H.T.; Ibaceta, R.R.; Perata, P. Tomato fruits: A good target for iodine biofortification. Front. Plant Sci. 2013, 4, 205. [Google Scholar] [CrossRef] [PubMed]
- Landini, M.; Gonzali, S.; Perata, P. Iodine biofortification in tomato. J. Plant Nutri. Soil Sci. 2011, 174, 480–486. [Google Scholar] [CrossRef]
- Weng, H.-X.; Yan, A.-L.; Hong, C.-L.; Xie, L.-L.; Qin, Y.-C.; Cheng, C.Q. Uptake of different species of iodine by water spinach and its effect to growth. Biol. Trace Elem. Res. 2008, 124, 184–194. [Google Scholar] [CrossRef] [PubMed]
- Blasco, B.; Rios, J.; Cervilla, L.; Sánchez-Rodrigez, E.; Ruiz, J.; Romero, L. Iodine biofortification and antioxidant capacity of lettuce: Potential benefits for cultivation and human health. Ann. Appl. Biol. 2008, 152, 289–299. [Google Scholar] [CrossRef]
- Zhu, Y.G.; Huang, Y.Z.; Hu, Y.; Liu, Y.X. Iodine uptake by spinach (Spinacia oleracea L.) plants grown in solution culture: Effects of iodine species and solution concentrations. Environ. Int. 2003, 29, 33–37. [Google Scholar] [CrossRef]
- Muramatsu, Y.; Christoffers, D.; Ohmomo, Y. Influence of chemical forms on iodine uptake by plant. J. Radiat Res. 1993, 24, 326–338. [Google Scholar] [CrossRef] [PubMed]
- Incrocci, L.; Carmassi, G.; Maggini, R.; Poli, C.; Saidov, D.; Tamburini, C.; Pardossi, A. Iodine accumulation and tolerance in sweet basil (Ocimum basilicum L.) with green or purple leaves grown in floating system technique. Front. Plant Sci. 2019, 10, 1494. [Google Scholar] [CrossRef]
- Weng, H.; Hong, C.; Yan, A.; Ji, Z. Biogeochemical transport of iodine and its quantitative model. Sci. China Earth Sci. 2013, 56, 1599–1606. [Google Scholar] [CrossRef]
- Hong, C.L.; Weng, H.X.; Qin, Y.C.; Yan, A.L.; Xie, L.L. Transfer of iodine from soil to vegetables by applying exogenous iodine. Agron. Sustain. Dev. 2008, 28, 575–583. [Google Scholar] [CrossRef]
- Caffagni, A.; Pecchioni, N.; Meriggi, P.; Bucci, V.; Sabatini, E.; Acciarri, N.; Ciriaci, T.; Pulcini, L.; Felicioni, N.; Beretta, M. Iodine uptake and distribution in horticultural and fruit tree species. Ital. J. Agron. 2012, 7, e32. [Google Scholar] [CrossRef]
- Smoleń, S.; Kowalska, I.; Sady, W. Assessment of biofortification with iodine and selenium of lettuce cultivated in the NFT hydroponic system. Sci. Hortic. 2014, 166, 9–16. [Google Scholar] [CrossRef]
- Leyva, R.; Sánchez-Rodríguez, E.; Ríos, J.J.; Rubio-Wilhelmi, M.M.; Romero, L.; Ruiz, J.M.; Blasco, B. Beneficial effects of exogenous iodine in lettuce plants subjected to salinity stress. Plant Sci. 2011, 181, 195–202. [Google Scholar] [CrossRef]
- Neha, G.; Bajpai, M.S.; Majumdar, R.S.; Mishra, P.K. Response of iodine on antioxidant levels of Glycine max L. grown under Cd2+ stress. Adv. Biol. Res. 2015, 9, 40–48. [Google Scholar]
- Sabatino, L.; D’Anna, F.; Iapichino, G.; Moncada, A.; D’Anna, E.; De Pasquale, C. Interactive effects of genotype and molybdenum supply on yield and overall fruit quality of tomato. Front. Plant Sci. 2019, 9, 1922. [Google Scholar] [CrossRef] [PubMed]
- Consentino, B.B.; Rouphael, Y.; Ntatsi, G.; De Pasquale, C.; Iapichino, G.; D’Anna, F.; La Bella, S.; Sabatino, L. Agronomic performance and fruit quality in greenhouse grown eggplant are interactively modulated by iodine dosage and grafting. Sci. Hortic. 2022, 295, 110891. [Google Scholar] [CrossRef]
- Lawson, P.G.; Daum, D.; Czauderna, R.; Meuser, H.; Härtling, J.W. Soil versus foliar iodine fertilization as a biofortification strategy for field-grown vegetables. Front. Plant Sci. 2015, 6, 450. [Google Scholar] [CrossRef]
- Strzetelski, P.; Smoleń, S.; Rożek, S.; Sady, W. The effect of differentiated fertilization and foliar application of iodine on yielding and antioxidant properties in radish (Raphanus sativus L.). Plants. Ecol. Chem. Eng. A 2010, 17, 1189–1195. [Google Scholar]
- Smolen, S.; Rozek, R.; Ledwozyw-Smolen, I.; Strzetelski, P. Preliminary evaluation of the influence of soil fertilization and foliar nutrition with iodine on the efficiency of iodine biofortification and chemical composition of lettuce. J. Elem. 2011, 16, 613–622. [Google Scholar] [CrossRef]
- He, Z.L.; Yang, X.E.; Stoffella, P.J. Trace elements in agroecosystems and impacts on the environment. J. Trace Elem. Med. Biol. 2005, 19, 125–140. [Google Scholar] [CrossRef]
- Kaiser, B.N.; Gridley, K.L.; Ngaire Brady, J.; Phillips, T.; Tyerman, S.D. The role of molybdenum in agricultural plant production. Ann. Bot. 2005, 96, 745–754. [Google Scholar] [CrossRef]
