
Expression Profiling of Heat Shock Protein Genes as Putative Early Heat-Responsive Members in Lettuce



Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Material and Growth Conditions
2.2. Morphometric Analysis
2.3. RNA Preparation and Transcriptome Sequencing Analysis
2.4. Data Analysis Quality Control
2.5. Differentially Expressed Genes (DEG)
2.6. Quantitative RT-PCR (qRT-PCR) Analysis
2.7. Phylogenetic Analysis
3. Results
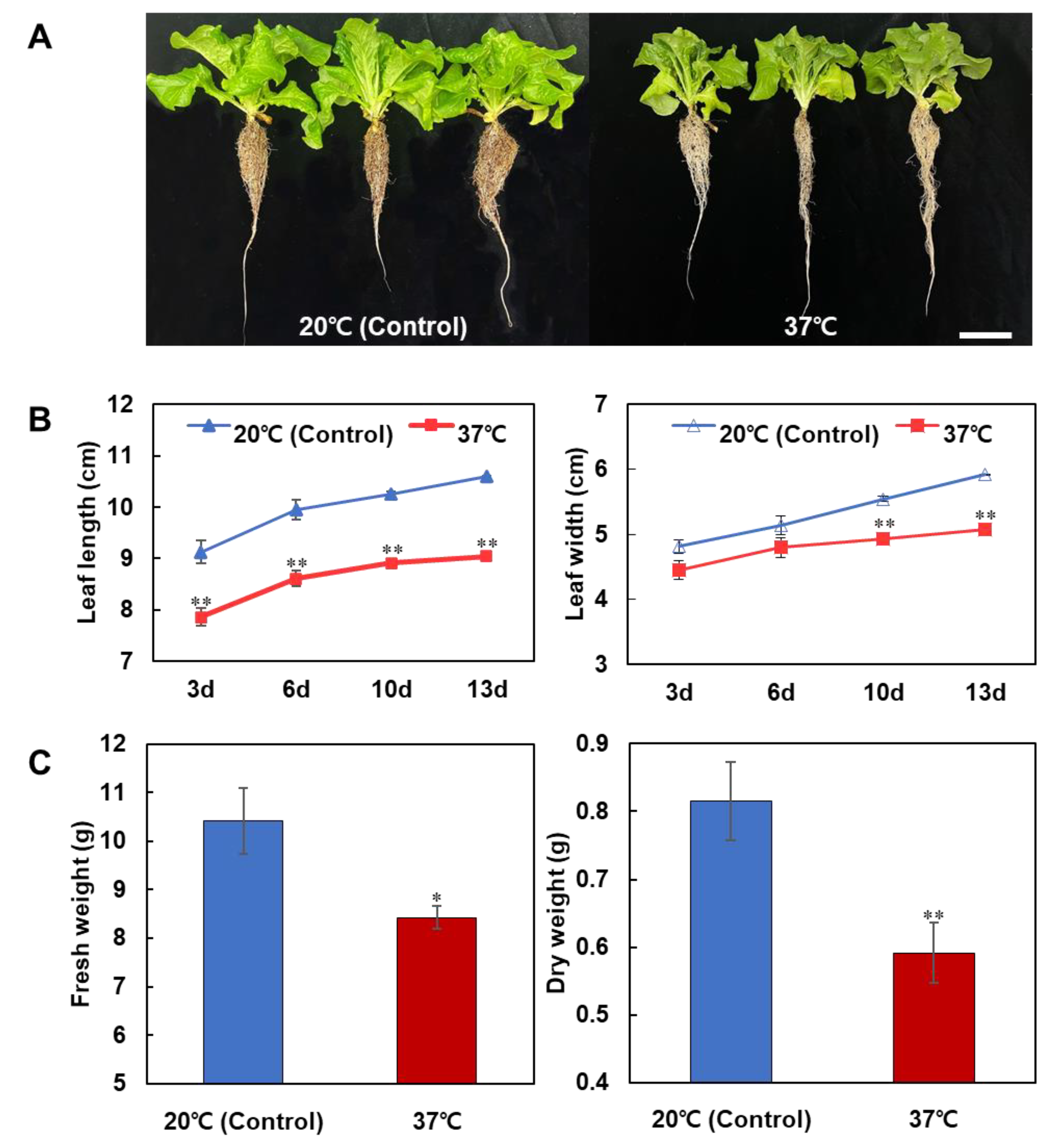
3.1. Effects of Heat Stress on the Early Vegetative Growth of Lettuce
3.2. Heat Treatment Induced the Expression of a Heat Shock Protein Gene, LsHSP70-2711 in Lettuce
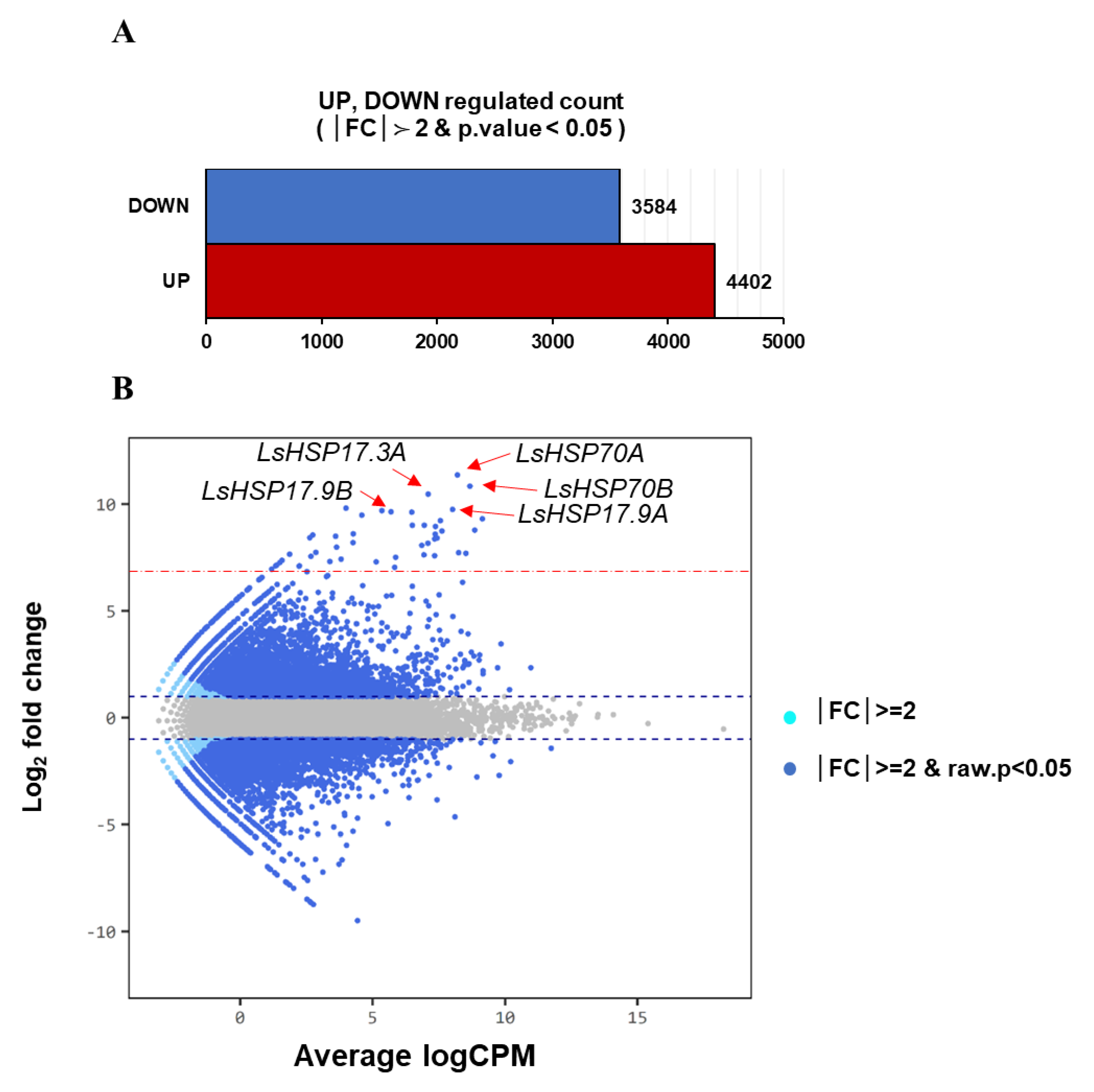
3.3. Transcriptome Profiling of Lettuce with Heat Treatment
3.4. Differentially Expressed Gene Analysis
3.5. Experimental Validation of Gene Expression by qRT-PCR
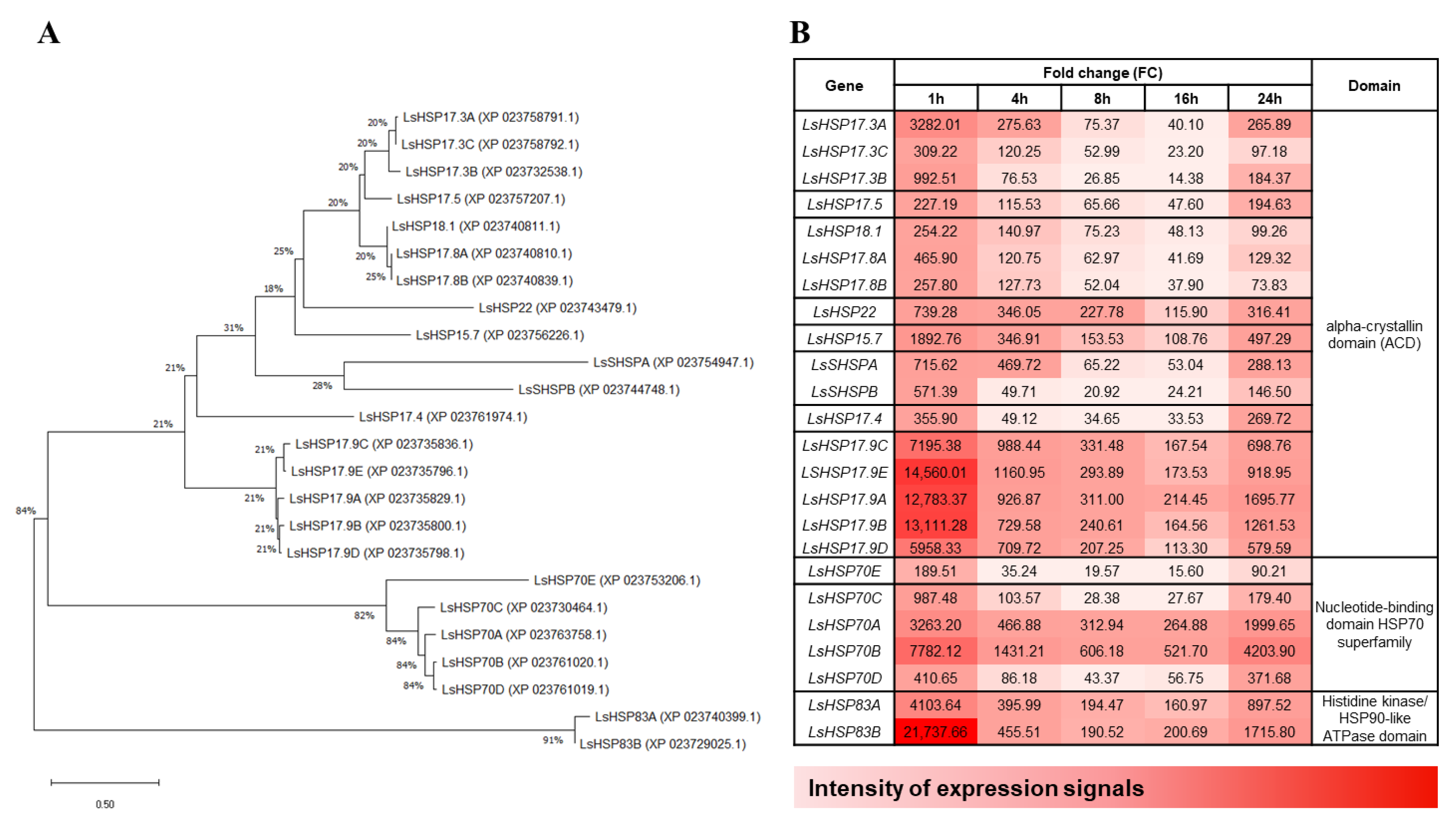
3.6. Time-Course Gene Expression Patterns of the Lettuce HSPs and Their Phylogenetic Relationships
4. Discussion
4.1. Effects of Heat Stress Treatment on the Early Vegetative Growth of Lettuce
4.2. Heat Stress Treatment Increased the Expression of LsHSP70-2711 in Lettuce
4.3. Transcriptome Profiling of Heat-Treated Lettuce and DEG Analysis
4.4. Temporal Expression Patterns of Lettuce HSP Genes under Heat Stress
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Shatilov, M.V.; Razin, A.F.; Ivanova, M.I. Analysis of the world lettuce market. IOP Conf. Ser. Earth Environ. Sci. 2019, 395, 012053. [Google Scholar] [CrossRef]
- Wallace, R.; Wszelaki, A.; Miles, C.; Cowan, J.; Martin, J.; Roozen, J.; Gundersen, B.; Inglis, D. Lettuce Yield and Quality When Grown in High Tunnel and Open-Field Production Systems Under Three Diverse Climates. HortTechnology 2012, 22, 659–668. [Google Scholar] [CrossRef]
- Lee, A.-C.; Liao, F.-S.; Lo, H.-F. Temperature, Daylength, and Cultivar Interact to Affect the Growth and Yield of Lettuce Grown in High Tunnels in Subtropical Regions. HortScience 2015, 50, 1412. [Google Scholar] [CrossRef]
