
Exogenous Application of Chitosan Alleviate Salinity Stress in Lettuce (Lactuca sativa L.)

Abstract
:1. Introduction
2. Materials and Methods
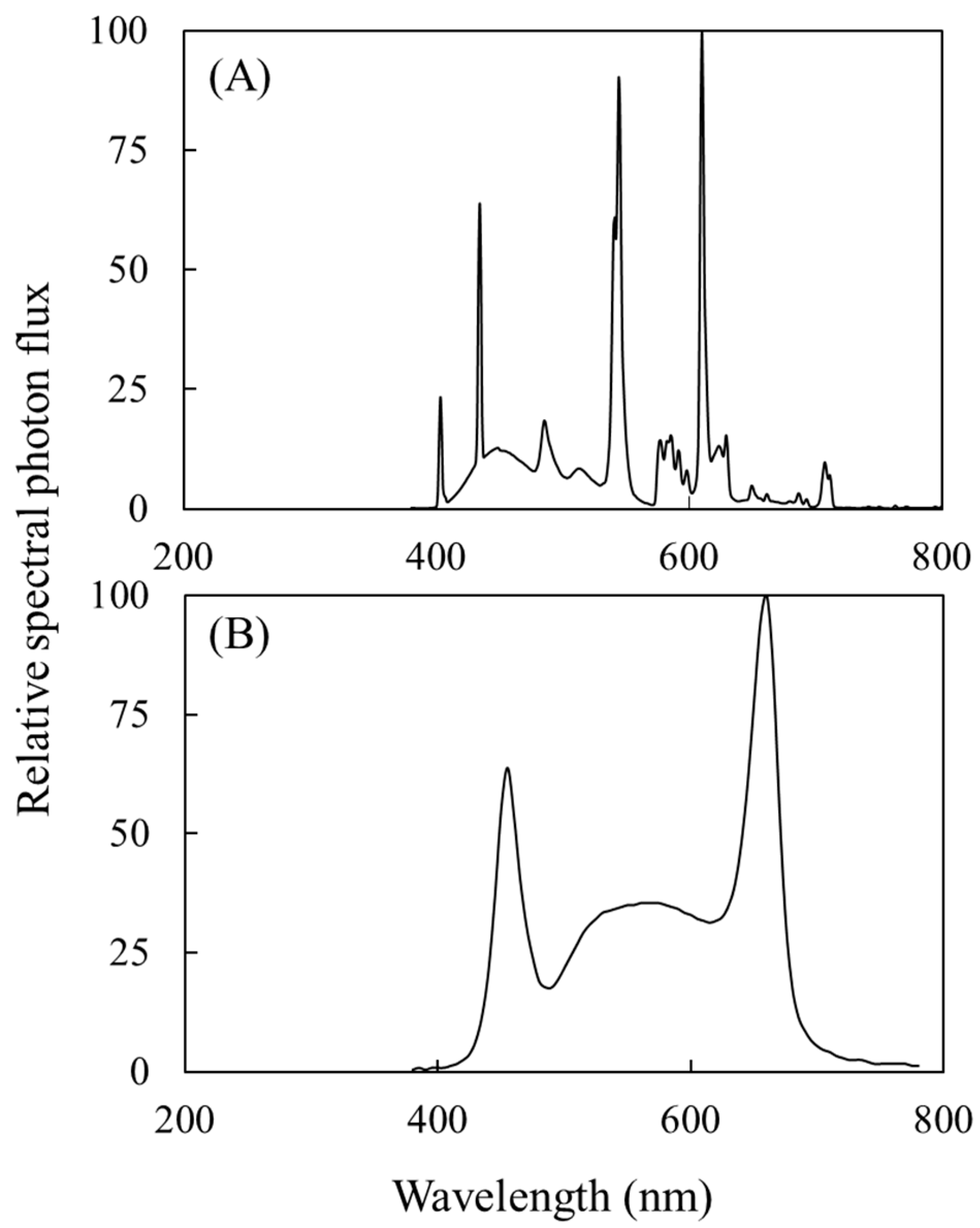
2.1. Plant Materials and Treatments
2.2. Plant Growth Analysis
2.3. Estimation of Leaf Relative Water Content and Electrolyte Leakage
2.4. Determination of the Potassium and Sodium Contents in Lettuce Leaves