- Gupta, U. (Ed.) Bioavalability of Molybdate Depends on the Soil Properties. In Molybdenum in Agriculture; Cambridge University Press: Cambridge, MA, USA, 1997; pp. 71–91. [Google Scholar]
- Tejada-Jiménez, M.; Chamizo-Ampudia, A.; Galván, A.; Fernández, E.; Llamas, Á. Molybdenum metabolism in plants. Metallomics 2013, 5, 1191–1203. [Google Scholar] [CrossRef]
- Leimkühler, S.; Iobbi-Nivol, C. Bacterial molybdoenzymes: Old enzymes for new purposes. FEMS Microbiol. Rev. 2016, 40, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Mendel, R.R.; Kruse, T. Cell biology of molybdenum in plants and humans. Biochim. et Biophys. Acta (BBA)-Mol. Cell Res. 2012, 1823, 1568–1579. [Google Scholar] [CrossRef] [PubMed]
- Stiefel, E.I. The biogeochemistry of molybdenum and tungsten. Met. Ions Biol. Syst. 2002, 39, 1–29. [Google Scholar]
- Campbell, W.H. Nitrate reductase structure, function and regulation: Bridging the gap between biochemistry and physiology. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1999, 50, 277–303. [Google Scholar] [CrossRef] [PubMed]
- Seo, M.; Akaba, S.; Oritani, T.; Delarue, M.; Bellini, C.; Caboche, M.; Koshiba, T. Higher activity of an aldehyde oxidase in the auxin-overproducing superroot1 mutant of Arabidopsis thaliana. Plant Physiol. 1998, 116, 687–693. [Google Scholar] [CrossRef] [PubMed]
- Seo, M.; Peeters, A.J.M.; Koiwai, H.; Oritani, T.; Marion-Poll, A.; Zeevaart, J.A.D.; Koornneef, M.; Kamiya, Y.; Koshiba, T. The Arabidopsis aldehyde oxidase 3 (AAO3) gene product catalyzes the final step in abscisic acid biosynthesis in leaves. Proc. Natl. Acad. Sci. USA 2000, 97, 12908–12913. [Google Scholar] [CrossRef]
- Min, Y.; Hu, C.-X.; Wang, Y.-H. Effects of molybdenum on the intermediates of chlorophyll biosynthesis in winter wheat cultivars under low temperature. Agric. Sci. China 2006, 5, 670–677. [Google Scholar]
- Smith, K.S.; Balistrieri, L.S.; Smith, S.M.; Severson, R.C. Distribution and Mobility of Molybdenum in the Terrestrial Environment; Cambridge University Press: Cambridge, UK, 1997. [Google Scholar]
- Kovács, B.; Puskás-Preszner, A.; Huzsvai, L.; Lévai, L.; Bódi, É. Effect of molybdenum treatment on molybdenum concentration and nitrate reduction in maize seedlings. Plant Physiol. Biochem. 2015, 96, 38–44. [Google Scholar] [CrossRef]
- van Gestel, C.A.; McGrath, S.P.; Smolders, E.; Ortiz, M.D.; Borgman, E.; Verweij, R.A.; Buekers, J.; Oorts, K. Effect of long-term equilibration on the toxicity of molybdenum to soil organisms. Environ. Pollut. 2012, 162, 1–7. [Google Scholar] [CrossRef]
- Luk, E.; Jensen, L.T.; Culotta, V.C. The many highways for intracellular trafficking of metals. J.Biol. Inorg. Chem. 2003, 8, 803–809. [Google Scholar] [CrossRef]
- Dhaliwal, S.S.; Sharma, V.; Shukla, A.K.; Kaur, J.; Verma, V.; Kaur, M.; Singh, P.; Brestic, M.; Gaber, A.; Hossain, A. Interactive effects of molybdenum, zinc and iron on the grain yield, quality, and nodulation of cowpea (Vigna unguiculata (L.) Walp.) in North-Western India. Molecules 2022, 27, 3622. [Google Scholar] [CrossRef]
- Moncada, A.; Miceli, A.; Sabatino, L.; Iapichino, G.; D’Anna, F.; Vetrano, F. Effect of molybdenum rate on yield and quality of lettuce, escarole, and curly endive grown in a floating system. Agronomy 2018, 8, 171. [Google Scholar] [CrossRef]
- Mondy, N.I.; Munshi, C.B. Effect of soil and foliar application of molybdenum on the glycoalkaloid and nitrate concentration of potatoes. J. Agric. Food Chem. 1993, 41, 256–258. [Google Scholar] [CrossRef]
- Jena, B.; Nayak, R.; Bhol, R.; Sahoo, S. Soil and foliar application of molybdenum on yield, biochemical quality of green gram (Vigna Radiata L.) grown in Fe rich soils. Bangladesh J. Bot. 2022, 51, 689–696. [Google Scholar] [CrossRef]
- Floor, G.H.; Román-Ross, G. Selenium in volcanic environments: A review. Appl. Geochem. 2012, 27, 517–531. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Bhuyan, M.B.; Raza, A.; Hawrylak-Nowak, B.; Matraszek-Gawron, R.; Al Mahmud, J.; Nahar, K.; Fujita, M. Selenium in plants: Boon or bane? Environ. Exp. Bot. 2020, 178, 104170. [Google Scholar] [CrossRef]
- Chen, L.; Yang, F.; Xu, J.; Hu, Y.; Hu, Q.; Zhang, Y.; Pan, G. Determination of selenium concentration of rice in China and effect of fertilization of selenite and selenate on selenium content of rice. J. Agric. Food Chem. 2002, 50, 5128–5130. [Google Scholar] [CrossRef] [PubMed]
