


Genome-Wide Identification and Expression Analysis of the PME and PMEI Gene Families in Diospyros kaki: A Bioinformatics Study


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Identification of PME and PMEI Genes
2.2. Phylogenetic Analysis and Interaction Network Construction
2.3. Motif Prediction and Gene Structure Analysis of PME and PMEI Genes
2.4. Chromosomal Distribution, Gene Duplication, and Kaks Calculation Analysis
2.5. Promoter Cis-Regulatory Elements Analysis
2.6. In Silico Expression Analysis of PME and PMEI Genes
3. Results
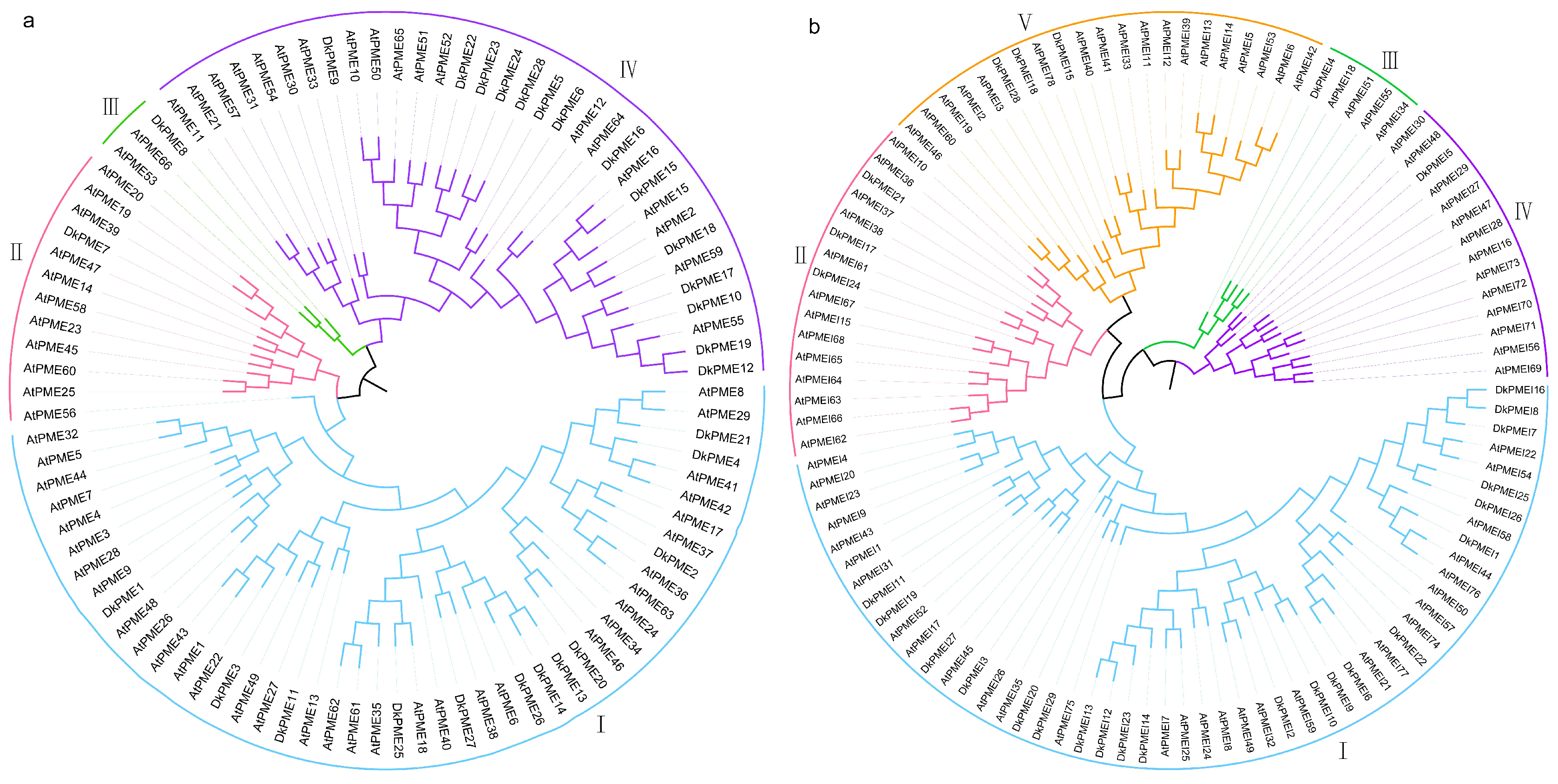
3.1. Identification and Phylogenetic Analysis of PME and PMEI Genes in D. kaki
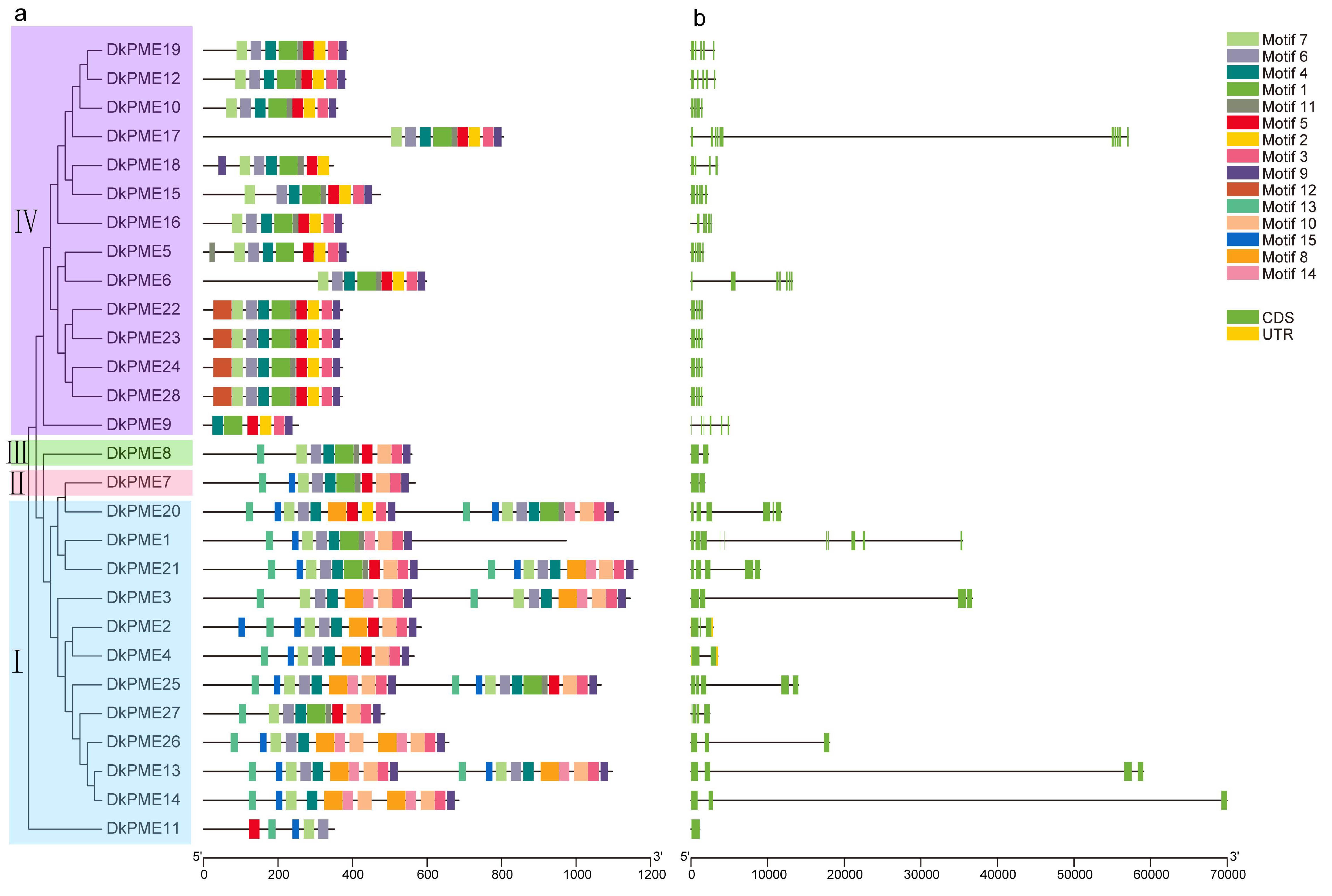
3.2. Conserved Motifs and Structural Analysis
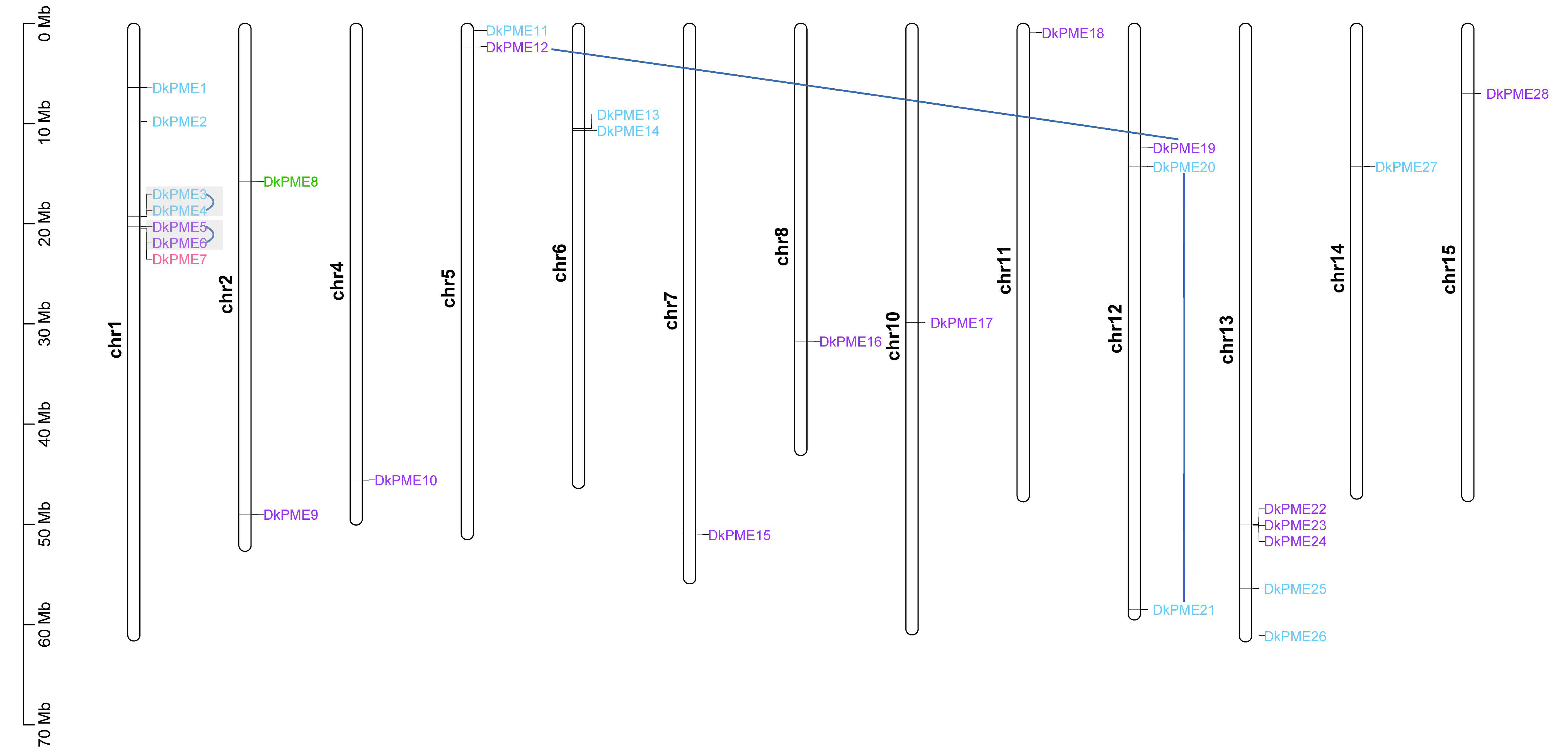
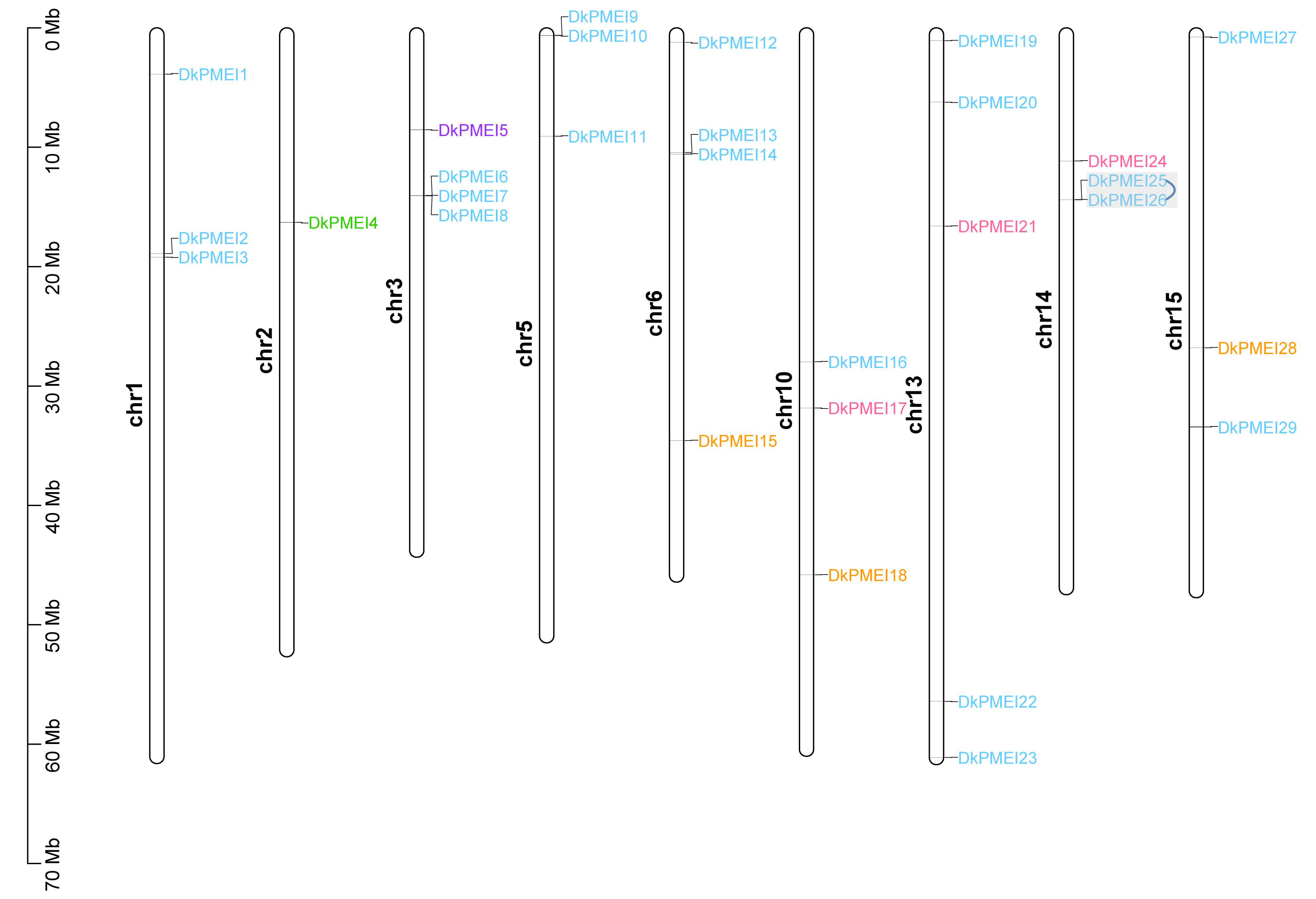
3.3. Chromosomal Locations, Gene Duplication, and Ka/Ks of PME and PMEI Genes
3.4. Synteny Analysis of PME and PMEI Genes
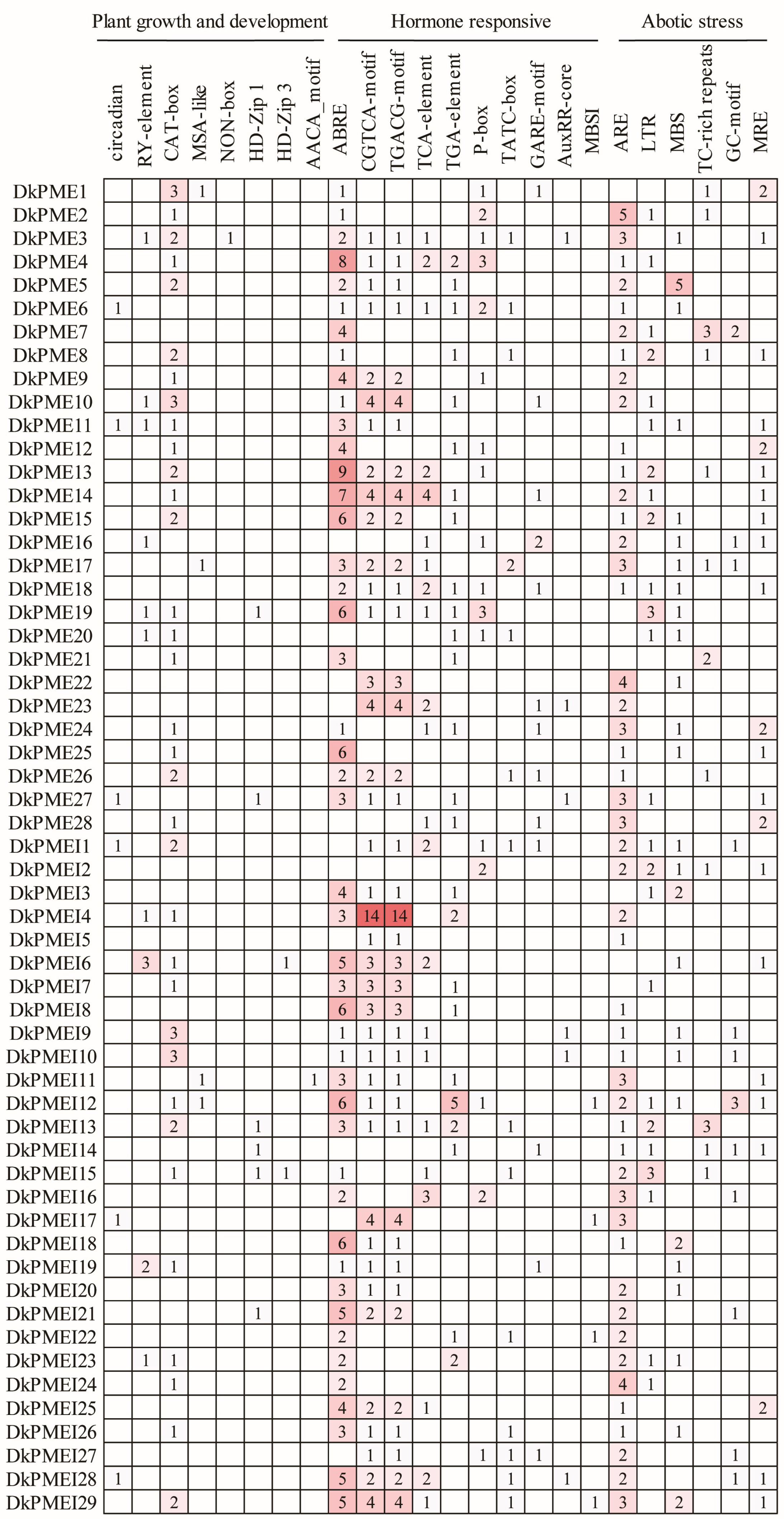
3.5. Cis-Element Analysis of PME and PMEI Genes
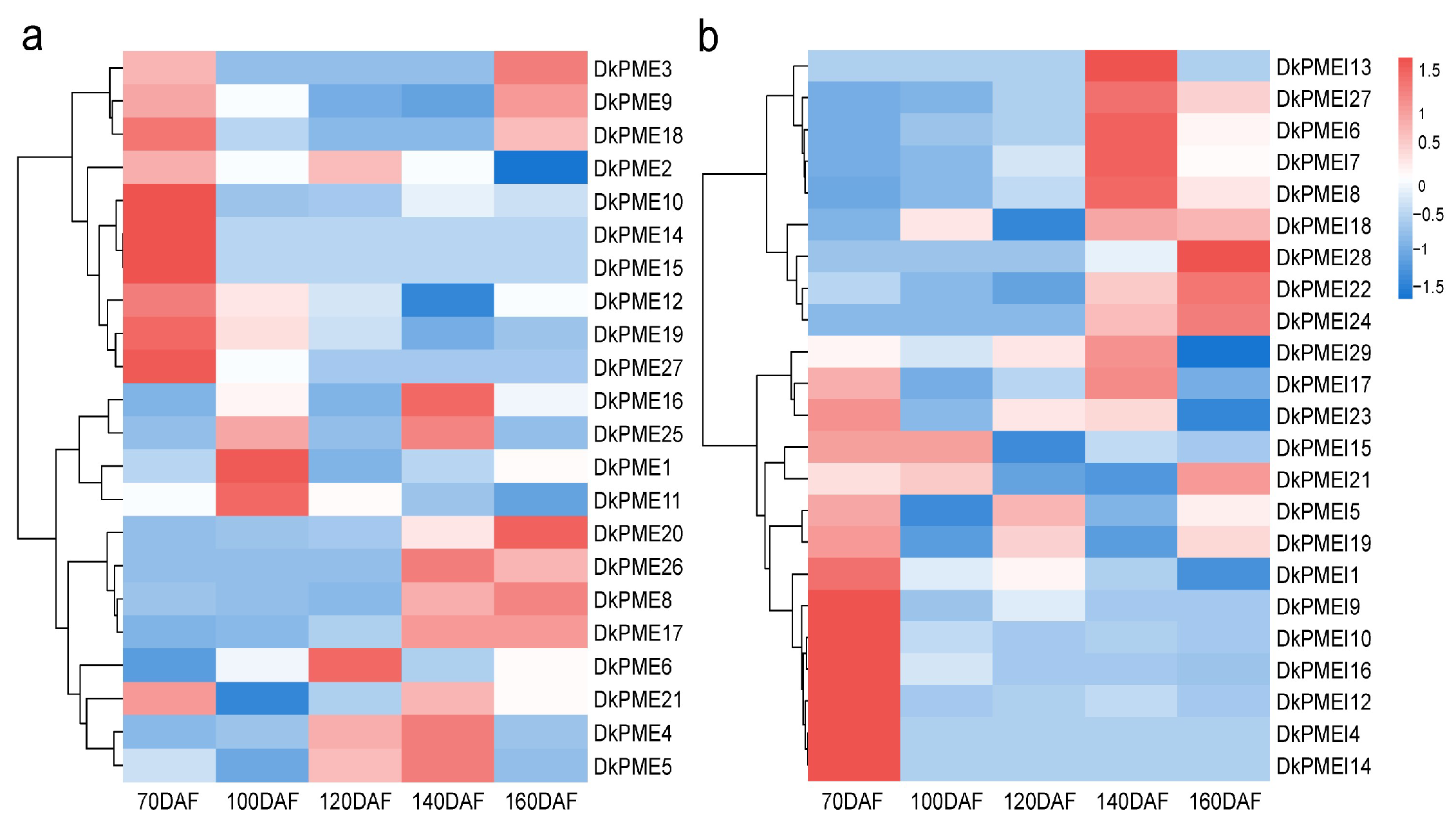
3.6. Expression Patterns of DkPME and DkPMEI Genes during Fruit Development
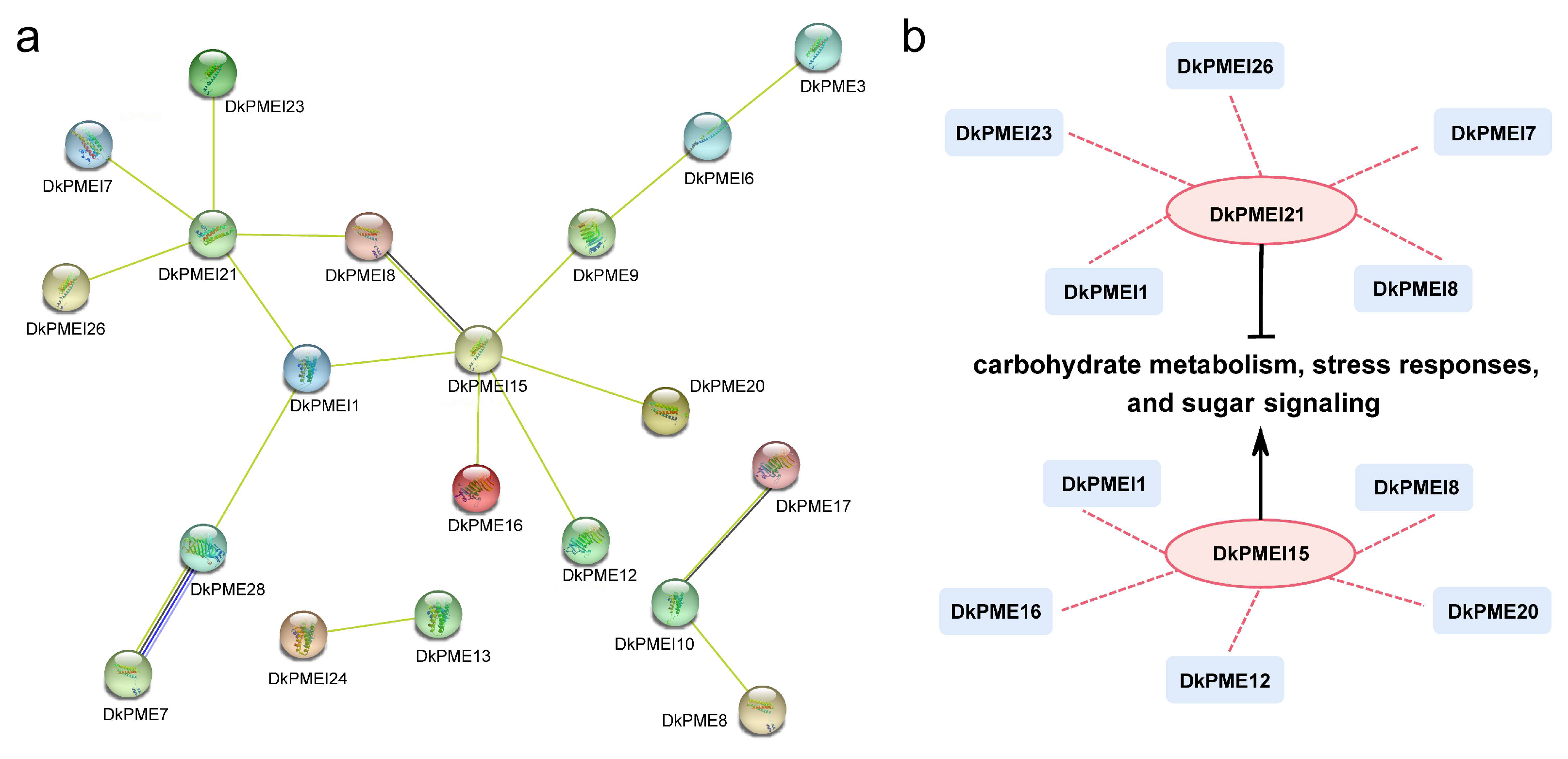
3.7. Protein Interaction Analysis of DkPMEs and DkPMEIs
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhao, Y.; Man, Y.; Wen, J.; Guo, Y.; Lin, J. Advances in imaging plant cell walls. Trends Plant Sci. 2019, 24, 867–878. [Google Scholar] [CrossRef]
- Daher, F.B.; Braybrook, S.A. How to let go: Pectin and plant cell adhesion. Front. Plant Sci. 2015, 6, 523. [Google Scholar] [CrossRef]
