
Identification of a Monosomic Alien Chromosome Addition Line Responsible for the Purple Color Trait in Heading Chinese Cabbage
 ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Growth Conditions
2.3. Analysis of Anthocyanin Content
2.3.1. Analysis of Anthocyanin Content by Colorimetric Method
2.3.2. HPLC–MS/MS Analyses
2.4. Fluorescence In Situ Hybridization (FISH)
2.4.1. Chromosome Preparation
2.4.2. DNA Probe Preparation
2.4.3. FISH
2.4.4. Image Processing
2.5. Real-Time Quantitative PCR (RT-qPCR)
2.6. Genotype Identification
2.7. Yeast-One Hybridization
3. Results
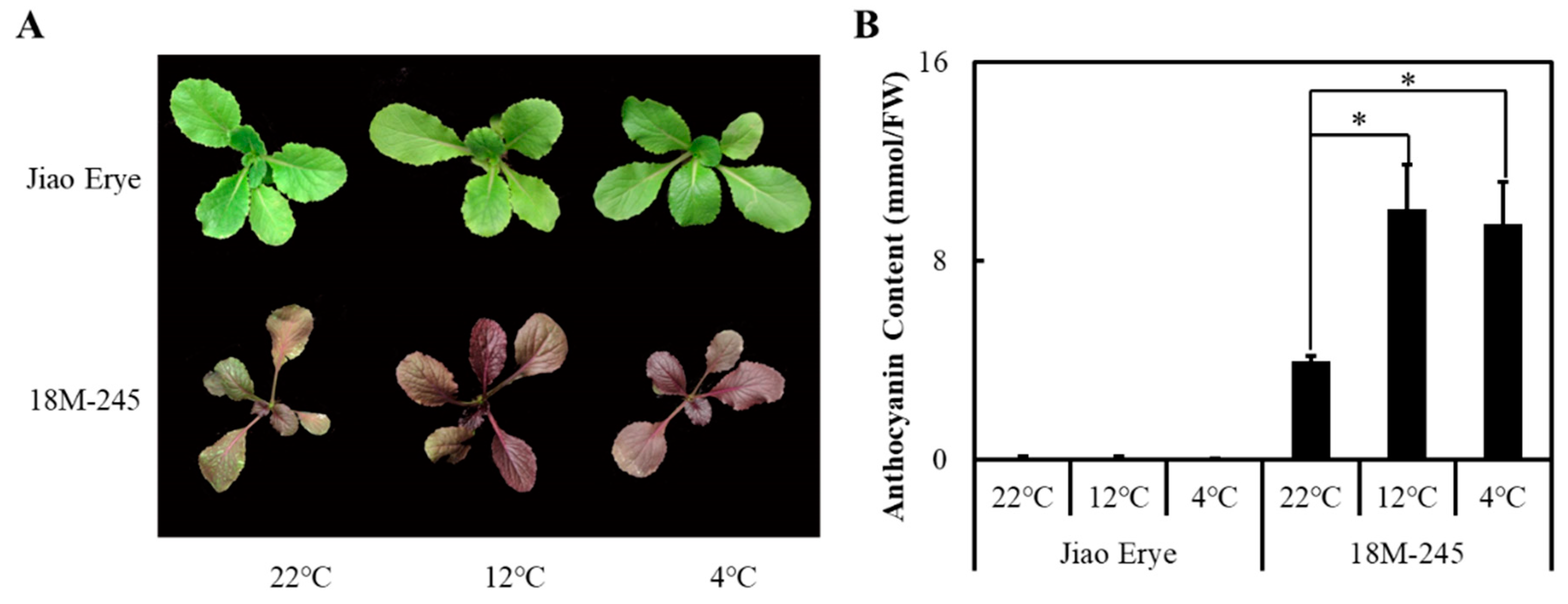
3.1. Generation of a New Purple Chinese Cabbage Germplasm
3.2. Anthocyanin Content in the 18M-245 Leaves
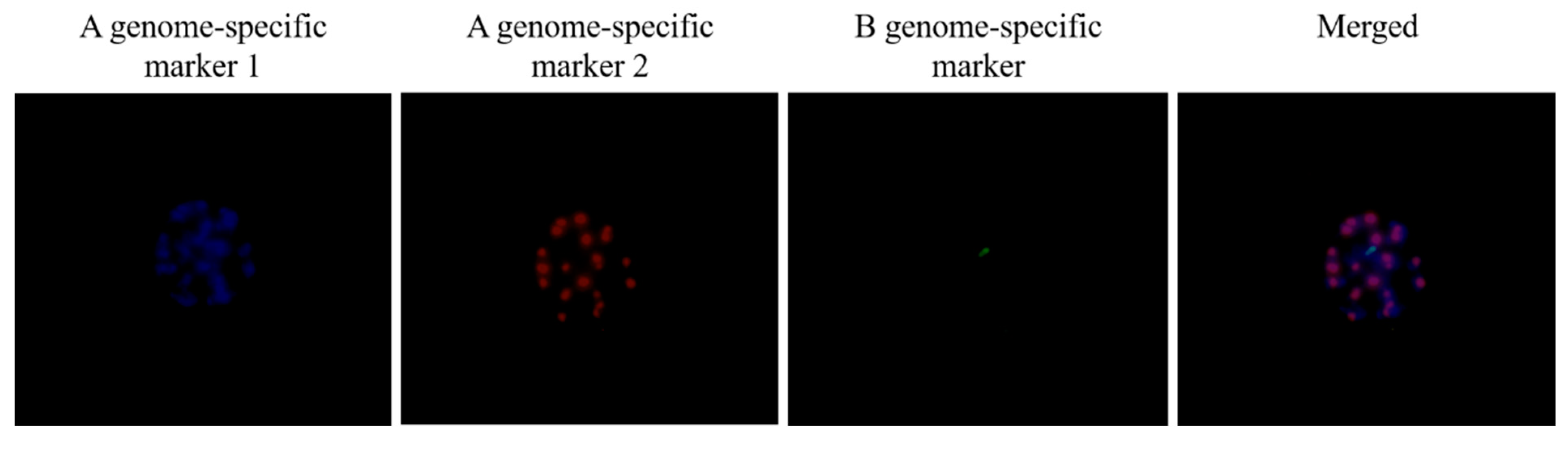
3.3. Identification of a Monosomic Alien Chromosome Addition Line in 18M-245
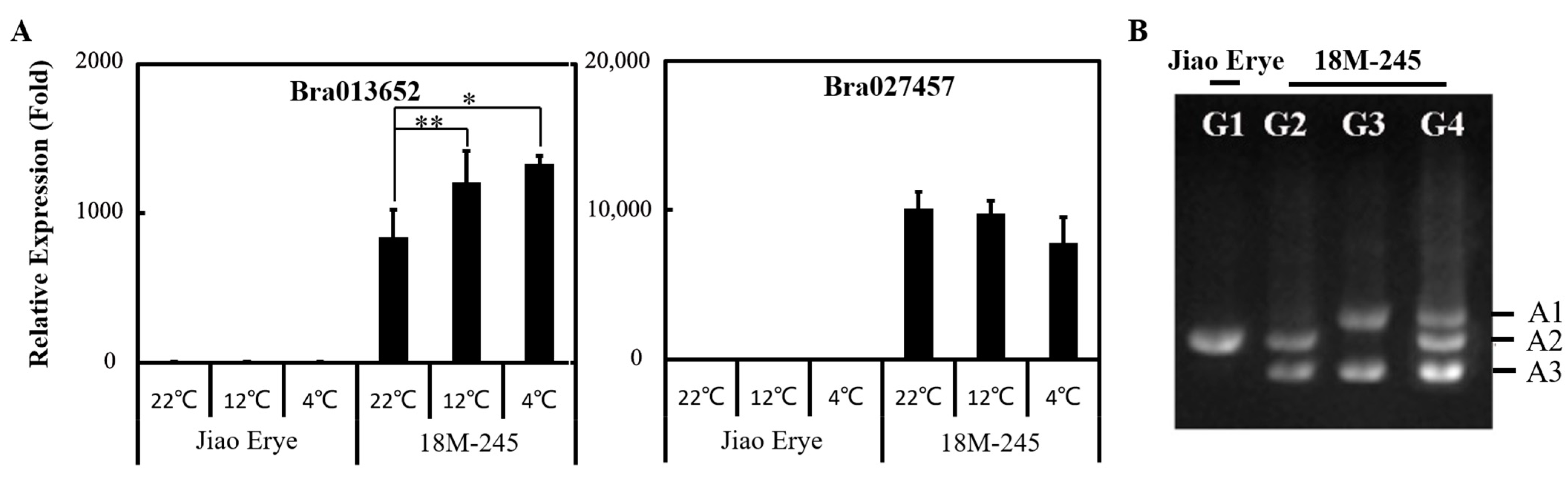
3.4. LDOX Transferred from the Brassica B Genome Is Highly Expressed in 18M-245
3.5. BjuB014115 Is Necessary for the Purple Color Trait in 18M-245
3.6. BjuB014115 Is Regulated by MYB and bHLH Transcription Factors from the Brassica A Genome
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Buer, C.S.; Imin, N.; Djordjevic, M.A. Flavonoids: New roles for old molecules. J. Integr. Plant Biol. 2010, 52, 98–111. [Google Scholar] [CrossRef] [PubMed]
- He, J.; Giusti, M.M. Anthocyanins: Natural colorants with health-promoting properties. Annu. Rev. Food Sci. Technol. 2010, 1, 163–187. [Google Scholar] [CrossRef] [PubMed]
- Winkel-Shirley, B. It takes a garden. How work on diverse plant species has contributed to an understanding of flavonoid metabolism. Plant Physiol. 2001, 127, 1399–1404. [Google Scholar] [CrossRef] [PubMed]
- Koes, R.; Verweij, W.; Quattrocchio, F. Flavonoids: A colorful model for the regulation and evolution of biochemical pathways. Trends Plant Sci. 2005, 10, 236–242. [Google Scholar] [CrossRef] [PubMed]
- Naing, A.H.; Kim, C.K. Roles of R2R3-MYB transcription factors in transcriptional regulation of anthocyanin biosynthesis in horticultural plants. Plant Mol. Biol. 2018, 98, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Pelletier, M.K.; Burbulis, I.E.; Winkel-Shirley, B. Disruption of specific flavonoid genes enhances the accumulation of flavonoid enzymes and end-products in Arabidopsis seedlings. Plant Mol. Biol. 1999, 40, 45–54. [Google Scholar] [CrossRef]
