
Selection and Validation of miRNA Reference Genes by Quantitative Real-Time PCR Analysis in Paeonia suffruticosa

 , ,
, ,
Abstract
1. Introduction
2. Methods
2.1. Plant Materials
2.2. Total miRNA Isolation and cDNA Synthesis
2.3. Selection of Candidate Reference miRNAs and Primer Design
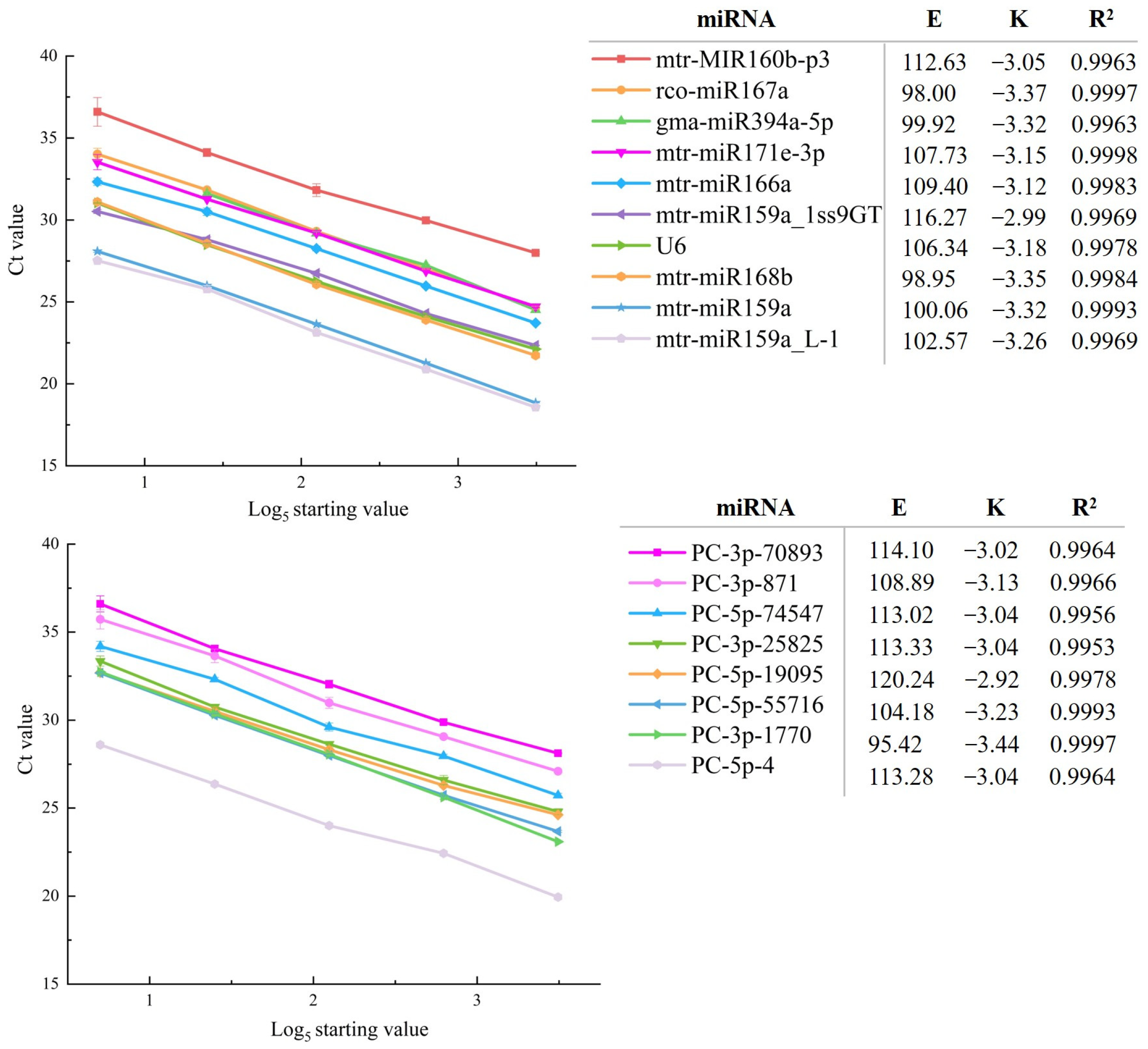
2.4. Drawing of Standard Curve and Verification of Primer Amplification Efficiency
2.5. qRT-PCR of Candidate Reference miRNAs
2.6. Assessment of Expression Stability of Candidate Reference miRNAs
2.7. Validation of Reference miRNAs
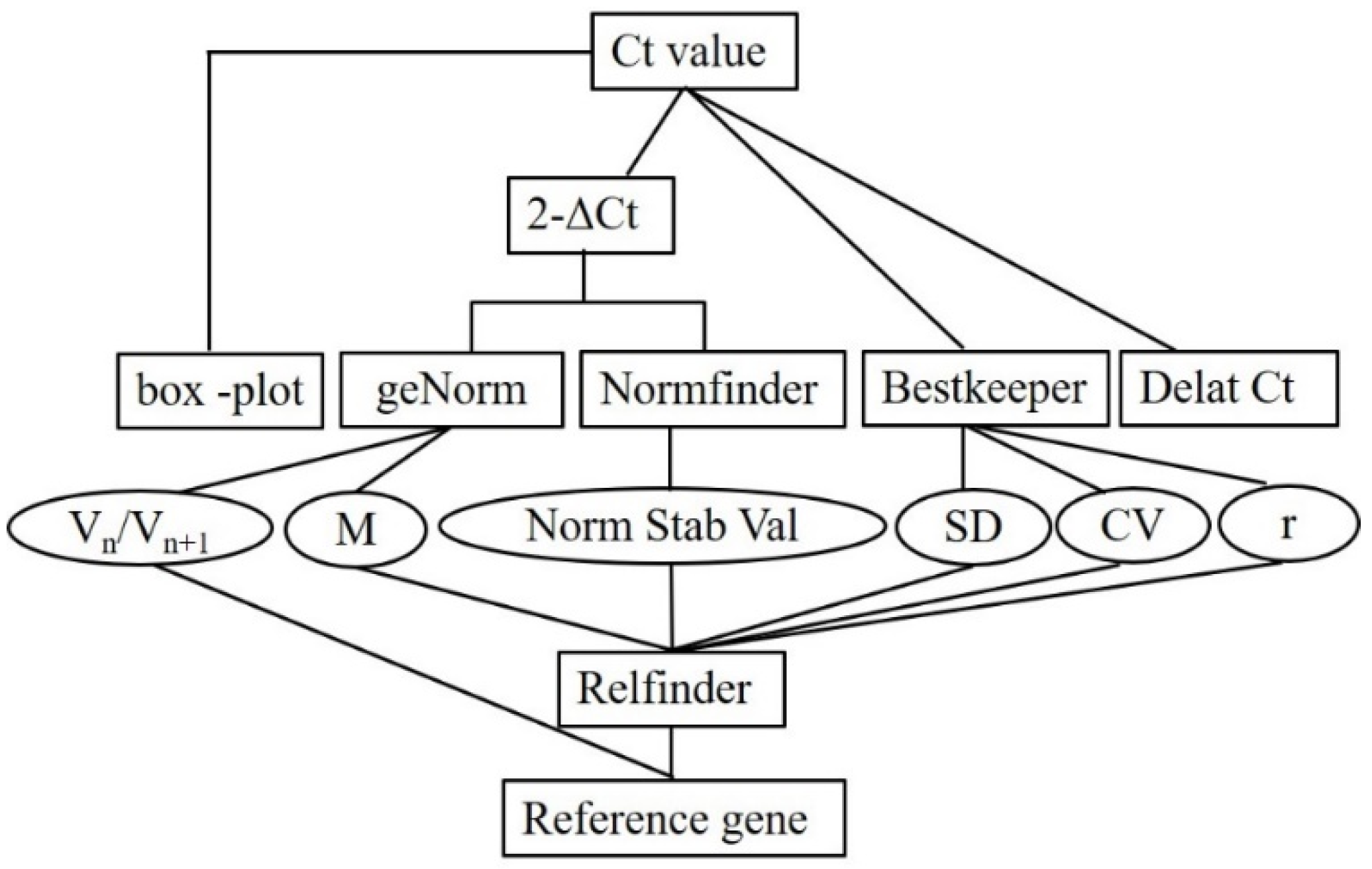
2.8. Date Analyses
3. Results
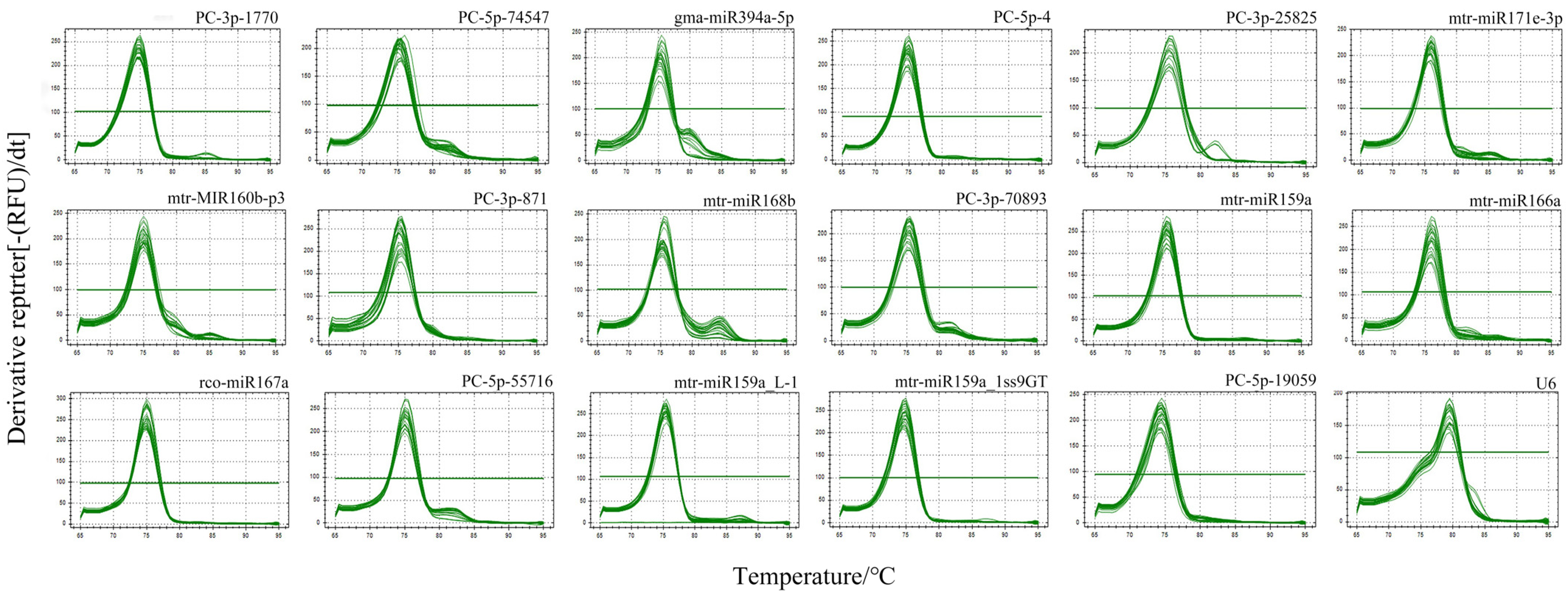
3.1. Evaluation of Primer Specificity and Test of Amplification Efficiency
3.2. Expression Levels of Candidate Reference miRNAs
3.3. Stability Analysis of Candidate Reference miRNAs
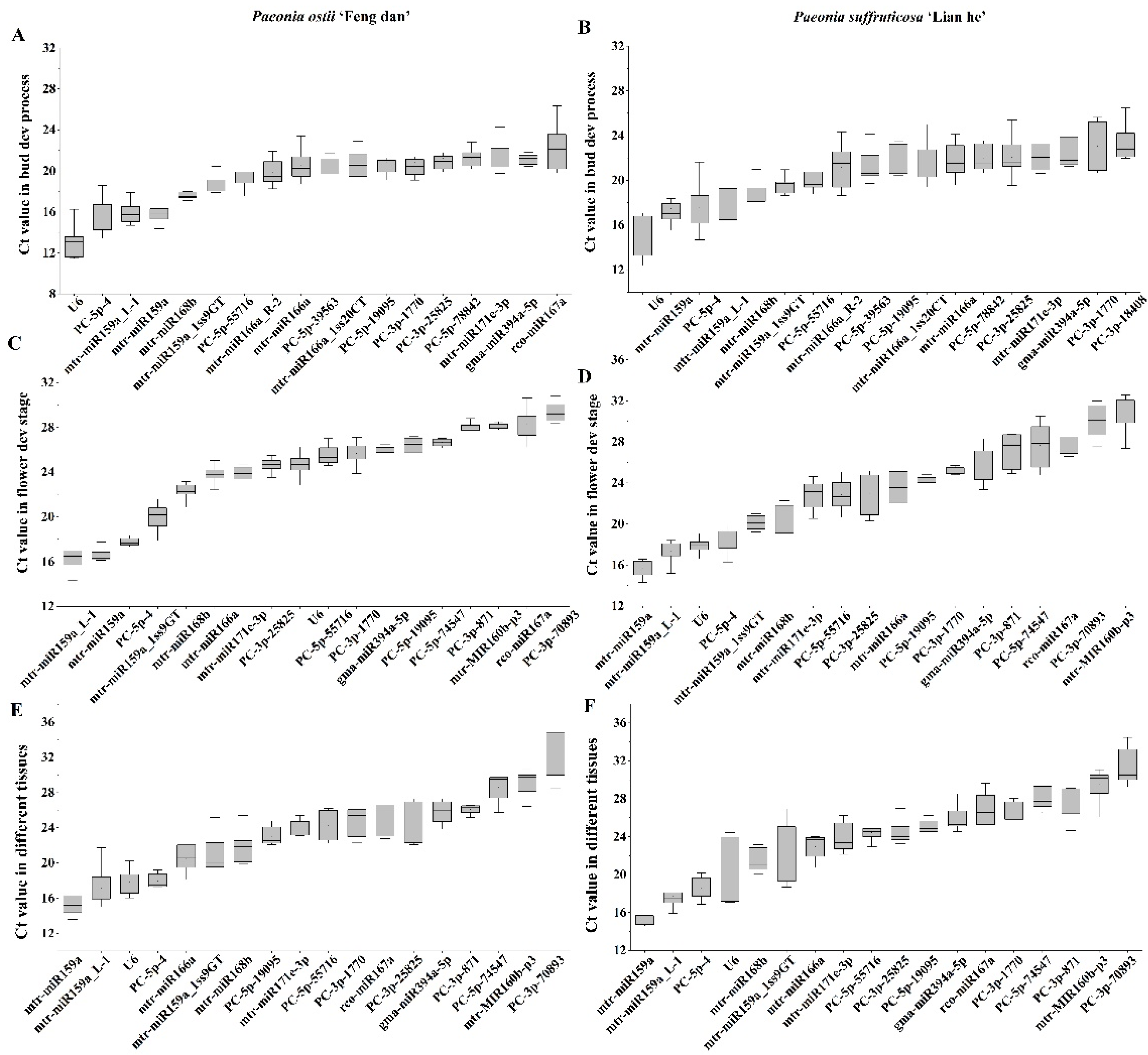
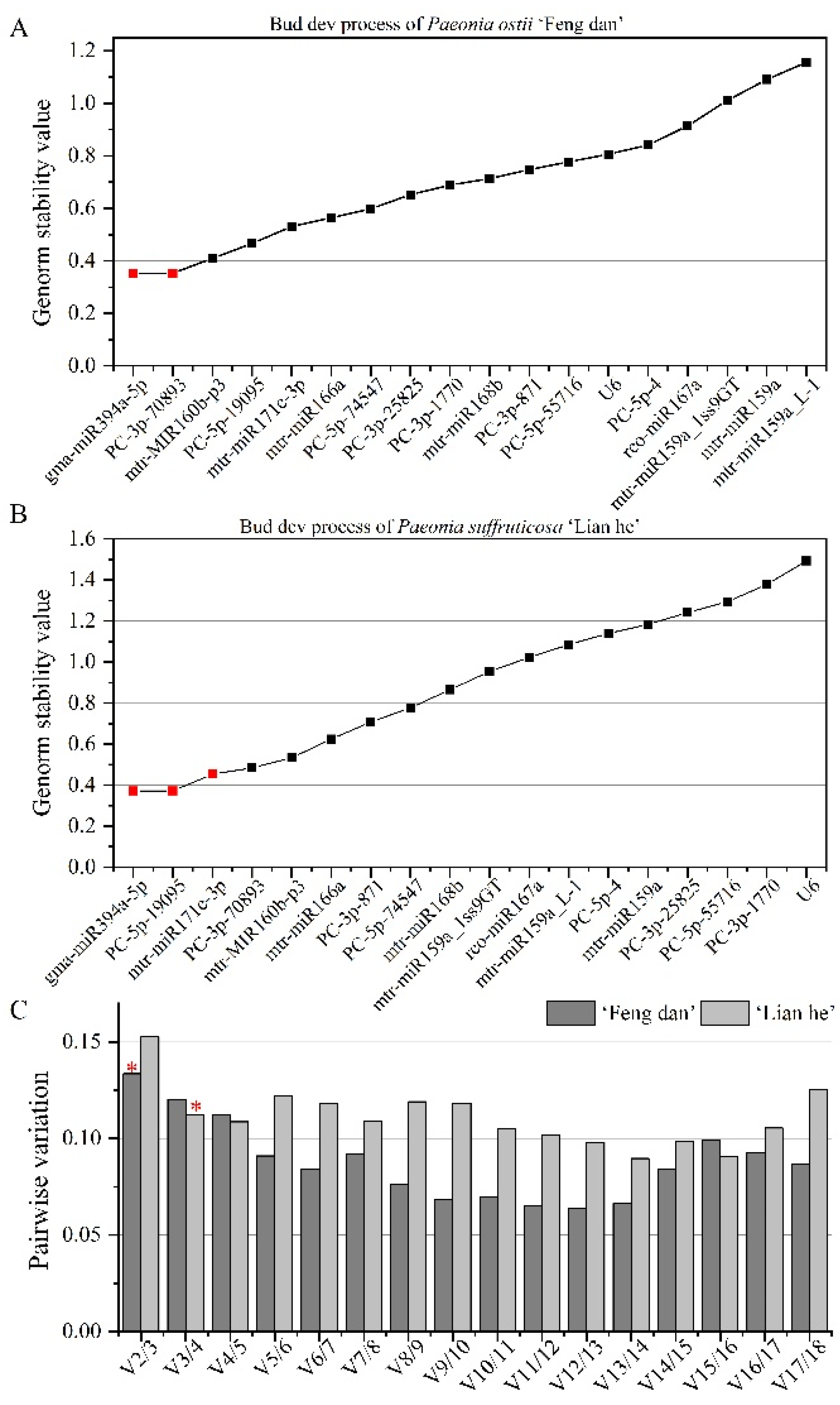
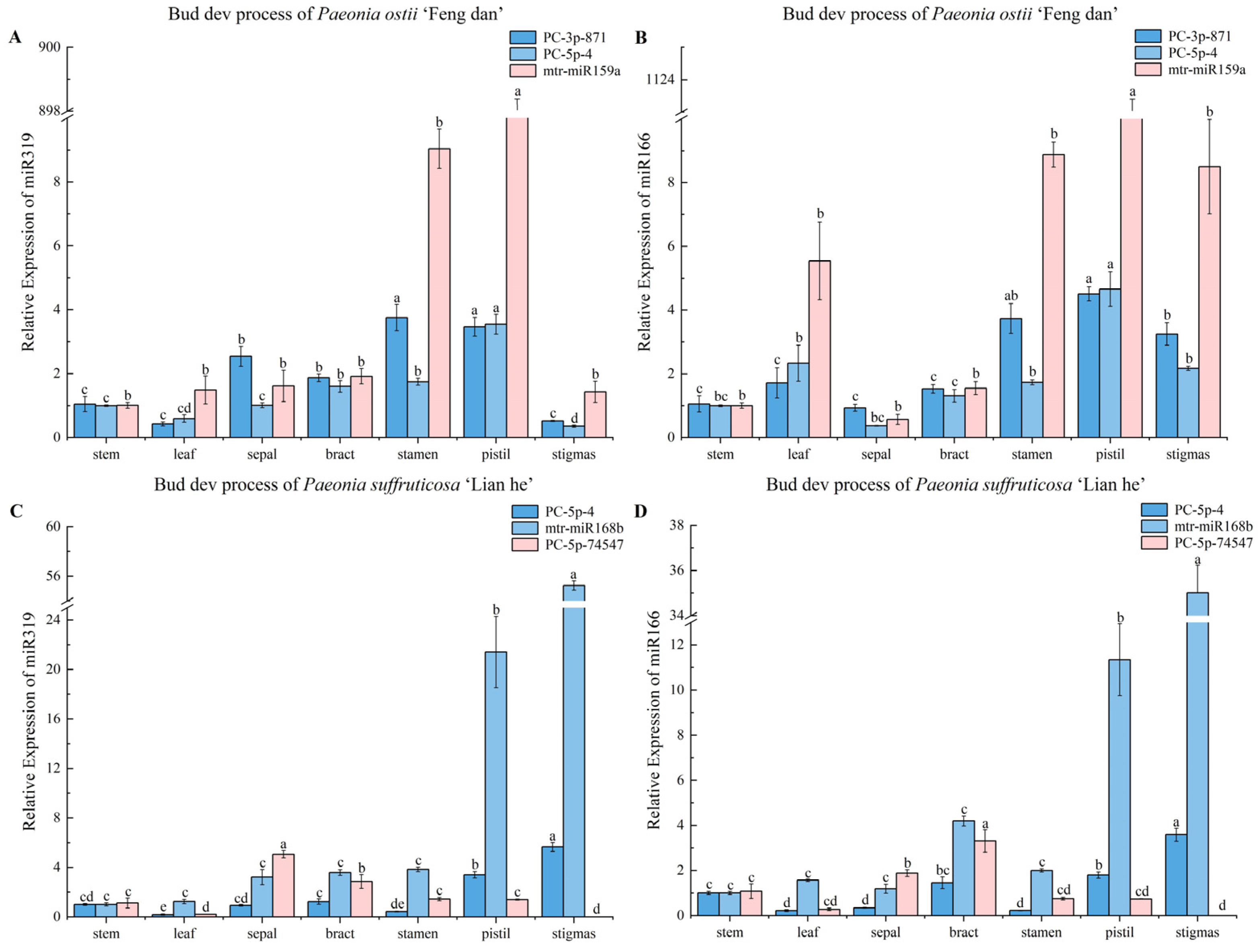
3.3.1. Stability Analysis of Candidate Reference miRNAs during the Bud Development Process
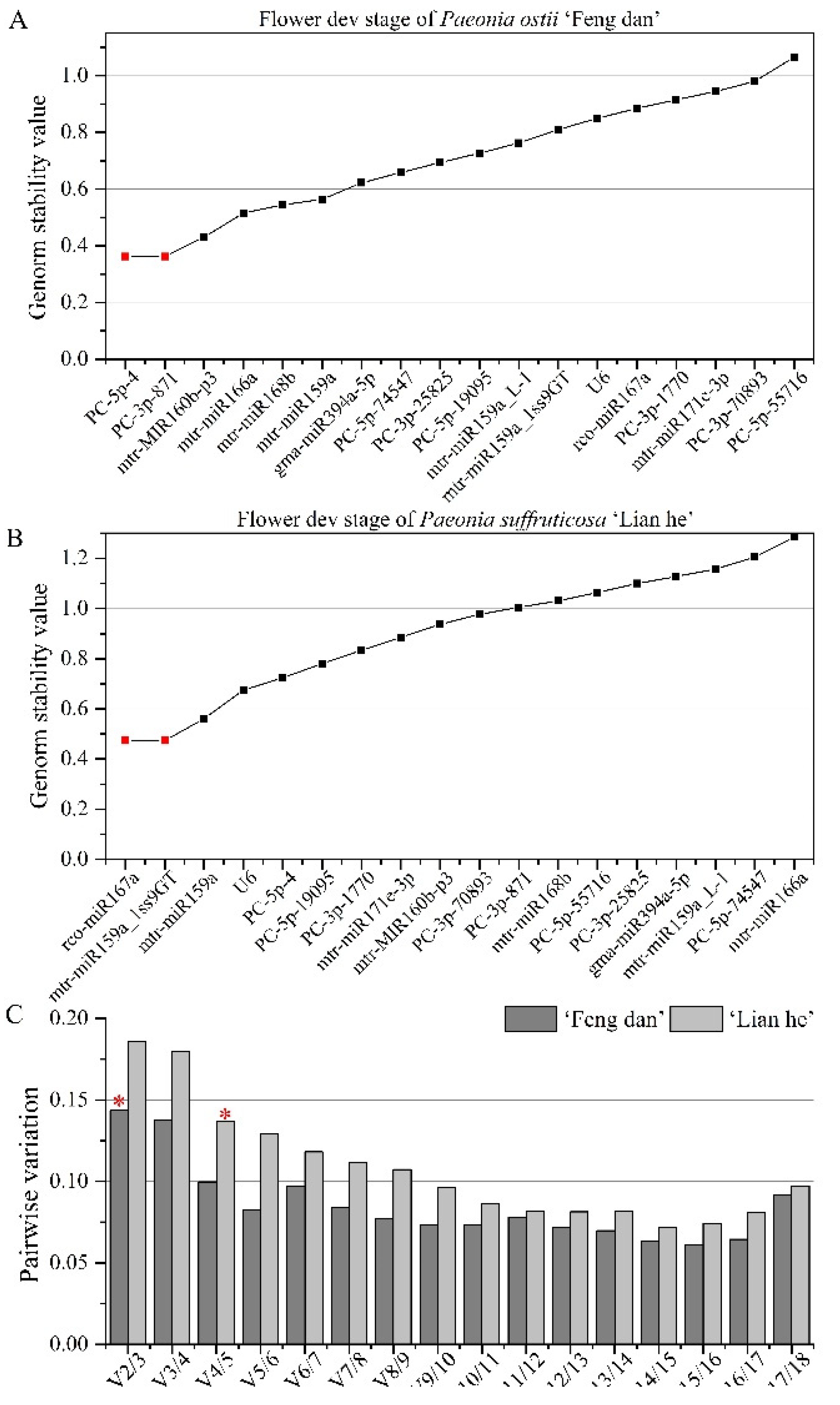
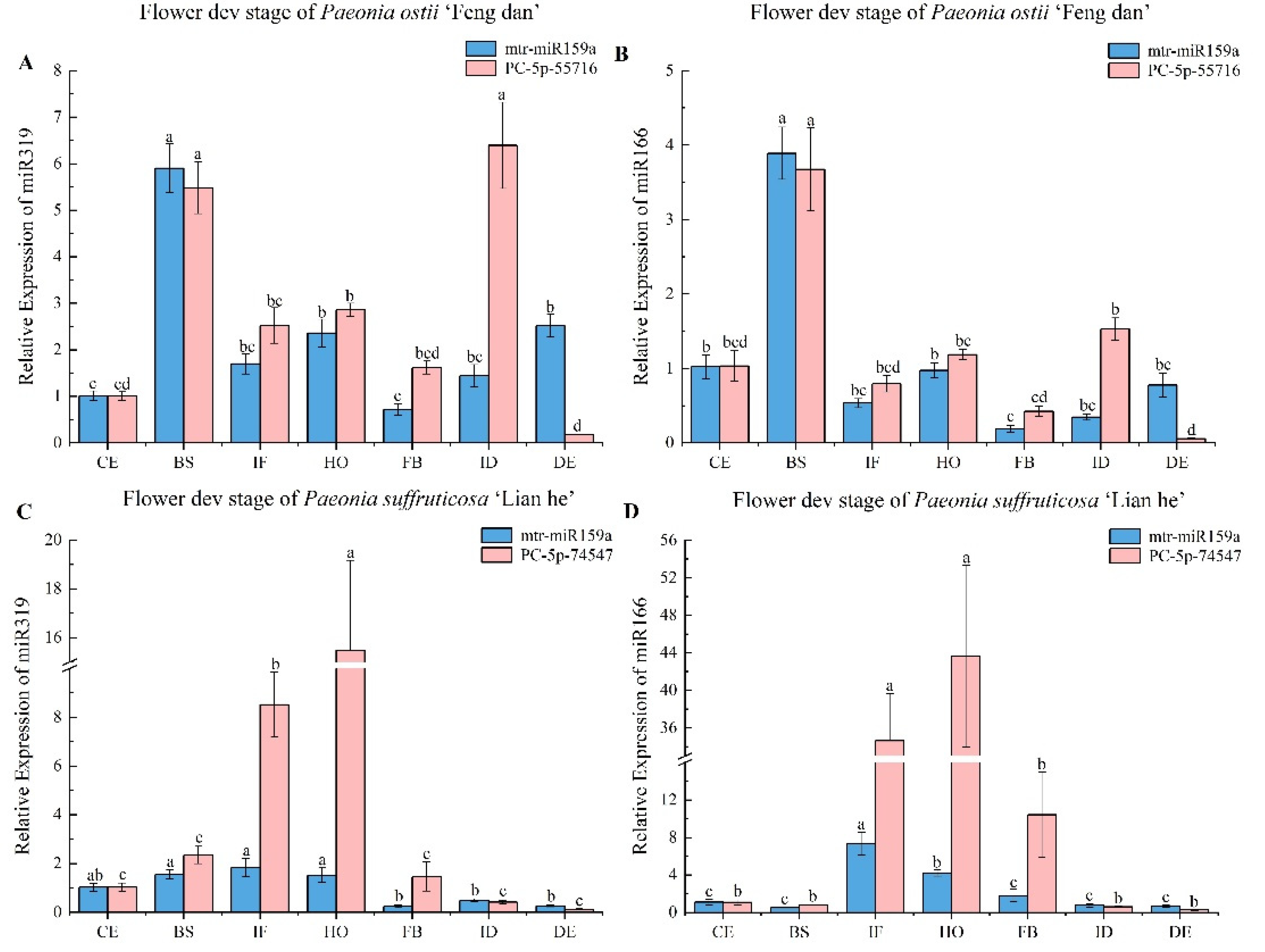
3.3.2. Stability Analysis of Candidate Reference miRNAs at Different Flower Developmental Stages
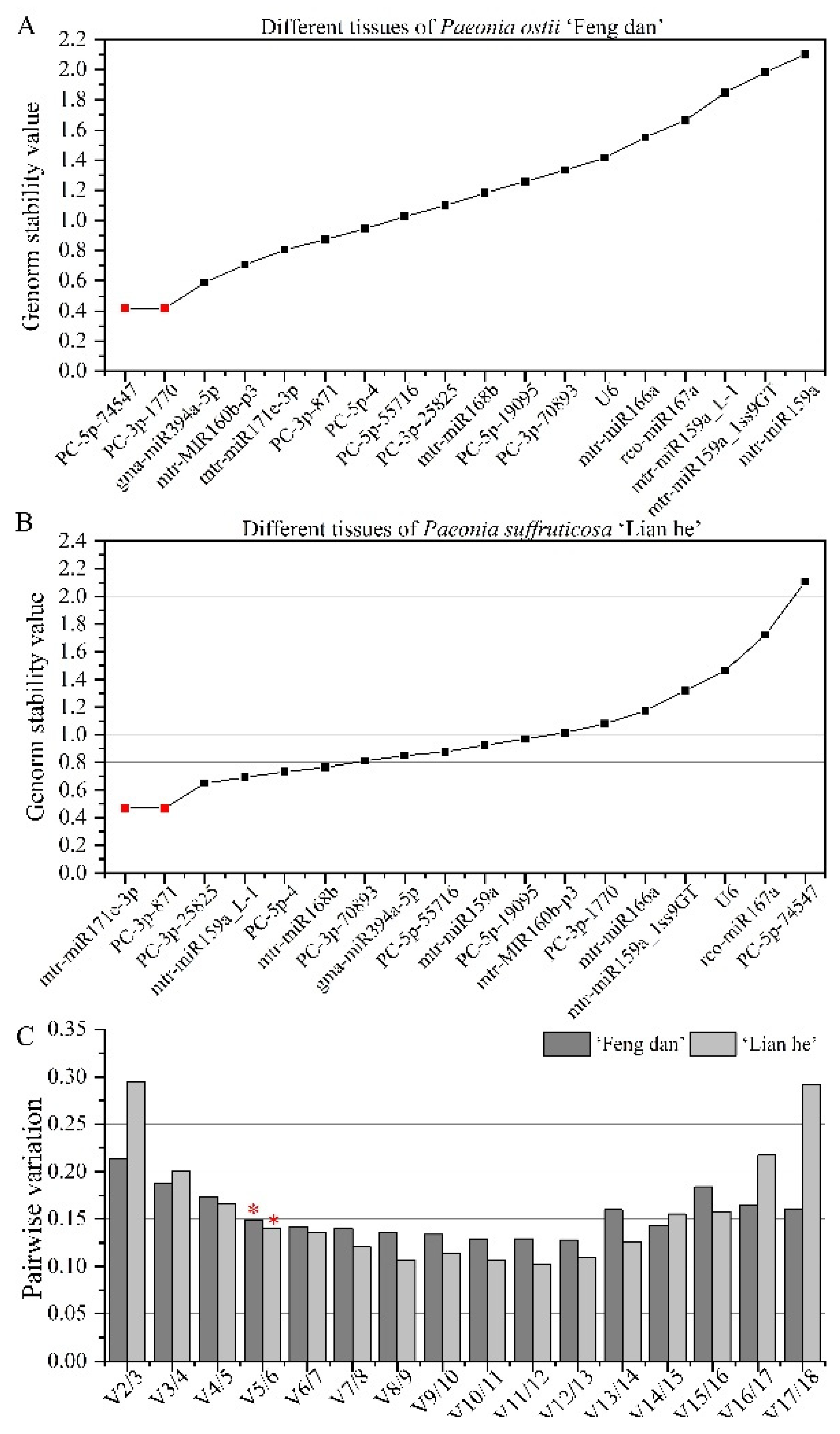
3.3.3. Stability Analysis of Candidate Reference miRNAs at Different Tissues
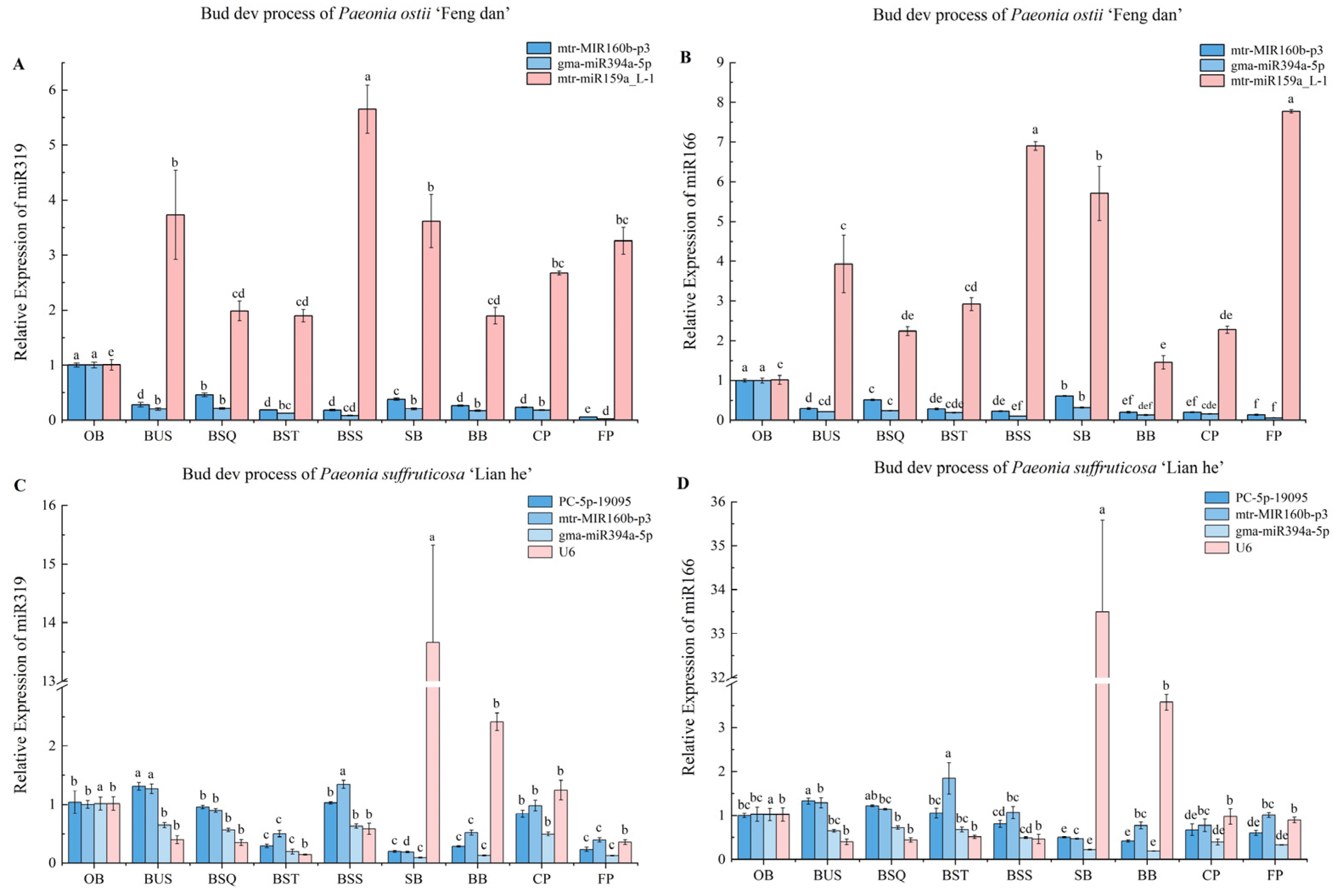
3.4. Validation of Reference miRNAs
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhang, L.; Guo, D.L.; Guo, L.L.; Guo, Q.; Wang, H.F.; Hou, X.G. Construction of a high-density genetic map and QTLs mapping with GBS from the interspecific F1 population of P. ostii ‘Fengdan Bai’ and P. suffruticosa ‘Xin Riyuejin’. Sci. Hortic. 2019, 246, 190–200. [Google Scholar] [CrossRef]
- Liu, Z.Y.; Shi, Y.T.; Xue, Y.Q.; Wang, X.P.; Huang, Z.; Xue, J.Q.; Zhang, X.X. Non-structural carbohydrates coordinate tree peony flowering both as energy substrates and as sugar signaling triggers, with the bracts playing an essential role. Plant Physiol. Biochem. 2021, 159, 80–88. [Google Scholar] [CrossRef] [PubMed]
- Hou, X.G.; Guo, Q.; Wei, W.Q.; Guo, L.L.; Guo, D.L.; Zhang, L. Screening of genes related to early and late flowering in tree peony based on bulked segregant RNA sequencing and verification by quantitative real-time PCR. Molecules 2018, 23, 689. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.J.; Shen, J.J.; Wang, C.; Wang, Z.Y.; Guo, L.L.; Hou, X.G. Characterization of PsmiR319 during flower development in early-and late-flowering tree peonies cultivars. Plant Signal. Behav. 2022, 17, 2120303. [Google Scholar] [CrossRef]
