


Identification and Expression Analysis of the bHLH Gene Family Members in Diospyros kaki

 , ,
, ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. RNA Sequencing
2.3. Identification of bHLH Transcription Factors (TFs) in the D. kaki Transcriptome
2.4. Multiple Sequence Alignment, Conserved Motif Identification, and Phylogenetic Analysis
2.5. Gene Ontology Annotation
2.6. Protein Interaction Network Analysis
2.7. Calculating Ka and Ks of the Homologous bHLH Gene Pairs
2.8. In Silico Analysis of bHLH Genes in Different Tissues
3. Results
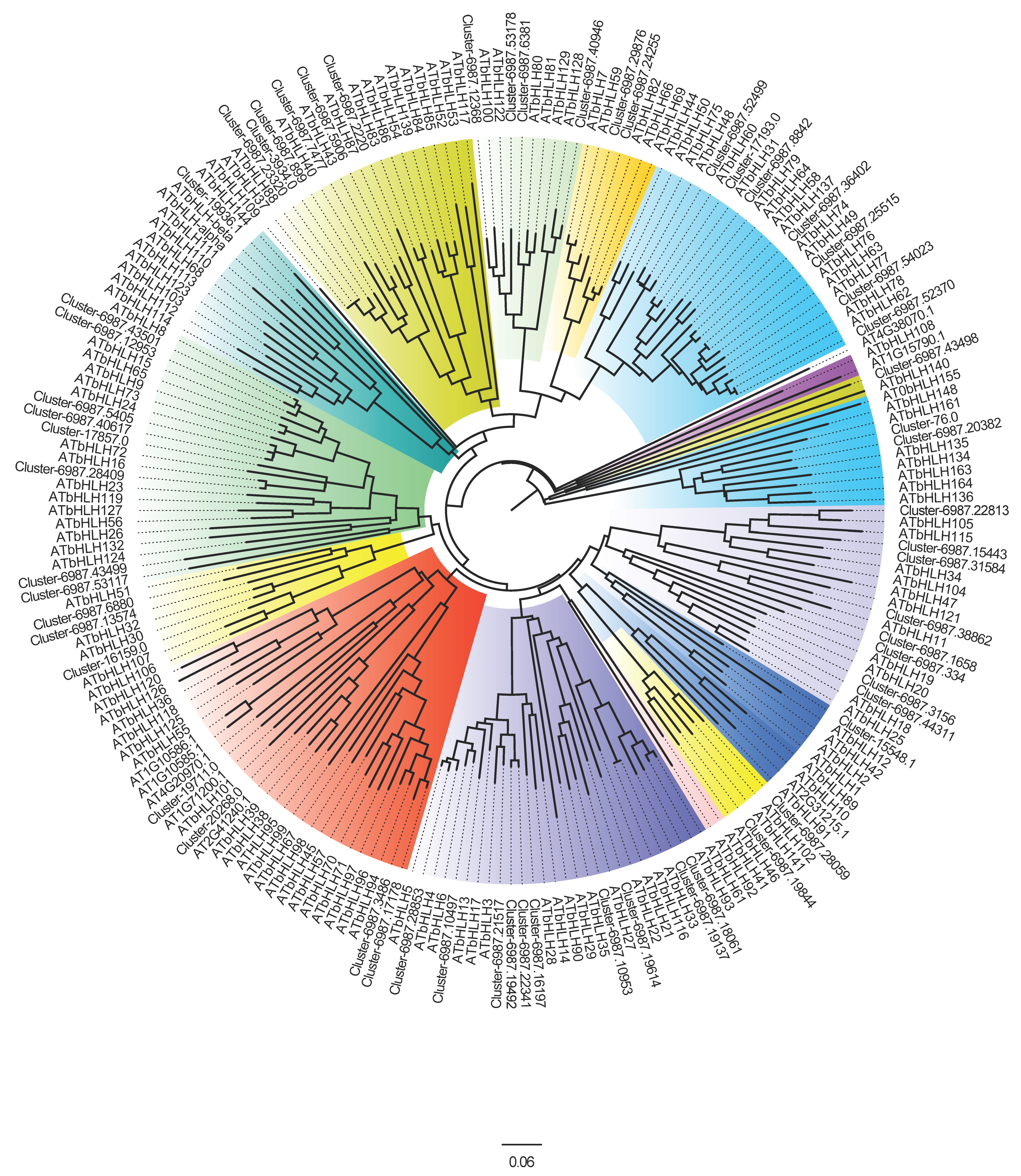
3.1. Identification and Phylogenetic Analysis of bHLH Genes in A. thaliana and D. kaki
3.2. Phylogenetic Analysis of the bHLH Proteins
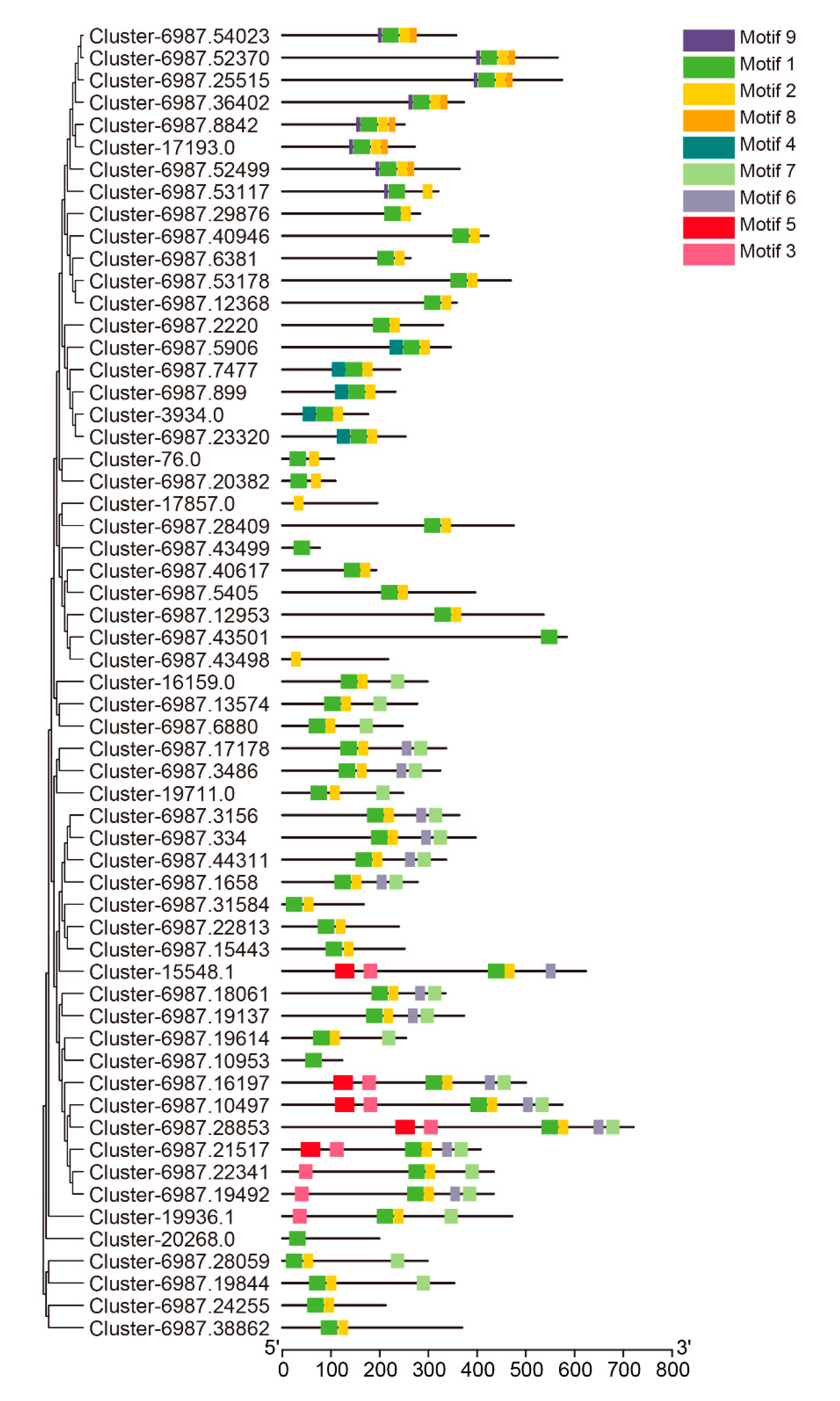
3.3. Conserved Motif Analysis
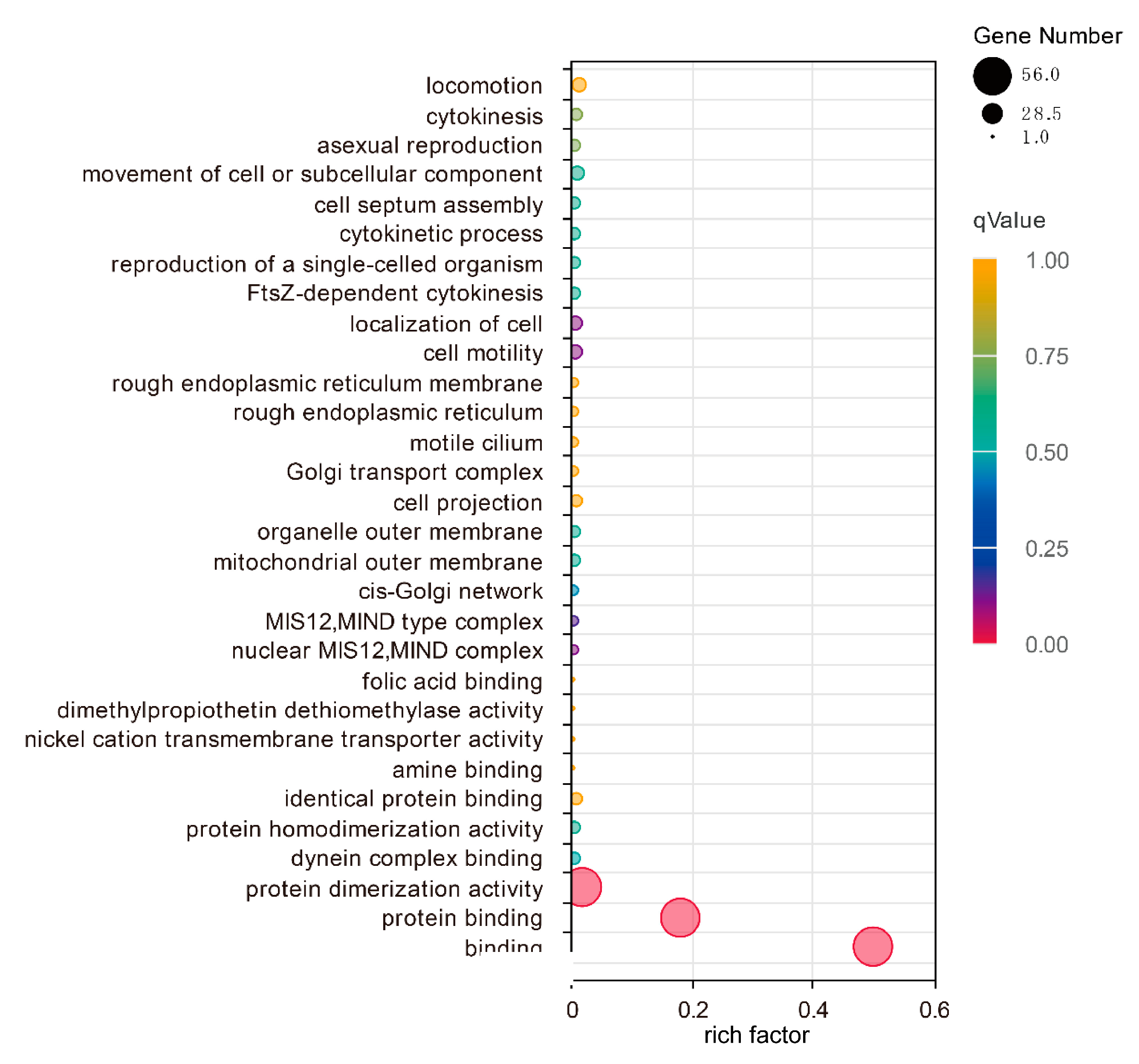
3.4. Gene Ontology Annotation
3.5. Selection Pressure and Differentiation Time of the Homologous bHLH Genes
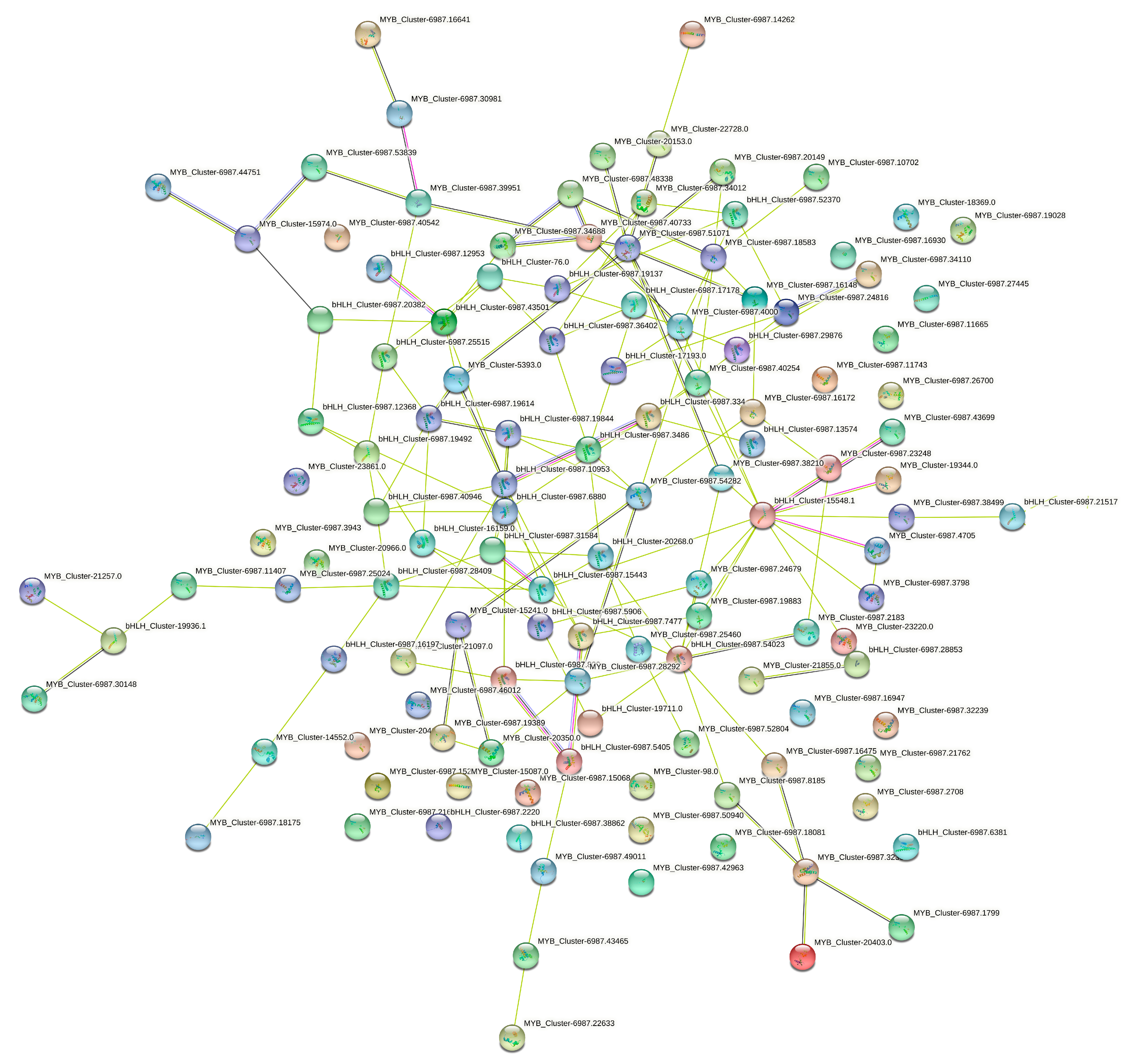
3.6. bHLH and MYB Protein–Protein Interaction Network Analysis
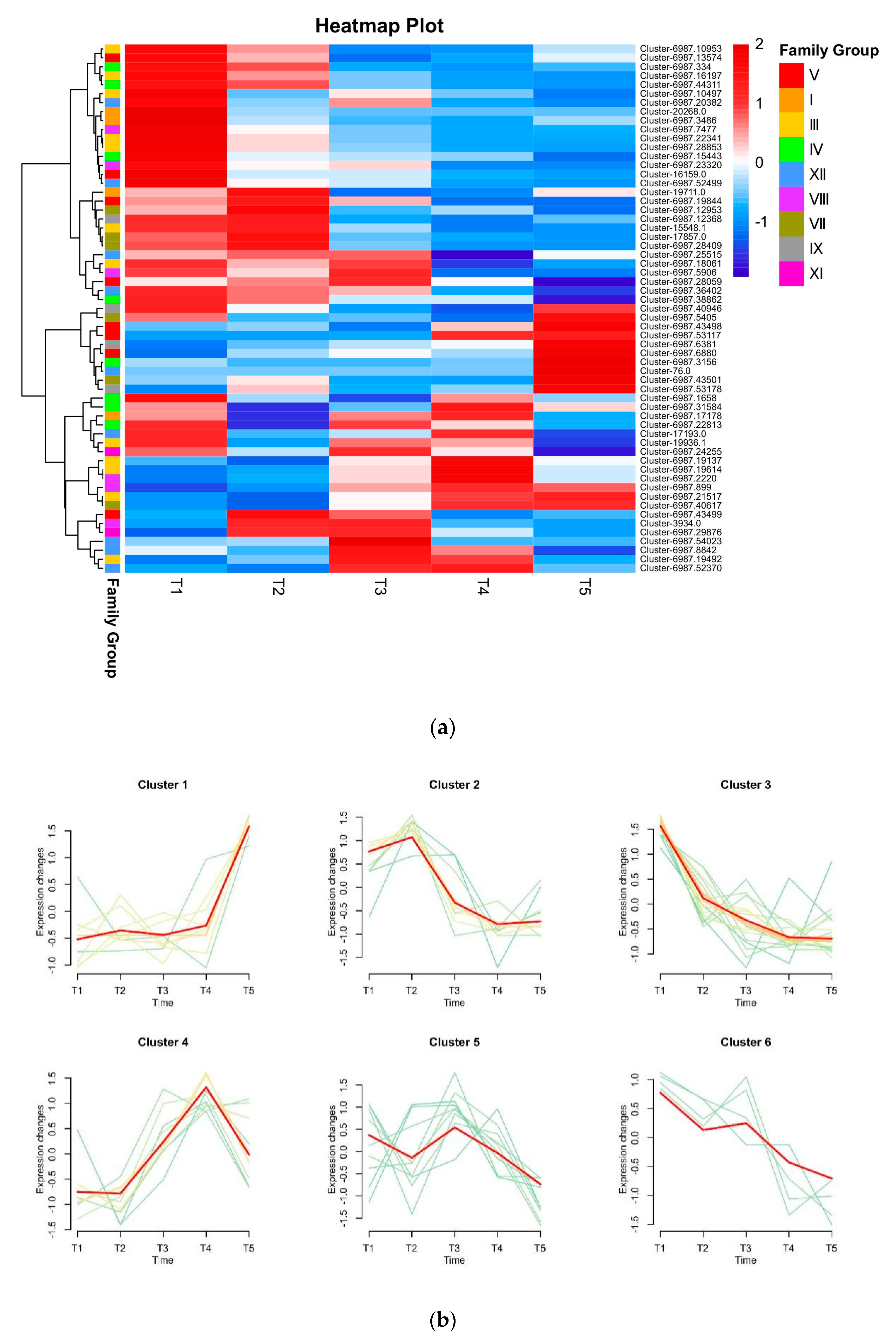
3.7. In Silico Analysis of bHLH Genes in D. kaki Fruit at Five Different Stages
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Castillon, A.; Shen, H.; Huq, E. Phytochrome interacting factors: Central players in phytochrome-mediated light signaling networks. Trends Plant Sci. 2007, 12, 514–521. [Google Scholar] [CrossRef]
- Duek, P.D.; Fankhauser, C. HFR1, a putative bHLH transcription factor, mediates both phytochrome a and cryptochrome signaling. Plant J. 2003, 34, 827–836. [Google Scholar] [CrossRef] [PubMed]
- Kondou, Y.; Nakazawa, M.; Kawashima, M.; Ichikawa, T.; Yoshizumi, T.; Suzuki, K.; Ishikawa, A.; Koshi, T.; Matsui, R.; Muto, S. RETARDED GROWTH OF EMBRYO1, a new basic helix-loop helix protein, in endosperm to control EMBRYO growth. Plant Physiol. 2008, 147, 1924–1935. [Google Scholar] [CrossRef]
- Toledo-Ortiz, G.; Huq, E.; Quail, P.H. The Arabidopsis basic/helix-loop-helix transcription factor family. Plant Cell 2003, 15, 1749–1770. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Peng, Z.; Kong, N.; Lu, R.; Pei, Y.; Huang, C.; Ma, H. Genome-wide identification and characterization of the Potato bHLH transcription factor family. Genes 2018, 9, 54. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.; Fan, H.J.; Ling, H.Q. Genome-wide identification and characterization of the bHLH gene family in tomato. BMC Genom. 2015, 16, 9. [Google Scholar] [CrossRef]
