
Temperature and GA3 as Modulating Factors in the Biosynthesis of Alkaloids during Imbibition and Early Development of Annona x atemoya Mabb. cv. ‘Gefner’ Seedlings

 ,
,  , , , and
, , , and
Abstract

1. Introduction
2. Materials and Methods
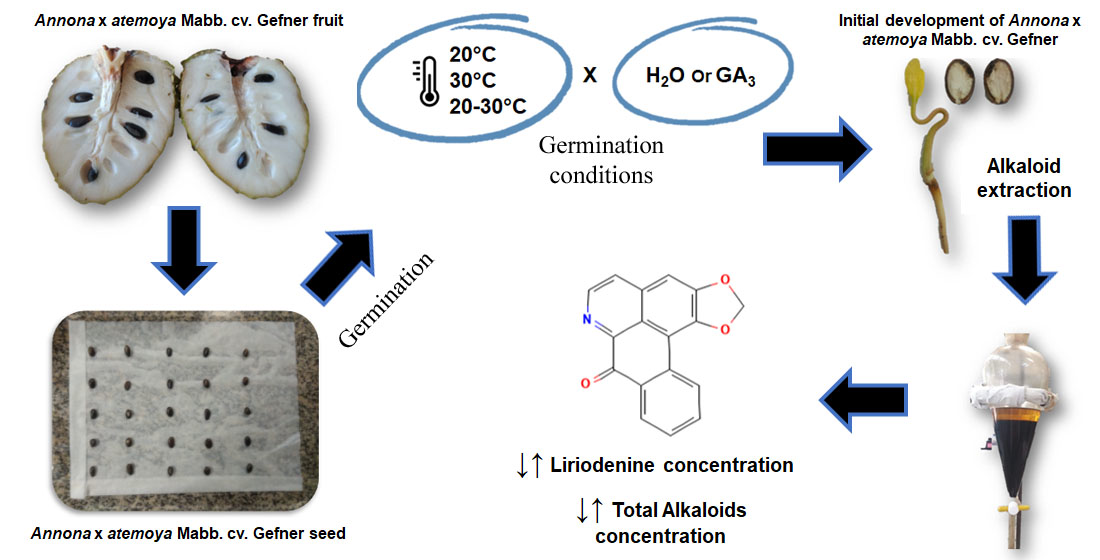
2.1. Plant Material
2.2. Experimental Design and Experiment Implementation
2.3. Germination Test
2.4. Extraction and Quantification of Total Alkaloids and Liriodenine
2.5. Statistical Analysis
3. Results and Discussion
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- De-la-Cruz-Chacón, I.; López-Fernández, N.Y.; Riley-Saldaña, C.A.; Castro-Moreno, M.; González-Esquinca, A.R. Antifungal Activity in Vitro of Sapranthus microcarpus (Annonaceae) against Phytopathogens. Acta Bot. Mex. 2019, 126, 1–9. [Google Scholar] [CrossRef]
- dos Santos, R.C.; Chagas, E.A.; de Melo Filho, A.A.; Takahashi, J.A.; Montero, I.F.; Chagas, P.C.; Ribeiro, P.R.E.; de Melo, C.G.R. Bioactive Extracts from Annona hypoglauca. Chem. Eng. Trans. 2018, 64, 289–294. [Google Scholar] [CrossRef]
- González-Esquinca, A.R.; De-La-Cruz-Chacón, I.; Castro-Moreno, M.; Orozco-Castillo, J.A.; Riley-Saldaña, C.A. Alkaloids and Acetogenins in Annonaceae Development: Biological Considerations. Rev. Bras. De Frutic. 2014, 36, 1–16. [Google Scholar] [CrossRef]
- Levorato Vinche, A.D.; de-La-Cruz-Chacón, I.; González-Esquinca, A.R.; de Fátima da Silva, J.; Ferreira, G.; dos Santos, D.C.; Garces, H.G.; de Oliveira, D.V.M.; Marçon, C.; de Souza Cavalcante, R.; et al. Antifungal Activity of Liriodenine on Agents of Systemic Mycoses, with Emphasis on the Genus Paracoccidioides. J. Venom. Anim. Toxins Incl. Trop. Dis. 2020, 26, e20200023. [Google Scholar] [CrossRef] [PubMed]
- Rinaldi, M.V.N.; Díaz, I.E.C.; Suffredini, I.B.; Moreno, P.R.H. Alkaloids and Biological Activity of Beribá (Annona hypoglauca). Rev. Bras. De Farmacogn. 2017, 27, 77–83. [Google Scholar] [CrossRef]
- Tamfu, A.N.; Ceylan, O.; Fru, G.C.; Ozturk, M.; Duru, M.E.; Shaheen, F. Antibiofilm, antiquorum sensing and antioxidant activity of secondary metabolites from seeds of Annona senegalensis, Persoon. Microb. Pathog. 2020, 144, 104191. [Google Scholar] [CrossRef]
- Lúcio, A.S.S.C.; Almeida, J.R.G.D.S.; da-Cunha, E.V.L.; Tavares, J.F.; Barbosa Filho, J.M. Alkaloids of the Annonaceae: Occurrence and a Compilation of Their Biological Activities. Alkaloids Chem. Biol. 2015, 74, 233–409. [Google Scholar]
