
Biostimulatory Action of Vegetal Protein Hydrolysate and the Configuration of Fruit Physicochemical Characteristics in Grafted Watermelon

 ,
,  ,
,  ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Growing Conditions
2.2. Biostimulant Characteristics and Application
2.3. Fruit Age Monitoring, Sampling Procedures, Yield, and Morphometric Traits
2.4. Fruit Pulp Texture and Colorimetry
2.5. Soluble Carbohydrates, Lycopene, and Titratable Acidity
2.6. Mineral Analysis
2.7. Experimental Design and Statistical Analysis
3. Results and Discussion
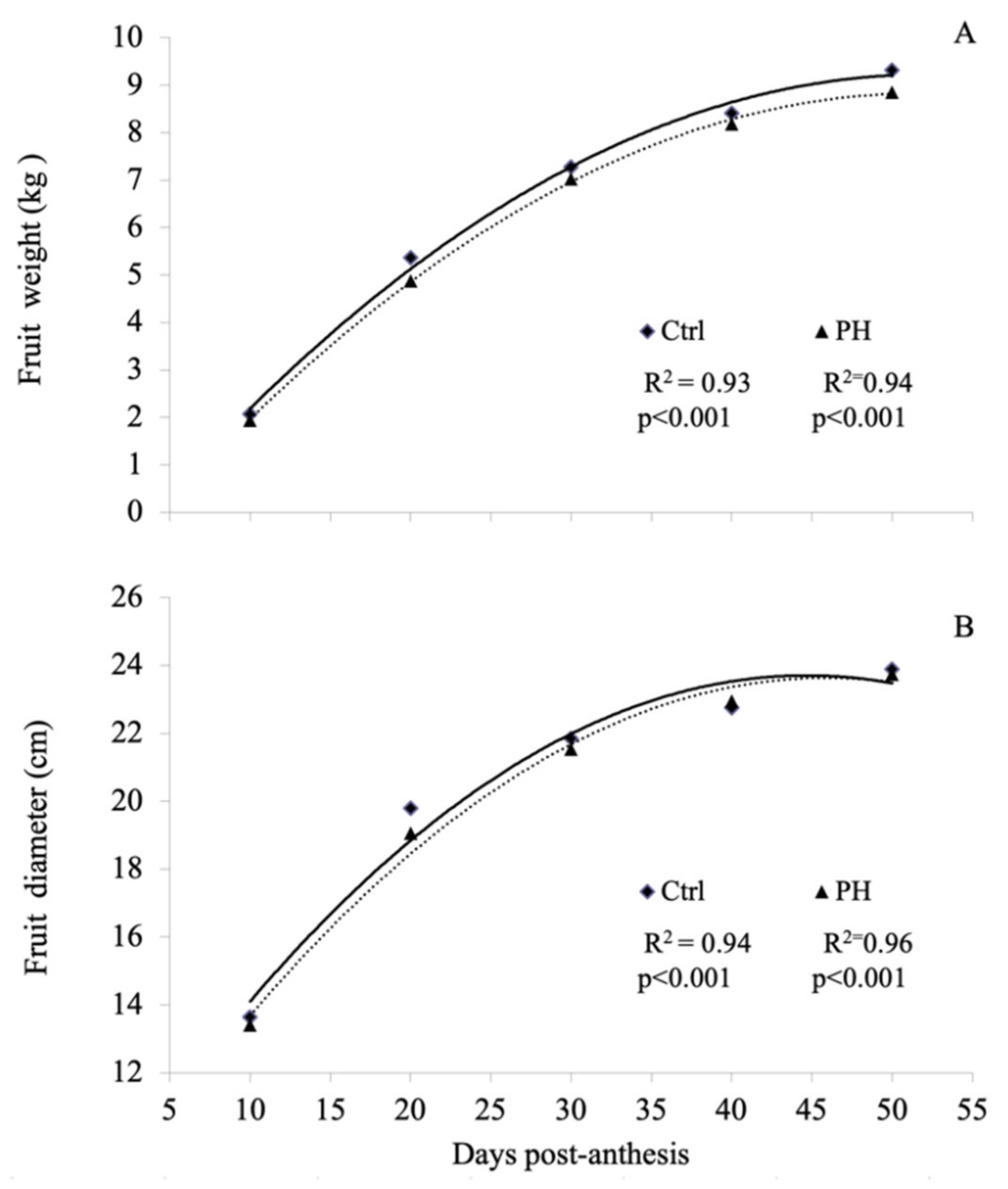
3.1. Yield Components and Fruit Morphological Characteristics
3.2. Fruit Pulp Texture and Rind Thickness
3.3. Colorimetry
3.4. Lycopene Content
3.5. Soluble Carbohydrates
3.6. Sweetness Index and Titratable Acidity
3.7. Mineral Content
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kyriacou, M.C.; Rouphael, Y.; Colla, G.; Zrenner, R.; Schwarz, D. Vegetable Grafting: The Implications of a Growing Agronomic Imperative for Vegetable Fruit Quality and Nutritive Value. Front. Plant Sci. 2017, 8, 741. [Google Scholar] [CrossRef]
- Rouphael, Y.; Kyriacou, M.C.; Colla, G. Vegetable Grafting: A Toolbox for Securing Yield Stability under Multiple Stress Conditions. Front. Plant Sci. 2018, 8, 2255. [Google Scholar] [CrossRef] [Green Version]
- Schwarz, D.; Rouphael, Y.; Colla, G.; Venema, J.H. Grafting as a Tool to Improve Tolerance of Vegetables to Abiotic Stresses: Thermal Stress, Water Stress and Organic Pollutants. Sci. Hortic. 2010, 127, 162–171. [Google Scholar] [CrossRef]
- Colla, G.; Hoagland, L.; Ruzzi, M.; Cardarelli, M.; Bonini, P.; Canaguier, R.; Rouphael, Y. Biostimulant Action of Protein Hydrolysates: Unraveling Their Effects on Plant Physiology and Microbiome. Front. Plant Sci. 2017, 8, 2202. [Google Scholar] [CrossRef] [Green Version]
- Colla, G.; Nardi, S.; Cardarelli, M.; Ertani, A.; Lucini, L.; Canaguier, R.; Rouphael, Y. Protein Hydrolysates as Biostimulants in Horticulture. Sci. Hortic. 2015, 196, 28–38. [Google Scholar] [CrossRef]
- Du Jardin, P. Plant Biostimulants: Definition, Concept, Main Categories and Regulation. Sci. Hortic. 2015, 196, 3–14. [Google Scholar] [CrossRef] [Green Version]
- Yakhin, O.I.; Lubyanov, A.A.; Yakhin, I.A.; Brown, P.H. Biostimulants in Plant Science: A Global Perspective. Front. Plant Sci. 2017, 7, 2049. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rouphael, Y.; Colla, G. Biostimulants in Agriculture. Front. Plant Sci. 2020, 11, 40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kyriacou, M.C.; Soteriou, G.A.; Rouphael, Y.; Siomos, A.S.; Gerasopoulos, D. Configuration of Watermelon Fruit Quality in Response to Rootstock-mediated Harvest Maturity and Postharvest Storage. J. Sci. Food Agric. 2016, 96, 2400–2409. [Google Scholar] [CrossRef]