- Hawkesford, M.J.; Zhao, F.-J. Strategies for increasing the selenium content of wheat. J. Cereal Sci. 2007, 46, 282–292. [Google Scholar] [CrossRef]
- Schiavon, M.; dall’Acqua, S.; Mietto, A.; Pilon-Smits, E.A.; Sambo, P.; Masi, A.; Malagoli, M. Selenium fertilization alters the chemical composition and antioxidant constituents of tomato (Solanum lycopersicon L.). J. Agric. Food Chem. 2013, 61, 10542–10554. [Google Scholar] [CrossRef]
- Fordyce, F.M. Selenium Deficiency and Toxicity in the Environment. In Essentials of Medical Geology; Springer: Berlin/Heidelberg, Germany, 2013; pp. 375–416. [Google Scholar]
- Bodnar, M.; Konieczka, P.; Namiesnik, J. The properties, functions, and use of selenium compounds in living organisms. J. Environ. Sci. Health Part C 2012, 30, 225–252. [Google Scholar] [CrossRef]
- Boyd, R. Selenium stories. Nat. Chem. 2011, 3, 570. [Google Scholar] [CrossRef]
- Hossain, A.; Skalicky, M.; Brestic, M.; Maitra, S.; Sarkar, S.; Ahmad, Z.; Vemuri, H.; Garai, S.; Mondal, M.; Bhatt, R.; et al. Selenium biofortification: Roles, mechanisms, responses and prospects. Molecules 2021, 26, 881. [Google Scholar] [CrossRef]
- Germ, M.; Stibilj, V. Selenium and plants. Acta Agric. Slov. 2007, 89, 65–71. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Nahar, K.; Alam, M.; Fujita, M. Modulation of antioxidant machinery and the methylglyoxal detoxification system in selenium-supplemented Brassica napus seedlings confers tolerance to high temperature stress. Biol. Trace Elem. Res. 2014, 161, 297–307. [Google Scholar] [CrossRef] [PubMed]
- Pannico, A.; El-Nakhel, C.; Graziani, G.; Kyriacou, M.C.; Giordano, M.; Soteriou, G.A.; Zarrelli, A.; Ritieni, A.; De Pascale, S.; Rouphael, Y. Selenium biofortification impacts the nutritive value, polyphenolic content, and bioactive constitution of variable microgreens genotypes. Antioxidants 2020, 9, 272. [Google Scholar] [CrossRef]
- Puccinelli, M.; Malorgio, F.; Incrocci, L.; Rosellini, I.; Pezzarossa, B. Effects of individual and simultaneous selenium and iodine biofortification of baby-leaf lettuce plants grown in two different hydroponic systems. Horticulturae 2021, 7, 590. [Google Scholar] [CrossRef]
- Puccinelli, M.; Malorgio, F.; Pintimalli, L.; Rosellini, I.; Pezzarossa, B. Biofortification of lettuce and basil seedlings to produce selenium enriched leafy vegetables. Horticulturae 2022, 8, 801. [Google Scholar] [CrossRef]
- Nascimento, C.S.; Nascimento, C.S.; Lopes, G.; Carrasco, G.; Gratão, P.L.; Cecílio Filho, A.B. Biofortified rocket (Eruca sativa) with selenium by using the nutrient film technique. Horticulturae 2022, 8, 1088. [Google Scholar] [CrossRef]
- Abdalla, M.A.; Wick, J.E.; Famuyide, I.M.; McGaw, L.J.; Mühling, K.H. Selenium enrichment of green and red lettuce and the induction of radical scavenging potential. Horticulturae 2021, 7, 488. [Google Scholar] [CrossRef]
- Pannico, A.; El-Nakhel, C.; Kyriacou, M.C.; Giordano, M.; Stazi, S.R.; De Pascale, S.; Rouphael, Y. Combating micronutrient deficiency and enhancing food functional quality through selenium fortification of select lettuce genotypes grown in a closed soilless system. Front. Plant Sci. 2019, 10, 1495. [Google Scholar] [CrossRef]
- White, P.; Bowen, H.; Parmaguru, P.; Fritz, M.; Spracklen, W.; Spiby, R.; Meacham, M.; Mead, A.; Harriman, M.; Trueman, L. Interactions between selenium and sulphur nutrition in Arabidopsis thaliana. J. Exp. Bot. 2004, 55, 1927–1937. [Google Scholar] [CrossRef] [PubMed]
- El-Ramady, H.; Faizy, S.E.-D.; Abdalla, N.; Taha, H.; Domokos-Szabolcsy, É.; Fari, M.; Elsakhawy, T.; Omara, A.E.-D.; Shalaby, T.; Bayoumi, Y.; et al. Selenium and nano-selenium biofortification for human health: Opportunities and challenges. Soil Syst. 2020, 4, 57. [Google Scholar] [CrossRef]
- Terry, N.; Zayed, A.; De Souza, M.; Tarun, A. Selenium in higher plants. Annu. Rev. Plant Biol. 2000, 51, 401–432. [Google Scholar] [CrossRef] [PubMed]
- White, P.J. Selenium accumulation by plants. Ann. Bot. 2016, 117, 217–235. [Google Scholar] [CrossRef]
- Li, H.F.; McGrath, S.P.; Zhao, F.J. Selenium uptake, translocation and speciation in wheat supplied with selenate or selenite. New Phytol. 2008, 178, 92–102. [Google Scholar] [CrossRef] [PubMed]
- Dennert, G.; Zwahlen, M.; Brinkman, M.; Vinceti, M.; Zeegers, M.P.; Horneber, M. Selenium for preventing cancer. Cochrane Database Syst. Rev. 2011, 5, CD005195. [Google Scholar]