- Anderson, C.T. We be jammin’: An update on pectin biosynthesis, trafficking, and dynamics. J. Exp. Bot. 2015, 67, 495–502. [Google Scholar] [CrossRef] [PubMed]
- Caffall, K.H.; Mohnen, D. The structure, function, and biosynthesis of plant cell wall pectic polysaccharides. Carbohydr. Res. 2009, 344, 1879–1900. [Google Scholar] [CrossRef]
- Sénéchal, F.; Wattier, C.; Rustérucci, C.; Pelloux, J. Homogalacturonan-modifying enzymes: Structure, expression, and roles in plants. J. Exp. Bot. 2014, 65, 5125–5160. [Google Scholar] [CrossRef]
- Di, M.A.; Giovane, A.; Raiola, A.; Camardella, L.; Bonivento, D.; De, L.G.; Cervone, F.; Bellincampi, D.; Tsernoglou, D. Structural basis for the interaction between pectin methylesterase and a specific inhibitor protein. Plant Cell 2005, 17, 849–858. [Google Scholar] [CrossRef]
- Balestrieri, C.; Castaldo, D.; Giovane, A.; Quagliuolo, L.; Servillo, L. Aglycoprotein inhibitor of pectin methylesterase in kiwi fruit (Actinidia chinensis). Eur. J. Med. Chem. 1990, 193, 183–187. [Google Scholar] [CrossRef]
- Micheli, F. Pectin methylesterases: Cell wall enzymes with important roles in Plant Physiol. Trends Plant Sci. 2001, 6, 414–419. [Google Scholar] [CrossRef]
- Jolie, R.P.; Duvetter, T.; Van Loey, A.M.; Hendrickx, M.E. Pectin methylesterase and its proteinaceous inhibitor: A review. Carbohydr. Res. 2010, 345, 2583–2595. [Google Scholar] [CrossRef]
- Louvet, R.; Cavel, E.; Gutierrez, L.; Guénin, S.; Roge, D.; Gillet, F.; Guerineau, F.; Pelloux, J. Comprehensive expression profiling of the pectin methylesterase gene family during silique development in Arabidopsis thaliana. Planta 2006, 224, 782–791. [Google Scholar] [CrossRef]