- Porter, J.R. Rising temperatures are likely to reduce crop yields. Nature 2005, 436, 174. [Google Scholar] [CrossRef]
- NOAA. State of the Climate: Global Climate Report for Annual 2020; National Centers for Environmental Information: Asheville, NC, USA, 2021. [Google Scholar]
- Mearns, L.O.; Katz, R.W.; Schneider, S.H. Extreme high-temperature events: Changes in their probabilities with changes in mean temperature. J. Appl. Meteorol. Climatol. 1984, 23, 1601–1613. [Google Scholar] [CrossRef]
- Peng, S.; Huang, J.; Sheehy, J.E.; Laza, R.C.; Visperas, R.M.; Zhong, X.; Centeno, G.S.; Khush, G.S.; Cassman, K.G. Rice yields decline with higher night temperature from global warming. Proc. Natl. Acad. Sci. USA 2004, 101, 9971–9975. [Google Scholar] [CrossRef] [PubMed]
- Nascimento, W.M. Preventing thermoinhibition in a thermosensitive lettuce genotype by seed imbibition at low temperature. Sci. Agric. 2003, 60, 477–480. [Google Scholar] [CrossRef][Green Version]
- Wien, H.C. The Physiology of Vegetable Crops; CAB International: Wallingford, UK, 1997. [Google Scholar]
- Wu, G.; Zhang, C.; Chu, L.-Y.; Shao, H.-B. Responses of higher plants to abiotic stresses and agricultural sustainable development. J. Plant Interact. 2007, 2, 135–147. [Google Scholar] [CrossRef]
- Wahid, A.; Gelani, S.; Ashraf, M.; Foolad, M.R. Heat tolerance in plants: An overview. Environ. Exp. Bot. 2007, 61, 199–223. [Google Scholar] [CrossRef]
- Mittler, R. Abiotic stress, the field environment and stress combination. Trends Plant Sci 2006, 11, 15–19. [Google Scholar] [CrossRef]
- Rai, G.K.; Kumar, R.R.; Bagati, S. Abiotic Stress Tolerance Mechanisms in Plants; CRC Press: Boca Raton, FL, USA, 2021; Volume 370. [Google Scholar]
- Bhattacharya, A. Chapter 3—Effect of High Temperature on Protein Metabolism in Plants. In Effect of High Temperature on Crop Productivity and Metabolism of Macro Molecules; Bhattacharya, A., Ed.; Academic Press: Cambridge, MA, USA, 2019; pp. 217–309. [Google Scholar]