- Lyons, G.H.; Stangoulis, J.C.R.; Graham, R.D. Exploiting micronutrient interaction to optimize biofortification programs: The case for inclusion of selenium and iodine in the Harvest Plust programs. Nutr. Rev. 2004, 62, 247–252. [Google Scholar] [CrossRef]
- Ramos, S.J.; Faquin, V.; Guilherme, L.R.G.; Castro, E.M.; Ávila, F.W.; Carvalho, G.S.; Bastos, C.E.A.; Oliveira, C. Selenium biofortification and antioxidant activity in lettuce plants fed with selenate and selenite. Plant Soil Environ. 2010, 56, 584–588. [Google Scholar] [CrossRef]
- Sabatino, L.; Ntatsi, G.; Iapichino, G.; D’Anna, F.; De Pasquale, C. Effect of selenium enrichment and type of application on yield, functional quality and mineral composition of curly endive grown in a hydroponic System. Agronomy 2019, 9, 207. [Google Scholar] [CrossRef]
- Shalaby, T.; Bayoumi, Y.; Alshaal, T.; Elhawat, N.; Sztrik, A.; El-Ramady, H. Selenium fortification induces growth, antioxidant activity, yield and nutritional quality of lettuce in salt-affected soil using foliar and soil applications. Plant Soil 2017, 421, 245–258. [Google Scholar] [CrossRef]
- Newman, R.; Waterland, N.; Moon, Y.; Tou, J.C. Selenium biofortification of agricultural crops and effects on plant nutrients and bioactive compounds important for human health and disease prevention—A Review. Plant Foods Hum. Nutr. 2019, 74, 449–460. [Google Scholar] [CrossRef]
- Pilon-smits, E.A.H.; Winkel, L.H.E.; Lin, Z. Selenium in Plants, 1st ed.; Springer: Cham, Switzerland; Midtown Manhattan, NY, USA, 2017. [Google Scholar]
- Schiavon, M.; Pilon-Smits, E.A.H. The fascinating facets of plant selenium accumulation—Biochemistry, physiology, evolution and ecology. New Phytol. 2017, 213, 1582–1596. [Google Scholar] [CrossRef] [PubMed]
- White, P.J.; Bowen, H.C.; Marshall, B.; Broadley, M.R. Extraordinarily high leaf selenium to sulfur ratios define ‘Se-accumulator’plants. Annal. Bot. 2007, 100, 111–118. [Google Scholar] [CrossRef] [PubMed]
- Pilon-Smits, E.A.; Le Duc, D.L. Phytoremediation of selenium using transgenic plants. Curr. Opin. Biotechnol. 2009, 20, 207–212. [Google Scholar] [CrossRef]
- Cooke, J.; DeGabriel, J.L.; Hartley, S.E. The functional ecology of plant silicon. Funct. Ecol. 2016, 30, 1270–1276. [Google Scholar] [CrossRef]
- Coskun, D.; Britto, D.T.; Huynh, W.Q.; Kronzucker, H.J. The role of silicon in higher plants under salinity and drought stress. Fron. Plant Sci. 2016, 7, 1072. [Google Scholar] [CrossRef]
- Coskun, D.; Deshmukh, R.; Sonah, H.; Menzies, J.G.; Reynolds, O.; Ma, J.F.; Kronzucker, H.J.; Bélanger, R.R. The controversies of silicon's role in plant biology. New Phytol. 2019, 221, 67–85. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Wang, R.; Mur, L.A.J.; Ruan, J.; Shen, Q.; Guo, S. Functions of silicon in plant drought stress responses. Hortic. Res. 2021, 8, 254. [Google Scholar] [CrossRef]
- Liang, Y.; Hua, H.; Zhu, Y.G.; Zhang, J.; Cheng, C.; R÷ mheld, V. Importance of plant species and external silicon concentration to active silicon uptake and transport. New Phytol. 2006, 172, 63–72. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, E.; Ma, J.; Miyake, Y. The possibility of silicon as an essential element for higher plants. J. Agric. Food Chem. 1990, 2, 99–102. [Google Scholar]
- Richmond, K.E.; Sussman, M. Got silicon? The non-essential beneficial plant nutrient. Curr. Opin. Plant Biol. 2003, 6, 268–272. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.F.; Yamaji, N. Silicon uptake and accumulation in higher plants. Trends Plant Sci. 2006, 11, 392–397. [Google Scholar] [CrossRef] [PubMed]
- Puppe, D.; Sommer, M. Experiments, uptake mechanisms, and functioning of silicon foliar fertilization—A review focusing on maize, rice, and wheat. Adv. Agron. 2018, 152, 1–49. [Google Scholar]
- Epstein, E. The anomaly of silicon in plant biology. Proc. Natl. Acad. Sci. USA 1994, 91, 11–17. [Google Scholar] [CrossRef]
- Hodson, M.; White, P.; Mead, A.; Broadley, M. Phylogenetic variation in the silicon composition of plants. Ann. Bot. 2005, 96, 1027–1046. [Google Scholar] [CrossRef]
- Hameed, A.; Sheikh, M.A.; Jamil, A.; Basra, S.M.A. Seed priming with sodium silicate enhances seed germination and seedling growth in wheat (Triticum aestivum L.) under water deficit stress induced by polyethylene glycol. Pak. J. Life Soc. Sci. 2013, 11, 19–24. [Google Scholar]