- Li, Z.X.; Wu, L.M.; Wang, C.; Wang, Y.; He, L.G.; Wang, Z.J.; Ma, X.F.; Bai, F.X.; Feng, G.Z.; Liu, J.H.; et al. Characterization of pectin methylesterase gene family and its possible role in juice sac granulation in navel orange (Citrus sinensis Osbeck). BMC Genom. 2022, 23, 185. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Gao, Y.; Wang, S.; Zhang, Q.; Yang, S. Genome-wide identification of PME genes, evolution and expression analyses in soybean (Glycine max L.). BMC Plant Biol. 2021, 21, 578. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; He, H.; He, L.F. Genome-wide analysis of the pectin methylesterase gene family in potato. Potato Res. 2021, 64, 1–19. [Google Scholar] [CrossRef]
- Kagan, Z.V.; Tieman, D.M.; Marlow, S.J.; Handa, A.K. Differential regulation of polygalacturonase and pectin methylesterase gene expression during and after heat stress in ripening tomato (Lycopersicon esculentum Mill.) fruits. Plant Mol. Biol. 1995, 29, 1101–1110. [Google Scholar] [CrossRef] [PubMed]
- Wen, B.; Ström, A.; Tasker, A.; West, G.; Tucker, G.A. Effect of silencing the two major tomato fruit pectin methylesterase isoforms on cell wall pectin metabolism. Plant Biol. 2013, 15, 1025–1032. [Google Scholar] [CrossRef] [PubMed]
- Guénin, S.; Hardouin, J.; Paynel, F.; Müller, K.; Mongelard, G.; Driouich, A.; Lerouge, P.; Kermode, A.; Lehner, A.; Mollet, J.C.; et al. AtPME3, a ubiquitous cell wall pectin methylesterase of Arabidopsis thaliana, alters the metabolism of cruciferin seed storage proteins during post-germinative growth of seedlings. J. Exp. Bot. 2017, 68, 1083–1095. [Google Scholar] [CrossRef]