- Mao, K.; Dong, Q.; Li, C.; Liu, C.; Ma, F. Genome wide identification and characterization of apple bHLH transcription factors and expression analysis in response to drought and salt stress. Front. Plant Sci. 2017, 8, 480. [Google Scholar] [CrossRef]
- Li, X.; Duan, X.; Jiang, H.; Sun, Y.; Tang, Y.; Yuan, Z.; Guo, J.; Liang, W.; Chen, L.; Yin, J.; et al. Genome-wide analysis of basic/helix-loop-helix transcription factor family in rice and Arabidopsis. Plant Physiol. 2006, 141, 1167–1184. [Google Scholar] [CrossRef]
- Song, S.; Qi, T.; Fan, M.; Zhang, X.; Gao, H.; Huang, H.; Wu, D.; Guo, H.; Xie, D. The bHLH subgroup IIId factors negatively regulate jasmonate-mediated plant defense and development. PLoS Genet. 2013, 9, e1003653. [Google Scholar] [CrossRef]
- Liu, X.; An, X.; Liu, X.; Hu, D.; Wang, X.; You, C.; Hao, Y. MdSnRK1.1 interacts with MdJAZ18 to regulate sucrose-induced anthocyanin and proanthocyanidin accumulation in apple. J. Exp. Bot. 2017, 68, 2977–2990. [Google Scholar] [CrossRef]
- Chandler, V.L.; Radicella, J.P.; Robbins, T.P.; Chen, J.; Turks, D. Two regulatory genes of the maize anthocyanin pathway are homologous: Isolation of B utilizing R genomic sequences. Plant Cell 1989, 1, 1175–1183. [Google Scholar] [PubMed]
- Payne, C.T.; Zhang, F.; Lloyd, A.M. GL3 encodes a bHLH protein that regulates trichome development in Arabidopsis through interaction with GL1 and TTG1. Genetics 2000, 156, 1349–1362. [Google Scholar] [CrossRef]
- Li, M.; Sun, L.; Gu, H.; Cheng, D.; Guo, X.; Chen, R.C.; Wu, Z.; Jiang, J.; Fan, X.; Chen, J. Genome-wide characterization and analysis of bHLH transcription factors related to anthocyanin biosynthesis in spine grapes (Vitis davidii). Sci. Rep. 2021, 11, 6863. [Google Scholar] [CrossRef]
- Zhang, F.; Gonzalez, A.; Zhao, M.; Payne, C.T.; Lloyd, A. A network of redundant bHLH proteins functions in all TTG1-dependent pathways of Arabidopsis. Development 2003, 130, 4859–4869. [Google Scholar] [CrossRef] [PubMed]
- Neuffer, M.G.; Coe, E.H.; Wessler, S.R. Mutants of Maize, 1st ed.; Cold Spring Harbor Laboratory Press: New York, NY, USA, 1997; pp. 54–59. [Google Scholar]
- Petroni, K.; Cominelli, E.; Consonni, G.; Gusmaroli, G.; Gavazzi, G.; Tonelli, C. The developmental expression of the maize regulatory gene Hopi determines germination-dependent anthocyanin accumulation. Genetics 2000, 155, 323–336. [Google Scholar] [CrossRef] [PubMed]
- Rahim, M.A.; Busatto, N.; Trainotti, L. Regulation of anthocyanin biosynthesis in peach fruits. Planta 2014, 240, 913–929. [Google Scholar] [CrossRef]