- Shi, Y.; Zhang, Y.; Yao, H.; Wu, J.; Sun, H.; Gong, H. Silicon improves seed germination and alleviates oxidative stress of bud seedlings in tomato under water deficit stress. Plant Physiol. Biochem. 2014, 78, 27–36. [Google Scholar] [CrossRef]
- Zargar, S.M.; Agnihotri, A. Impact of silicon on various agro-morphological and physiological parameters in maize and revealing its role in enhancing water stress tolerance. Emir. J. Food Agric. 2013, 25, 138–141. [Google Scholar]
- Zia, Z.; Bakhat, H.F.; Saqib, Z.A.; Shah, G.M.; Fahad, S.; Ashraf, M.R.; Hammad, H.M.; Naseem, W.; Shahid, M. Effect of water management and silicon on germination, growth, phosphorus and arsenic uptake in rice. Ecotoxicol. Environ. Saf. 2017, 144, 11–18. [Google Scholar] [CrossRef]
- Chen, W.; Yao, X.; Cai, K.; Chen, J. Silicon alleviates drought stress of rice plants by improving plant water status, photosynthesis and mineral nutrient absorption. Biol. Trace Elem. Res. 2011, 142, 67–76. [Google Scholar] [CrossRef]
- Emam, M.M.; Khattab, H.E.; Helal, N.M.; Deraz, A.E. Effect of selenium and silicon on yield quality of rice plant grown under drought stress. Aust. J. Crop Sci. 2014, 8, 596–605. [Google Scholar]
- Ming, D.; Pei, Z.; Naeem, M.; Gong, H.; Zhou, W. Silicon alleviates PEG-induced water-deficit stress in upland rice seedlings by enhancing osmotic adjustment. J. Agron. Crop Sci. 2012, 198, 14–26. [Google Scholar] [CrossRef]
- Valentinuzzi, F.; Pii, Y.; Borruso, L.; Mimmo, T.; Puglisi, E.; Trevisan, M.; Cesco, S. Epiphytic microbial community and post-harvest characteristics of strawberry fruits as affected by plant nutritional regime with silicon. Agronomy 2021, 11, 2407. [Google Scholar] [CrossRef]
- Hidalgo-Santiago, L.; Navarro-León, E.; López-Moreno, F.J.; Arjó, G.; González, L.M.; Ruiz, J.M.; Blasco, B. The application of the silicon-based biostimulant Codasil® offset water deficit of lettuce plants. Sci. Hortic. 2021, 285, 110177. [Google Scholar] [CrossRef]
- da Silva, D.L.; de Mello Prado, R.; Tenesaca, L.F.L.; da Silva, J.L.F.; Mattiuz, B.-H. Silicon attenuates calcium deficiency in rocket plants by increasing the production of non-enzymatic antioxidants compounds. Sci. Hortic. 2021, 285, 110169. [Google Scholar] [CrossRef]
- D’Imperio, M.; Montesano, F.F.; Renna, M.; Leoni, B.; Buttaro, D.; Parente, A.; Serio, F. NaCl stress enhances silicon tissue enrichment of hydroponic “baby leaf” chicory under biofortification process. Sci. Hortic. 2018, 235, 258–263. [Google Scholar] [CrossRef]
- Pilon, C.; Soratto, R.P.; Broetto, F.; Fernandes, A.M. Foliar or soil applications of silicon alleviate water-deficit stress of potato plants. Agron. J. 2014, 106, 2325–2334. [Google Scholar] [CrossRef]
- Kaushik, P.; Saini, D.K. Silicon as a vegetable crops modulator—A review. Plant 2019, 8, 148. [Google Scholar] [CrossRef]
- Hoffmann, J.; Berni, R.; Hausman, J.-F.; Guerriero, G. A Review on the beneficial role of silicon against salinity in non-accumulator crops: Tomato as a model. Biomolecules 2020, 10, 1284. [Google Scholar] [CrossRef]
- Li, Y. The Effects of Silicon Nutrition on Hydroponically Grown Lettuce, Bok Choy and Basil. Ph.D. Thesis, School of Graduate Studies, Rutgers The State University of New Jersey, Newark, NJ, USA, 2020. [Google Scholar]
- Lyons, G. Biofortification of cereals with foliar selenium and iodine could reduce hypothyroidism. Front. Plant Sci. 2018, 9, 730. [Google Scholar] [CrossRef]
- Golob, A.; Kroflič, A.; Jerše, A.; Kacjan Maršić, N.; Šircelj, H.; Stibilj, V.; Germ, M. Response of pumpkin to different concentrations and forms of selenium and iodine, and their combinations. Plants 2020, 9, 899. [Google Scholar] [CrossRef] [PubMed]
- Germ, M.; Stibilj, V.; Šircelj, H.; Jerše, A.; Kroflič, A.; Golob, A.; Maršić, N.K. Biofortification of common buckwheat microgreens and seeds with different forms of selenium and iodine. J. Sci. Food Agric. 2019, 99, 4353–4362. [Google Scholar] [CrossRef] [PubMed]
- Germ, M.; Kacjan-Maršić, N.; Kroflič, A.; Jerše, A.; Stibilj, V.; Golob, A. Significant accumulation of iodine and selenium in chicory (Cichorium intybus L. var. foliosum Hegi) leaves after foliar spraying. Plants 2020, 9, 1766. [Google Scholar] [CrossRef] [PubMed]
- Smoleń, S.; Baranski, R.; Ledwożyw-Smoleń, I.; Skoczylas, Ł.; Sady, W. Combined biofortification of carrot with iodine and selenium. Food Chem. 2019, 300, 125202. [Google Scholar] [CrossRef]