- Zhang, L.; Song, C.W.; Guo, D.L.; Guo, L.L.; Hou, X.G.; Wang, H.F. Identification of differentially expressed miRNAs and their target genes in response to brassinolide treatment on flowering of tree peony (Paeonia ostii). Plant Signal. Behav. 2022, 17, 2056364. [Google Scholar] [CrossRef]
- Wang, J.; Long, Y.; Zhang, J.W.; Xue, M.D.; Huang, G.G.; Huang, K.; Yuan, Q.H.; Pei, X.W. Combined analysis and miRNA expression profiles of the flowering related genes in common wild rice (oryza rufipogon Griff.). Genes Genom. 2018, 40, 835–845. [Google Scholar] [CrossRef]
- Zhao, C.Z.; Li, T.T.; Zhao, Y.H.; Zhang, B.H.; Li, A.Q.; Zhao, S.Z.; Hou, L.; Xia, H.; Fan, S.J.; Qiu, J.J.; et al. Integrated small RNA and mRNA expression profiles reveal miRNAs and their target genes in response to Aspergillus flavus growth in peanut seeds. BMC Plant Biol. 2020, 20, 215. [Google Scholar] [CrossRef]
- Willmann, M.R.; Poethig, R.S. Conservation and evolution of miRNA regulatory programs in plant development. Curr. Opin. Plant Biol. 2007, 10, 503–511. [Google Scholar] [CrossRef]
- Curaba, J.; Spriggs, A.; Taylor, J.; Li, Z.Y.; Helliwell, C. miRNA regulation in the early development of barley seed. BMC Plant Biol. 2012, 12, 1–16. [Google Scholar] [CrossRef]
- Choudhary, A.; Kumar, A.; Kaur, H.; Kaur, N. MiRNA: The taskmaster of plant world. Biologia 2021, 76, 1551–1567. [Google Scholar] [CrossRef]
- Spanudakis, E.; Jackson, S. The role of microRNAs in the control of flowering time. J. Exp. Bot. 2014, 65, 365–380. [Google Scholar] [CrossRef]
- Li, X.Y.; Guo, F.; Ma, S.Y.; Zhu, M.Y.; Pan, W.H.; Bian, H.W. Regulation of flowering time via miR172-mediated APETALA2-like expression in ornamental gloxinia (Sinningia speciosa). J. Zhejiang Univ. -Sci. B 2019, 20, 322–331. [Google Scholar] [CrossRef]
- Lee, Y.S.; Lee, D.Y.; Cho, L.H.; An, G. Rice miR172 induces flowering by suppressing OsIDS1 and SNB, two AP2 genes that negatively regulate expression of Ehd1 and florigens. Rice 2014, 7, 1–13. [Google Scholar] [CrossRef]
- Glazińska, P.; Zienkiewicz, A.; Wojciechowski, W.; Kopcewicz, J. The putative miR172 target gene InAPETALA2-like is involved in the photoperiodic flower induction of Ipomoea nil. J. Plant Physiol. 2009, 166, 1801–1813. [Google Scholar] [CrossRef]
- Zhao, Q.; Sun, C.; Liu, D.D.; Hao, Y.J.; You, C.X. Ectopic expression of the apple Md-miR172e gene alters flowering time and floral organ identity in Arabidopsis. Plant Cell Tissue Organ Cult. 2015, 123, 535–546. [Google Scholar] [CrossRef]
- Tang, M.Y.; Bai, X.; Niu, L.J.; Chai, X.; Chen, M.S.; Xu, Z.F. miR172 regulates both vegetative and reproductive development in the perennial woody plant Jatropha curcas. Plant Cell Physiol. 2018, 59, 2549–2563. [Google Scholar] [CrossRef]