- Bosch, M.; Hepler, P.K. Pectin methylesterases and pectin dynamics in pollen tubes. Plant Cell 2005, 17, 3219–3226. [Google Scholar] [CrossRef]
- Jiang, L.X.; Yang, S.L.; Xie, L.F.; Li, F.; Puah, C.S.; Zhang, X.Q.; Yang, W.C.; Sundaresan, V.; Ye, D. Vanguard1 encodes a pectin methylesterase that enhances pollen tube growth in the arabidopsis style and transmitting tract. Plant Cell 2005, 17, 584–596. [Google Scholar] [CrossRef]
- Pilling, J.; Willmitzer, L.; Fisahn, J. Expression of a Petunia inflata pectin methyl esterase in Solanum tuberosum L. enhances stem elongation and modifies cation distribution. Planta 2000, 210, 391–399. [Google Scholar] [CrossRef]
- Yan, J.; He, H.; Fang, L.; Zhang, A. Pectin methylesterase31 positively regulates salt stress tolerance in Arabidopsis. Biochem. Biophys. Res. Commun. 2018, 496, 497–501. [Google Scholar] [CrossRef]
- Angelica, G.; Vincenzo, L.; Stefania, L.G.; Daniela, Z.; Eleonora, F.; Nathan, R.; Olga, A.Z.; Elisabetta, D.A.; Linda, M.; Daniela, B.; et al. Cell wall features transferred from common into durum wheat to improve Fusarium Head Blight resistance. Plant Sci. 2018, 274, 121–128. [Google Scholar] [CrossRef]
- Kohli, P.; Kalia, M.; Gupta, R. Pectin Methylesterases: A Review. J. Bioprocess. Biotech. 2015, 5, 1. [Google Scholar] [CrossRef]
- Giovane, A.; Balestrieri, C.; Quagliuolo, L.; Castaldo, D.; Servillo, L.A. Glycoprotein inhibitor of pectin methylesterase in kiwi fruit. Eur. J. Med. Chem. 1995, 233, 926–929. [Google Scholar] [CrossRef]
- Müller, K.; Levesque, T.G.; Fernandes, A.; Wormit, A.; Bartels, S.; Usadel, B.; Kermode, A. Overexpression of a pectin methylesterase inhibitor in Arabidopsis thaliana leads to altered growth morphology of the stem and defective organ separation. Plant Signal. Behav. 2013, 8, e26464. [Google Scholar] [CrossRef]