- Gupta, S.C.; Sharma, A.; Mishra, M.; Mishra, R.K.; Chowdhuri, D.K. Heat shock proteins in toxicology: How close and how far? Life Sci. 2010, 86, 377–384. [Google Scholar] [CrossRef]
- Hu, W.; Hu, G.; Han, B. Genome-wide survey and expression profiling of heat shock proteins and heat shock factors revealed overlapped and stress specific response under abiotic stresses in rice. Plant Sci. 2009, 176, 583–590. [Google Scholar] [CrossRef]
- Kotak, S.; Larkindale, J.; Lee, U.; von Koskull-Döring, P.; Vierling, E.; Scharf, K.D. Complexity of the heat stress response in plants. Curr. Opin. Plant Biol. 2007, 10, 310–316. [Google Scholar] [CrossRef]
- Morimoto, R.I.; Santoro, M.G. Stress-inducible responses and heat shock proteins: New pharmacologic targets for cytoprotection. Nat. Biotechnol. 1998, 16, 833–838. [Google Scholar] [CrossRef] [PubMed]
- Park, S.; Shi, A.; Mou, B. Genome-wide identification and expression analysis of the CBF/DREB1 gene family in lettuce. Sci. Rep. 2020, 10, 5733. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Xu, S.; Cheng, Y.; Peng, Z.; Han, J. Transcriptome profiling of anthocyanin-related genes reveals effects of light intensity on anthocyanin biosynthesis in red leaf lettuce. PeerJ 2018, 6, e4607. [Google Scholar] [CrossRef] [PubMed]
- Reyes-Chin-Wo, S.; Wang, Z.; Yang, X.; Kozik, A.; Arikit, S.; Song, C.; Xia, L.; Froenicke, L.; Lavelle, D.O.; Truco, M.-J.; et al. Genome assembly with in vitro proximity ligation data and whole-genome triplication in lettuce. Nat. Commun. 2017, 8, 14953. [Google Scholar] [CrossRef]
- Borowski, J.M.; Galli, V.; da Silva Messias, R.; Perin, E.C.; Buss, J.H.; dos Anjos e Silva, S.D.; Rombaldi, C.V. Selection of candidate reference genes for real-time PCR studies in lettuce under abiotic stresses. Planta 2014, 239, 1187–1200. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Liu, R.; Su, Z.; Zhou, H.; Huang, Q.; Fan, S.; Liu, C.; Han, Y. LsHSP70 is induced by high temperature to interact with calmodulin, leading to higher bolting resistance in lettuce. Sci. Rep. 2020, 10, 15155. [Google Scholar] [CrossRef]