- Yu, Z.X.; Wang, L.J.; Zhao, B.; Shan, C.M.; Zhang, Y.H.; Chen, D.F.; Chen, X.Y. Progressive regulation of sesquiterpene biosynthesis in Arabidopsis and Patchouli (Pogostemon cablin) by the miR156-targeted SPL transcription factors. Mol. Plant 2015, 8, 98–110. [Google Scholar] [CrossRef]
- Chuck, G.; Cigan, A.M.; Saeteurn, K.; Hake, S. The heterochronic maize mutant Corngrass1 results from overexpression of a tandem microRNA. Nat. Genet. 2007, 39, 544–549. [Google Scholar] [CrossRef]
- Zhang, X.H.; Zou, Z.; Zhang, J.H.; Zhang, Y.Y.; Han, Q.Q.; Hu, T.X.; Xu, X.G.; Liu, H.; Ye, Z.B. Over-expression of sly-miR156a in tomato results in multiple vegetative and reproductive trait alterations and partial phenocopy of the sft mutant. FEBS Lett. 2011, 585, 435–439. [Google Scholar] [CrossRef]
- Zhang, T.Q.; Wang, J.W.; Zhou, C.M. The role of miR156 in developmental transitions in Nicotiana tabacum. Sci. China Life Sci. 2015, 58, 253–260. [Google Scholar] [CrossRef]
- Feng, S.J.; Xu, Y.M.; Guo, C.K.; Zheng, J.R.; Zhou, B.Y.; Zhang, Y.T.; Ding, Y.; Zhang, L.; Zhu, Z.J.; Wang, H.S.; et al. Modulation of miR156 to identify traits associated with vegetative phase change in tobacco (Nicotiana tabacum). J. Exp. Bot. 2016, 67, 1493–1504. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.W.; Czech, B.; Weigel, D. miR156-regulated SPL transcription factors define an endogenous flowering pathway in Arabidopsis thaliana. Cell 2009, 138, 738–749. [Google Scholar] [CrossRef] [PubMed]
- Li, X.Y.; Bian, H.W.; Song, D.F.; Ma, S.Y.; Han, N.N.; Wang, J.H.; Zhu, M.Y. Flowering time control in ornamental gloxinia (Sinningia speciosa) by manipulation of miR159 expression. Ann. Bot. 2013, 111, 791–799. [Google Scholar] [CrossRef] [PubMed]
- Zhu, L.; Li, S.S.; Ma, Q.Y.; Wen, J.; Yan, K.Y.; Li, Q.Z. The acer palmatum TCP transcription factor ApTCP2 controls leaf morphogenesis, accelerates senescence, and affects flowering via miR319 in Arabidopsis thaliana. J. Plant Growth Regul. 2022, 41, 244–256. [Google Scholar] [CrossRef]
- Han, J.G.; Zhang, T.R.; Li, J.; Hu, Y.H. Identification of miRNA responsive to early flowering in tree peony (Paeonia ostii) by high-throughput sequencing. J. Hortic. Sci. Biotechnol. 2021, 96, 297–310. [Google Scholar] [CrossRef]
- Xu, M.Y.; Zhang, L.; Li, W.W.; Hu, X.L.; Wang, M.B.; Fan, Y.L.; Zhang, C.Y.; Wang, L. Stress-induced early flowering is mediated by miR169 in Arabidopsis thaliana. J. Exp. Bot. 2014, 65, 89–101. [Google Scholar] [CrossRef]
- Bernardi, Y.; Ponso, M.A.; Belén, F.; Vegetti, A.C.; Dotto, M.C. MicroRNA miR394 regulates flowering time in Arabidopsis thaliana. Plant Cell Rep. 2022, 41, 1375–1388. [Google Scholar] [CrossRef]