- Fallik, E.; Ziv, C. How Rootstock/Scion Combinations Affect Watermelon Fruit Quality after Harvest? J. Sci. Food Agric. 2020, 100, 3275–3282. [Google Scholar] [CrossRef]
- Kyriacou, M.C.; Soteriou, G. Quality and Postharvest Performance of Watermelon Fruit in Response to Grafting on Interspecific Cucurbit Rootstocks. J. Food Qual. 2015, 38, 21–29. [Google Scholar] [CrossRef]
- Fathy, M.A.; Gabr, M.A.; El Shall, S.A. Effect of Humic Acid Treatments on ‘Canino’ apricot Growth, Yield and Fruit Quality. N. Y. Sci. J. 2010, 3, 109–115. [Google Scholar]
- Soppelsa, S.; Kelderer, M.; Casera, C.; Bassi, M.; Robatscher, P.; Matteazzi, A.; Andreotti, C. Foliar Applications of Biostimulants Promote Growth, Yield and Fruit Quality of Strawberry Plants Grown under Nutrient Limitation. Agronomy 2019, 9, 483. [Google Scholar] [CrossRef] [Green Version]
- Barrajón-Catalán, E.; Álvarez-Martínez, F.J.; Borras, F.; Perez, D.; Herrero, N.; Ruiz, J.J.; Micol, V. Metabolomic Analysis of the Effects of a Commercial Complex Biostimulant on Pepper Crops. Food Chem. 2020, 310, 125818. [Google Scholar] [CrossRef] [PubMed]
- Gonçalves, B.; Morais, M.C.; Sequeira, A.; Ribeiro, C.; Guedes, F.; Silva, A.P.; Aires, A. Quality Preservation of Sweet Cherry cv. ‘staccato’ by Using Glycine-Betaine or Ascophyllum nodosum. Food Chem. 2020, 322, 126713. [Google Scholar] [CrossRef]
- Colla, G.; Cardarelli, M.; Bonini, P.; Rouphael, Y. Foliar Applications of Protein Hydrolysate, Plant and Seaweed Extracts Increase Yield but Differentially Modulate Fruit Quality of Greenhouse Tomato. HortScience 2017, 52, 1214–1220. [Google Scholar] [CrossRef]
- Rouphael, Y.; Colla, G.; Giordano, M.; El-Nakhel, C.; Kyriacou, M.C.; De Pascale, S. Foliar Applications of a Legume-Derived Protein Hydrolysate Elicit Dose-Dependent Increases of Growth, Leaf Mineral Composition, Yield and Fruit Quality in Two Greenhouse Tomato Cultivars. Sci. Hortic. 2017, 226, 353–360. [Google Scholar]
- Ertani, A.; Pizzeghello, D.; Francioso, O.; Sambo, P.; Sanchez-Cortes, S.; Nardi, S. Capsicum chinensis L. Growth and Nutraceutical Properties Are Enhanced by Biostimulants in a Long-Term Period: Chemical and Metabolomic Approaches. Front. Plant Sci. 2014, 5, 375. [Google Scholar] [CrossRef] [Green Version]
- Devi, P.; Perkins-Veazie, P.; Miles, C. Impact of Grafting on Watermelon Fruit Maturity and Quality. Horticulturae 2020, 6, 97. [Google Scholar] [CrossRef]