- Honório, A.B.M.; De-La-cruz-chacón, I.; Martínez-Vázquez, M.; da Silva, M.R.; Campos, F.G.; Martin, B.C.; da Silva, G.C.; Fernandes Boaro, C.S.; Ferreira, G. Impact of Drought and Flooding on Alkaloid Production in Annona crassiflora Mart. Horticulturae 2021, 7, 414. [Google Scholar] [CrossRef]
- Ovile Mimi, C.; De-la-Cruz-Chacón, I.; Caixeta Sousa, M.; Aparecida Ribeiro Vieira, M.; Ortiz Mayo Marques, M.; Ferreira, G.; Silvia Fernandes Boaro, C. Chemophenetics as a Tool for Distinguishing Morphotypes of Annona emarginata (Schltdl.) H. Rainer. Chem. Biodivers. 2021, 18, 1–14. [Google Scholar] [CrossRef]
- Corrêa, P.L.C.; De-la-Cruz-Chacón, I.; Sousa, M.C.; Vieira, M.A.R.; Campos, F.G.; Marques, M.O.M.; Boaro, C.S.F.; Ferreira, G. Effect of Nitrogen Sources on Photosynthesis and Biosynthesis of Alkaloids and Leaf Volatile Compounds in Annona sylvatica A. St.-Hil. J. Soil Sci. Plant Nutr. 2022, 22, 956–970. [Google Scholar] [CrossRef]
- Mannino, G.; Gentile, C.; Porcu, A.; Agliassa, C.; Caradonna, F.; Bertea, C.M. Chemical Profile and Biological Activity of Cherimoya (Annona cherimola Mill.) and Atemoya (Annona atemoya) Leaves. Molecules 2020, 25, 2612. [Google Scholar] [CrossRef] [PubMed]
- De-La-Cruz Chacón, I.; Riley-Saldaña, C.A.; González-Esquinca, A.R. Secondary Metabolites during Early Development in Plants. Phytochem. Rev. 2013, 12, 47–64. [Google Scholar] [CrossRef]
- de la Cruz-Chacón, I.; González-Esquinca, A.R.; Fefer, P.G.; Garcia, L.F.J. Liriodenine, Early Antimicrobial Defence in Annona diversifolia. Z. Für Nat. Sect. C J. Biosci. 2011, 66, 377–384. [Google Scholar] [CrossRef]
- Rabêlo, S.V.; Costa, E.V.; Barison, A.; Dutra, L.M.; Nunes, X.P.; Tomaz, J.C.; Oliveira, G.G.; Lopes, N.P.; Santos, M.D.F.C.; Almeida, J.R.G.d.S. Alkaloids Isolated from the Leaves of Atemoya (Annona cherimola × Annona squamosa). Braz. J. Pharmacogn. 2015, 25, 419–421. [Google Scholar] [CrossRef]
- De La Cruz Chacón, I.; González-Esquinca, A.R. Liriodenine Alkaloid in Annona diversifolia during Early Development. Nat. Prod. Res. 2012, 26, 42–49. [Google Scholar] [CrossRef]
- Sousa, M.C.; Bronzatto, A.C.; González-Esquinca, A.R.; Campos, F.G.; Dalanhol, S.J.; Boaro, C.S.F.; Martins, A.L.; da Silva Almeida, J.R.G.; Costa, E.V.; De-la-Cruz-Chacón, I.; et al. The Production of Alkaloids in Annona cacans Seedlings Is Affected by the Application of GA4+7 + 6-Benzyladenine. Biochem. Syst. Ecol. 2019, 84, 47–51. [Google Scholar] [CrossRef]
- Castro-Moreno, M.; Tinoco-Ojangurén, C.L.; Cruz-Ortega, M.D.R.; González-Esquinca, A.R. Influence of Seasonal Variation on the Phenology and Liriodenine Content of Annona lutescens (Annonaceae). J. Plant Res. 2013, 126, 529–537. [Google Scholar] [CrossRef]