- Fahad, S.; Bajwa, A.A.; Nazir, U.; Anjum, S.A.; Farooq, A.; Zohaib, A.; Sadia, S.; Nasim, W.; Adkins, S.; Saud, S.; et al. Crop Production under Drought and Heat Stress: Plant Responses and Management Options. Front. Plant Sci. 2017, 8, 1147. [Google Scholar] [CrossRef]
- Barnabás, B.; Jäger, K.; Fehér, A. The effect of drought and heat stress on reproductive processes in cereals. Plant Cell Env. 2008, 31, 11–38. [Google Scholar] [CrossRef] [PubMed]
- Demirevska-Kepova, K.; Holzer, R.; Simova-Stoilova, L.; Feller, U. Heat stress effects on ribulose-1,5-bisphosphate carboxylase/oxygenase, Rubisco binding protein and Rubisco activase in wheat leaves. Biol. Plant. 2005, 49, 521–525. [Google Scholar] [CrossRef]
- Hatfield, J.L.; Prueger, J.H. Temperature extremes: Effect on plant growth and development. Weather Clim. Extrem. 2015, 10, 4–10. [Google Scholar] [CrossRef]
- Holmes, S.C.; Wells, D.E.; Pickens, J.M.; Kemble, J.M. Selection of Heat Tolerant Lettuce (Lactuca sativa L.) Cultivars Grown in Deep Water Culture and Their Marketability. Horticulturae 2019, 5, 50. [Google Scholar] [CrossRef]
- Al-Whaibi, M.H. Plant heat-shock proteins: A mini review. J. King Saud Univ. Sci. 2011, 23, 139–150. [Google Scholar] [CrossRef]
- Ul Haq, S.; Khan, A.; Ali, M.; Khattak, A.M.; Gai, W.X.; Zhang, H.X.; Wei, A.M.; Gong, Z.H. Heat Shock Proteins: Dynamic Biomolecules to Counter Plant Biotic and Abiotic Stresses. Int. J. Mol. Sci. 2019, 20, 5321. [Google Scholar] [CrossRef] [PubMed]
- Timperio, A.M.; Egidi, M.G.; Zolla, L. Proteomics applied on plant abiotic stresses: Role of heat shock proteins (HSP). J. Proteom. 2008, 71, 391–411. [Google Scholar] [CrossRef] [PubMed]
- Keller, M.; Bokszczanin, K.L.; Bostan, H.; Bovy, A.; Chaturvedi, P.; Chen, Y.; Chiusano, M.L.; Firon, N.; Fragkostefanakis, S.; Iannacone, R.; et al. The coupling of transcriptome and proteome adaptation during development and heat stress response of tomato pollen. BMC Genom. 2018, 19, 447. [Google Scholar] [CrossRef]