- Hichri, I.; Heppel, S.C.; Pillet, J.; Leon, C.; Czemmel, S.; Delrot, S.; Virginie, L.; Bogs, J. The basic helix-loop-helix transcription factor MYC1 is involved in the gegulation of the flavonoid biosynthesis pathway in grapevine. Mol. Plant 2010, 3, 509–523. [Google Scholar] [CrossRef]
- Wada, T.; Kunihiro, A.; Tominaga-Wada, R.; Takaya, M. Arabidopsis CAPRICE (MYB) and GLABRA3 (bHLH) control tomato (Solanum lycopersicum) anthocyanin biosynthesis. PLoS ONE 2014, 9, e109093. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, Q.; Pu, T.; Suo, Y.; Han, W.; Diao, S.; Li, H.; Sun, P.; Fu, J. Transcriptomic profiling analysis to identify genes associated with PA biosynthesis and insolubilization in the late stage of fruit development in C-PCNA persimmon. Sci. Rep. 2022, 12, 19140. [Google Scholar] [CrossRef]
- Punta, M.; Coggill, P.C.; Eberhardt, R.Y.; Mistry, J.; Tate, J.; Boursnell, C.; Pang, N.; Forslund, K.; Ceric, G.; Clements, J. The Pfam protein families database. Nucleic Acids Res. 2004, 28, 263–266. [Google Scholar] [CrossRef]
- Wilkins, M.R.; Gasteiger, E.; Bairoch, A.; Sanchez, J.C.; Williams, K.L.; Appel, R.D.; Hochstrasser, D.F. Protein identification and analysis tools in the ExPASy server. Methods Mol. Biol. 1999, 112, 531–552. [Google Scholar] [PubMed]
- Jin, J.P.; Tian, F.; Yang, D.C.; Meng, Y.Q.; Kong, L.; Luo, J.C.; Gao, G. PlantTFDB 4.0: Toward a central hub for transcription factors and regulatory interactions in plants. Nucleic Acids Res. 2017, 45, D1040–D1045. [Google Scholar] [CrossRef] [PubMed]
- Jin, J.P.; He, K.; Tang, X.; Li, Z.; Lv, L.; Zhao, Y.; Luo, J.C.; Gao, G. An Arabidopsis transcriptional regulatory map reveals distinct functional and evolutionary features of novel transcription factors. Mol. Biol. Evol. 2015, 32, 1767–1773. [Google Scholar] [CrossRef] [PubMed]
- Jin, J.P.; Zhang, H.; Kong, L.; Gao, G.; Luo, J.C. PlantTFDB 3.0: A portal for the functional and evolutionary study of plant transcription factors. Nucleic Acids Res. 2014, 42, D1182–D1187. [Google Scholar] [CrossRef]
- Larkin, M.A.; Blackshields, G.; Brown, N.P.; Chenna, R.; Mcgettigan, P.A.; Mcwilliam, H.; Valentin, F.; Wallace, I.M.; Wilm, A.; Lopez, R. Clustal W and Clustal X version 2.0. Bioinformatics 2007, 21, 2947–2948. [Google Scholar] [CrossRef]
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. MEGA5: Molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol. Biol. Evol. 2011, 28, 2731–2739. [Google Scholar] [CrossRef]