2.5. Estimation of the Proline, Soluble Sugar, and Ascorbic Acid Contents
2.6. Examination of H2O2 Content, O2− Generation, and Malondialdehyde Content
2.7. Enzyme Assays
2.8. Statistical Analysis
3. Results
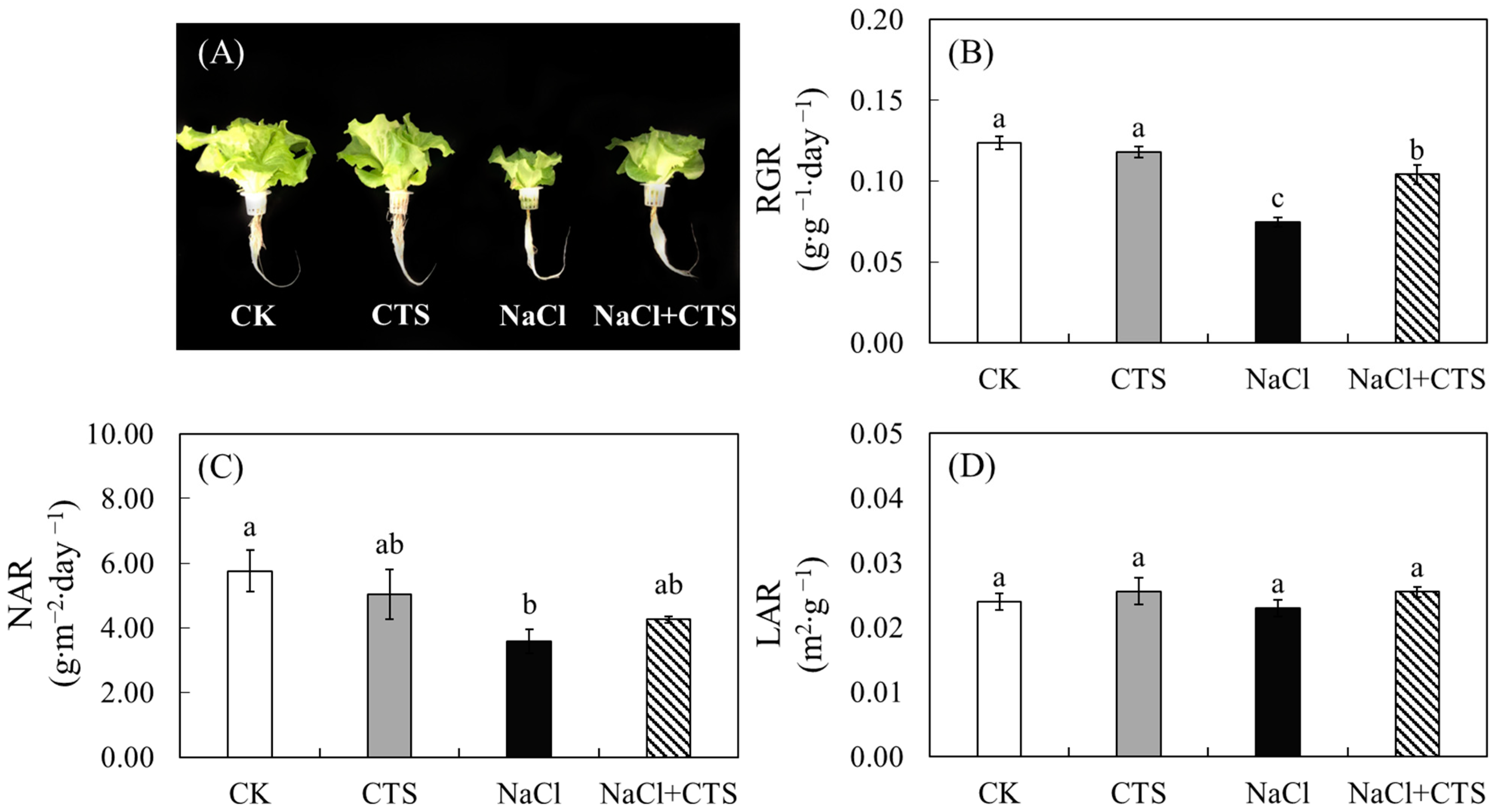
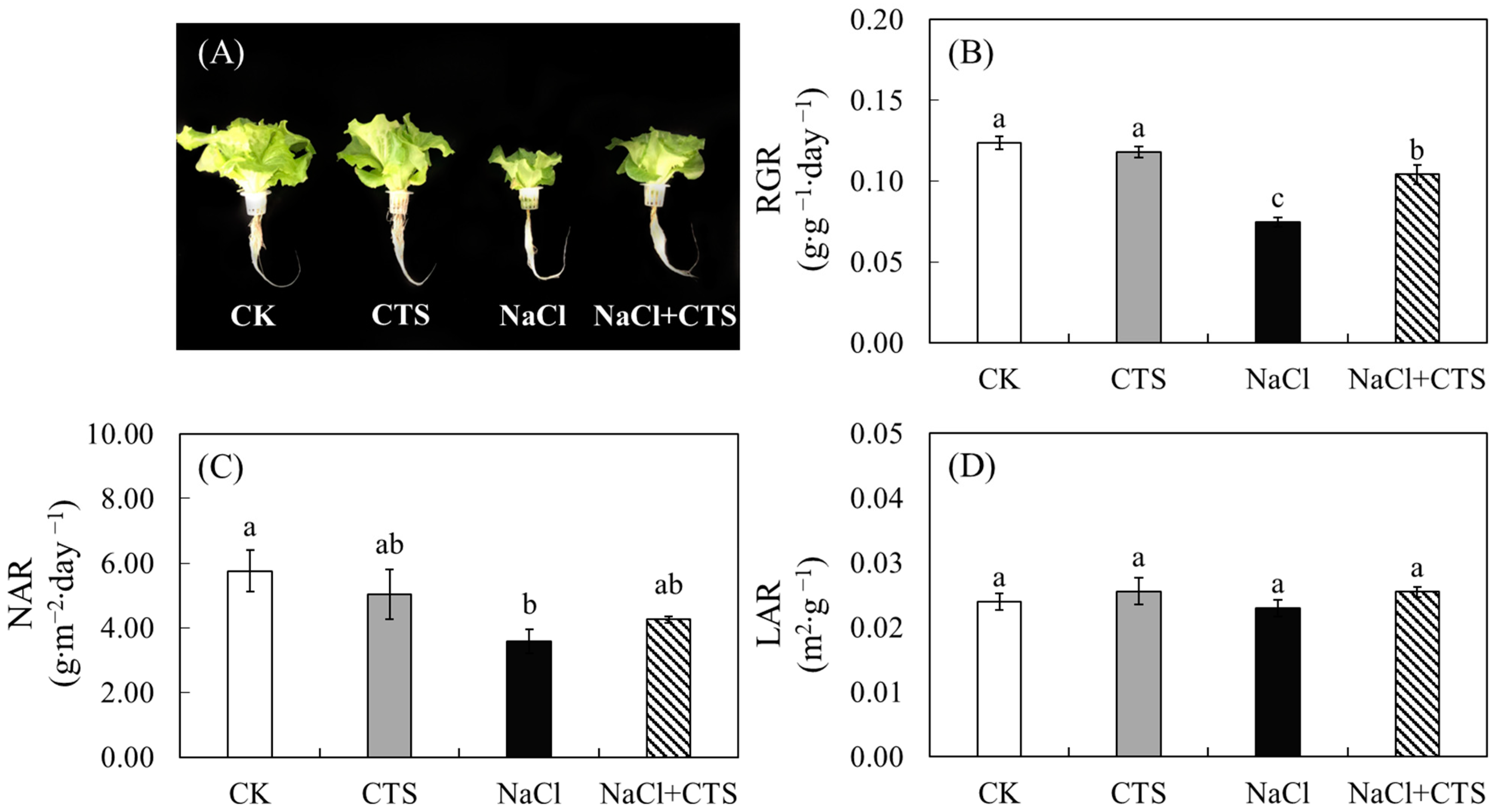
3.1. Exogenous CTS Improved the Growth and Biomass of Lettuce under NaCl Stress