- Riley-Saldaña, C.A.; Cruz-Ortega, M.D.R.; Martínez Vázquez, M.; De-La-Cruz-Chacón, I.; Castro-Moreno, M.; González-Esquinca, A.R. Acetogenins and Alkaloids during the Initial Development of Annona muricata L. (Annonaceae). Z. Für Nat. Sect. C J. Biosci. 2017, 72, 497–506. [Google Scholar] [CrossRef]
- Riley-saldaña, C.A.; Cruz-ortega, M.R. Do Colletotrichum gloeosporioides and Rhizopus stolonifer Induce Alkaloidal and Antifungal Responses in Annona muricata Seedlings? Z. Für Nat. Sect. C J. Biosci. 2022, 77, 1–7. [Google Scholar] [CrossRef]
- Christie, P.J.; Alfenito, M.R.; Walbot, V. Impact of Low-Temperature Stress on General Phenylpropanoid and Anthocyanin Pathways: Enhancement of Transcript Abundance and Anthocyanin Pigmentation in Maize Seedlings. Planta 1994, 194, 541–549. [Google Scholar] [CrossRef]
- Elgorashi, E.E.; Drewes, S.E.; Van Staden, J. Organ-to-Organ and Seasonal Variation in Alkaloids from Crinum macowanii. Fitoterapia 2002, 73, 490–495. [Google Scholar] [CrossRef]
- Gobbo-Neto, L.; Lopes, N.P. Plantas Medicinais: Fatores de Influência No Conteúdo de Metabólitos Secundários. Quim. Nova 2007, 30, 374–381. [Google Scholar] [CrossRef]
- Roca-Pérez, L.; Boluda, R.; Gavidia, I.; Pérez-Bermúdez, P. Seasonal Cardenolide Production and Dop5βr Gene Expression in Natural Populations of Digitalis obscura. Phytochemistry 2004, 65, 1869–1878. [Google Scholar] [CrossRef] [PubMed]
- Wallaart, T.E.; Pras, N.; Beekman, A.C.; Quax, W.J. Seasonal Variation of Artemisinin and Its Biosynthetic Precursors in Plants of Artemisia annua of Different Geographical Origin: Proof for the Existence of Chemotypes. Planta Med. 2000, 66, 57–62. [Google Scholar] [CrossRef]
- Zucareli, V.; Ferreira, G.; Silvério, E.R.V.; Amaro, A.C.E. Luz e Temperatura Na Germinação de Sementes de Annona squamosa L. Rev. Bras. De Biociências 2007, 5, 840–842. [Google Scholar]
- Ferreira, G.; De-La-cruz-chacón, I.; González-Esquinca, A.R. Changes in Hormonal Balance as Key to Reserve Degradation after Dormancy Overcoming in Annona macroprophyllata and Annona purpurea Seeds. Rev. Bras. Frutic. 2020, 42, 1–11. [Google Scholar] [CrossRef]
- de Oliveira, M.C.; Ferreira, G.; Guimarães, V.F.; Dias, G.B. Germinação de Sementes de Atemoia (Annona cherimola Mill. × A. squamosa L.) CV “Gefner” Submetidas a Tratamentos Com Ácido Giberélico (GA3) e Ethephon. Rev. Bras. Frutic. 2010, 32, 27. [Google Scholar] [CrossRef]