- Frioni, T.; Sabbatini, P.; Tombesi, S.; Norrie, J.; Poni, S.; Gatti, M.; Palliotti, A. Effects of a Biostimulant Derived from the Brown Seaweed Ascophyllum nodosum on Ripening Dynamics and Fruit Quality of Grapevines. Sci. Hortic. 2018, 232, 97–106. [Google Scholar]
- Tarantino, A.; Lops, F.; Disciglio, G.; Lopriore, G. Effects of Plant Biostimulants on Fruit Set, Growth, Yield and Fruit Quality Attributes of ‘Orange Rubis®’Apricot (Prunus armeniaca L.) Cultivar in Two Consecutive Years. Sci. Hortic. 2018, 239, 26–34. [Google Scholar] [CrossRef]
- Soteriou, G.A.; Kyriacou, M.C. Rootstock-Mediated Effects on Watermelon Field Performance and Fruit Quality Characteristics. Int. J. Veg. Sci. 2015, 21, 344–362. [Google Scholar] [CrossRef]
- Paul, K.; Sorrentino, M.; Lucini, L.; Rouphael, Y.; Cardarelli, M.; Bonini, P.; Miras Moreno, M.B.; Reynaud, H.; Canaguier, R.; Trtílek, M. A Combined Phenotypic and Metabolomic Approach for Elucidating the Biostimulant Action of a Plant-Derived Protein Hydrolysate on Tomato Grown under Limited Water Availability. Front. Plant Sci. 2019, 10, 493. [Google Scholar] [CrossRef] [PubMed]
- McGuire, R.G. Reporting of Objective Color Measurements. HortScience 1992, 27, 1254–1255. [Google Scholar] [CrossRef] [Green Version]
- Fish, W.W.; Davis, A.R. The Effects of Frozen Storage Conditions on Lycopene Stability in Watermelon Tissue. J. Agric. Food Chem. 2003, 51, 3582–3585. [Google Scholar] [CrossRef] [PubMed]
- Kyriacou, M.C.; Soteriou, G.A.; Rouphael, Y. Modulatory Effects of Interspecific and Gourd Rootstocks on Crop Performance, Physicochemical Quality, Bioactive Components and Postharvest Performance of Diploid and Triploid Watermelon Scions. Agronomy 2020, 10, 1396. [Google Scholar] [CrossRef]
- Maynard, D.N. Watermelons: Characteristics, Production, and Marketing; ASHS Press: Ashcroft, BC, Canada, 2001; ISBN 0-9707546-1-2. [Google Scholar]
- Walters, S.A. Influence of Plant Density and Cultivar on Mini Triploid Watermelon Yield and Fruit Quality. HortTechnology 2009, 19, 553–557. [Google Scholar] [CrossRef] [Green Version]
- Sinnott, E.W. A Developmental Analysis of the Relation between Cell Size and Fruit Size in Cucurbits. Am. J. Bot. 1939, 26, 179–189. [Google Scholar] [CrossRef]
- Fukuoka, N.; Masuda, D.; Kanamori, Y. Effect of Timing of High Temperature Treatment around the Fruit on Sugar Accumulation in Watermelon (Citrullus Lanatus (Thunb.) Matsum. et Nakai). Environ. Control Biol. 2008, 46, 241–248. [Google Scholar] [CrossRef] [Green Version]