- Lepiniec, L.; Debeaujon, I.; Routaboul, J.M.; Baudry, A.; Pourcel, L.; Nesi, N.; Caboche, M. Genetics and biochemistry of seed flavonoids. Annu. Rev. Plant Biol. 2006, 57, 405–430. [Google Scholar] [CrossRef]
- Yan, H.; Pei, X.; Zhang, H.; Li, X.; Zhang, X.; Zhao, M.; Chiang, V.L.; Sederoff, R.R.; Zhao, X. MYB-mediated regulation of anthocyanin biosynthesis. Int. J. Mol. Sci. 2021, 22, 3103. [Google Scholar] [CrossRef]
- Xu, W.; Grain, D.; Bobet, S.; Le Gourrierec, J.; Thévenin, J.; Kelemen, Z.; Lepiniec, L.; Dubos, C. Complexity and robustness of the flavonoid transcriptional regulatory network revealed by comprehensive analyses of MYB-bHLH-WDR complexes and their targets in Arabidopsis seed. New Phytol. 2014, 202, 132–144. [Google Scholar] [CrossRef]
- Zhou, H.; Lin-Wang, K.; Wang, F.; Espley, R.V.; Ren, F.; Zhao, J.; Ogutu, C.; He, H.; Jiang, Q.; Allan, A.C.; et al. Activator-type R2R3-MYB genes induce a repressor-type R2R3-MYB gene to balance anthocyanin and proanthocyanidin accumulation. New Phytol. 2019, 221, 1919–1934. [Google Scholar] [CrossRef]
- Stracke, R.; Ishihara, H.; Huep, G.; Barsch, A.; Mehrtens, F.; Niehaus, K.; Weisshaar, B. Differential regulation of closely related R2R3-MYB transcription factors controls flavonol accumulation in different parts of the Arabidopsis thaliana seedling. Plant J. Cell Mol. Biol. 2007, 50, 660–677. [Google Scholar] [CrossRef]
- Zhang, F.; Gonzalez, A.; Zhao, M.; Payne, C.T.; Lloyd, A. A network of redundant bHLH proteins functions in all TTG1-dependent pathways of Arabidopsis. Development 2003, 130, 4859–4869. [Google Scholar] [CrossRef] [PubMed]
- Baudry, A.; Caboche, M.; Lepiniec, L. TT8 controls its own expression in a feedback regulation involving TTG1 and homologous MYB and bHLH factors, allowing a strong and cell-specific accumulation of flavonoids in Arabidopsis thaliana. Plant J. Cell Mol. Biol. 2006, 46, 768–779. [Google Scholar] [CrossRef]