- Zhang, J.; Gai, M.Z.; Xue, B.Y.; Jia, N.N.; Wang, C.X.; Wang, J.X.; Sun, H.M. The use of miRNAs as reference genes for miRNA expression normalization during Lilium somatic embryogenesis by real-time reverse transcription PCR analysis. Plant Cell Tissue Organ Cult. 2017, 129, 105–118. [Google Scholar] [CrossRef]
- Kou, S.J.; Wu, X.M.; Liu, Z.; Liu, Y.L.; Xu, Q.; Guo, W.W. Selection and validation of suitable reference genes for miRNA expression normalization by quantitative RT-PCR in citrus somatic embryogenic and adult tissues. Plant Cell Rep. 2012, 31, 2151–2163. [Google Scholar] [CrossRef]
- Tang, F.; Chu, L.W.; Shu, W.B.; He, X.J.; Wang, L.J.; Lu, M.J. Selection and validation of reference genes for quantitative expression analysis of miRNAs and mRNAs in Poplar. Plant Methods 2019, 15, 1–15. [Google Scholar] [CrossRef]
- Borowski, J.M.; Galli, V.; da Silva Messias, R.; Perin, E.C.; Buss, J.H.; dos Anjos e Silva, S.D.; Rombaldi, C.V. Selection of candidate reference genes for real-time PCR studies in lettuce under abiotic stresses. Planta 2014, 239, 1187–1200. [Google Scholar] [CrossRef]
- Luo, M.; Gao, Z.; Li, H.; Li, Q.; Zhang, C.X.; Xu, W.P.; Song, S.R.; Ma, C.; Wang, S.P. Selection of reference genes for miRNA qRT-PCR under abiotic stress in grapevine. Sci. Rep. 2018, 8, 4444. [Google Scholar] [CrossRef]
- Zhang, L.C.; Yang, X.Q.; Yin, Y.Y.; Wang, J.X.; Wang, Y.W. Identification and validation of miRNA reference genes in poplar under pathogen stress. Mol. Biol. Rep. 2021, 48, 3357–3366. [Google Scholar] [CrossRef]
- Peltier, H.J.; Latham, G.J. Normalization of microRNA expression levels in quantitative RT-PCR assays: Identification of suitable reference RNA targets in normal and cancerous human solid tissues. Rna 2008, 14, 844–852. [Google Scholar] [CrossRef]
- Verstraeten, B.; De Smet, L.; Kyndt, T.; De Meyer, T. Selection of miRNA reference genes for plant defence studies in rice (Oryza sativa). Planta 2019, 250, 2101–2110. [Google Scholar] [CrossRef]
- Ferdous, J.; Li, Y.; Reid, N.; Langridge, P.; Shi, B.J.J.; Tricker, P.J. Identification of reference genes for quantitative expression analysis of microRNAs and mRNAs in barley under various stress conditions. PLoS ONE 2015, 10, e0118503. [Google Scholar] [CrossRef]
- Machado, R.D.; Christoff, A.P.; Loss-Morais, G.; Margis-Pinheiro, M.; Margis, R.; Körbes, A.P. Comprehensive selection of reference genes for quantitative gene expression analysis during seed development in Brassica napus. Plant Cell Rep. 2015, 34, 1139–1149. [Google Scholar] [CrossRef]