- Zhu, X.; Tang, C.; Li, Q.H.; Qiao, X.; Li, X.; Cai, Y.L.; Wang, P.; Sun, Y.Y.; Zhang, H.; Zhang, S.L.; et al. Characterization of the pectin methylesterase inhibitor gene family in Rosaceae and role of PbrPMEI23/39/41 in methylesterified pectin distribution in pear pollen tube. Planta 2021, 253, 118. [Google Scholar] [CrossRef]
- Nguyen, H.P.; Jeong, H.Y.; Kim, H.; Kim, Y.C.; Lee, C. Molecular and biochemical characterization of rice pectin methylesterase inhibitors (OsPMEIs). Plant Physiol. Biochem. 2016, 101, 105–112. [Google Scholar] [CrossRef]
- Li, B.; Wang, H.; He, S.; Ding, Z.T.; Wang, Y.; Li, N.N.; Hao, X.Y.; Wang, L.; Yang, Y.J.; Qian, W.J. Genome-Wide Identification of the PMEI Gene Family in Tea Plant and Functional Analysis of CsPMEI2 and CsPMEI4 Through Ectopic Overexpression. Front. Plant Sci. 2022, 12, 807514. [Google Scholar] [CrossRef]
- Ren, A.; Ahmed, R.I.; Chen, H.; Han, L.H.; Sun, J.H.; Ding, A.M.; Guo, Y.F.; Kong, Y.Z. Genome-wide identification, characterization and expression patterns of the pectin methylesterase inhibitor genes in sorghum bicolor. Genes 2019, 10, 755. [Google Scholar] [CrossRef]
- Pinzon-Latorre, D.; Deyholos, M.K. Pectinmethylesterases (PME) and Pectinmethylesterase Inhibitors (PMEI) enriched during phloem fiber development in Flax (Linum usitatissimum). PLoS ONE 2014, 9, e105386. [Google Scholar] [CrossRef]
- Liu, T.T.; Yu, H.; Xiong, X.P.; Yu, Y.J.; Yue, X.Y.; Liu, J.L.; Cao, J.S. Genome-wide identification and characterization of pectin methylesterase inhibitor genes in Brassica oleracea. Int. J. Mol. Sci. 2018, 19, 3338. [Google Scholar] [CrossRef]