- Ferreira, G.; De-La-Cruz-Chacón, I.; González-Esquinca, A.R. Superação Da Dormência de Sementes de Annona macroprophyllata e Annona purpurea Com o Uso de Reguladores Vegetais. Rev. Bras. Frutic. 2016, 38, 1–10. [Google Scholar] [CrossRef]
- Rego, C.H.Q.; Cardoso, F.B.; Cotrim, M.F.; Cândido, A.C.D.S.; Alves, C.Z. Ácido Giberélico Auxilia Na Superação Da Dormência Fisiológica E Expressão De Vigor Das Sementes De Graviola. J. Neotrop. Agric. 2018, 5, 83–86. [Google Scholar] [CrossRef]
- Costa, P.N.; Bueno, S.S.C.; Ferreira, G. Germination Phases of Annona emarginata (SCHLTDL.) H. Rainer Seeds under Different Temperatures. Rev. Bras. Frutic. 2011, 33, 253–260. [Google Scholar] [CrossRef]
- Oliveira, I.V.d.M.; Andrade, R.A.D.; Martins, A.B.G. Influência Da Temperatura Na Germinação de Sementes de Annona montana. Rev. Bras. Frutic. 2005, 27, 344–345. [Google Scholar] [CrossRef]
- Gomes, R.L.; Passos, J.R.d.S.; Gimenez, J.I.; Sousa, M.C.; De-Pieri-Oliveira, M.d.F.; Mimi, C.O.; Ferreira, G. Optimum Sample Size in the Germination of Atemoya Seeds (Annona × atemoya Mabb.). J. Agric. Sci. 2019, 11, 239. [Google Scholar] [CrossRef]
- Costa, J.R. Técnicas Experimentais Aplicadas Às Ciências Agrárias. Embrapa Agrobiol. 2003, 54, 1–54. [Google Scholar]
- Stenzel, N.M.C.; Murata, I.M.; Neves, C.S.V.J. Superação Da Dormência Em Sementes de Atemóia e Fruta-Do-Conde. Rev. Bras. Frutic. 2003, 25, 305–308. [Google Scholar] [CrossRef]
- Braga, J.F.; Ferreira, G.; Pinho, S.Z.d.; Braga, L.F.; Sousa, M.P. Germination of Atemoya (Annona cherimola Mill. × A. squamosa L.) Cv. Gefner Seeds Subjected to Treatments with Plant Growth Regulators. Int. J. Sci. Nat. 2010, 1, 120–126. [Google Scholar]

{kind=link}
| Total Alkaloid Concentration | Liriodenine Concentration | ||||
|---|---|---|---|---|---|
| Temperature | H2O | GA3 | H2O | GA3 | |
| Seed coat | 20 °C | 0.779 Aa * | 1.771 Aa | ND | ND |
| 30 °C | 1.553 Aa | 1.346 Aa | ND | ND | |
| 20–30 °C | 1.481 Aa | 1.833 Aa | ND | ND | |
| Endosperm | 20 °C | 13.857 BCa | 19.272 Aa | 0.000 Ab | 0.661 Aa |
| 30 °C | 22.777 Aba | 14.205 Ab | 0.199 Aa | 0.239 ABa | |
| 20–30 °C | 8.665 Ca | 11.248 Aa | 0.000 Aa | 0.000 Ba | |
| Seed Imbibition H2O | Seed Imbibition GA3 | |||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Alkaloids | Seed Coat | Endosperm | Hypocotyl | Cotyledon | Root | Sead Coat | Endosperm | Hypocotyl | Cotyledon | Root | ||||||||||||||||||||
| 20 | 30 | 20–30 | 20 | 30 | 20–30 | 20 | 30 | 20–30 | 20 | 30 | 20–30 | 20 | 30 | 20–30 | 20 | 30 | 20–30 | 20 | 30 | 20–30 | 20 | 30 | 20–30 | 20 | 30 | 20–30 | 20 | 30 | 20–30 | |
| Asimilobine | x | x | x | x | x | x | x | x | x | x | ||||||||||||||||||||
