
Study on Heat Resistance of Peony Using Photosynthetic Indexes and Rapid Fluorescence Kinetics

Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Materials and Experimental Design
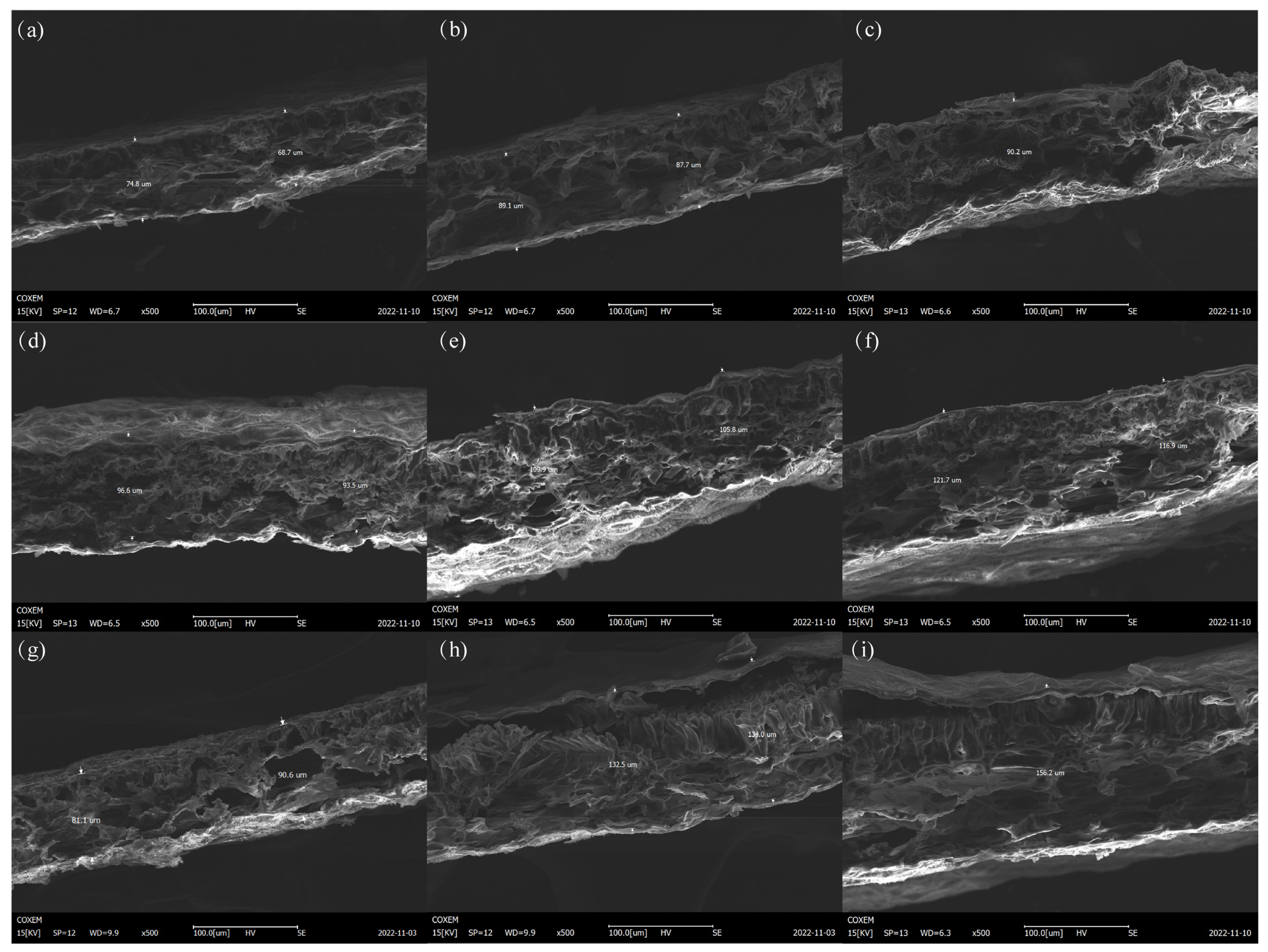
2.2. Electron Microscope Observation
2.3. Determination of Photosynthetic Parameters
2.4. Determination of Chlorophyll Fluorescence Kinetic Parameters
2.5. Statistical Analyses
3. Results
3.1. Changes in Leaf Morphology of Different Species of Peony under High-Temperature Stress
3.2. Effects of High-Temperature Stress on Photosynthetic Characteristic Curve Parameters of Different Varieties of Peony Leaves
3.3. Effects of High-Temperature Stress on the Rapid Chlorophyll Fluorescence Induction Kinetic Curves of Different Species of Peony
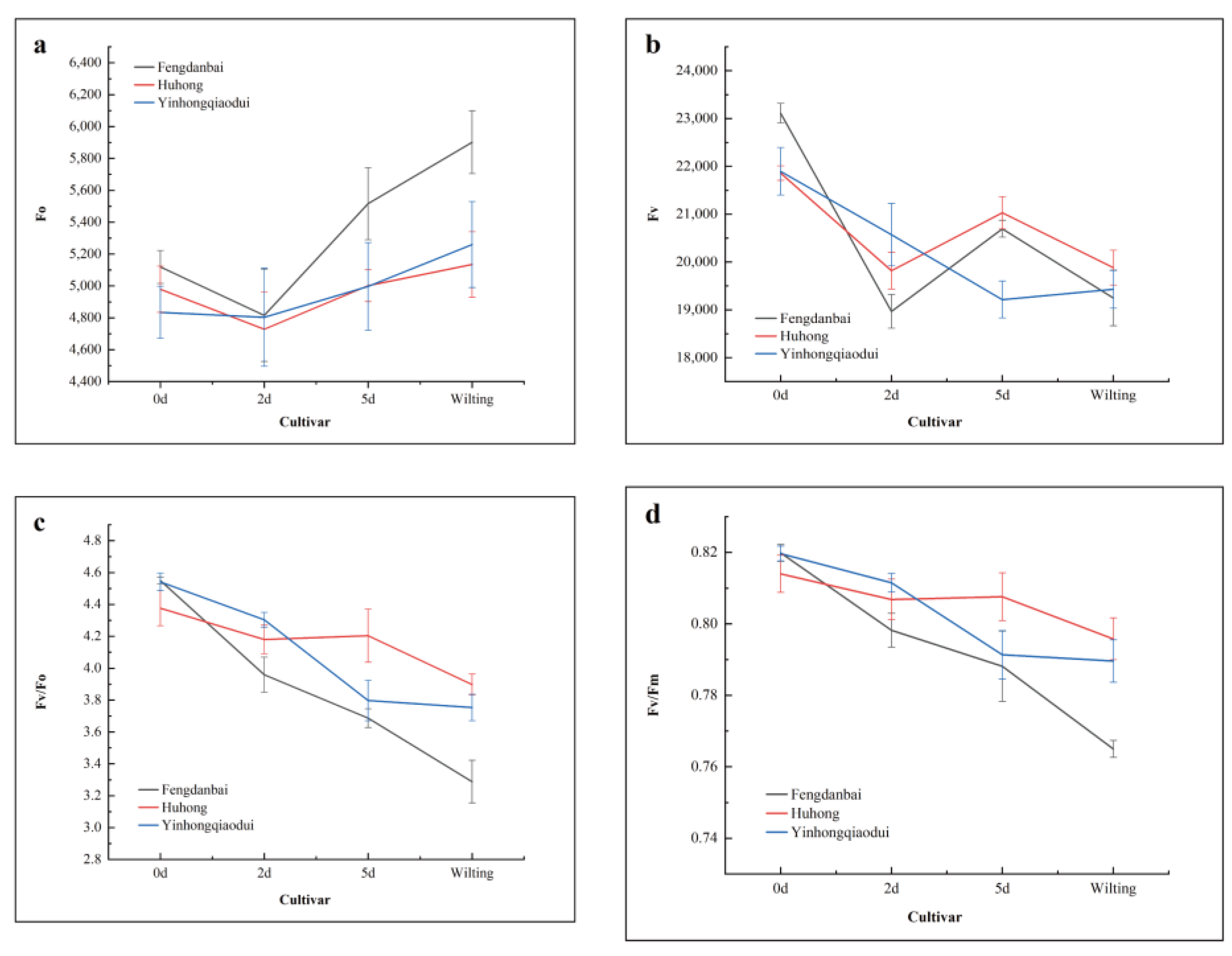
3.4. Effects of High-Temperature Stress on the Basic Parameters of Chlorophyll Fluorescence of Different Varieties of Peony Leaves
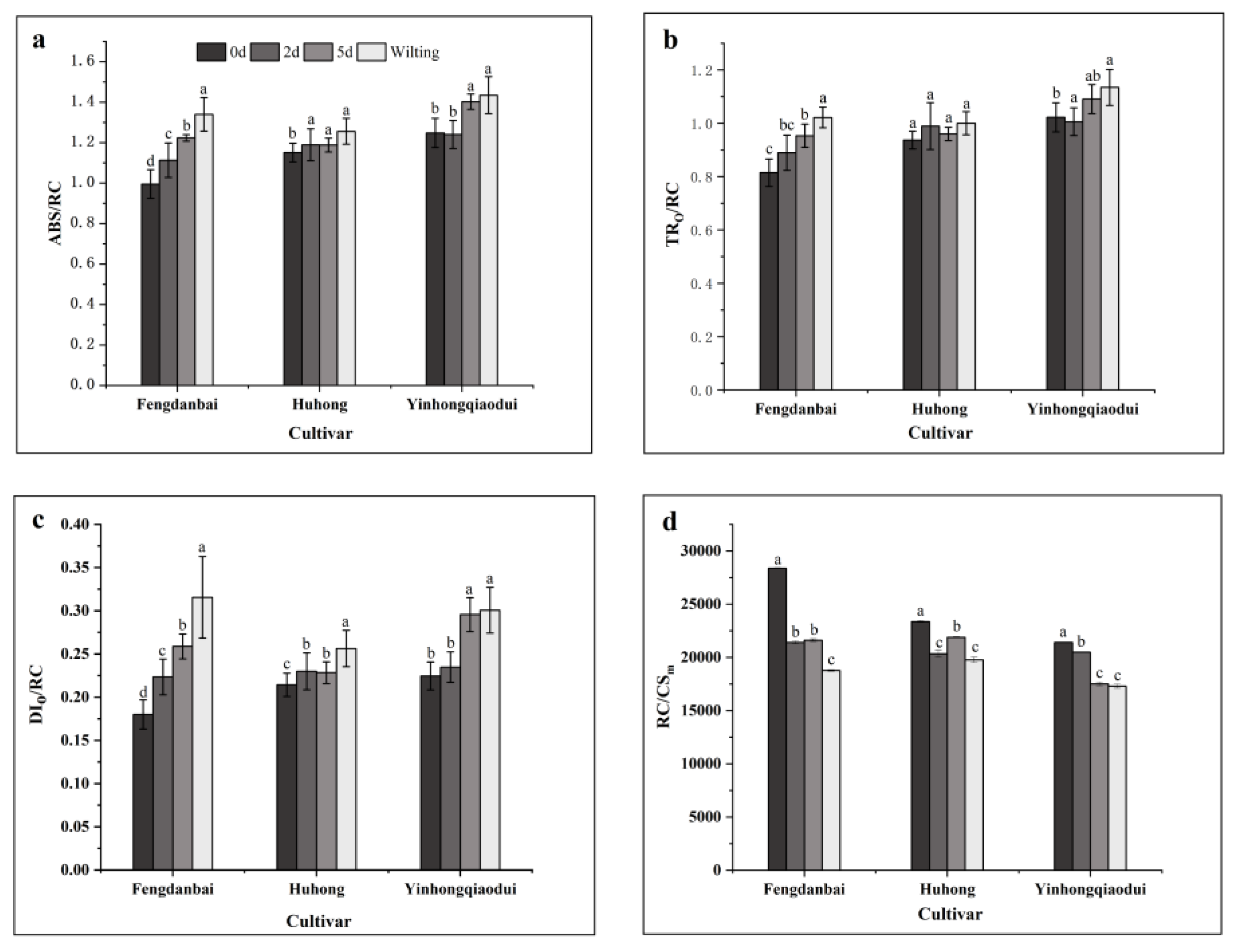
3.5. Effects of High-Temperature Stress on the Activity of PSII Reaction Centers of Peony Leaves of Different Species
3.6. Effects of High-Temperature Stress on the Sides of PSII Receptors in Leaves of Different Peony Species
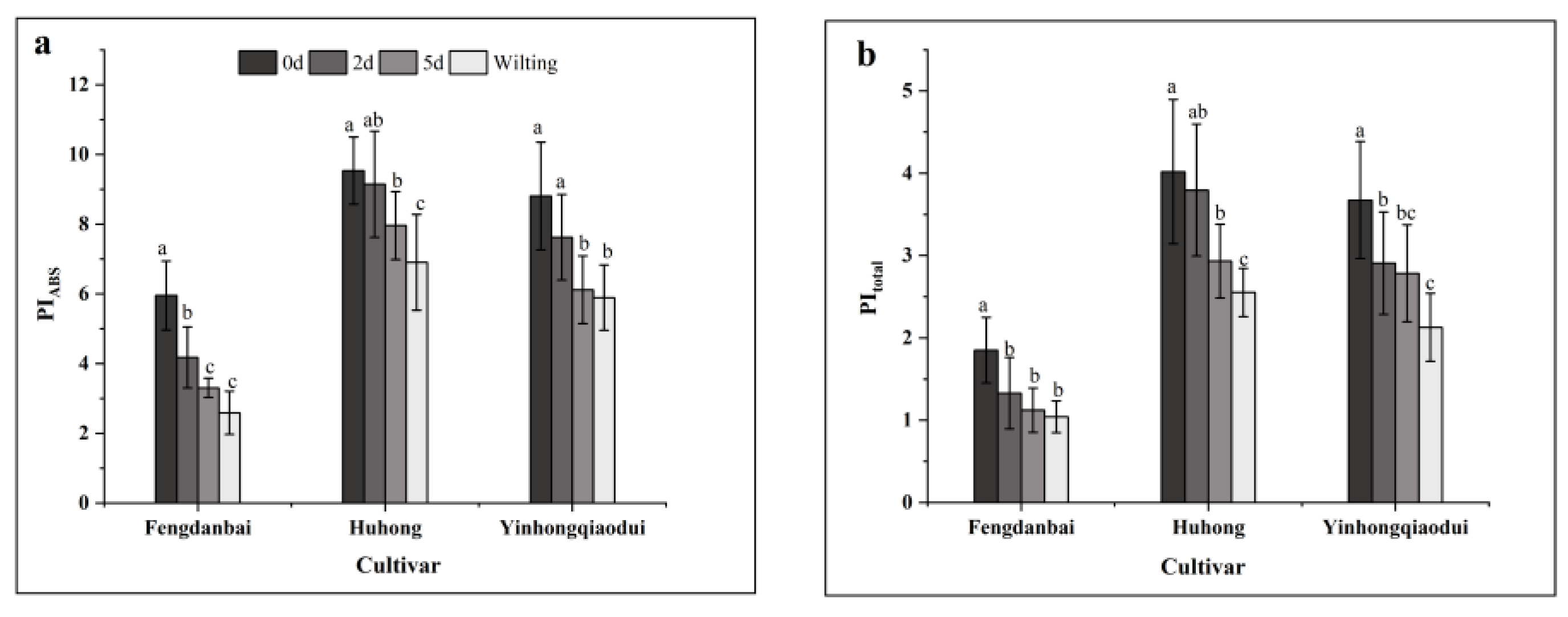
3.7. Effects of High-Temperature Stress on the Photochemical Performance Index of Different Varieties of Peony Leaves
3.8. Correlation of Chlorophyll Fluorescence Parameters of Peony Leaves under High-Temperature Stress
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wang, G. Distribution of peony in China. Bonsai Flowers Trees (Flower Gard.) 2000, 3, 5. [Google Scholar]
- Shen, S. Peony Should Be the Leader in the Industrialization of Famous Flowers—The Reaserch of Peony Industrialiazation of Heze. Master’s Thesis, Shandong University, Jinan, China, 2008. [Google Scholar]
- Zhou, D.; Zhang, M. High yield cultivation technique of peony. Mod. Agric. 2004, 3, 31. [Google Scholar]
- Wang, J. Genetic Diversity of Paeonia Ostii and Germplasm Resources of Tree Peony Cultivars from Chinese Jiangnan Area. Ph.D. Thesis, Beijing Forestry University, Beijing, China, 2009. [Google Scholar]
- Ashraf, M.H.; Harris, P.J. Photosynthesis under stressful environments: An overview. Photosynth. Int. J. Photosynth. Res. 2013, 51, 163–190. [Google Scholar] [CrossRef]
- Song, Q.; Feng, P.; Liu, Y.; Yang, X. The research progress of the mechanism on PSII assemble and repair circulation. Plant Physiol. Commun. 2019, 55, 133–140. [Google Scholar]