- Liu, T.T.; Yu, H.; Xiong, X.P.; Yu, Y.J.; Yue, X.Y.; Liu, J.L.; Cao, J.S. Genome-wide identification, molecular evolution, and expression profiling analysis of pectin methylesterase inhibitor genes in Brassica campestris ssp. chinensis. Int. J. Mol. Sci. 2018, 19, 1338. [Google Scholar] [CrossRef]
- Lionetti, V.; Cervone, F.; Bellincampi, D. Methyl esterification of pectin plays a role during plant–pathogen interactions and affects plant resistance to diseases. J. Plant Physiol. 2012, 169, 1623–1630. [Google Scholar] [CrossRef]
- Sénéchal, F.; Mareck, A.; Marcelo, P.; Lerouge, P.; Pelloux, J. Arabidopsis PME17 activity can be controlled by pectin methylesterase Inhibitor4. Plant Signal. Behav. 2015, 10, e983351. [Google Scholar] [CrossRef] [PubMed]
- Müller, K.; Levesque, T.G.; Bartels, S.; Weitbrecht, K.; Wormit, A.; Usadel, B.; Haughn, G.; Kermode, A.R. Demethylesterification of cell wall pectins in arabidopsis plays a role in seed germination. Plant Physiol. 2012, 161, 305–316. [Google Scholar] [CrossRef]
- Nguyen, H.P.; Jeong, H.Y.; Jeon, S.H.; Kim, D.; Lee, C. Rice pectin methylesterase inhibitor28 (OsPMEI28) encodes a functional PMEI and its overexpression results in a dwarf phenotype through increased pectin methylesterification levels. J. Plant Physiol. 2017, 208, 17–25. [Google Scholar] [CrossRef]
- Luo, Z.; Wang, R. Persimmon in China: Domestication and traditional utilizations of genetic resources. JSTOR 2008, 22, 239–243. Available online: http://www.jstor.org/stable/42883463 (accessed on 6 December 2022).