- Feyissa, D.N.; Løvdal, T.; Olsen, K.M.; Slimestad, R.; Lillo, C. The endogenous GL3, but not EGL3, gene is necessary for anthocyanin accumulation as induced by nitrogen depletion in Arabidopsis rosette stage leaves. Planta 2009, 230, 747–754. [Google Scholar] [CrossRef] [PubMed]
- Appelhagen, I.; Jahns, O.; Bartelniewoehner, L.; Sagasser, M.; Weisshaar, B.; Stracke, R. Leucoanthocyanidin Dioxygenase in Arabidopsis thaliana: Characterization of mutant alleles and regulation by MYB-BHLH-TTG1 transcription factor complexes. Gene 2011, 484, 61–68. [Google Scholar] [CrossRef] [PubMed]
- Albert, N.W.; Davies, K.M.; Lewis, D.H.; Zhang, H.; Montefiori, M.; Brendolise, C.; Boase, M.R.; Ngo, H.; Jameson, P.E.; Schwinn, K.E. A conserved network of transcriptional activators and repressors regulates anthocyanin pigmentation in eudicots. Plant Cell 2014, 26, 962–980. [Google Scholar] [CrossRef]
- Dubos, C.; Le Gourrierec, J.; Baudry, A.; Huep, G.; Lanet, E.; Debeaujon, I.; Routaboul, J.M.; Alboresi, A.; Weisshaar, B.; Lepiniec, L. MYBL2 is a new regulator of flavonoid biosynthesis in Arabidopsis thaliana. Plant J. Cell Mol. Biol. 2008, 55, 940–953. [Google Scholar] [CrossRef]
- Matsui, K.; Umemura, Y.; Ohme-Takagi, M. AtMYBL2, a protein with a single MYB domain, acts as a negative regulator of anthocyanin biosynthesis in Arabidopsis. Plant J. Cell Mol. Biol. 2008, 55, 954–967. [Google Scholar] [CrossRef]
- Hayashi, K.; Matsumoto, S.; Tsukazaki, H.; Kondo, T.; Kubo, N.; Hirai, M. Mapping of a novel locus regulating anthocyanin pigmentation in Brassica rapa. Breed. Sci. 2010, 60, 76–80. [Google Scholar] [CrossRef]
- Wang, W.; Zhang, D.; Yu, S.; Liu, J.; Wang, D.; Zhang, F.; Yu, Y.; Zhao, X.; Lu, G.; Su, T. Mapping the BrPur gene for purple leaf color on linkage group A03 of Brassica rapa. Euphytica 2014, 199, 293–302. [Google Scholar] [CrossRef]
- He, Q.; Wu, J.; Xue, Y.; Zhao, W.; Li, R.; Zhang, L. The novel gene BrMYB2, located on chromosome A07, with a short intron 1 controls the purple-head trait of Chinese cabbage (Brassica rapa L.). Hortic. Res. 2020, 7, 97. [Google Scholar] [CrossRef] [PubMed]