- Kang, W.-H.; Sim, Y.M.; Koo, N.; Nam, J.-Y.; Lee, J.; Kim, N.; Jang, H.; Kim, Y.-M.; Yeom, S.-I. Transcriptome profiling of abiotic responses to heat, cold, salt, and osmotic stress of Capsicum annuum L. Sci. Data 2020, 7, 17. [Google Scholar] [CrossRef]
- Yang, Z.; Li, W.; Su, X.; Ge, P.; Zhou, Y.; Hao, Y.; Shu, H.; Gao, C.; Cheng, S.; Zhu, G.; et al. Early Response of Radish to Heat Stress by Strand-Specific Transcriptome and miRNA Analysis. Int. J. Mol. Sci. 2019, 20, 3321. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Jiang, B.; Liu, W.; Lin, Y.E.; Liang, Z.; He, X.; Peng, Q. Transcriptome Analyses Provide Novel Insights into Heat Stress Responses in Chieh-Qua (Benincasa hispida Cogn. var. Chieh-Qua How). Int. J. Mol. Sci. 2019, 20, 883. [Google Scholar] [CrossRef] [PubMed]
- Tripp, J.; Mishra, S.K.; Scharf, K.D. Functional dissection of the cytosolic chaperone network in tomato mesophyll protoplasts. Plant Cell Environ. 2009, 32, 123–133. [Google Scholar] [CrossRef]
- Derocher, A.E.; Helm, K.W.; Lauzon, L.M.; Vierling, E. Expression of a Conserved Family of Cytoplasmic Low Molecular Weight Heat Shock Proteins during Heat Stress and Recovery. Plant Physiol. 1991, 96, 1038–1047. [Google Scholar] [CrossRef]
- Usman, M.G.; Rafii, M.; Ismail, M.; Malek, M.A.; Latif, M.; Oladosu, Y. Heat Shock Proteins: Functions and Response against Heat Stress in Plants. Int. J. Sci. Technol. Res. 2014, 3, 204–218. [Google Scholar]
- Ye, S.; Yu, S.; Shu, L.; Wu, J.; Wu, A.; Luo, L. Expression profile analysis of 9 heat shock protein genes throughout the life cycle and under abiotic stress in rice. Chin. Sci. Bull. 2012, 57, 336–343. [Google Scholar] [CrossRef]
- Mansfield, M.A.; Key, J.L. Synthesis of the Low Molecular Weight Heat Shock Proteins in Plants 1. Plant Physiol. 1987, 84, 1007–1017. [Google Scholar] [CrossRef] [PubMed]
- Nagao, R.T.; Czarnecka, E.; Gurley, W.B.; Schöffl, F.; Key, J.L. Genes for low-molecular-weight heat shock proteins of soybeans: Sequence analysis of a multigene family. Mol. Cell Biol. 1985, 5, 3417–3428. [Google Scholar] [CrossRef]