- Wang, B.; Tian, Z.; Ying, B.; Ma, Y.; Zuo, Z. Photosynthetic abilities in Cinnamomum camphora with high temperature stress. J. Zhejiang AF Univ. 2019, 36, 47–53. [Google Scholar]
- Xu, H.; Song, B.; Gu, Z. Advances in heat tolerance mechanisms of plants. Jiangsu J. Agric. Sci. 2020, 36, 243–250. [Google Scholar]
- Zahid, K.R.; Ali, F.; Shah, F.; Younas, M.; Shah, T.; Shahwar, D.; Hassan, W.; Ahmad, Z.; Qi, C.; Lu, Y.; et al. Response and Tolerance Mechanism of Cotton Gossypium hirsutum L. to Elevated Temperature Stress: A Review. Front. Plant Sci. 2016, 7, 937. [Google Scholar] [CrossRef] [PubMed]
- Xiao, F.; Yang, Z.Q.; Lee, K.W. Photosynthetic and physiological responses to high temperature in grapevine (Vitis vinifera L.) leaves during the seedling stage. J. Hortic. Sci. Biotechnol. 2016, 92, 2–10. [Google Scholar] [CrossRef]
- Hao, Z.; Zhou, C.; Liu, D.; Wei, M.; Tao, J. Effects of High Temperature Stress on Photosynthesi, Chlorophyll Fluorescence and Ultrastructure of Herbaceous Peony (Paeonia lactiflora Pall.). Mol. Plant Breed. 2017, 15, 2359–2367. [Google Scholar]
- Force, L.; Critchley, C.; van Rensen, J.J.S. New fluorescence parameters for monitoring photosynthesis in plants. Photosynth. Res. 2003, 78, 17. [Google Scholar] [CrossRef]
- Zhang, S. A Discussion on Chlorophyll Fluorescence Kinetics Parameters and Their Significance. Chin. Bull. Bot. 1999, 16, 444–448. [Google Scholar]
- Ren, S.; Li, Y.; Zou, J. Influence of High Temperature Stress on Chlorophyll Fluorescence Parameters in Four Kinds of Orchidaceae. North. Hortic. 2017, 12, 70–74. [Google Scholar]
- Zheng, Y.; He, T.; Chen, L.; Chen, L.; Rong, J.; Zheng, Y. Effects of high temperature stress on photosynthesis and chlorophyll fluorescence of Rhododendron hybridum. J. Fujian Agric. For. Univ. Nat. Sci. Ed. 2012, 41, 608–615. [Google Scholar]
- Chen, M.; Tang, Y. Chlorophyll fluorescence characteristics of Amaranthus tricolor L. under high temperature stress. Chin. J. Ecol. 2013, 32, 1813–1818. [Google Scholar]
- Yang, C.; Li, X.; Du, S. Damage mechanism of photosystems in winter wheat under high temperature. Chin. J. Eco-Agric. 2021, 30, 399–408. [Google Scholar] [CrossRef]
- Liu, J. Effects of High Temperature and Drought Stress on PSII Function and Light Distribution in Peony Leaves with Different Resistance. North. Hortic. 2019, 11, 8. [Google Scholar] [CrossRef]