- Guo, N.; Wu, J.; Zheng, S.; Cheng, F.; Liu, B.; Liang, J.; Cui, Y.; Wang, X. Anthocyanin profile characterization and quantitative trait locus mapping in zicaitai (Brassica rapa L. ssp. chinensis var. purpurea). Mol. Breed. 2015, 35, 113. [Google Scholar] [CrossRef]
- Zhang, X.; Zhang, K.; Wu, J.; Guo, N.; Liang, J.; Wang, X.; Cheng, F. QTL-Seq and sequence assembly rapidly mapped the gene BrMYBL2.1 for the purple trait in Brassica rapa. Sci. Rep. 2020, 10, 2328. [Google Scholar] [CrossRef] [PubMed]
- Sun, R.; Zhang, S.; Zhang, S.; Fei, L. Research on creation of Purple Chinese Cabbage germplasm. Acta Hortic. Sin. 2006, 33, 1032. [Google Scholar]
- Zhang, M.K.; Zhang, L.G.; Gong, Z.H.; Hui, M.X. Screening RAPD markers linked to purple trait of Chinese Cabbage and its chromosome location. Acta Bot. Boreali-Occident. Sin. 2008, 7, 82–88. [Google Scholar]
- Lee, H.; Oh, I.-N.; Kim, J.; Jung, D.; Cuong, N.P.; Kim, Y.; Lee, J.; Kwon, O.; Park, S.U.; Lim, Y.; et al. Phenolic compound profiles and their seasonal variations in new red-phenotype head-forming Chinese cabbages. LWT 2018, 90, 433–439. [Google Scholar] [CrossRef]
- Rameneni, J.J.; Choi, S.R.; Chhapekar, S.S.; Kim, M.S.; Singh, S.; Yi, S.Y.; Oh, S.H.; Kim, H.; Lee, C.Y.; Oh, M.H.; et al. Red Chinese cabbage transcriptome analysis reveals structural genes and multiple transcription factors regulating reddish purple color. Int. J. Mol. Sci. 2020, 21, 2901. [Google Scholar] [CrossRef]
- Xie, L.; Li, F.; Zhang, S.; Zhang, H.; Qian, W.; Li, P.; Zhang, S.; Sun, R. Mining for candidate genes in an introgression line by using RNA sequencing: The anthocyanin overaccumulation phenotype in Brassica. Front. Plant Sci. 2016, 7, 1245. [Google Scholar] [CrossRef]
- Zhou, R.N.; Shi, R.; Jiang, S.M.; Yin, W.B.; Wang, H.H.; Chen, Y.H.; Hu, J.; Wang, R.R.; Zhang, X.Q.; Hu, Z.M. Rapid EST isolation from chromosome 1R of rye. BMC Plant Biol. 2008, 8, 28. [Google Scholar] [CrossRef]
- Jiang, S.M.; Yin, W.B.; Hu, J.; Shi, R.; Zhou, R.N.; Chen, Y.H.; Zhou, G.H.; Wang, R.R.; Song, L.Y.; Hu, Z.M. Isolation of expressed sequences from a specific chromosome of Thinopyrum intermedium infected by BYDV. Genome 2009, 52, 68–76. [Google Scholar] [CrossRef]