3.2. Effects of NaCl and CTS on the RWC, EL, and the Contents of Potassium and Sodium of the Lettuce Leaves
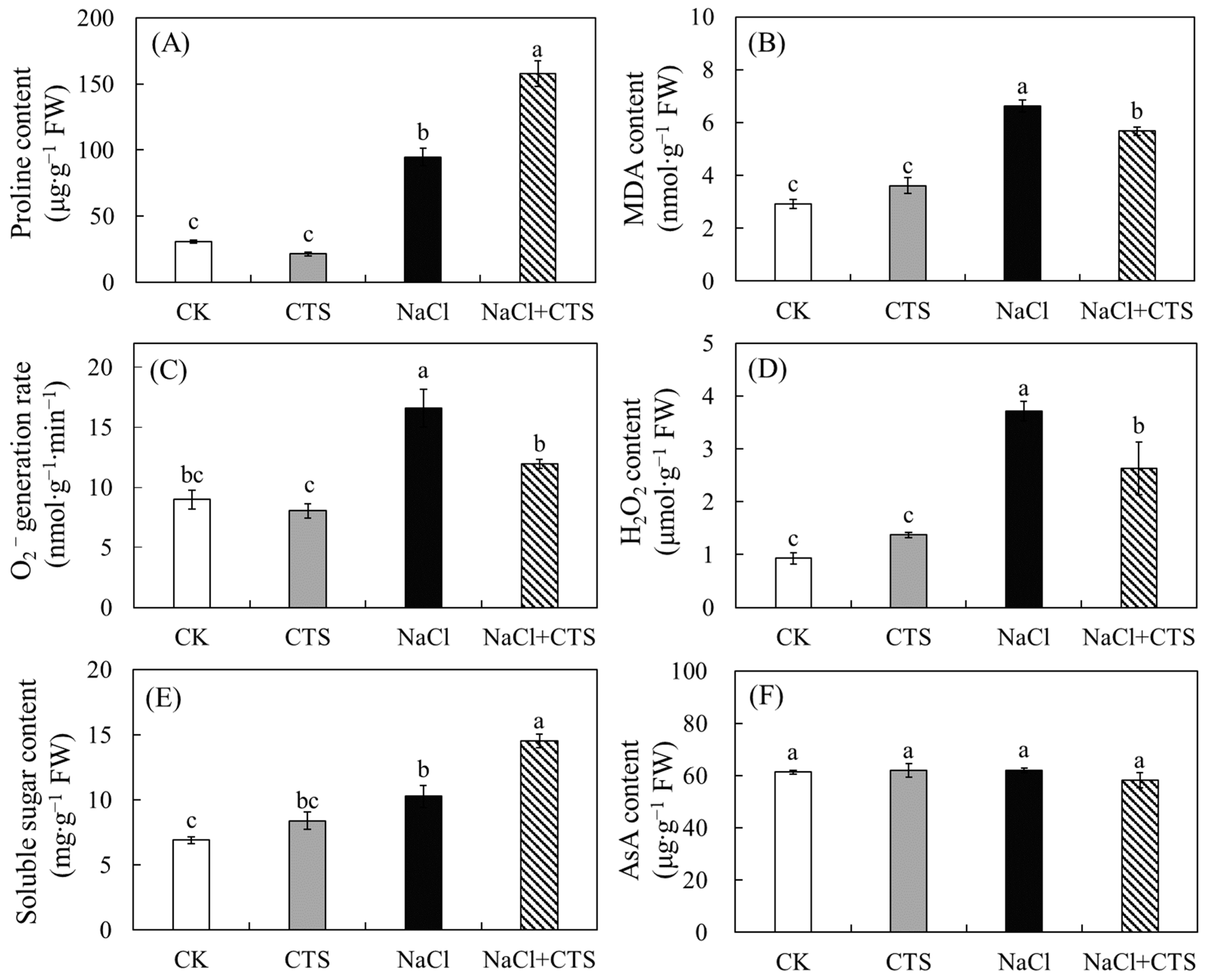
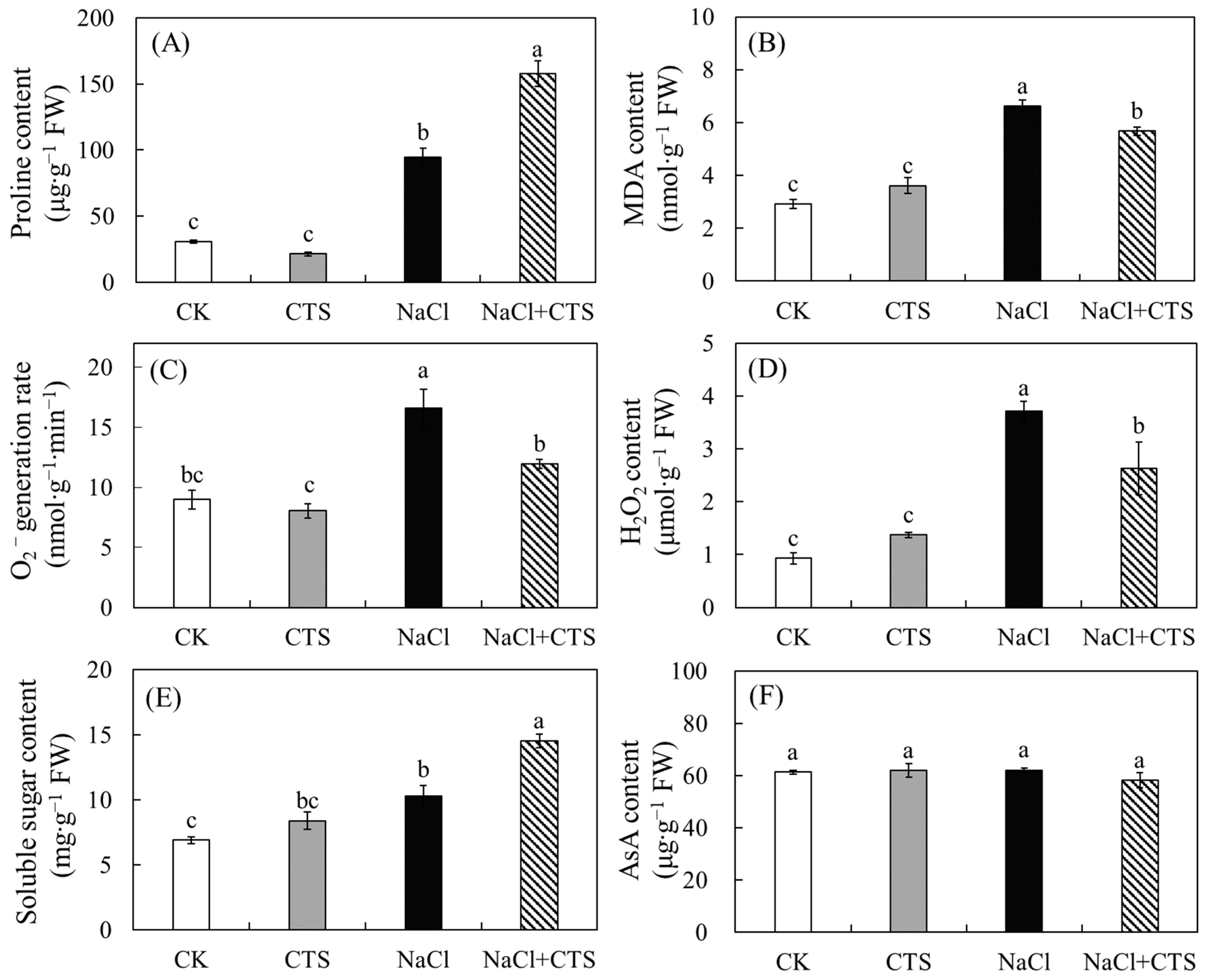
3.3. Effects of NaCl and CTS on the Proline Content, MDA Content, O2− Generation, H2O2 Content, Soluble Sugar Content, and AsA Content of the Lettuce Leaves
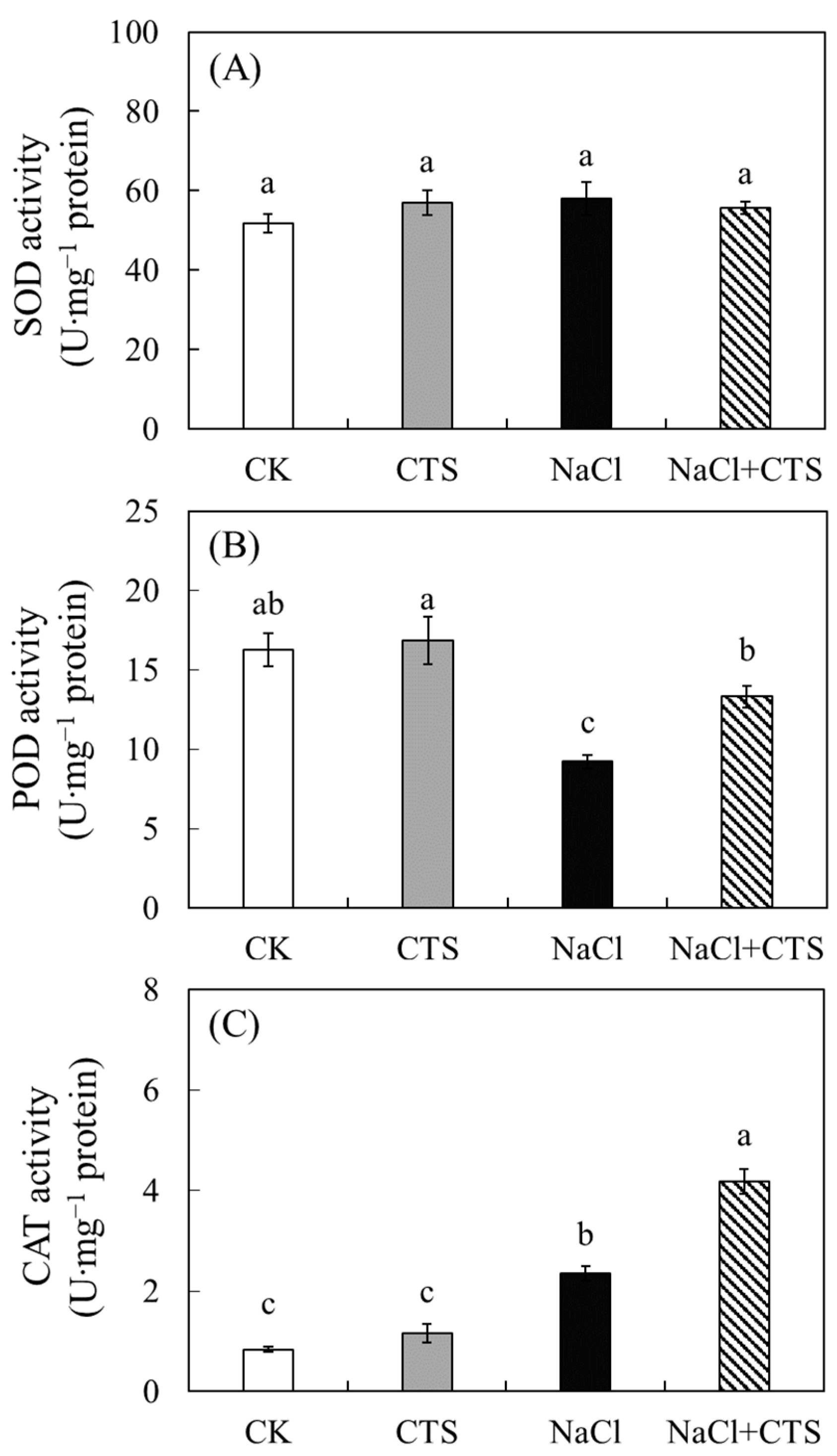