- Liu, C.; Chen, D.; Gai, S. Effects of high-and low temperature stress on the leaf PSII functions and physiological characteristics of tree peony (Paeonia suffruticosa cv. ‘Roufurong’). Chin. J. Appl. Ecol. 2012, 23, 7. [Google Scholar]
- Qian, G.; Kong, X.; Zhang, S. Physiological response of three peony cultivars to high temperature stress. Jiangsu Agric. Sci. 2017, 45, 3. [Google Scholar]
- Luo, J.; Han, J.; Wang, Y. Response of Heat Stress on the Physiological Biochemistry of Paeonia suffruticosa. J. Yangtze Univ. Nat. Sci. Ed. 2011, 8, 4. [Google Scholar]
- Wu, S.; Jin, X.; Zhang, M.; Zhang, F.; Luo, F. Effects of Exogenous Abscisic Acid on Heat Tolerance in Tree Peony Seedlings under High Temperature Stress. Adv. Ornam. Hortic. China 2018, 2018, 354–360. [Google Scholar]
- Ren, Z.; Chen, F.; Shu, C.; Li, X.; Liu, K.; Ji, X. Effects of Exogenous 2,4-epibrassinolide on Heat Resistance of Peony. J. Jianghan Univ. Nat. Sci. Ed. 2018, 46, 446–453. [Google Scholar]
- Ji, W.; Luo, H.; Song, Y.; Hong, E.; Li, Z.; Lin, B.; Fan, C.; Wang, H.; Song, X.; Jin, S.; et al. Changes in Photosynthetic Characteristics of Paeonia suffruticosa under High Temperature Stress. Agronomy 2022, 12, 1203. [Google Scholar] [CrossRef]
- Ye, Z.; Suggett, D.J.; Robakowski, P.; Kang, H. A mechanistic model for the photosynthesis-light response based on the photosynthetic electron transport of photosystem II in C3and C4species. New Phytol. 2013, 199, 110–120. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Cheng, X. Physiological responses and regulatory pathways of transcription factors in plants under drought, high-salt, and low temperature stresses. Plant Nutr. Fertil. Sci. 2017, 23, 1565–1574. [Google Scholar]
- Wahid, A.; Gelani, S.; Ashraf, M.; Foolad, M.R. Heat tolerance in plants: An overview. Environ. Exp. Bot. 2007, 61, 199–223. [Google Scholar] [CrossRef]
- Zhang, S.; Ling, J. Effects of High Temperature Stress on Physiological Property of Grape Leaf in Greenhouse. Sci. Technol. Qinghai Agric. For. 2017, 2, 18–20+31. [Google Scholar]
- Wang, Y. Studies on Shade Tolerance and Application of Some Main Plant Species Used in Beijing. Ph.D. Thesis, Beijing Forestry University, Beijing, China, 1996. [Google Scholar]
- Zuo, L. Study on the Application of Tree Peony in Gardens. Master’s Thesis, Beijing Forestry University, Beijing, China, 2005. [Google Scholar]