- Kyriacou, M.C.; Leskovar, D.I.; Colla, G.; Rouphael, Y. Watermelon and Melon Fruit Quality: The Genotypic and Agro-Environmental Factors Implicated. Sci. Hortic. 2018, 234, 393–408. [Google Scholar] [CrossRef]
- Soteriou, G.A.; Kyriacou, M.C.; Siomos, A.S.; Gerasopoulos, D. Evolution of Watermelon Fruit Physicochemical and Phytochemical Composition during Ripening as Affected by Grafting. Food Chem. 2014, 165, 282–289. [Google Scholar] [CrossRef]
- Fernando, D.; Milagrosa, S.; Francisco, C.; Francisco, M. Biostimulant Activity of Trichoderma saturnisporum in Melon (Cucumis melo). Hortscience 2018, 53, 810–815. [Google Scholar] [CrossRef]
- Soppelsa, S.; Kelderer, M.; Casera, C.; Bassi, M.; Robatscher, P.; Andreotti, C. Use of Biostimulants for Organic Apple Production: Effects on Tree Growth, Yield, and Fruit Quality at Harvest and during Storage. Front. Plant Sci. 2018, 9, 1342. [Google Scholar] [CrossRef]
- Trandel, M.A.; Johanningsmeier, S.; Schultheis, J.; Gunter, C.; Perkins-Veazie, P. Cell Wall Polysaccharide Composition of Grafted ‘Liberty’ Watermelon with Reduced Incidence of Hollow Heart Defect. Front. Plant Sci. 2021, 12, 278. [Google Scholar] [CrossRef]
- San Bautista, A.; Calatayud, A.; Nebauer, S.G.; Pascual, B.; Maroto, J.V.; López-Galarza, S. Effects of Simple and Double Grafting Melon Plants on Mineral Absorption, Photosynthesis, Biomass and Yield. Sci. Hortic. 2011, 130, 575–580. [Google Scholar] [CrossRef] [Green Version]
- Liu, N.; Yang, J.; Guo, S.; Xu, Y.; Zhang, M. Genome-Wide Identification and Comparative Analysis of Conserved and Novel MicroRNAs in Grafted Watermelon by High-Throughput Sequencing. PLoS ONE 2016, 8, e57359. [Google Scholar] [CrossRef] [PubMed]
- Soteriou, G.A.; Siomos, A.S.; Gerasopoulos, D.; Rouphael, Y.; Georgiadou, S.; Kyriacou, M.C. Biochemical and Histological Contributions to Textural Changes in Watermelon Fruit Modulated by Grafting. Food Chem. 2017, 237, 133–140. [Google Scholar] [CrossRef]
- Anees, M.; Gao, L.; Umer, M.J.; Yuan, P.; Zhu, H.; Lu, X.; He, N.; Gong, C.; Kaseb, M.O.; Zhao, S. Identification of Key Gene Networks Associated with Cell Wall Components Leading to Flesh Firmness in Watermelon. Front. Plant Sci. 2021, 12, 1164. [Google Scholar] [CrossRef] [PubMed]
- Proietti, S.; Rouphael, Y.; Colla, G.; Cardarelli, M.; De Agazio, M.; Zacchini, M.; Rea, E.; Moscatello, S.; Battistelli, A. Fruit Quality of Mini-watermelon as Affected by Grafting and Irrigation Regimes. J. Sci. Food Agric. 2008, 88, 1107–1114. [Google Scholar] [CrossRef]