- Heim, M.A.; Jakoby, M.; Werber, M.; Martin, C.; Weisshaar, B.; Bailey, P.C. The basic helix-loop-helix transcription factor family in plants: A genome-wide study of protein structure and functional diversity. Mol. Biol. Evol. 2003, 20, 735–747. [Google Scholar] [CrossRef]
- Bailey, T.L.; Johnson, J.; Grant, C.E.; Noble, W.S. The MEME suite. Nucleic Acids Res. 2015, 43, W39–W49. [Google Scholar] [CrossRef]
- Szklarczyk, D.; Gable, A.L.; Lyon, D.; Junge, A.; Wyder, S.; Huerta-Cepas, J.; Simonovic, M.; Doncheva, N.T.; Morris, J.H.; Bork, P. STRING v11: Protein-protein association networks with increased coverage, supporting functional discovery in genome-wide experimental datasets. Nucleic Acids Res. 2018, 47, D607–D613. [Google Scholar] [CrossRef]
- Li, W.H.; Gojobori, T.; Nei, M. Pseudogenes as a paradigm of neutral evolution. Nature 1981, 292, 237–239. [Google Scholar] [CrossRef]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Xia, R. TBtools: An integrative toolkit developed for interactive analyses of big biological data. Mol. Plant. 2020, 13, 1194–1202. [Google Scholar] [CrossRef]
- Wang, J.; Sun, P.; Li, Y.; Liu, Y.; Yu, J.; Ma, X.; Sun, S.; Yang, N.; Xia, R.; Lei, T. Hierarchically aligning 10 legume genomes establishes a family-level genomics platform. Plant Physiol. 2017, 174, 1981–2016. [Google Scholar] [CrossRef]
- Li, J.F.; Miao, B.B.; Wang, S.X.; Dong, W.X.; Hou, S.; Si, C.C.; Wang, M.J. Hiplot: A comprehensive and easy-to-use web service for boosting publication-ready biomedical data visualization. Brief Bioinform. 2022, 23, bbac261. [Google Scholar] [CrossRef] [PubMed]
- Escaray, F.J.; Passeri, V.; Perea-García, A.; Antonelli, C.J.; Damiani, F.; Ruiz, O.A.; Paolocci, F. The R2R3-MYB TT2b and the bHLH TT8 genes are the major regulators of proanthocyanidin biosynthesis in the leaves of Lotus species. Planta 2017, 246, 243–261. [Google Scholar] [CrossRef]
- Li, Y.; Shan, X.; Zhou, L.; Gao, R.; Yang, S.; Wang, S.; Wang, L.; Gao, X. The R2R3-MYB factor FhMYB5 from Freesia hybrida contributes to the regulation of anthocyanin and proanthocyanidin biosynthesis. Front. Plant Sci. 2019, 9, 1935. [Google Scholar] [CrossRef]
- Li, C.; Qiu, J.; Huang, S.; Yin, J.; Yang, G. AaMYB3 interacts with AabHLH1 to regulate proanthocyanidin accumulation in Anthurium andraeanum (Hort.)—Another strategy to modulate pigmentation. Hortic. Res. 2019, 6, 14. [Google Scholar] [CrossRef]