- Qiu, Y.; Wang, W.; Hu, X. Relationship between Leaf Anatomical Structure and Heat Resistance of Rhododendron simsii. For. Environ. 2021, 37, 69–81. [Google Scholar]
- Crafts-Brandner, S.J.; Salvucci, M.E. Rubisco activase constrains the photosynthetic potential of leaves at high temperature and CO2. Proc. Natl. Acad. Sci. USA 2000, 97, 13430–13435. [Google Scholar] [CrossRef]
- Szymańska, R.; Ślesak, I.; Orzechowska, A.; Kruk, J. Physiological and biochemical responses to high light and temperature stress in plants. Environ. Exp. Bot. 2017, 139, 165–177. [Google Scholar] [CrossRef]
- Mathur, S.; Jajoo, A. Photosynthesis: Limitations in Response to High Temperature Stress. J. Photochem. Photobiol. B 2014, 137, 116–126. [Google Scholar] [CrossRef]
- Zhang, Y.; Yang, Z.; Lu, S. Response of Photosynthetic Characteristics of Leaves of Protected Chrysanthemum Variety ‘Jinbeidahon’ to High Temperature Stress. North. Hortic. 2021, 2021, 9. [Google Scholar]
- Xu, C.; Wang, M.T.; Yang, Z.Q.; Han, W.; Zheng, S.H. Effects of high temperature on photosynthetic physiological characteristics of strawberry seedlings in greenhouse and construction of stress level. Chin. J. Appl. Ecol. 2021, 32, 10. [Google Scholar]
- Wu, A.J.; Xu, W.Z.; Guo, Y.L.; Chen, J.; Li, S.; Xu, B.C. Photosynthetic Light-response Curves of Lespedeza davurica under Different Water and Fertilization Conditions. Acta Agrestia Sin. 2015, 23, 8. [Google Scholar]
- Zhou, Y.; Zhang, R.; Wang, S.X.; Wang, F.T.; Qi, Y. Comparative analysis on responses of vegetation productivity relative to different drought monitor patterns in Karst regions of southwestern China. Appl. Ecol. Environ. Res. 2019, 17, 85–105. [Google Scholar] [CrossRef]
- Wang, X.; Jin, W. Determination and analysis of Cypripedium japonicum shade tolerance. J. Nanjing For. Univ. Nat. Sci. Ed. 2014, 57, 57–61. [Google Scholar]
- Zhao, H.; Yang, Z.; Wang, M.; Wei, T.; Wang, L.; Sun, Q.; Zhang, X. Effects of high temperature and high humidity stress and restoration on the fast fluorescence induction dynamics of tomato leaves. Chin. J. Ecol. 2019, 38, 2405–2413. [Google Scholar]
- Bunce, J.A. Responses of stomatal conductance to light, humidity and temperature in winter wheat and barley grown at three concentrations of carbon dioxide in the field. Glob. Chang. Biol. 2000, 6, 371–382. [Google Scholar] [CrossRef]