- Teng, S.; Keurentjes, J.; Bentsink, L.; Koornneef, M.; Smeekens, S. Sucrose-specific induction of anthocyanin biosynthesis in Arabidopsis requires the MYB75/PAP1 gene. Plant Physiol. 2005, 139, 1840–1852. [Google Scholar] [CrossRef]
- Lei, L.; Li, Y.; Wang, Q.; Xu, J.; Chen, Y.; Yang, H.; Ren, D. Activation of MKK9-MPK3/MPK6 enhances phosphate acquisition in Arabidopsis thaliana. New Phytol. 2014, 203, 1146–1160. [Google Scholar] [CrossRef]
- Guzzo, F.; Campagnari, E.; Levi, M. A new FISH protocol with increased sensitivity for physical mapping with short probes in plants. J. Exp. Bot. 2000, 51, 965–970. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Zhao, H.; Zhang, F.; Yangjun, Y.U.; Zhao, X.; Shuancang, Y.U.; Wang, W.; Tongbing, S.U.; Guixiang, L.U. Identification and RNA-seq of new Purple Chinese Cabbage 15NG28 progenies. Acta Agric. Boreali-Sin. 2017, 6, 14–24. [Google Scholar]
- Yang, T.; Ma, H.; Zhang, J.; Wu, T.; Song, T.; Tian, J.; Yao, Y. Systematic identification of long noncoding RNAs expressed during light-induced anthocyanin accumulation in apple fruit. Plant J. 2019, 100, 572–590. [Google Scholar] [CrossRef]
- He, Q.; Ren, Y.; Zhao, W.; Li, R.; Zhang, L. Low temperature promotes anthocyanin biosynthesis and related gene expression in the seedlings of Purple Head Chinese Cabbage (Brassica rapa L.). Genes 2020, 11, 81. [Google Scholar] [CrossRef] [PubMed]
- Baudry, A.; Heim, M.A.; Dubreucq, B.; Caboche, M.; Weisshaar, B.; Lepiniec, L. TT2, TT8, and TTG1 synergistically specify the expression of BANYULS and proanthocyanidin biosynthesis in Arabidopsis thaliana. Plant J. Cell Mol. Biol. 2004, 39, 366–380. [Google Scholar] [CrossRef]
- Liu, C.; Jun, J.H.; Dixon, R.A. MYB5 and MYB14 Play Pivotal Roles in Seed Coat Polymer Biosynthesis in Medicago truncatula. Plant Physiol. 2014, 165, 1424–1439. [Google Scholar] [CrossRef]
- Liu, Z.; Wang, Y.; Fan, K.; Li, Z.; Jia, Q.; Lin, W.; Zhang, Y. Phytochrome-interacting factor 4 (PIF4) negatively regulates anthocyanin accumulation by inhibiting PAP1 transcription in Arabidopsis seedlings. Plant Sci. 2021, 303, 110788. [Google Scholar] [CrossRef]