- Sun, W.; Van Montagu, M.; Verbruggen, N. Small heat shock proteins and stress tolerance in plants. Biochim. Biophys. Acta Gene Struct. Expr. 2002, 1577, 1–9. [Google Scholar] [CrossRef]
- Seo, J.S.; Lee, Y.-M.; Park, H.G.; Lee, J.-S. The intertidal copepod Tigriopus japonicus small heat shock protein 20 gene (Hsp20) enhances thermotolerance of transformed Escherichia coli. Biochem. Biophys. Res. Commun. 2006, 340, 901–908. [Google Scholar] [CrossRef]
- Ferguson, D.L.; Guikema, J.A.; Paulsen, G.M. Ubiquitin Pool Modulation and Protein Degradation in Wheat Roots during High Temperature Stress. Plant Physiol. 1990, 92, 740–746. [Google Scholar] [CrossRef] [PubMed]
- Mogk, A.; Bukau, B. Role of sHsps in organizing cytosolic protein aggregation and disaggregation. Cell Stress Chaperones 2017, 22, 493–502. [Google Scholar] [CrossRef] [PubMed]
- Yu, A.; Li, P.; Tang, T.; Wang, J.; Chen, Y.; Liu, L. Roles of Hsp70s in Stress Responses of Microorganisms, Plants, and Animals. BioMed Res. Int. 2015, 2015, 510319. [Google Scholar] [CrossRef] [PubMed]
- Hartl, F.U.; Hayer-Hartl, M. Molecular chaperones in the cytosol: From nascent chain to folded protein. Science 2002, 295, 1852–1858. [Google Scholar] [CrossRef]
- Mayer, M.P.; Bukau, B. Hsp70 chaperones: Cellular functions and molecular mechanism. Cell Mol. Life Sci. 2005, 62, 670–684. [Google Scholar] [CrossRef]
- Zhu, X.; Zhao, X.; Burkholder, W.F.; Gragerov, A.; Ogata, C.M.; Gottesman, M.E.; Hendrickson, W.A. Structural analysis of substrate binding by the molecular chaperone DnaK. Science 1996, 272, 1606–1614. [Google Scholar] [CrossRef]
- Chakafana, G.; Zininga, T.; Shonhai, A. The Link That Binds: The Linker of Hsp70 as a Helm of the Protein’s Function. Biomolecules 2019, 9, 543. [Google Scholar] [CrossRef]
- Szabo, A.; Langer, T.; Schröder, H.; Flanagan, J.; Bukau, B.; Hartl, F.U. The ATP hydrolysis-dependent reaction cycle of the Escherichia coli Hsp70 system DnaK, DnaJ, and GrpE. Proc. Natl. Acad. Sci. USA 1994, 91, 10345–10349. [Google Scholar] [CrossRef]