- Akagi, T.; Ikegami, A.; Tsujimoto, T.; Kobayashi, S.; Sato, A.; Kono, A.; Yonemori, K. DkMyb4 is a myb transcription factor involved in proanthocyanidin biosynthesis in persimmon fruit. Plant Physiol. 2009, 151, 2028–2045. [Google Scholar] [CrossRef]
- Chen, W.; Xiong, Y.; Xu, L.; Zhang, Q.; Luo, Z. An integrated analysis based on transcriptome and proteome reveals deastringency-related genes in CPCNA persimmon. Sci. Rep. 2017, 7, 44671. [Google Scholar] [CrossRef] [PubMed]
- Su, F.; Jia, H.; Zhang, Q.; Luo, Z. Isolation and characterization of a basic Helix-Loop-Helix transcription factor gene potentially involved in proanthocyanidin biosynthesis regulation in persimmon (Diospyros kaki Thunb.). Sci. Hortic. 2012, 136, 115–121. [Google Scholar] [CrossRef]
- Feyissa, D.N.; Lovdal, T.; Olsen, K.M.; Slimestad, R.; Lillo, C. The endogenous GL3, but not EGL3, gene is necessary for anthocyanin accumulation asinduced by nitrogen depletion in Arabidopsis rosette stage leaves. Planta 2009, 230, 747–754. [Google Scholar] [CrossRef]
- Xu, W.; Grain, D.; Le Gourrierec, J.; Harscoët, E.; Berger, A.; Jauvion, V.; Dubos, C. Regulation of flavonoid biosynthesis involves an unexpected complex transcriptional regulation of TT8 expression, in Arabidopsis. New Phytol. 2013, 198, 59–70. [Google Scholar] [CrossRef] [PubMed]
- Baudry, A.; Heim, M.A.; Dubreucq, B.; Caboche, M.; Weisshaar, B.; Lepiniec, L. TT2, TT8, and TTG1 synergistically specify the expression of BANYULS and proanthocyanidin biosynthesis in Arabidopsis thaliana. Plant J. 2004, 39, 366–380. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| GeneID | pI | MW | aa | Instability Index | GRAVY | Description | Family Group |
|---|---|---|---|---|---|---|---|
| Cluster-15548.1 | 5.92 | 70,353.3 | 622 | 57.15 | −0.493 | bHLH-MYC and R2R3-MYB transcription factors N-terminal | IIIf |
| Cluster-16159.0 | 8.59 | 33,034.44 | 297 | 70.62 | −0.511 | Helix–loop–helix DNA-binding domain | Vb |
| Cluster-17193.0 | 7.14 | 29,350.72 | 271 | 44.77 | −0.648 | Helix–loop–helix DNA-binding domain | XII |
| Cluster-17857.0 | 10.79 | 21,843.76 | 194 | 60.57 | −0.242 | Helix–loop–helix DNA-binding domain | VIIb |
| Cluster-19711.0 | 9.26 | 27,915.6 | 247 | 42.38 | −0.266 | Helix–loop–helix DNA-binding domain | Ib |
| Cluster-19936.1 | 5.94 | 52,936.87 | 472 | 53.04 | −0.464 | Helix–loop–helix DNA-binding domain | X |
| Cluster-20268.0 | 9.42 | 22,594.14 | 199 | 58.5 | −0.573 | Helix–loop–helix DNA-binding domain | Ib |
| Cluster-3934.0 | 10.3 | 19,350.09 | 175 | 64.74 | −0.582 | Helix–loop–helix DNA-binding domain | VIIIb |