- Xu, H.; Zou, Q.; Yang, G.; Jiang, S.; Fang, H.; Wang, Y.; Zhang, J.; Zhang, Z.; Wang, N.; Chen, X. MdMYB6 regulates anthocyanin formation in apple both through direct inhibition of the biosynthesis pathway and through substrate removal. Hortic. Res. 2020, 7, 72. [Google Scholar] [CrossRef]
- Lelivelt, C.L.C.; Lange, W.; Dolstra, O. Intergeneric crosses for the transfer of resistance to the beet cyst nematode from Raphanus sativus to Brassica napus. Euphytica 1993, 68, 111–120. [Google Scholar] [CrossRef]
- Zhang, S.; Li, P.; Qian, W.; Zhang, S.; Li, F.; Zhang, H.; Wang, X.; Sun, R. Mapping and expression profiling reveal an inserted fragment from purple mustard involved anthocyanin accumulation in Chinese cabbage. Euphytica 2016, 212, 83–95. [Google Scholar] [CrossRef]
- Gould, K.S. Nature’s swiss army knife: The diverse protective roles of anthocyanins in leaves. J. Biomed. Biotechnol. 2004, 2004, 314–320. [Google Scholar] [CrossRef]
- Nakabayashi, R.; Saito, K. Integrated metabolomics for abiotic stress responses in plants. Curr. Opin. Plant Biol. 2015, 24, 10–16. [Google Scholar] [CrossRef]
- Jiang, H.; Zhou, L.J.; Gao, H.N.; Wang, X.F.; Li, Z.W.; Li, Y.Y. The transcription factor MdMYB2 influences cold tolerance and anthocyanin accumulation by activating SUMO E3 ligase MdSIZ1 in apple. Plant Physiol. 2022, 189, 2044–2060. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Niu, Y.; Zheng, Y. Multiple functions of MYB transcription factors in abiotic stress responses. Int. J. Mol. Sci. 2021, 22, 6125. [Google Scholar] [CrossRef]
- Gonzalez, A.; Zhao, M.; Leavitt, J.M.; Lloyd, A.M. Regulation of the anthocyanin biosynthetic pathway by the TTG1/bHLH/Myb transcriptional complex in Arabidopsis seedlings. Plant J. Cell Mol. Biol. 2008, 53, 814–827. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Class | Compound | Content (ng/gDW) | Fold Change (18M-245 vs. Jiao Erye) | p Value | |
|---|---|---|---|---|---|
| Jiao Erye | 18M-245 | ||||
| Cyanidin | Cyanidin-3-O-glucoside | 31.88 | 26.39 | 0.83 | 0.702 |
| Cyanidin | Cyanidin-3,5,3’-O-triglucoside | 0.01 | 17.81 | 1191.97 | 0.005 |
| Cyanidin | Cyanidin-3-O-(6-O-p-coumaroyl)-glucoside | 0.00 | 5.95 | 0.148 | |