- Conner, T.W.; LaFayette, P.R.; Nagao, R.T.; Key, J.L. Sequence and Expression of a HSP83 from Arabidopsis thaliana. Plant Physiol. 1990, 94, 1689–1695. [Google Scholar] [CrossRef][Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Total Read Bases | Total Processed Reads | GC (%) | QC20 (%) | QC30 (%) | Mapped Reads | Mapped Ratio (%) | |
|---|---|---|---|---|---|---|---|
| Optimal temperature (20 ℃) | 7,571,262,233 | 75,171,018 | 45.05 | 99.13 | 96.64 | 66,549,755 | 88.53 |
| High temperature (37 ℃) | 6,763,559,164 | 67,144,596 | 44.00 | 99.18 | 96.77 | 64,208,699 | 95.63 |
| Gene Name | Gene Symbol | Description | Fold Change (High Temperature) /Optimal Temperature) | High Temperature FPKM | Optimal Temperature FPKM |
|---|---|---|---|---|---|
| LsHSP70A | LOC111912265 | heat shock cognate 70 kDa protein 2-like | 2652.36 | 253.41 | 0.08 |
| LsHSP70B | LOC111909436 | heat shock cognate 70 kDa protein 2-like | 1842.68 | 346.31 | 0.16 |
| LsHSP17.3A | LOC111907241 | 17.3 kDa class I heat shock protein-like | 1430.81 | 314.21 | 0.18 |
| LsHSP17.9A | LOC111883736 | 17.9 kDa class II heat shock protein-like | 867.15 | 575.13 | 0.59 |
| LsHSP17.9B | LOC111883707 | 17.9 kDa class II heat shock protein-like | 804.25 | 118.73 | 0.11 |
| LsHSP70C | LOC111878184 | heat shock 70 kDa protein | 797 | 73.23 | 0.08 |
| LsSHSPA | LOC111903402 | small heat shock protein, chloroplastic | 722.28 | 44.25 | 0.05 |
| LsHSP83A | LOC111888460 | heat shock protein 83-like | 642.29 | 430.69 | 0.61 |
| LsHSP17.9C | LOC111883742 | 17.9 kDa class II heat shock protein-like | 599.83 | 382.16 | 0.55 |
| LsHSP17.9D | LOC111883705 | 17.9 kDa class II heat shock protein-like | 521.53 | 297.66 | 0.49 |
| LsHSP70D | LOC111909434 | heat shock cognate 70 kDa protein 2-like | 500.86 | 134.68 | 0.24 |
| LsHSP83B | LOC111876685 | heat shock protein 83 | 430.44 | 146.87 | 0.30 |
| LsHSP17.3B | LOC111880347 | 17.3 kDa class I heat shock protein-like | 388.74 | 329.52 | 0.75 |
| LsHSP17.8A | LOC111888905 | 17.8 kDa class I heat shock protein-like | 378 | 15.51 | 0.01 |