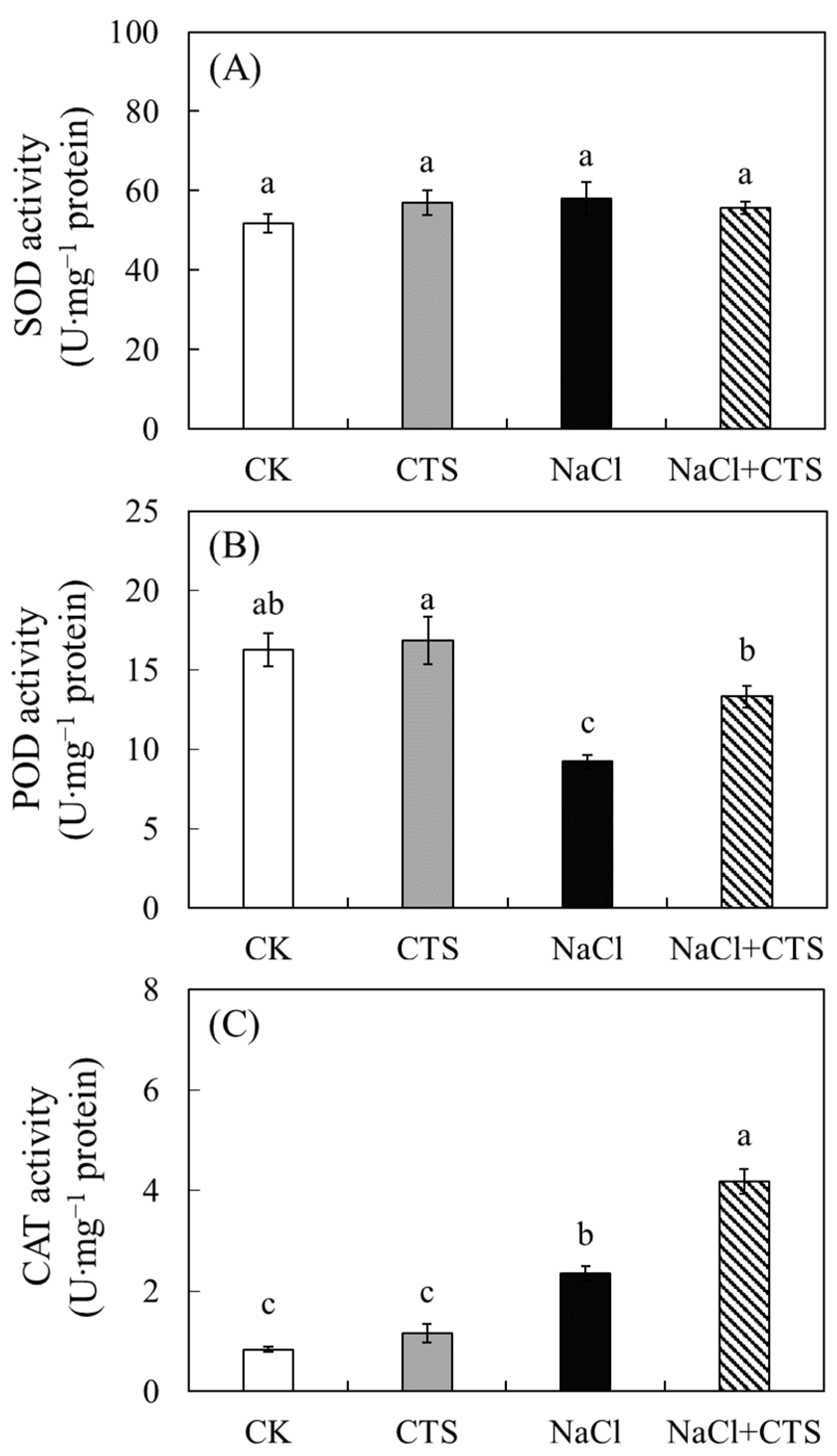
3.4. Effects of NaCl and CTS on the Antioxidant Enzyme Activity in the Lettuce Leaves
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Shahid, S.A.; Zaman, M.; Heng, L. Introduction to Soil Salinity, Sodicity and Diagnostics Techniques. In Guideline for Salinity Assessment, Mitigation and Adaptation Using Nuclear and Related Techniques; Springer Nature: Vienna, Switzerland, 2018. [Google Scholar]