| Cyanidin | Cyanidin-3-O-sophoroside | 0.00 | 3.29 | 0.003 | |
| Cyanidin | Cyanidin-3,5-O-diglucoside | 0.00 | 2.47 | 0.001 | |
| Cyanidin | Cyanidin-3-(6′′-caffeylsophoroside)-5-glucoside | 0.00 | 1.19 | 0.002 | |
| Cyanidin | Cyanidin-3-O-5-O-(6-O-coumaroyl)-diglucoside | 0.00 | 0.99 | 0.019 | |
| Cyanidin | Cyanidin-3-(6-O-p-caffeoyl)-glucoside | 0.70 | 0.81 | 0.817 | |
| Cyanidin | Cyanidin-3-O-sambubioside-5-O-glucoside | 0.00 | 0.03 | 0.022 | |
| Cyanidin | Cyanidin-3-O-xyloside | 0.00 | 0.02 | 0.048 | |
| Cyanidin | Cyanidin-3-O-(6′′-ferulylsophoroside)-5-glucoside | 0.01 | 0.01 | 1.16 | 0.922 |
| Cyanidin | Cyanidin-3-O-sambubioside | 0.00 | 0.01 | 0.012 | |
| Pelargonidin | Pelargonidin-3-sophoroside-5-glucoside | 0.00 | 0.05 | 0.009 | |
| Pelargonidin | Pelargonidin-3-O-galactoside | 0.29 | 0.04 | 0.14 | 0.167 |
| Pelargonidin | Pelargonidin-3-O-glucoside | 0.00 | 0.03 | 0.007 | |
| Pelargonidin | Pelargonidin-3-O-sophoroside | 0.00 | 0.03 | 0.010 | |
| Pelargonidin | Pelargonidin-3-O-sophoroside-5-O-(malonyl)-glucoside | 0.00 | 0.02 | 0.040 | |
| Pelargonidin | Pelargonidin-3-O-(6-O-p-coumaroyl)-glucoside | 0.00 | 0.01 | 0.324 | |
| Pelargonidin | Pelargonidin-3-O-rutinoside | 0.01 | 0.00 | 0.15 | 0.165 |
| Peonidin | Peonidin-3,5-O-diglucoside | 0.01 | 0.06 | 4.44 | 0.018 |
| Peonidin | Peonidin-3-O-glucoside | 0.00 | 0.05 | 0.013 | |
| Peonidin | Peonidin-3-sophoroside-5-glucoside | 0.00 | 0.05 | 0.023 | |
| Peonidin | Peonidin-3-O-5-O-(6-O-coumaroyl)-diglucoside | 0.00 | 0.02 | 0.023 | |
| Peonidin | Peonidin-3-O-(6-O-p-coumaroyl)-glucoside | 0.00 | 0.01 | 0.015 | |
| Peonidin | Peonidin-3-(caffeoyl-glucosyl-glucoside)-5-glucoside | 0.00 | 0.00 | 0.044 | |
| Delphinidin | Delphinidin-3-O-sophoroside | 0.66 | 0.54 | 0.82 | 0.832 |
| Delphinidin | Delphinidin-3,5-O-diglucoside | 0.08 | 0.12 | 1.41 | 0.649 |
| Delphinidin | Delphinidin-3-O-(6-O-malonyl-beta-D-glucoside) | 0.00 | 0.04 | 8.78 | 0.088 |
| Delphinidin | Delphinidin-3-O-sambubioside | 0.00 | 0.01 | 0.016 | |
| Petunidin | Petunidin-3-O-(6-O-malonyl-beta-D-glucoside) | 0.03 | 0.04 | 1.29 | 0.763 |
| Petunidin | Petunidin-3-O-glucoside | 0.00 | 0.01 | 0.218 | |