| Cluster-6987.10497 | 5.7 | 63,669.96 | 574 | 54.13 | −0.656 | bHLH-MYC and R2R3-MYB transcription factors N-terminal | IIIe |
| Cluster-6987.10953 | 4.52 | 13,939.64 | 123 | 64.48 | −0.468 | Helix–loop–helix DNA-binding domain | IIIc |
| Cluster-6987.12368 | 8.32 | 39,649.34 | 357 | 55.24 | −0.618 | Helix–loop–helix DNA-binding domain | Ib |
| Cluster-6987.12953 | 5.85 | 57,668.09 | 535 | 75.43 | −0.549 | Helix–loop–helix DNA-binding domain | VIIa |
| Cluster-6987.13574 | 6.54 | 30,618.32 | 276 | 47.1 | −0.551 | Helix–loop–helix DNA-binding domain | Vb |
| Cluster-6987.15443 | 5.9 | 27,730.39 | 250 | 56.36 | −0.534 | Helix–loop–helix DNA-binding domain | IVc |
| Cluster-6987.16197 | 5.42 | 54,988.88 | 499 | 52.74 | −0.378 | bHLH-MYC and R2R3-MYB transcription factors N-terminal | IIIe |
| Cluster-6987.1658 | 9.56 | 30,647.86 | 277 | 62.46 | −0.626 | Helix–loop–helix DNA-binding domain | IVa |
| Cluster-6987.17178 | 5.4 | 37,385.92 | 335 | 62.07 | −0.523 | Helix–loop–helix DNA-binding domain | Ia |
| Cluster-6987.18061 | 4.62 | 37,271.96 | 334 | 53.27 | −0.478 | Helix–loop–helix DNA-binding domain | IIIb |
| Cluster-6987.19137 | 4.97 | 42,060.74 | 373 | 52.02 | −0.515 | Helix–loop–helix DNA-binding domain | IIIb |
| Cluster-6987.19492 | 8.2 | 47,950.04 | 433 | 50.54 | −0.527 | bHLH-MYC and R2R3-MYB transcription factors N-terminal | IIId |
| Cluster-6987.19614 | 6.29 | 28,597.66 | 253 | 62.5 | −0.385 | Helix–loop–helix DNA-binding domain | IIIc |
| Cluster-6987.19844 | 5.22 | 39,283.15 | 352 | 65.05 | −0.841 | Helix–loop–helix DNA-binding domain | Va |
| Cluster-6987.20382 | 9.79 | 12,432.12 | 108 | 82.07 | −0.719 | Helix–loop–helix DNA-binding domain | XII |
| Cluster-6987.21517 | 6.4 | 44,681.21 | 406 | 54.06 | −0.488 | bHLH-MYC and R2R3-MYB transcription factors N-terminal | IIId |
| Cluster-6987.2220 | 8.23 | 37,059.51 | 329 | 63.87 | −0.243 | Helix–loop–helix DNA-binding domain | VIIIb |
| Cluster-6987.22341 | 9.17 | 47,990.27 | 433 | 44.05 | −0.563 | bHLH-MYC and R2R3-MYB transcription factors N-terminal | IIId |
| Cluster-6987.22813 | 6.73 | 26,070.62 | 238 | 52.85 | −0.642 | Helix–loop–helix DNA-binding domain | IVc |
| Cluster-6987.23320 | 9.32 | 28,111.55 | 252 | 59.86 | −0.379 | Helix–loop–helix DNA-binding domain | VIIIb |
| Cluster-6987.24255 | 5.97 | 23,415.91 | 211 | 55.17 | −0.237 | Helix–loop–helix DNA-binding domain | XI |
| Cluster-6987.25515 | 5.39 | 62,110.75 | 574 | 50.52 | −0.48 | Helix–loop–helix DNA-binding domain | XII |