- Golob, A.; Novak, T.; Maršić, N.K.; Šircelj, H.; Stibilj, V.; Jerše, A.; Kroflič, A.; Germ, M. Biofortification with selenium and iodine changes morphological properties of Brassica oleracea L. var. gongylodes and increases their contents in tubers. Plant Physiol. Biochem. 2020, 150, 234–243. [Google Scholar] [CrossRef]
- Sahin, O. Combined biofortification of soilless grown lettuce with iodine, selenium and zinc and its effect on essential and non-essential elemental composition. J. Plant Nutr. 2020, 44, 673–678. [Google Scholar] [CrossRef]
- Şahin, Ö. Combined Iodine, Iron and Zinc Biofortification of Tomato Fruit. J. Inst. Sci. Technol. 2020, 10, 2242–2251. [Google Scholar] [CrossRef]
- Golubkina, N.; Gomez, L.D.; Kekina, H.; Cozzolino, E.; Simister, R.; Tallarita, A.; Torino, V.; Koshevarov, A.; Cuciniello, A.; Maiello, R.; et al. Joint Selenium-iodine supply and arbuscular mycorrhizal fungi inoculation affect yield and quality of chickpea seeds and residual biomass. Plants 2020, 9, 804. [Google Scholar] [CrossRef]
- Verlinden, S. Microgreens: Definitions, product types, and production practices. Hortic. Rev. 2020, 47, 85–124. [Google Scholar]
- Di Gioia, F.; Petropoulos, S.A.; Ozores-hampton, M.; Morgan, K.; Rosskopf, E.N. Zinc and Iron agronomic biofortification of brassicaceae microgreens. Agronomy 2019, 9, 677. [Google Scholar]
- Puccinelli, M.; Pezzarossa, B.; Pintimalli, L.; Malorgio, F. Selenium biofortification of three wild species, Rumex acetos L., Plantago coronopus L., and Portulaca oleracea L., grown as microgreens. Agronomy 2021, 11, 1155. [Google Scholar] [CrossRef]
- Newman, R.G.; Moon, Y.; Sams, C.E.; Tou, J.C.; Waterland, N.L. Biofortification of sodium selenate improves dietary mineral contents and antioxidant capacity of culinary herb microgreens. Front. Plant Sci. 2021, 12, 716437. [Google Scholar] [CrossRef]
- Mezeyová, I.; Hegedűsová, A.; Golian, M.; Andrejiová, A.; Šlosár, M.; Mezey, J. Influence of microgreens biofortification with selenium on their quantitative and qualitative parameters. Agronomy 2022, 12, 1096. [Google Scholar] [CrossRef]
- Van Der Straeten, D.; Bhullar, N.K.; de Steur, H.; Gruissem, W.; MacKenzie, D.; Pfeiffer, W.; Qaim, M.; Slamet-Loedin, I.; Strobbe, S.; Tohme, J.; et al. Multiplying the efficiency and impact of biofortification through metabolic engineering. Nat. Commun. 2020, 11, 5203. [Google Scholar] [CrossRef]
- Lowe, N.M. The global challenge of hidden hunger: Perspectives from the field. Proc. Nutr. Soc. 2021, 80, 283–289. [Google Scholar] [CrossRef]
- Saltzman, A.; Birol, E.; Oparinde, A.; Andersson, M.S.; Asare-Marfo, D.; Diressie, M.T.; Gonzalez, C.; Lividini, K.; Moursi, M.; Zeller, M. Availability, production, and consumption of crops biofortified by plant breeding: Current evidence and future potential. Ann. N. Y. Acad. Sci. 2017, 1390, 104–114. [Google Scholar] [CrossRef]
- Srivastav, P.; Vutukuru, M.; Ravindran, G.; Awad, M.M. Biofortification—Present scenario, possibilities and challenges: A scientometric approach. Sustainability 2022, 14, 11632. [Google Scholar] [CrossRef]
- Alfthan, G.; Eurola, M.; Ekholm, P.; Venäläinen, E.R.; Root, T.; Korkalainen, K.; Hartikainen, H.; Salminen, P.; Hietaniemi, V.; Aspila, P.; et al. Effects of nationwide addition of selenium to fertilizers on foods, and animal and human health in Finland: From deficiency to optimal selenium status of the population. J. Trace Elem. Med. Biol. 2015, 31, 142–147. [Google Scholar] [CrossRef]
- Baldassano, S.; Accardi, G.; Aiello, A.; Buscemi, S.; Di Miceli, G.; Galimberti, D.; Candore, G.; Ruisi, P.; Caruso, C.; Vasto, S. Fibres as functional foods and the effects on gut hormones: The example of β-glucans in a single arm pilot study. J. Funct. Foods 2018, 47, 264–269. [Google Scholar] [CrossRef]
- Muthayya, S.; Rah, J.H.; Sugimoto, J.D.; Roos, F.F.; Kraemer, K.; Black, R.E. The global hidden hunger indices and maps: An advocacy tool for action. PLoS ONE 2013, 8, e67860. [Google Scholar] [CrossRef]
- Trumbo, P.; Yates, A.A.; Schlicker, S.; Poos, M. Dietary reference intakes: Vitamin A, vitamin K, arsenic, boron, chromium, copper, iodine, iron, manganese, molybdenum, nickel, silicon, vanadium, and zinc. J. Am. Diet. Assoc. 2001, 101, 3–294. [Google Scholar] [CrossRef]
- Tsongas, T.A.; Meglen, R.R.; Walravens, P.A.; Chappel, W.R. Molybdenum in the diet: An estimate of average daily intake in the United States. Am. J. Clin. Nutr. 1980, 33, 1103–1107. [Google Scholar] [CrossRef]