| Petunidin | Petunidin-3-O-sambubioside | 0.00 | 0.00 | 0.00 | |
| Malvidin | Malvidin-3-O-(6-O-malonyl-beta-D-glucoside) | 0.00 | 0.05 | Inf | 0.012 |
| Malvidin | Malvidin | 0.00 | 0.00 | 0.23 | 0.525 |
| flavonoid | Kaempferol-3-O-rutinoside | 0.00 | 18.12 | 0.020 | |
| flavonoid | Quercetin-3-O-glucoside | 5.63 | 2.23 | 0.40 | 0.539 |
| flavonoid | Naringenin-7-O-glucoside | 0.21 | 0.13 | 0.65 | 0.760 |
| Gene ID | Gene Name | Green Reads Count | Purple Reads Count | log2Ratio (Green/Purple) | p-Value | FDR |
|---|---|---|---|---|---|---|
| Bra027796 | BrEGL3 | 90 | 176 | −1.06819 | 0.457398 | 1 |
| Bra037887 | BrTTG8 | 3 | 4408 | −10.5618 | 6.57 × 10−7 | 0.00207 |
| Bra009770 | BtTTG1 | 528 | 1069 | −1.11916 | 0.427705 | 1 |
| Bra007957 | BrMYBL2-1 | 4 | 463 | −6.911 | 0.000222 | 0.038438 |
| Bra016164 | BrMYBL2-2 | 45 | 2875 | −6.0951 | 0.000392 | 0.055515 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xin, X.; Zhang, D.; Zhao, H.; Su, T.; Zhao, X.; Wang, W.; Li, P.; Yu, Y.; Wang, J.; Yu, S.; et al. Identification of a Monosomic Alien Chromosome Addition Line Responsible for the Purple Color Trait in Heading Chinese Cabbage. Horticulturae 2023, 9, 146. https://doi.org/10.3390/horticulturae9020146
Xin X, Zhang D, Zhao H, Su T, Zhao X, Wang W, Li P, Yu Y, Wang J, Yu S, et al. Identification of a Monosomic Alien Chromosome Addition Line Responsible for the Purple Color Trait in Heading Chinese Cabbage. Horticulturae. 2023; 9(2):146. https://doi.org/10.3390/horticulturae9020146
Chicago/Turabian StyleXin, Xiaoyun, Deshuang Zhang, Hong Zhao, Tongbing Su, Xiuyun Zhao, Weihong Wang, Peirong Li, Yangjun Yu, Jiao Wang, Shuancang Yu, and et al. 2023. "Identification of a Monosomic Alien Chromosome Addition Line Responsible for the Purple Color Trait in Heading Chinese Cabbage" Horticulturae 9, no. 2: 146. https://doi.org/10.3390/horticulturae9020146
APA StyleXin, X., Zhang, D., Zhao, H., Su, T., Zhao, X., Wang, W., Li, P., Yu, Y., Wang, J., Yu, S., & Zhang, F. (2023). Identification of a Monosomic Alien Chromosome Addition Line Responsible for the Purple Color Trait in Heading Chinese Cabbage. Horticulturae, 9(2), 146. https://doi.org/10.3390/horticulturae9020146