- Krause, G.H.; Weis, E. ChlorophyII fluorescence and photosynthesis: The basics. Annu. Rev. Plant Physiol. 1991, 42, 313–349. [Google Scholar] [CrossRef]
- Yan, Z.; Ma, T.; Guo, S.; Liu, R.; Li, M. Leaf anatomy, photosynthesis and chlorophyll fluorescence of lettuce as influenced by arbuscular mycorrhizal fungi under high temperature stress. Sci. Hortic. 2021, 280, 109933. [Google Scholar] [CrossRef]
- Li, P.; Gao, H.; Reto, J. Strasser Application of the Fast Chlorophyll Fluorescence Induction Dynamics Analysis in Photosynthesis Study. J. Plant Physiol. Mol. Biol. 2005, 31, 559–566. [Google Scholar]
- Murata, N.; Takahashi, S.; Nishiyama, Y.; Allakhverdiev, S.I. Photoinhibition of photosystemII under environmental stress. Biochim. Biophys. Acta-Bioenerg. 2007, 1767, 414–421. [Google Scholar] [CrossRef] [PubMed]
- Jin, L.Q.; Che, X.K.; Zhang, Z.S.; Gao, H.Y. The Relationship between the Changes in Wk and Different Damage Degree of PSII Donor Side and Acceptor Side under High Temperature with High Light in Cucumber. Plant Physiol. Commun. 2015, 51, 969–976. [Google Scholar]
- Chang, C.; Zhang, D.; Hao, X. Effects of elevated CO2 concentration and increased temperature on the photosynthesis and fast chlorophyll fluorescence of winter wheat leaves. Plant Physiol. Commun. 2021, 57, 10. [Google Scholar]
- Liu, C.; Yuan, Y.; Gai, S.; Zhang, Y.; Liu, C.; Zheng, G. Effects of Strong Light Coupled with High Temperature Treatment on Energy Transfer Between PSII and PSI in Tree Peony Leaves. Acta Hortic. Sin. 2014, 41, 8. [Google Scholar]
- Henmi, T.; Miyao, M.; Yamamoto, Y. Release and reactive-oxygen-mediated damage of the oxygen-evolving complex subunits of PSII during photoinhibition. Plant Cell Physiol. 2004, 45, 243–250. [Google Scholar] [CrossRef]
- Yuan, J.L.; Ma, C.; Feng, Y.L.; Zhang, J.; Yang, F.Q.; Li, Y.J. Response of chlorophyll fluorescence transient in leaves of wheats with different drought resistances to drought stresses and rehydration. Plant Physiol. Commun. 2018, 54, 11. [Google Scholar]
- Strasser, B.J.; Strasser, R.J. Heat tolerance evaluation of different varieties of Heuchera spp. based on chlorophyll fluorescence parameters. Measuring Fast Fluorescence Transients to Address Environmental Questions: The JIP Test. In Photosynthesis: From Light to Biosphere; Mathis, P., Ed.; KAP Press: Dordrecht, The Netherlands, 1995; Volume 5, pp. 977–980. [Google Scholar]