- EFSA (European Food Safety Authority). Scientific opinion on dietary reference values for molybdenum. EFSA J. 2013, 11, 3333. [Google Scholar] [CrossRef]
- Martin, K.R. The chemistry of silica and its potential health benefits. J. Nutr. Health Aging 2007, 11, 94–97. [Google Scholar]
- Ahmed, T.; Hossain, M.; Sanin, K.I. Global burden of maternal and child undernutrition and micronutrient deficiencies. Ann. Nutr. Metab. 2012, 61, 8–17. [Google Scholar] [CrossRef]
- Amato, A.; Baldassano, S.; Cortis, C.; Cooper, J.; Proia, P. Physical activity, nutrition, and bone health. Hum. Mov. 2018, 19, 1–10. [Google Scholar] [CrossRef]
- Amato, A.; Proia, P.; Caldara, G.F.; Alongi, A.; Ferrantelli, V.; Baldassano, S. Analysis of body perception, preworkout meal habits and bone resorption in child gymnasts. Int. J. Environ. Res. Public Health 2021, 18, 2184. [Google Scholar] [CrossRef]
- Proia, P.; Amato, A.; Drid, P.; Korovljev, D.; Vasto, S.; Baldassano, S. The impact of diet and physical activity on bone health in children and adolescents. Front. Endocrinol. 2021, 12, 704647. [Google Scholar] [CrossRef]
- Olson, R.; Gavin-Smith, B.; Ferraboschi, C.; Kraemer, K. Food fortification: The advantages, disadvantages and lessons from sight and life programs. Nutrients 2021, 13, 1118. [Google Scholar] [CrossRef]
- Amato, A.; Baldassano, S.; Vasto, S.; Schirò, G.; Davì, C.; Drid, P.; Dos Santos Mendes, F.A.; Caldarella, R.; D’Amelio, M.; Proia, P. Effects of a resistance training protocol on physical performance, body composition, bone metabolism, and systemic homeostasis in patients diagnosed with Parkinson’s disease: A pilot study. Int. J. Environ. Res. Public Health 2022, 19, 13022. [Google Scholar] [CrossRef]
- Vasto, S.; Amato, A.; Proia, P.; Baldassano, S. Is the secret in the gut? SuperJump activity improves bone remodeling and glucose homeostasis by GLP-1 and GIP peptides in eumenorrheic women. Biology 2022, 11, 296. [Google Scholar] [CrossRef] [PubMed]
- Vasto, S.; Amato, A.; Proia, P.; Caldarella, R.; Cortis, C.; Baldassano, S. Dare to jump: The effect of the new high impact activity SuperJump on bone remodeling. A new tool to maintain fitness during COVID-19 home confinement. Biol. Sport 2022, 39, 1011–1019. [Google Scholar] [CrossRef] [PubMed]
- Juste Contin Gomes, M.; Stampini Duarte Martino, H.; Tako, E. Effects of iron and zinc biofortified foods on gut microbiota in vivo (Gallus gallus): A systematic review. Nutrients 2021, 13, 189. [Google Scholar] [CrossRef]
- Dias, D.M.; Kolba, N.; Binyamin, D.; Ziv, O.; Regini Nutti, M.; Martino, H.S.D.; Glahn, R.P.; Koren, O.; Tako, E. Iron biofortified carioca bean (Phaseolus vulgaris L.)—Based brazilian diet delivers more absorbable iron and affects the gut microbiota in vivo (Gallus gallus). Nutrients 2018, 10, 1970. [Google Scholar] [CrossRef] [PubMed]
- Reed, S.; Neuman, H.; Glahn, R.P.; Koren, O.; Tako, E. Characterizing the gut (Gallus gallus) microbiota following the consumption of an iron biofortified Rwandan cream seeded carioca (Phaseolus vulgaris L.) bean-based diet. PLoS ONE 2017, 12, e0182431. [Google Scholar] [CrossRef] [PubMed]
- Beasley, J.T.; Johnson, A.A.; Kolba, N.; Bonneau, J.P.; Glahn, R.P.; Ozeri, L.; Koren, O.; Tako, E. Nicotianamine-chelated iron positively affects iron status, intestinal morphology and microbial populations in vivo (Gallus gallus). Sci. Rep. 2020, 10, 2297. [Google Scholar] [CrossRef]
- Reed, S.; Knez, M.; Uzan, A.; Stangoulis, J.C.; Glahn, R.P.; Koren, O.; Tako, E. Alterations in the gut (Gallus gallus) microbiota following the consumption of zinc biofortified wheat (Triticum aestivum)-based diet. J. Agric. Food Chem. 2018, 66, 6291–6299. [Google Scholar] [CrossRef]
- D’Imperio, M.; Brunetti, G.; Gigante, I.; Serio, F.; Santamaria, P.; Cardinali, A.; Colucci, S.; Minervini, F. Integrated in vitro approaches to assess the bioaccessibility and bioavailability of silicon-biofortified leafy vegetables and preliminary effects on bone. Vitr. Cell. Dev. Biol. Anim. 2017, 53, 217–224. [Google Scholar] [CrossRef]
- Mottiar, Y.; Altosaar, I. Iodine sequestration by amy lose to combat iodine deficiency disorders. Trends Food Sci. Technol. 2011, 22, 335–340. [Google Scholar] [CrossRef]
- Vasto, S.; Di Gaudio, F.; Raso, M.; Sabatino, L.; Caldarella, R.; De Pasquale, C.; Di Rosa, L.; Baldassano, S. Impact on glucose homeostasis: Is food biofortified with molybdenum a workable solution? a two-arm study. Nutrients 2022, 14, 1351. [Google Scholar] [CrossRef]