| LsHSP15.7 | LOC111904729 | 15.7 kDa heat shock protein, peroxisomal | 364.15 | 32.8 | 0.08 |
| LsHSP17.8B | LOC111888937 | 17.8 kDa class I heat shock protein-like | 346.51 | 14.04 | 0.01 |
| LSHSP17.9E | LOC111883704 | 17.9 kDa class II heat shock protein-like | 343.39 | 375.45 | 1.00 |
| LsSHSPB | LOC111892899 | small heat shock protein, chloroplastic | 331.61 | 306.43 | 0.82 |
| LsHSP70E | LOC111901569 | stromal 70 kDa heat shock-related protein, chloroplastic-like | 288.39 | 106.59 | 0.33 |
| LsHSP17.3C | LOC111907242 | 17.3 kDa class I heat shock protein | 212.52 | 617.27 | 2.65 |
| LsHSP17.5 | LOC111905717 | 17.5 kDa class I heat shock protein-like | 209.04 | 793.21 | 3.46 |
| LsHSP22 | LOC111891650 | 22.0 kDa class IV heat shock protein-like | 160.13 | 19.88 | 0.11 |
| LsHSP17.4 | LOC111910362 | 17.4 kDa class III heat shock protein | 157.84 | 91.11 | 0.51 |
| LsHSP18.1 | LOC111888907 | 18.1 kDa class I heat shock protein-like | 139.04 | 10.47 | 0.04 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kang, Y.; Jang, S.-W.; Lee, H.J.; Barchenger, D.W.; Jang, S. Expression Profiling of Heat Shock Protein Genes as Putative Early Heat-Responsive Members in Lettuce. Horticulturae 2021, 7, 312. https://doi.org/10.3390/horticulturae7090312
Kang Y, Jang S-W, Lee HJ, Barchenger DW, Jang S. Expression Profiling of Heat Shock Protein Genes as Putative Early Heat-Responsive Members in Lettuce. Horticulturae. 2021; 7(9):312. https://doi.org/10.3390/horticulturae7090312
Chicago/Turabian StyleKang, Yeeun, Suk-Woo Jang, Hee Ju Lee, Derek W. Barchenger, and Seonghoe Jang. 2021. "Expression Profiling of Heat Shock Protein Genes as Putative Early Heat-Responsive Members in Lettuce" Horticulturae 7, no. 9: 312. https://doi.org/10.3390/horticulturae7090312
APA StyleKang, Y., Jang, S.-W., Lee, H. J., Barchenger, D. W., & Jang, S. (2021). Expression Profiling of Heat Shock Protein Genes as Putative Early Heat-Responsive Members in Lettuce. Horticulturae, 7(9), 312. https://doi.org/10.3390/horticulturae7090312