- Perkins-Veazie, P.; Collins, J.K.; Davis, A.R.; Roberts, W. Carotenoid Content of 50 Watermelon Cultivars. J. Agric. Food Chem. 2006, 54, 2593–2597. [Google Scholar] [CrossRef] [PubMed]
- Perkins-Veazie, P.; Collins, J.K.; Pair, S.D.; Roberts, W. Lycopene Content Differs among Red-fleshed Watermelon Cultivars. J. Sci. Food Agric. 2001, 81, 983–987. [Google Scholar] [CrossRef]
- Ilahy, R.; Tlili, I.; Siddiqui, M.W.; Hdider, C.; Lenucci, M.S. Inside and beyond Color: Comparative Overview of Functional Quality of Tomato and Watermelon Fruits. Front. Plant Sci. 2019, 10, 769. [Google Scholar] [CrossRef] [Green Version]
- Magwaza, L.S.; Opara, U.L. Analytical Methods for Determination of Sugars and Sweetness of Horticultural Products—A Review. Sci. Hortic. 2015, 184, 179–192. [Google Scholar] [CrossRef]
- Roussos, P.A.; Denaxa, N.; Damvakaris, T. Strawberry Fruit Quality Attributes after Application of Plant Growth Stimulating Compounds. Sci. Hortic. 2009, 119, 138–146. [Google Scholar] [CrossRef]
- Brown, A.C., Jr.; Summers, W.L. Carbohydrate Accumulation and Color Development in Watermelon. J. Am. Soc. Hortic. Sci. 1985, 110, 683–687. [Google Scholar]
- Rouphael, Y.; Cardarelli, M.; Colla, G.; Rea, E. Yield, Mineral Composition, Water Relations, and Water Use Efficiency of Grafted Mini-Watermelon Plants under Deficit Irrigation. HortScience 2008, 43, 730–736. [Google Scholar] [CrossRef] [Green Version]
- Ertani, A.; Schiavon, M.; Nardi, S. Transcriptome-Wide Identification of Differentially Expressed Genes in Solanum lycopersicon L. in Response to an Alfalfa-Protein Hydrolysate Using Microarrays. Front. Plant Sci. 2017, 8, 1159. [Google Scholar] [CrossRef] [Green Version]
- Huang, Y.; Li, J.; Hua, B.; Liu, Z.; Fan, M.; Bie, Z. Grafting onto Different Rootstocks as a Means to Improve Watermelon Tolerance to Low Potassium Stress. Sci. Hortic. 2013, 149, 80–85. [Google Scholar] [CrossRef]
- Zhong, Y.; Chen, C.; Nawaz, M.A.; Jiao, Y.; Zheng, Z.; Shi, X.; Xie, W.; Yu, Y.; Guo, J.; Zhu, S. Using Rootstock to Increase Watermelon Fruit Yield and Quality at Low Potassium Supply: A Comprehensive Analysis from Agronomic, Physiological and Transcriptional Perspective. Sci. Hortic. 2018, 241, 144–151. [Google Scholar] [CrossRef]
- Gruda, N.; Savvas, D.; Colla, G.; Rouphael, Y. Impacts of Genetic Material and Current Technologies on Product Quality of Selected Greenhouse Vegetables—A Review. Eur. J. Hortic. Sci. 2018, 83, 319–328. [Google Scholar] [CrossRef]
- Bulgari, R.; Franzoni, G.; Ferrante, A. Biostimulants Application in Horticultural Crops under Abiotic Stress Conditions. Agronomy 2019, 9, 306. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