| Cluster-6987.28059 | 6.69 | 32,511.04 | 297 | 49.9 | −0.725 | Helix–loop–helix DNA-binding domain | Va |
| Cluster-6987.28409 | 6.37 | 50,973.69 | 474 | 52.91 | −0.489 | Helix–loop–helix DNA-binding domain | VIIb |
| Cluster-6987.28853 | 6.35 | 79,061.33 | 720 | 50.46 | −0.568 | bHLH-MYC and R2R3-MYB transcription factors N-terminal | IIIe |
| Cluster-6987.29876 | 9.95 | 30,252.81 | 282 | 34.57 | −0.016 | Helix–loop–helix DNA-binding domain | XI |
| Cluster-6987.3156 | 6.15 | 39,930.5 | 362 | 54.46 | −0.3 | Helix–loop–helix DNA-binding domain | IVa |
| Cluster-6987.31584 | 9.57 | 19,032.99 | 166 | 48.38 | −0.74 | Helix–loop–helix DNA-binding domain | IVc |
| Cluster-6987.334 | 5.87 | 44,580.72 | 397 | 51.11 | −0.452 | Helix–loop–helix DNA-binding domain | IVa |
| Cluster-6987.3486 | 5.49 | 35,928.41 | 323 | 64.54 | −0.522 | Helix–loop–helix DNA-binding domain | Ia |
| Cluster-6987.36402 | 6.3 | 40,786.68 | 372 | 49.98 | −0.615 | Helix–loop–helix DNA-binding domain | XII |
| Cluster-6987.38862 | 6.3 | 40,736.53 | 368 | 56.85 | −0.835 | Helix–loop–helix DNA-binding domain | IVb |
| Cluster-6987.40617 | 5.62 | 21,615.02 | 192 | 93.67 | −0.755 | Helix–loop–helix DNA-binding domain | VIIb |
| Cluster-6987.40946 | 9.61 | 45,283.41 | 422 | 52.67 | −0.574 | Helix–loop–helix DNA-binding domain | Ⅸ |
| Cluster-6987.43498 | 6.29 | 23,122.8 | 216 | 48.76 | −0.123 | Helix–loop–helix DNA-binding domain | VIIIb |
| Cluster-6987.43499 | 9.44 | 8641.81 | 76 | 77.43 | −0.779 | Helix–loop–helix DNA-binding domain | VIIa |
| Cluster-6987.43501 | 6.01 | 62,262.63 | 584 | 46.58 | −0.517 | Helix–loop–helix DNA-binding domain | VIIa |
| Cluster-6987.44311 | 6.1 | 36,970.1 | 335 | 37.89 | −0.31 | Helix–loop–helix DNA-binding domain | IVa |
| Cluster-6987.52370 | 8.67 | 61,252.56 | 564 | 58.26 | −0.61 | Helix–loop–helix DNA-binding domain | XII |
| Cluster-6987.52499 | 5.91 | 39,534.24 | 363 | 49.32 | −0.596 | Helix–loop–helix DNA-binding domain | XII |
| Cluster-6987.53117 | 9.11 | 35,833.89 | 319 | 55.41 | −0.522 | Helix–loop–helix DNA-binding domain | VIIa |
| Cluster-6987.53178 | 8.65 | 51,919.33 | 468 | 51.1 | −0.707 | Helix–loop–helix DNA-binding domain | Ⅸ |
| Cluster-6987.54023 | 7.2 | 40,160.13 | 356 | 52.22 | −0.714 | Helix–loop–helix DNA-binding domain | XII |
| Cluster-6987.5405 | 6.5 | 43,291.31 | 395 | 56.64 | −0.564 | Helix–loop–helix DNA-binding domain | VIIb |
| Cluster-6987.5906 | 5.84 | 38,000.7 | 345 | 59.52 | −0.532 | Helix–loop–helix DNA-binding domain | VIIIb |
| Cluster-6987.6381 | 5.76 | 28,349.49 | 262 | 65.3 | −0.589 | Helix–loop–helix DNA-binding domain | Ⅸ |