| Discretine | ||||||||||||||||||||||||||||||
| Lanuginosine | x | x | x | x | x | x | x | x | x | x | x | x | x | x | x | x | x | x | x | x | x | x | x | x | x | x | x | x | x | |
| Laurotetanine | x | x | x | x | x | x | x | x | x | x | x | x | x | x | x | x | x | x | x | x | x | x | ||||||||
| Liriodenine | x | x | x | x | x | x | x | x | x | x | x | x | x | x | x | x | x | x | x | x | x | x | x | x | x | x | x | x | x | x |
| N-metil-Lauroteanine | x | x | x | x | x | x | x | x | x | x | x | x | x | x | x | x | x | x | ||||||||||||
| Norglaucine | x | |||||||||||||||||||||||||||||
| Oxoglaucine | x | x | x | x | x | x | ||||||||||||||||||||||||
| Reticuline | x | |||||||||||||||||||||||||||||
| Xylopine | x | x | x | x | x | x | x | x | x | x | x | x | x | x | x | x | x | x | x | x | x | x | x | |||||||
| Xylopinine | x | x | x | x | x | x | x | x | x | x | x | x | x | x | x | x | x | x | x | x | x | x | x | |||||||
| Alkaloids | Seed Dry | Seed Imbibition H2O | Seed Imbibition GA3 | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Endosperm | Seed Coat | Endosperm | Seed Coat | Endosperm | Seed Coat | |||||||||
| 20 | 30 | 20–30 | 20 | 30 | 20–30 | 20 | 30 | 20–30 | 20 | 30 | 20–30 | |||
| Asimilobine | x | x | x | x | x | x | x | x | x | x | ||||
| Discretine | ||||||||||||||
| Lanuginosine | x | x | ||||||||||||
| Laurotetanine | x | x | x | x | x | |||||||||
| Liriodenine | x | x | x | x | x | x | x | x | x | x | x | x | ||
| N-metil-Lauroteanine | ||||||||||||||
| Norglaucine | ||||||||||||||
| Oxoglaucine | x | x | x | |||||||||||
| Reticuline | ||||||||||||||
| Xylopine | x | |||||||||||||
| Xylopinine | x | x | x | x | x | x | ||||||||
| Temperature | H2O | GA3 | |
|---|---|---|---|
| % G | 20 °C | 15 Cb * | 71 Aa |
| 30 °C | 42 Bb | 71 Aa | |
| 20–30 °C | 55 ABa | 62 Aa | |
| % D | 20 °C | 59 Aa | 10 Ab |
| 30 °C | 35 Ba | 0 Ab | |
| 20–30 °C | 25 Ba | 0 Ab | |
| % M | 20 °C | 26 Aa | 19 Aa |
| 30 °C | 23 Aa | 29 Aa | |
| 20–30 °C | 20 Aa | 38 Aa | |
| GSI | 20 °C | 0.209 Cb | 0.805 Ca |
| 30 °C | 1.063 Bb | 2.155 ABa | |
| 20–30 °C | 1.618 ABa | 1.676 Ba | |
| MGT | 20 °C | 18.75 Ab | 25.685 Aa |
| 30 °C | 13.219 Ba | 9.268 Bb | |
| 20–30 °C | 10.504 Ba | 10.202 Ba |
| Temperature | H2O | GA3 | |
|---|---|---|---|
| Seed coat | 20 °C | 27.291 Aa * | 0.818 Ab |
| 30 °C | 5.169 BCa | 0.375 Aa | |
| 20–30 °C | 4.035 Ca | 2.17 Aa | |
| Endosperm | 20 °C | 168.218 Aa | 13.609 Bb |
| 30 °C | 46.133 BCa | 28.24 Ba | |
| 20–30 °C | 34.151 Cb | 83.5 Aa | |
| Root | 20 °C | 147.039 Ca | 44.782 Bb |