- Giordani, E.; Doumett, S.; Nin, S.; Del Bubba, M. Selected primary and secondary metabolites in fresh persimmon (Diospyros kaki Thunb.): A review of analytical methods and current knowledge of fruit composition and health benefits. Food Res. Int. 2011, 44, 1752–1767. [Google Scholar] [CrossRef]
- Taira, S.; Ono, M.; Matsumoto, N. Reduction of persimmon astringency by complex formation between pectin and tannins. Postharvest Biol. Technol. 1997, 12, 265–271. [Google Scholar] [CrossRef]
- Wang, Y.; Li, K.K.; Li, C.M. Effects of interaction between pectin and tannin on the deastringency of different varieties of persimmons during maturing. Xiandai Shipin Keji 2019, 35, 87–94. [Google Scholar] [CrossRef]
- Awad, M. Persimmon Pectinmethylesterase: Extraction and variation during ripening. J. Food Sci. 1985, 50, 1643–1645. [Google Scholar] [CrossRef]
- Punta, M.; Coggill, P.C.; Eberhardt, R.Y.; Mistry, J.; Tate, J.; Boursnell, C.; Pang, N.; Forslund, K.; Ceric, G.; Clements, J.; et al. The Pfam protein families database. Nucleic Acids Res. 2011, 40, D290–D301. [Google Scholar] [CrossRef] [PubMed]
- Letunic, I.; Doerks, T.; Bork, P. SMART 7: Recent updates to the protein domain annotation resource. Nucleic Acids Res. 2011, 40, D302–D305. [Google Scholar] [CrossRef] [PubMed]
- Julio, C. 2-D Proteome Analysis Protocols (ed Andrew J. Link). Protein Sci. 1999, 8, 531–552. [Google Scholar] [CrossRef]
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. MEGA5: Molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol. Biol. Evol. 2011, 28, 2731–2739. [Google Scholar] [CrossRef]
- Larkin, M.A.; Blackshields, G.; Brown, N.P.; Chenna, R.; McGettigan, P.A.; McWilliam, H.; Valentin, F.; Wallace, I.M.; Wilm, A.; Lopez, R.; et al. Clustal W and Clustal X version 2.0. Bioinformatics 2007, 23, 2947–2948. [Google Scholar] [CrossRef] [PubMed]
- Szklarczyk, D.; Gable, A.L.; Nastou, K.C.; Lyon, D.; Kirsch, R.; Pyysalo, S.; Doncheva, N.T.; Legeay, M.; Fang, T.; Bork, P.; et al. The STRING database in 2021: Customizable protein–protein networks, and functional characterization of user-uploaded gene/measurement sets. Nucleic Acids Res. 2020, 49, D605–D612. [Google Scholar] [CrossRef] [PubMed]
- Bailey, T.L.; Johnson, J.; Grant, C.E.; Noble, W.S. The MEME Suite. Nucleic Acids Res. 2015, 43, W39–W49. [Google Scholar] [CrossRef]
- Chen, C.J.; Hao, C.; Yi, Z.; Hannah, R.T.; Margaret, H.F.; He, Y.H.; Xia, R. TBtools: An Integrative Toolkit Developed for Interactive Analyses of Big Biological Data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef]
- Wang, Y.P.; Tang, H.B.; Debarry, J.D.; Tan, X.; Li, J.P.; Wang, X.Y.; Lee, T.; Jin, H.Z.; Marler, B.; Guo, H.; et al. MCScanX: A toolkit for detection and evolutionary analysis of gene synteny and collinearity. Nucleic Acids Res. 2012, 40, e49. [Google Scholar] [CrossRef]
- Rombauts, S.; Déhais, P.; Van, M.; Rouzé, P. PlantCARE, a plant cis-acting regulatory element database. Nucleic Acids Res. 1999, 27, 295–296. [Google Scholar] [CrossRef]
- Li, J.F.; Miao, B.B.; Wang, S.X.; Dong, W.X.; Hou, S.; Si, C.C.; Wang, M.J. Hiplot: A comprehensive and easy-to-use web service for boosting publication-ready biomedical data visualization. Brief Bioinform. 2022, 23, bbac261. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.J.; Yuan, D.J.; Gao, W.H.; Li, Y.; Tan, J.F.; Zhang, X.L. A comparative genome analysis of PME and PMEI families reveals the evolution of pectin metabolism in plant cell walls. PLoS ONE 2013, 8, e72082. [Google Scholar] [CrossRef] [PubMed]
- Li, W.J.; Shang, H.H.; Ge, Q.; Zou, C.S.; Cai, J.; Wang, D.J.; Fan, S.M.; Zhang, Z.; Deng, X.Y.; Tan, Y.N.; et al. Genome-wide identification, phylogeny, and expression analysis of pectin methylesterases reveal their major role in cotton fiber development. BMC Genom. 2016, 17, 1000. [Google Scholar] [CrossRef]
- Li, Z.X.; Wang, C.; Long, D.; Jiang, Y.C.; He, L.G.; Wang, Z.J.; Ma, X.F.; Bai, F.X.; Liu, J.H.; Wu, L.M.; et al. Genome-wide identification, bioinformatics characterization and functional analysis of pectin methylesterase inhibitors related to low temperature-induced juice sac granulation in navel orange (Citrus sinensis Osbeck). Sci. Hortic. 2022, 298, 110983. [Google Scholar] [CrossRef]
- Zhu, Q.G.; Xu, Y.; Yang, Y.; Guan, C.F.; Zhang, Q.Y.; Huang, J.W.; Grierson, D.; Chen, K.S.; Gong, B.C.; Yin, X.R. The persimmon (Diospyros oleifera Cheng) genome provides new insights into the inheritance of astringency and ancestral evolution. Hortic. Res. 2019, 6, 15. [Google Scholar] [CrossRef]
- De, S.R.; Sabaghian, E.; Li, Z.; Saeys, Y.; Van-de, P.Y. Coordinated functional divergence of genes after genome duplication in Arabidopsis thaliana. Plant Cell 2017, 29, 2786–2800. [Google Scholar] [CrossRef]