- Sun, Y.; Li, S.; Zhang, Y. Heat tolerance evaluation of different varieties of Heuchera spp. based on chlorophyll fluorescence parameters. J. Cent. South Univ. For. Technol. 2021, 41, 11. [Google Scholar]
- Tao, M.; Feng, X.; He, Y.; Zhang, J.; Bai, X.; Yang, G.; Wei, Y. Time-series monitoring of transgenic maize seedlings phenotyping exhibiting glyphosate tolerance. Processes 2022, 10, 2206. [Google Scholar] [CrossRef]
- Stirbet, A.D.; Strasser, R.J. Numerical simulation of the in vivo fluorescence in plants. Math. Comput. Simul. 1996, 42, 245–253. [Google Scholar] [CrossRef]
- Mathur, S.; Mehta, P.; Jajoo, A. Effects of dual stress (high salt and high temperature) on the photochem ical efficiency of wheat leaves (Triticum aestivum). Physiol. Mol. Biol. Plants 2013, 19, 179–188. [Google Scholar] [CrossRef]
- Stefanov, D.; Petkova, V.; Denev, I.D. Screening for heat tolerance in common bean (Phaseolus vulgaris L.) lines and cultivars using JIP-test. Sci. Hortic. 2010, 128, 1–6. [Google Scholar] [CrossRef]
- Teng, Z.; Zhang, H.; Dai, X. Effects of drought stress on PSII photochemical activity in leaves of Morus alba. Acta Agric. Zhejiangensis 2016, 28, 1–8. [Google Scholar]
- Zhang, L.; Liu, J. Effects of heat stress on photosynthetic electron transport in a marine cyanobacterium Arthrospira sp. J. Appl. Phycol. 2016, 28, 757–763. [Google Scholar] [CrossRef]
- Mu, X.; Ma, Z.; Zhang, L.; Fu, L.; Liu, T.; Ding, Y.; Xia, L.; Zhang, F.; Zhang, J.; Qi, S.; et al. Responses of photosynthetic fluorescence characteristics, pollination, and yield components of maize cultivars to high temperature during flowering. Chin. J. Eco-Agric. 2022, 30, 57–71. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cultivar | Origin | Time to Leaf Wilting (d) | Time of Plant Death (d) |
|---|---|---|---|
| Fengdanbai | Shandong | 6 | 9 |
| Huhong | Shandong | 16 | / |
| Yinhongqiaodui | Shandong | 9 | 12 |
| Fengdanbai (μm) | Huhong (μm) | Yinhongqiaodui (μm) | |
|---|---|---|---|
| Untreated | 71.47 ± 3.09 c | 86.43 ± 4.86 c | 95.30 ± 1.61 c |
| Wilting | 88.07 ± 0.91 b | 132.10 ± 2.13 b | 107.47 ± 2.15 b |
| death | 91.43 ± 1.23 a | 150.53 ± 1.96 a | 119.47 ± 2.42 a |
| Cultivar | Stress Days | LSP | LCP | Rd | Pnmax | AQE |
|---|---|---|---|---|---|---|
| (μmol·m−2·s−1) | (μmol·m−2·s−1) | (μmol·m−2·s−1) | (μmol·m−2·s−1) | (μmol·m−2·s−1) | ||
| Fengdanbai | 0 | 526 ± 22.79 b | 0.3383 ± 0.10 d | 0.0286 ± 0.11 d | 5.9489 ± 0.14 a | 0.0175 ± 0.0001 a |