- Mendoza, C. Effect of genetically modified low phytic acid plants on mineral absorption. Int.J. Food Sci. Technol. 2002, 37, 759–767. [Google Scholar] [CrossRef]
- Lin, L.; Ockenden, I.; Lott, J.N.A. The concentrations and distribution of phytic acid phosphorus and other mineral nutrients in wild-type and low phytic acid1-1 (lpa1-1) corn (Zea mays L.) grains and grain parts. Can. J. Bot. 2005, 83, 131–141. [Google Scholar] [CrossRef]
- Murgia, I.; Arosio, P.; Tarantino, D.; Soave, C. Biofortification for combating ‘hidden hunger’for iron. Trends Plant Sci. 2012, 17, 47–55. [Google Scholar] [CrossRef] [PubMed]
- Singh, U.; Praharaj, C.S.; Chaturvedi, S.K.; Bohra, A. Biofortification: Introduction, Approaches, Limitations, and Challenges in Biofortification of Food Crops; Springer: Berlin, Germany, 2016; pp. 3–18. [Google Scholar]
- Frossard, E.; Bucher, M.; Mächler, F.; Mozafar, A.; Hurrell, R. Potential for increasing the content and bioavailability of Fe, Zn and Ca in plants for human nutrition. J. Sci. Food Agric. 2000, 80, 861–879. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Microelement | Most Suitable Vegetable Crops for Biofortification |
|---|---|
| Fe | Rocket, red cabbage, and mustard microgreens |
| Zn | Rocket, red cabbage, mustard microgreens, broccoli, carrot, kale, and lettuce |
| I | Basil, cabbage, carrot, Chinese cabbage, cowpea, lettuce, mizuna, mustard, onion, potato, and tomato |
| Mo | Spinach and lettuce |
| Se | Basil, broccoli, carrot, chard, endive, garlic, Indian mustard, lettuce, onion, radish, spinach, and turnip |
| Si | Basil, chicory, mizuna, strawberry, and Swiss chard |
| Fe (mg/kg dw) | Zn (mg/kg dw) | |||
|---|---|---|---|---|
| Vegetables | Non-Biofortified | Biofortified | Non-Biofortified | Biofortified |
| Potato | 11 | 23 | 15 | 42 |
| Tomato | 448 | 656 | 15 | 30 |
| Lettuce | 180 | 400 | 145 | 220 |
| Carrot | 464 | 680 | 116.2 | 207.5 |
| Onion | No data | No data | 25.5 | 33.5 |
| I (mg/kg dw) | Mo (mg/kg dw) | |||
| Vegetables | Non-Biofortified | Biofortified | Non-Biofortified | Biofortified |
| Potato | 0.15 | 0.29 | No data | No data |
| Tomato | 0.03 | 0.23 | 0.02 | 0.07 |
| Lettuce | 20 | 780 | 50 | 580 |
| Carrot | 0.6 | 4.63 | No data | No data |
| Onion | 0.08 | 8.33 | No data | No data |
| Se (mg/kg dw) | Si (µg/g dw) | |||
| Vegetables | Non-Biofortified | Biofortified | Non-Biofortified | Biofortified |
| Potato | 5 | 27 | 26 | 50 |
| Tomato | 0.09 | 8.91 | No data | No data |
| Lettuce | 3 | 80 | No data | No data |
| Carrot | 2.21 | 10.97 | No data | No data |
| Onion | 0.03 | 9.96 | No data | No data |
| Microelement | RDA | UL |
|---|---|---|
| Fe | 8–18 mg [204] | 45 mg [204] |
| Zn | 9–14 mg [204] | 40 mg [204] |
| I | 150 µg [204] | 1100 µg [204] |
| Mo | 120–240 µg [205] | 600 µg [206] |
| Se | 55 µg [204] | 400 µg [204] |
| Si | Not established | 1700–1750 mg [207] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Consentino, B.B.; Ciriello, M.; Sabatino, L.; Vultaggio, L.; Baldassano, S.; Vasto, S.; Rouphael, Y.; La Bella, S.; De Pascale, S. Current Acquaintance on Agronomic Biofortification to Modulate the Yield and Functional Value of Vegetable Crops: A Review. Horticulturae 2023, 9, 219. https://doi.org/10.3390/horticulturae9020219
Consentino BB, Ciriello M, Sabatino L, Vultaggio L, Baldassano S, Vasto S, Rouphael Y, La Bella S, De Pascale S. Current Acquaintance on Agronomic Biofortification to Modulate the Yield and Functional Value of Vegetable Crops: A Review. Horticulturae. 2023; 9(2):219. https://doi.org/10.3390/horticulturae9020219
Chicago/Turabian StyleConsentino, Beppe Benedetto, Michele Ciriello, Leo Sabatino, Lorena Vultaggio, Sara Baldassano, Sonya Vasto, Youssef Rouphael, Salvatore La Bella, and Stefania De Pascale. 2023. "Current Acquaintance on Agronomic Biofortification to Modulate the Yield and Functional Value of Vegetable Crops: A Review" Horticulturae 9, no. 2: 219. https://doi.org/10.3390/horticulturae9020219
APA StyleConsentino, B. B., Ciriello, M., Sabatino, L., Vultaggio, L., Baldassano, S., Vasto, S., Rouphael, Y., La Bella, S., & De Pascale, S. (2023). Current Acquaintance on Agronomic Biofortification to Modulate the Yield and Functional Value of Vegetable Crops: A Review. Horticulturae, 9(2), 219. https://doi.org/10.3390/horticulturae9020219