- Panchy, N.; Lehti-Shiu, M.; Shiu, S.H. Evolution of gene duplication in plants. Plant Physiol. 2016, 171, 2294–2316. [Google Scholar] [CrossRef] [PubMed]
- Suo, Y.J.; Sun, P.; Cheng, H.H.; Han, W.J.; Diao, S.F.; Li, H.W.; Mai, Y.N.; Zhao, X.; Li, F.D.; Fu, J.M. A high-quality chromosomal genome assembly of Diospyros oleifera Cheng. GigaScience 2020, 1–10. [Google Scholar] [CrossRef]
- Sun, P.W.; Gao, Z.H.; Lv, F.F.; Yu, C.C.; Jin, Y.; Xu, Y.H.; Wei, J.H. Genome-wide analysis of basic helix–loop–helix (bHLH) transcription factors in Aquilaria sinensis. Sci. Rep. 2022, 12, 7194. [Google Scholar] [CrossRef]
- Zhang, P.P.; Wang, H.; Qin, X.; Chen, K.; Zhao, J.R.; Zhao, Y.X.; Yue, B. Genome-wide identification, phylogeny and expression analysis of the PME and PMEI gene families in maize. Sci. Rep. 2019, 9, 19918. [Google Scholar] [CrossRef]
- Wang, J.; Ling, L.; Cai, H.; Guo, C. Gene-wide identification and expression analysis of the PMEI family genes in soybean (Glycine max). 3 Biotech 2020, 10, 335. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Lyu, H.M.; Zhu, K.K.; Van-de, P.Y.; Cheng, Z.M.M. The emergence and evolution of intron-poor and intronless genes in intron-rich plant gene families. Plant J. 2021, 105, 1072–1082. [Google Scholar] [CrossRef] [PubMed]
- Omodele, I.; Christiaan, E.J.B.; Graeme, B. In silico analysis of cis-acting regulatory elements in 5′ regulatory regions of sucrose transporter gene families in rice (Oryza sativa Japonica) and Arabidopsis thaliana. Comput. Biol. Chem. 2010, 34, 268–283. [Google Scholar] [CrossRef]
- Xu, W.J.; Grain, D.; Bobet, S.; Le, G.; Thévenin, J.; Kelemen, Z.; Lepiniec, L.; Dubos, C. Complexity and robustness of the flavonoid transcriptional regulatory network revealed by comprehensive analyses of MYB–bHLH–WDR complexes and their targets in Arabidopsis seed. New Phytol. 2014, 202, 132–144. [Google Scholar] [CrossRef]
- Zhu, Q.G.; Gong, Z.Y.; Wang, M.M.; Li, X.; Grierson, D.; Yin, X.R.; Chen, K.S. A transcription factor network responsive to high CO2/hypoxia is involved in deastringency in persimmon fruit. J. Exp. Bot. 2018, 69, 2061–2070. [Google Scholar] [CrossRef]
- Damian, S.; Gable, A.L.; David, L.; Alexander, J.; Stefan, W.; Jaime, H.C.; Milan, S.; Nadezhda, T.D.; John, H.M.; Peer, B.; et al. STRING v11: Protein-protein association networks with increased coverage, supporting functional discovery in genome-wide experimental datasets. Nucleic Acids Res. 2018, 47, D607–D613. [Google Scholar] [CrossRef]
- Wang, J.; Wu, X.F.; Tang, Y.; Li, J.G.; Zhao, M.L. RNA-Seq provides new insights into the molecular events involved in “ball-skin versus bladder effect” on fruit cracking in litchi. Int. J. Mol. Sci. 2021, 22, 454. [Google Scholar] [CrossRef]
- Lionetti, V.; Raiola, A.; Mattei, B.; Bellincampi, D. The grapevine VvPMEI1 gene encodes a novel functional pectin methylesterase inhibitor associated to grape berry development. PLoS ONE 2015, 10, e0133810. [Google Scholar] [CrossRef]
- Reca, I.B.; Lionetti, V.; Camardella, L.; D’Avino, R.; Giardina, T.; Cervone, F.; Bellincampi, D. A functional pectin methylesterase inhibitor protein (SolyPMEI) is expressed during tomato fruit ripening and interacts with PME-1. Plant Mol. Biol. 2012, 79, 429–442. [Google Scholar] [CrossRef] [PubMed]
- Wen, B.; Zhang, F.; Wu, X.; Li, H. Characterization of the tomato (Solanum lycopersicum) pectin methylesterases: Evolution, activity of isoforms and expression during fruit ripening. Front. Plant Sci. 2020, 11, 238. [Google Scholar] [CrossRef]









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Q.; Pu, T.; Wang, Y.; Bai, Y.; Suo, Y.; Fu, J. Genome-Wide Identification and Expression Analysis of the PME and PMEI Gene Families in Diospyros kaki: A Bioinformatics Study. Horticulturae 2022, 8, 1159. https://doi.org/10.3390/horticulturae8121159
Zhang Q, Pu T, Wang Y, Bai Y, Suo Y, Fu J. Genome-Wide Identification and Expression Analysis of the PME and PMEI Gene Families in Diospyros kaki: A Bioinformatics Study. Horticulturae. 2022; 8(12):1159. https://doi.org/10.3390/horticulturae8121159
Chicago/Turabian StyleZhang, Qi, Tingting Pu, Yiru Wang, Yue Bai, Yujing Suo, and Jianmin Fu. 2022. "Genome-Wide Identification and Expression Analysis of the PME and PMEI Gene Families in Diospyros kaki: A Bioinformatics Study" Horticulturae 8, no. 12: 1159. https://doi.org/10.3390/horticulturae8121159
APA StyleZhang, Q., Pu, T., Wang, Y., Bai, Y., Suo, Y., & Fu, J. (2022). Genome-Wide Identification and Expression Analysis of the PME and PMEI Gene Families in Diospyros kaki: A Bioinformatics Study. Horticulturae, 8(12), 1159. https://doi.org/10.3390/horticulturae8121159