| 2 | 591 ± 12.01 a | 1.0768 ± 0.39 bc | 0.0325 ± 0.01 c | 1.3040 ± 0.05 d | 0.0025 ± 0.0003 d | |
| 5 | 567 ± 44.83 ab | 1.3513 ± 0.20 a | 0.0391 ± 0.02 b | 1.6740 ± 0.47 c | 0.0031 ± 0.0008 c | |
| 6 * | 502 ± 46.54 bc | 1.1913 ± 0.02 b | 0.0622 ± 0.01 a | 2.0135 ± 0.05 b | 0.0036 ± 0.0002 b | |
| Huhong | 0 | 501 ± 39.20 a | 6.3132 ± 1.50 c | 0.7930 ± 0.15 b | 3.8829 ± 0.79 b | 0.0100 ± 0.0022 ab |
| 2 | 476 ± 14.53 b | 7.1531 ± 0.28 b | 0.6958 ± 0.01 d | 4.3902 ± 0.06 a | 0.0114 ± 0.0001 a | |
| 5 | 486 ± 51.65 ab | 7.1175 ± 1.45 b | 0.7792 ± 0.17 bc | 3.5432 ± 0.14 c | 0.0106 ± 0.0031 a | |
| 16 * | 484 ± 49.89 ab | 7.9326 ± 1.64 a | 0.8982 ± 0.44 a | 3.7228 ± 0.10 b | 0.0062 ± 0.0053 b | |
| Yinhongqiaodui | 0 | 426 ± 21.21 c | 8.0760 ± 0.90 d | 0.8302 ± 0.09 a | 5.7344 ± 0.79 a | 0.0300 ± 0.0137 a |
| 2 | 543 ± 87.88 b | 14.7729 ± 0.89 c | 0.6056 ± 0.19 b | 3.9681 ± 0.81 b | 0.0203 ± 0.0032 b | |
| 5 | 612 ± 83.29 a | 38.7643 ± 2.70 a | 0.5933 ± 0.01 b | 0.2534 ± 0.02 d | 0.0048 ± 0.0006 c | |
| 9 * | 349 ± 21.24 d | 30.5128 ± 4.79 b | 0.6117 ± 0.16 b | 0.2834 ± 0.05 c | 0.0015 ± 0.0012 d |
| Index | LSP | Pnmax | AQE | LCP | Rd |
|---|---|---|---|---|---|
| LSP | 1 | ||||
| Pnmax | −0.184 | 1 | |||
| AQE | −0.193 | 0.821 ** | 1 | ||
| LCP | −0.112 | −0.537 | −0.173 | 1 | |
| Rd | −0.461 | 0.250 | 0.321 | 0.387 | 1 |
| Cultivar | Stress Days | Mo | Vj | Ψo | φEo |
|---|---|---|---|---|---|
| Fengdanbai | 0 | 0.359 ± 0.041 c | 0.438 ± 0.021 c | 0.562 ± 0.021 a | 0.452 ± 0.030 a |
| 2 | 0.427 ± 0.058 b | 0.467 ± 0.028 b | 0.533 ± 0.028 b | 0.422 ± 0.024 ab | |
| 5 | 0.453 ± 0.033 b | 0.479 ± 0.017 b | 0.527 ± 0.026 bc | 0.410 ± 0.012 b | |
| 6 * | 0.505 ± 0.020 a | 0.492 ± 0.019 a | 0.508 ± 0.019 c | 0.389 ± 0.021 c | |
| Huhong | 0 | 0.276 ± 0.010 ab | 0.293 ± 0.007 b | 0.707 ± 0.007 b | 0.577 ± 0.008 b |
| 2 | 0.297 ± 0.060 ab | 0.282 ± 0.015 b | 0.718 ± 0.015 a | 0.580 ± 0.009 a | |
| 5 | 0.297 ± 0.014 a | 0.307 ± 0.010 a | 0.693 ± 0.010 c | 0.560 ± 0.012 c | |
| 16 * | 0.297 ± 0.017 a | 0.310 ± 0.009 a | 0.690 ± 0.009 c | 0.503 ± 0.019 d | |
| Yinhongqiaodui | 0 | 0.304 ± 0.034 c | 0.296 ± 0.023 bc | 0.704 ± 0.023 a | 0.573 ± 0.030 a |
| 2 | 0.318 ± 0.033 bc | 0.315 ± 0.022 ab | 0.685 ± 0.022 b | 0.556 ± 0.016 b | |
| 5 | 0.331 ± 0.024 b | 0.310 ± 0.018 b | 0.690 ± 0.018 ab | 0.512 ± 0.124 c | |
| 9 * | 0.375 ± 0.030 a | 0.323 ± 0.020 a | 0.677 ± 0.020 bc | 0.544 ± 0.030 b |