- Guo, L.L.; Li, Y.Y.; Zhang, C.J.; Wang, Z.Y.; Carlson, J.E.; Yin, W.L.; Zhang, X.X.; Hou, X.G. Integrated analysis of miRNAome, transcriptome, and degradome reveals miRNA-target modules governing floral florescence, development and senescence across early-and late-flowering phenotypes in Paeonia suffruticosa. Front. Plant Sci. 2022, 13, 1082415. [Google Scholar] [CrossRef]
- Lyu, S.H.; Yu, Y.; Xu, S.R.; Cai, W.W.; Chen, G.X.; Chen, J.J.; Pan, D.M.; She, W.Q. Identification of appropriate reference genes for normalizing mirna expression in citrus infected by Xanthomonas citri subsp. citri. Genes 2019, 11, 17. [Google Scholar] [CrossRef]
- Silver, N.; Best, S.; Jiang, J.; Thein, S.L. Selection of housekeeping genes for gene expression studies in human reticulocytes using real-time PCR. BMC Mol. Biol. 2006, 7, 1–9. [Google Scholar] [CrossRef]
- Vandesompele, J.; De Preter, K.; Pattyn, F.; Poppe, B.; Van Roy, N.; De Paepe, A.; Speleman, F. Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of multiple internal control genes. Genome Biol. 2002, 3, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Andersen, C.L.; Jensen, J.L.; Ørntoft, T.F. Normalization of real-time quantitative reverse transcription-PCR data: A model-based variance estimation approach to identify genes suited for normalization, applied to bladder and colon cancer data sets. Cancer Res. 2004, 64, 5245–5250. [Google Scholar] [CrossRef] [PubMed]
- Pfaffl, M.W.; Tichopad, A.; Prgomet, C.; Neuvians, T.P. Determination of stable housekeeping genes, differentially regulated target genes and sample integrity: BestKeeper–Excel-based tool using pair-wise correlations. Biotechnol. Lett. 2004, 26, 509–515. [Google Scholar] [CrossRef] [PubMed]
- Li, N.; Yang, T.X.; Guo, Z.Y.; Wang, Q.S.; Chai, M.; Wu, M.B.; Li, X.Q.; Li, W.Y.; Li, G.X.; Tang, J.H.; et al. Maize microRNA166 inactivation confers plant development and abiotic stress resistance. Int. J. Mol. Sci. 2020, 21, 9506. [Google Scholar] [CrossRef] [PubMed]
- Xue, W.J.; Wang, L.S.; Li, X.Q.; Tang, M.W.; Li, J.Y.; Ding, B.; Kawabata, S.; Li, Y.H.; Zhang, Y. Evaluation of reference genes for quantitative PCR in Eustoma grandiflorum under different experimental conditions. Horticulturae 2022, 8, 164. [Google Scholar] [CrossRef]
- Li, F.; Sun, J.H.; Men, J.L.; Li, H.L.; Wang, G.; Wang, S.J.; Wang, J.B. Selection and validation of reference genes for RT-qPCR analysis in the pericarp of Litchi chinensis. Biol. Plant. 2022, 66, 103–111. [Google Scholar] [CrossRef]
- Li, Y.Y.; Luo, W.R.; Sun, Y.D.; Chang, H.C.; Ma, K.; Zhao, Z.X.; Lu, L. Identification and expression analysis of miR160 and their target genes in cucumber. Biochem. Genet. 2022, 60, 127–152. [Google Scholar] [CrossRef]