| 30 °C | 342.344 Aa | 119.647 Ab | |
| 20–30 °C | 173.474 BCa | 88.002 ABb | |
| Hypocotyl | 20 °C | 20.947 Cb | 33.22 Aa |
| 30 °C | 41.469 Ba | 21.755 Ab | |
| 20–30 °C | 51.603 ABa | 27.48 Ab | |
| Cotyledon | 20 °C | 122.053 Aa | 8.582 Bb |
| 30 °C | 84.985 Aa | 56.581 Bb | |
| 20–30 °C | 129.832 Aa | 41.967 Bb |
| Temperature | H2O | GA3 | |
|---|---|---|---|
| Seed coat | 20 °C | 0.000 Ba * | 0.144 Ca |
| 30 °C | 1.286 Aa | 0.956 ABa | |
| 20–30 °C | 0.359 Ba | 0.819 Ba | |
| Endosperm | 20 °C | 6.921 Aa | 2.112 Ba |
| 30 °C | 18.656 Aa | 13.116 Ba | |
| 20–30 °C | 7.011 Ab | 38.163 Aa | |
| Root | 20 °C | 27.453 Ba | 13.210 Aa |
| 30 °C | 127.831 Aa | 26.912 Ab | |
| 20–30 °C | 46.304 Ba | 22.658 Aa | |
| Hypocotyl | 20 °C | 20.328 Aa | 3.755 Bb |
| 30 °C | 6.868 Aa | 3.133 Ba | |
| 20–30 °C | 7.429 Aa | 2.591 Ba | |
| Cotyledon | 20 °C | 0.000 Bb | 25.084 Aa |
| 30 °C | 16.483 Ca | 21.890 ABa | |
| 20–30 °C | 30.932 Aa | 13.348 CBb |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
da Silva, G.C.; de-la-Cruz-Chacón, I.; Honório, A.B.M.; Martin, B.C.; Sousa, M.C.; Campos, F.G.; Boaro, C.S.F.; Ferreira, G. Temperature and GA3 as Modulating Factors in the Biosynthesis of Alkaloids during Imbibition and Early Development of Annona x atemoya Mabb. cv. ‘Gefner’ Seedlings. Horticulturae 2022, 8, 766. https://doi.org/10.3390/horticulturae8090766
da Silva GC, de-la-Cruz-Chacón I, Honório ABM, Martin BC, Sousa MC, Campos FG, Boaro CSF, Ferreira G. Temperature and GA3 as Modulating Factors in the Biosynthesis of Alkaloids during Imbibition and Early Development of Annona x atemoya Mabb. cv. ‘Gefner’ Seedlings. Horticulturae. 2022; 8(9):766. https://doi.org/10.3390/horticulturae8090766
Chicago/Turabian Styleda Silva, Gustavo Cabral, Ivan de-la-Cruz-Chacón, Ana Beatriz Marques Honório, Bruna Cavinatti Martin, Marília Caixeta Sousa, Felipe Girotto Campos, Carmen Sílvia Fernandes Boaro, and Gisela Ferreira. 2022. "Temperature and GA3 as Modulating Factors in the Biosynthesis of Alkaloids during Imbibition and Early Development of Annona x atemoya Mabb. cv. ‘Gefner’ Seedlings" Horticulturae 8, no. 9: 766. https://doi.org/10.3390/horticulturae8090766
APA Styleda Silva, G. C., de-la-Cruz-Chacón, I., Honório, A. B. M., Martin, B. C., Sousa, M. C., Campos, F. G., Boaro, C. S. F., & Ferreira, G. (2022). Temperature and GA3 as Modulating Factors in the Biosynthesis of Alkaloids during Imbibition and Early Development of Annona x atemoya Mabb. cv. ‘Gefner’ Seedlings. Horticulturae, 8(9), 766. https://doi.org/10.3390/horticulturae8090766