| Biostimulant | Yield | Fruit Number | Fruit Weight | Fruit Diameter |
|---|---|---|---|---|
| (t ha–1) | (Fruits ha–1) | (kg) | (cm) | |
| Control | 79.1 ± 4.6 | 10,181 ± 412 | 7.77 ± 0.3 | 22.76 ± 0.32 |
| PH | 80.6 ± 8.2 | 10,181 ± 548 | 7.80 ± 0.4 | 22.96 ± 0.20 |
| ns | ns | ns | ns |
| Source of Variance | Firmness | Rind | L * | C * | h° | a * | Lycopene | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| (kg-force) | (mm) | (0–100) | √ (a *2 + b *2) | (0–360 °) | (0–60) | (μg g−1 f.w.) | ||||||||
| Biostimulant | ||||||||||||||
| Control | 4.15 ± 0.17 | 9.42 ± 0.53 | 52.76 ± 2.71 | 26.11 ± 2.23 | 47.57 ± 5.96 | 19.96 ± 2.60 | 49.42 ± 7.31 | a | ||||||
| PH | 3.96 ± 0.15 | 9.41 ± 0.67 | 53.76 ± 2.98 | 25.53 ± 2.27 | 48.16 ± 6.17 | 19.51 ± 2.61 | 44.67 ± 6.71 | b | ||||||
| ns | ns | ns | ns | ns | ns | * | ||||||||
| Days post-anthesis | ||||||||||||||
| 10 | 4.91 ± 0.12 | a | 4.55 ± 0.31 | b | 77.42 ± 0.56 | a | 8.63 ± 0.28 | d | 103.06 ± 0.25 | a | –1.94 ± 0.07 | d | 0.27 ± 0.07 | e |
| 20 | 4.72 ± 0.10 | a | 10.16 ± 0.47 | a | 54.70 ± 0.65 | b | 20.55 ± 0.67 | c | 34.80 ± 0.96 | b | 16.88 ± 0.68 | c | 23.43 ± 1.52 | d |
| 30 | 3.65 ± 0.13 | b | 11.34 ± 0.35 | a | 45.95 ± 0.64 | c | 31.36 ± 0.58 | b | 32.03 ± 0.27 | c | 26.56 ± 0.43 | b | 55.96 ± 1.47 | c |
| 40 | 3.42 ± 0.11 | b | 10.68 ± 0.37 | a | 42.41 ± 0.64 | d | 36.35 ± 0.58 | a | 31.94 ± 0.47 | c | 30.79 ± 0.44 | a | 87.95 ± 2.55 | a |
| 50 | 3.38 ± 0.23 | b | 10.74 ± 0.66 | a | 42.57 ± 0.93 | d | 34.97 ± 0.66 | a | 33.0 ± 0.25 | bc | 29.27 ± 0.52 | a | 76.79 ± 2.97 | b |
| *** | *** | *** | *** | *** | *** | *** | ||||||||
| Source of Variance | SSC (%) | Fructose (μg mL−1) | Glucose (μg mL−1) | Sucrose (μg mL−1) | Total Sugars (μg mL−1) | Sweetness Index | TA (%) | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Biostimulant | ||||||||||||||
| Control | 10.48 ± 0.63 | 30.47 ± 1.74 | 24.75 ± 1.45 | 28.54 ± 4.95 | 83.76 ± 5.50 | 932 ± 58 | 0.17 ± 0.01 | |||||||
| PH | 10.34 ± 0.66 | 31.34 ± 2.16 | 25.70 ± 1.80 | 24.82 ± 4.89 | 81.86 ± 5.86 | 915 ± 63 | 0.16 ± 0.01 | |||||||
| ns | ns | ns | ns | ns | ns | ns | ||||||||
| Days post-anthesis | ||||||||||||||
| 10 | 4.95 ± 0.17 | c | 18.16 ± 0.91 | d | 16.28 ± 0.88 | d | 0.01 ± 0.01 | d | 34.45 ± 1.78 | c | 395 ± 20 | d | 0.16 ± 0.01 | b |
| 20 | 9.99 ± 0.20 | b | 40.42 ± 0.57 | a | 35.08 ± 0.66 | a | 4.66 ± 0.67 | d | 80.16 ± 1.59 | b | 920 ± 17 | c | 0.20 ± 0.01 | a |
| 30 | 12.37 ± 0.18 | a | 40.34 ± 0.96 | a | 31.35 ± 0.92 | b | 29.03 ± 2.29 | c | 100.71 ± 1.29 | a | 1137 ± 12 | a | 0.17 ± 0.01 | b |