- Zhang, X.P.; Shen, J.; Xu, Q.J.; Dong, J.; Song, L.R.; Wang, W.; Shen, F.F. Long noncoding RNA lncRNA354 functions as a competing endogenous RNA of miR160b to regulate ARF genes in response to salt stress in upland cotton. Plant Cell Environ. 2021, 44, 3302–3321. [Google Scholar] [CrossRef]
- Shen, X.X.; He, J.Q.; Ping, Y.K.; Guo, J.X.; Hou, N.; Cao, F.G.; Li, X.W.; Geng, D.L.; Wang, S.C.; Chen, P.X.; et al. The positive feedback regulatory loop of miR160-Auxin Response Factor 17-HYPONASTIC LEAVES 1 mediates drought tolerance in apple trees. Plant Physiol. 2022, 188, 1686–1708. [Google Scholar] [CrossRef]
- Ni, Z.Y.; Hu, Z.; Jiang, Q.Y.; Zhang, H. Overexpression of gma-MIR394a confers tolerance to drought in transgenic Arabidopsis thaliana. Biochem. Biophys. Res. Commun. 2012, 427, 330–335. [Google Scholar] [CrossRef]
- Song, J.B.; Gao, S.; Sun, D.; Li, H.; Shu, X.X.; Yang, Z.M. miR394 and LCR are involved in Arabidopsis salt and drought stress responses in an abscisic acid-dependent manner. BMC Plant Biol. 2013, 13, 1–16. [Google Scholar] [CrossRef]
- Song, J.B.; Gao, S.; Wang, Y.; Li, B.W.; Zhang, Y.L.; Yang, Z.M. miR394 and its target gene LCR are involved in cold stress response in Arabidopsis. Plant Gene 2016, 5, 56–64. [Google Scholar] [CrossRef]
- Millar, A.A.; Lohe, A.; Wong, G. Biology and function of miR159 in plants. Plants 2019, 8, 255. [Google Scholar] [CrossRef]
- Xian, Z.Q.; Huang, W.; Yang, Y.W.; Tang, N.; Zhang, C.; Ren, M.Z.; Li, Z.G. miR168 influences phase transition, leaf epinasty, and fruit development via SlAGO1s in tomato. J. Exp. Bot. 2014, 65, 6655–6666. [Google Scholar] [CrossRef]
- Zanca, A.S.; Vicentini, R.; Ortiz-Morea, F.A.; Del Bem, L.E.; da Silva, M.J.; Vincentz, M.; Nogueira, F.T. Identification and expression analysis of microRNAs and targets in the biofuel crop sugarcane. BMC Plant Biol. 2010, 10, 1–13. [Google Scholar] [CrossRef]
- Luo, X.N.; Luo, S.; Fu, Y.Q.; Kong, C.; Wang, K.; Sun, D.Y.; Li, M.C.; Yan, Z.G.; Shi, Q.Q.; Zhang, Y.L. Genome-wide identification and comparative profiling of micrornas reveal flavonoid biosynthesis in two contrasting flower color cultivars of tree peony. Front. Plant Sci. 2021, 12, 797799. [Google Scholar] [CrossRef]
- Jin, Q.J.; Xue, Z.Y.; Dong, C.L.; Wang, Y.J.; Chu, L.L.; Xu, Y.C. Identification and characterization of microRNAs from tree peony (Paeonia ostii) and their response to copper stress. PLoS ONE 2015, 10, e0117584. [Google Scholar] [CrossRef]
- Yin, D.D.; Li, S.S.; Shu, Q.Y.; Gu, Z.Y.; Wu, Q.; Feng, C.Y.; Xu, W.Z.; Wang, L.S. Identification of microRNAs and long non-coding RNAs involved in fatty acid biosynthesis in tree peony seeds. Gene 2018, 666, 72–82. [Google Scholar] [CrossRef]