| Cluster-6987.6880 | 8.4 | 26,783.4 | 246 | 44.78 | −0.373 | Helix–loop–helix DNA-binding domain | Vb |
| Cluster-6987.7477 | 6.06 | 27,388.11 | 241 | 57.5 | −0.591 | Helix–loop–helix DNA-binding domain | VIIIb |
| Cluster-6987.8842 | 6.52 | 27,533.95 | 251 | 55.93 | −0.668 | Helix–loop–helix DNA-binding domain | XII |
| Cluster-6987.899 | 9.19 | 25,377.21 | 231 | 66.65 | −0.442 | Helix–loop–helix DNA-binding domain | VIIIb |
| Cluster-76.0 | 9.5 | 12,131.7 | 105 | 70.54 | −0.77 | Helix–loop–helix DNA-binding domain | XII |
| Seq_1 | Seq_2 | Ka | Ks | Ka:Ks | Differentiation Time |
|---|---|---|---|---|---|
| Cluster-6987.17178 | Cluster-6987.3486 | 0.34702 | 1.659578 | 0.209101 | 136.031 |
| Cluster-6987.38862 | AtbHLH011 | 0.532813 | 3.251332 | 0.163875 | 266.5026 |
| Cluster-6987.5405 | AtbHLH024 | 0.605611 | 2.374446 | 0.255054 | 194.6267 |
| Cluster-6987.28409 | AtbHLH016 | 0.223524 | 2.046914 | 0.1092 | 167.7798 |
| Cluster-6987.25515 | AtbHLH049 | 0.372177 | 1.674926 | 0.222205 | 137.289 |
| Cluster-6987.36402 | AtbHLH074 | 0.444855 | 2.575702 | 0.172712 | 211.1231 |
| Cluster-6987.22813 | AtbHLH105 | 0.165583 | 2.098734 | 0.078897 | 172.0274 |
| Cluster-6987.22341 | Cluster-6987.19492 | 0.17841 | 0.939712 | 0.189856 | 77.02557 |
| Cluster-6987.21517 | AtbHLH003 | 0.344623 | 1.905672 | 0.18084 | 156.2026 |
| Cluster-6987.5906 | AtbHLH087 | 0.449166 | 2.025877 | 0.221714 | 166.0555 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Han, W.; Zhang, Q.; Suo, Y.; Li, H.; Diao, S.; Sun, P.; Huang, L.; Fu, J. Identification and Expression Analysis of the bHLH Gene Family Members in Diospyros kaki. Horticulturae 2023, 9, 380. https://doi.org/10.3390/horticulturae9030380
Han W, Zhang Q, Suo Y, Li H, Diao S, Sun P, Huang L, Fu J. Identification and Expression Analysis of the bHLH Gene Family Members in Diospyros kaki. Horticulturae. 2023; 9(3):380. https://doi.org/10.3390/horticulturae9030380
Chicago/Turabian StyleHan, Weijuan, Qi Zhang, Yujing Suo, Huawei Li, Songfeng Diao, Peng Sun, Lin Huang, and Jianmin Fu. 2023. "Identification and Expression Analysis of the bHLH Gene Family Members in Diospyros kaki" Horticulturae 9, no. 3: 380. https://doi.org/10.3390/horticulturae9030380
APA StyleHan, W., Zhang, Q., Suo, Y., Li, H., Diao, S., Sun, P., Huang, L., & Fu, J. (2023). Identification and Expression Analysis of the bHLH Gene Family Members in Diospyros kaki. Horticulturae, 9(3), 380. https://doi.org/10.3390/horticulturae9030380