| Fo | Fv | Fv/Fm | Fv/Fo | ABS/RC | TRo/RC | DIo/RC | RC/CSm | Ψo | φEo | Mo | Vj | PIABS | PItotal | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Fo | 1 | |||||||||||||
| Fv | 0.371 | 1 | ||||||||||||
| Fv/Fm | −0.321 | 0.730 ** | 1 | |||||||||||
| Fv/Fo | −0.330 | 0.727 ** | 0.999 ** | 1 | ||||||||||
| ABS/RC | −0.266 | −0.850 ** | −0.653 ** | −0.640 * | 1 | |||||||||
| TRo/RC | −0.059 | −0.660 * | −0.502 ** | −0.496 | 0.842 ** | 1 | ||||||||
| DIo/RC | 0.174 | −0.719 ** | −0.784 ** | −0.778 ** | 0.838 ** | 0.915 ** | 1 | |||||||
| RC/CSm | 0.196 | 0.860 ** | 0.628 ** | 0.627 * | −0.907 ** | −0.940 ** | −0.899 ** | 1 | ||||||
| Ψo | −0.636 * | −0.517 | 0.039 | 0.043 | 0.548 | 0.655 * | 0.403 | −0.669 * | 1 | |||||
| φEo | −0.661 * | −0.370 | 0.220 | 0.222 | 0.433 | 0.572 | 0.259 | −0.545 | 0.961 ** | 1 | ||||
| Mo | 0.613 * | 0.204 | −0.320 | −0.322 | −0.216 | −0.380 | 0.116 | 0.354 | −0.915 ** | −0.894 ** | 1 | |||
| Vj | 0.646 * | 0.519 | −0.041 | −0.044 | −0.551 | −0.649 * | −0.397 | 0.665 * | −1.000 ** | −0.961 ** | 0.915 ** | 1 | ||
| PIABS | −0.824 ** | −0.034 | 0.593 * | 0.598 * | −0.023 | −0.012 | −0.322 | −0.015 | 0.716 ** | 0.791 ** | −0.817 ** | −0.721 ** | 1 | |
| PItotal | −0.776 ** | −0.187 | 0.437 | 0.439 | 0.182 | 0.251 | −0.063 | −0.268 | 0.848 ** | 0.882 ** | −0.871 ** | −0.850 ** | 0.937 ** | 1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Z.; Ji, W.; Hong, E.; Fan, Z.; Lin, B.; Xia, X.; Chen, X.; Zhu, X. Study on Heat Resistance of Peony Using Photosynthetic Indexes and Rapid Fluorescence Kinetics. Horticulturae 2023, 9, 100. https://doi.org/10.3390/horticulturae9010100
Li Z, Ji W, Hong E, Fan Z, Lin B, Xia X, Chen X, Zhu X. Study on Heat Resistance of Peony Using Photosynthetic Indexes and Rapid Fluorescence Kinetics. Horticulturae. 2023; 9(1):100. https://doi.org/10.3390/horticulturae9010100
Chicago/Turabian StyleLi, Zhijun, Wen Ji, Erman Hong, Zhen Fan, Bangyu Lin, Xuanze Xia, Xia Chen, and Xiangtao Zhu. 2023. "Study on Heat Resistance of Peony Using Photosynthetic Indexes and Rapid Fluorescence Kinetics" Horticulturae 9, no. 1: 100. https://doi.org/10.3390/horticulturae9010100
APA StyleLi, Z., Ji, W., Hong, E., Fan, Z., Lin, B., Xia, X., Chen, X., & Zhu, X. (2023). Study on Heat Resistance of Peony Using Photosynthetic Indexes and Rapid Fluorescence Kinetics. Horticulturae, 9(1), 100. https://doi.org/10.3390/horticulturae9010100