- Zhang, Y.X.; Wang, Y.Y.; Gao, X.K.; Liu, C.Y.; Gai, S.P. Identification and characterization of microRNAs in tree peony during chilling induced dormancy release by high-throughput sequencing. Sci. Rep. 2018, 8, 4537. [Google Scholar] [CrossRef]
- Qi, S.; Yang, L.W.; Wen, X.H.; Hong, Y.; Song, X.B.; Zhang, M.M.; Dai, S.L. Reference gene selection for RT-qPCR analysis of flower development in Chrysanthemum morifolium and Chrysanthemum lavandulifolium. Front. Plant Sci. 2016, 7, 287. [Google Scholar] [CrossRef]
- Cortés, A.J.; López-Hernández, F. Harnessing crop wild diversity for climate change adaptation. Genes 2021, 12, 783. [Google Scholar] [CrossRef] [PubMed]
- Cortés, A.J.; López-Hernández, F.; Blair, M.W. Genome–environment associations, an innovative tool for studying heritable evolutionary adaptation in orphan crops and wild relatives. Front. Genet. 2022, 13, 910386. [Google Scholar] [CrossRef] [PubMed]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| miRNA Name | miRNA Mature Sequence | Forward Primer (5′-3′) |
|---|---|---|
| PC-3p-1770 | AGGTACTCCGTTCTCTCTCGTT | GCAGGTAGTCCGTTCTCTCTCG |
| PC-5p-74547 | AAAGTCGGATCGCCAGCAACATC | AGTCGGCTCGCCAGCAACAT |
| gma-miR394a-5p | TTGGCATTCTGTCCACCTCC | CCGCTTGGCATTCTGTCCACCTCC |
| PC-5p-4 | TTAATCAAGGGAATAGGGGCGCA | GCTTAATCAAGGGAATAGGGGCG |
| PC-3p-25825 | CGATTCTTCCACCCGGTCGGA | GCCATTCTTCCACCCGGTCG |
| mtr-miR171e-3p | TGATTGAGCCGCGCCAGTATC | CCGTGATTGAGCCGCGCCAGTAT |
| mtr-MIR160b-p3 | TATGAGGAGCCAAGCATATTG | GGCCGTATGAGGAGCCAAGCATATT |
| PC-3p-871 | TTCCCTGTTCTGGAGATCTAT | GCGGCGTTCCCTGTTCTGGAGATCTAT |
| mtr-miR168b | TCGCTTGGTGCAGGTCGGGAA | TCGCTTGGTGCAGGTCGGGAA |
| PC-3p-70893 | TTCAACCCAACTTCGTCTCTT | GCGGCGTTCAACCCAACTTCGTCTCTT |
| mtr-miR159a | TTTGGATTGAAGGGAGCTCTA | CCGCCGTTTGGATTGAAGGGAGC |
| mtr-miR166a | TCGGACCAGGCTTCATTCCCC | CCGTCGGACCAGGCTTCATTCCC |
| rco-miR167a | TGAAGCTGCCAGCATGATCTA | TCCGAACGCCAGCATGATCTA |
| PC-5p-55716 | CTATAGTCATCATCTGCCACAGGC | CGCCCTATAGTCATCATCTGCCACA |
| mtr-miR159a_L-1 | TTGGATTGAAGGGAGCTCAA | CGCCGTTGGATTGAAGGGAGC |
| mtr-miR159a_1ss9GT | TTTGGATTTAAGGGAGCTCTA | TGGTCGTGTTTGGATTTAAGGGAGC |
| PC-5p-19095 | AAAAGTCGGATCGCCAGCAACATC | CGCAAAAGTCGGATCGCCAGCAACATC |
| cpa-MIR319-p3_1ss20GT | CTGCCATCTCATGCATAAGT | GTCCTGCTGCCATCTCATGCAT |
| mtr-miR166g-5p | GGAATGTTGTCTGGCTCGAGG | CGGTGGGAATGTTGTCTGGCT |
| Ranking | Bud of ‘Feng dan’ | Bud of ‘Lian he’ | Flower of ‘Feng dan’ | Flower of ‘Lian he’ | Tissue of ‘Feng dan’ | Tissue of ‘Lian he’ |
|---|---|---|---|---|---|---|
| 1 | mtr-MIR160b-p3 | PC-5p-19095 | mtr-miR159a | mtr-miR159a | PC-3p-871 | PC-5p-4 |
| 2 | gma-miR394a-5p | gma-miR394a-5p | gma-miR394a-5p | rco-miR167a | PC-5p-4 | mtr-miR168b |