| 40 | 12.93 ± 0.18 | a | 29.76 ± 0.51 | b | 23.51 ± 0.64 | c | 51.57 ± 1.68 | b | 104.84 ± 1.08 | a | 1143 ± 10 | a | 0.13 ± 0.01 | c |
| 50 | 12.43 ± 0.14 | a | 23.62 ± 0.61 | c | 17.55 ± 0.63 | d | 57.58 ± 1.63 | a | 98.76 ± 0.97 | a | 1066 ± 9 | b | 0.15 ± 0.01 | bc |
| *** | *** | *** | *** | *** | *** | *** | ||||||||
| Source of Variance | K (g/kg d.m.) | Mg (g/kg d.m.) | Ca (g/kg d.m.) | P (g/kg d.m.) | DM (%) | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| Biostimulant | ||||||||||
| Control | 14.19 ± 0.40 | b | 1.49 ± 0.04 | 0.86 ± 0.07 | 1.71 ± 0.09 | 10.17 ± 0.6 | ||||
| PH | 15.70 ± 0.79 | a | 1.54 ± 0.03 | 1.03 ± 0.13 | 1.61 ± 0.06 | 10.86 ± 0.8 | ||||
| * | ns | ns | ns | ns | ||||||
| Days post-anthesis | ||||||||||
| 10 | 19.04 ± 1.53 | a | 1.34 ± 0.09 | b | 1.77 ± 0.17 | a | 1.15 ± 0.09 | b | 6.51 ± 1.62 | c |
| 20 | 14.97 ± 0.31 | b | 1.52 ± 0.04 | a | 1.01 ± 0.06 | b | 1.70 ± 0.08 | a | 9.53 ± 0.19 | b |
| 30 | 12.70 ± 0.35 | c | 1.55 ± 0.02 | a | 0.73 ± 0.04 | bc | 1.66 ± 0.07 | a | 12.15 ± 0.2 | a |
| 40 | 14.35 ± 0.39 | bc | 1.56 ± 0.04 | a | 0.59 ± 0.03 | c | 1.94 ± 0.07 | a | 12.54 ± 0.14 | a |
| 50 | 13.71 ± 0.37 | bc | 1.62 ± 0.04 | a | 0.63 ± 0.05 | c | 1.83 ± 0.07 | a | 12.16 ± 0.22 | a |
| *** | *** | *** | *** | *** |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Soteriou, G.A.; Rouphael, Y.; Emmanouilidou, M.G.; Antoniou, C.; Kyratzis, A.C.; Kyriacou, M.C. Biostimulatory Action of Vegetal Protein Hydrolysate and the Configuration of Fruit Physicochemical Characteristics in Grafted Watermelon. Horticulturae 2021, 7, 313. https://doi.org/10.3390/horticulturae7090313
Soteriou GA, Rouphael Y, Emmanouilidou MG, Antoniou C, Kyratzis AC, Kyriacou MC. Biostimulatory Action of Vegetal Protein Hydrolysate and the Configuration of Fruit Physicochemical Characteristics in Grafted Watermelon. Horticulturae. 2021; 7(9):313. https://doi.org/10.3390/horticulturae7090313
Chicago/Turabian StyleSoteriou, Georgios A., Youssef Rouphael, Maria G. Emmanouilidou, Chrystalla Antoniou, Angelos C. Kyratzis, and Marios C. Kyriacou. 2021. "Biostimulatory Action of Vegetal Protein Hydrolysate and the Configuration of Fruit Physicochemical Characteristics in Grafted Watermelon" Horticulturae 7, no. 9: 313. https://doi.org/10.3390/horticulturae7090313
APA StyleSoteriou, G. A., Rouphael, Y., Emmanouilidou, M. G., Antoniou, C., Kyratzis, A. C., & Kyriacou, M. C. (2021). Biostimulatory Action of Vegetal Protein Hydrolysate and the Configuration of Fruit Physicochemical Characteristics in Grafted Watermelon. Horticulturae, 7(9), 313. https://doi.org/10.3390/horticulturae7090313