| 3 | PC-3p-70893 | mtr-MIR160b-p3 | PC-5p-19095 | mtr-miR159a_1ss9GT | mtr-miR171e-3p | PC-5p-55716 |
| 4 | mtr-miR171e-3p | mtr-miR166a | mtr-MIR160b-p3 | mtr-miR171e-3p | gma-miR394a-5p | PC-3p-871 |
| 5 | PC-5p-19095 | mtr-miR168b | PC-5p-4 | PC-5p-4 | PC-3p-1770 | mtr-miR159a |
| 6 | PC-3p-871 | mtr-miR171e-3p | mtr-miR168b | PC-5p-19095 | PC-5p-74547 | PC-3p-25825 |
| 7 | mtr-miR166a | PC-3p-70893 | PC-3p-871 | U6 | mtr-MIR160b-p3 | mtr-miR171e-3p |
| 8 | mtr-miR168b | PC-3p-871 | mtr-miR166a | PC-3p-871 | PC-5p-19095 | mtr-miR159a_L-1 |
| 9 | PC-5p-74547 | mtr-miR159a_1ss9GT | PC-3p-25825 | PC-3p-70893 | PC-5p-55716 | PC-5p-19095 |
| 10 | PC-3p-1770 | rco-miR167a | PC-5p-74547 | PC-3p-1770 | U6 | gma-miR394a-5p |
| 11 | PC-3p-25825 | mtr-miR159a | mtr-miR159a_L-1 | mtr-MIR160b-p3 | mtr-miR168b | PC-3p-70893 |
| 12 | U6 | mtr-miR159a_L-1 | mtr-miR159a_1ss9GT | mtr-miR168b | mtr-miR166a | mtr-MIR160b-p3 |
| 13 | PC-5p-4 | PC-5p-74547 | U6 | gma-miR394a-5p | PC-3p-25825 | mtr-miR166a |
| 14 | PC-5p-55716 | PC-5p-4 | PC-3p-1770 | mtr-miR159a_L-1 | PC-3p-70893 | PC-3p-1770 |
| 15 | mtr-miR159a_1ss9GT | PC-5p-55716 | mtr-miR171e-3p | PC-5p-55716 | rco-miR167a | mtr-miR159a_1ss9GT |
| 16 | mtr-miR159a | PC-3p-25825 | rco-miR167a | PC-3p-25825 | mtr-miR159a_L-1 | U6 |
| 17 | rco-miR167a | PC-3p-1770 | PC-3p-70893 | mtr-miR166a | mtr-miR159a_1ss9GT | rco-miR167a |
| 18 | mtr-miR159a_L-1 | U6 | PC-5p-55716 | PC-5p-74547 | mtr-miR159a | PC-5p-74547 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, C.; Song, C.; Chen, L.; Ma, H.; Zhang, Y.; Guo, D.; Guo, L.; Hou, X. Selection and Validation of miRNA Reference Genes by Quantitative Real-Time PCR Analysis in Paeonia suffruticosa. Horticulturae 2023, 9, 148. https://doi.org/10.3390/horticulturae9020148
Zhang C, Song C, Chen L, Ma H, Zhang Y, Guo D, Guo L, Hou X. Selection and Validation of miRNA Reference Genes by Quantitative Real-Time PCR Analysis in Paeonia suffruticosa. Horticulturae. 2023; 9(2):148. https://doi.org/10.3390/horticulturae9020148
Chicago/Turabian StyleZhang, Chenjie, Chengwei Song, Linfeng Chen, Huili Ma, Yabing Zhang, Dalong Guo, Lili Guo, and Xiaogai Hou. 2023. "Selection and Validation of miRNA Reference Genes by Quantitative Real-Time PCR Analysis in Paeonia suffruticosa" Horticulturae 9, no. 2: 148. https://doi.org/10.3390/horticulturae9020148
APA StyleZhang, C., Song, C., Chen, L., Ma, H., Zhang, Y., Guo, D., Guo, L., & Hou, X. (2023). Selection and Validation of miRNA Reference Genes by Quantitative Real-Time PCR Analysis in Paeonia suffruticosa. Horticulturae, 9(2), 148. https://doi.org/10.3390/horticulturae9020148