- Deinlein, U.; Stephan, A.B.; Horie, T.; Luo, W.; Schroeder, J.I. Plant salt-tolerance mechanisms. Trends Plant Sci. 2014, 19, 371–379. [Google Scholar] [CrossRef] [Green Version]
- Ke, Q.; Wang, Z.; Ji, C.Y.; Jeong, J.C.; Lee, H.S.; Li, H.; Xu, B.; Deng, X.; Kwak, S.S. Transgenic poplar expressing codA exhibits enhanced growth and abiotic stress tolerance. Plant Physiol. Biochem. 2016, 100, 75–84. [Google Scholar] [CrossRef]
- Chen, M.; Yang, Z.; Liu, J.; Zhu, T.; Wei, X.; Fan, H.; Wang, B. Adaptation mechanism of salt excluders under saline conditions and its applications. Int. J. Mol. Sci. 2018, 19, 3668. [Google Scholar] [CrossRef] [Green Version]
- Azarmi-Atajan, F.; Sayyari-Zohan, M.H. Alleviation of salt stress in lettuce (Lactuca sativa L.) by plant growth-promoting rhizobacteria. J. Hortic. Postharvest Res. 2020, 3, 67–78. [Google Scholar]
- Sohaid, M.; Zahir, Z.; Khan, M.; Ans, M.; Asghar, H.; Yasin, S.; Al-Barakah, F. Comparative evaluation of different carrier-based multi-strain bacterial formulations to mitigate the salt stress in wheat. Saudi J. Biol. Sci. 2020, 27, 777–787. [Google Scholar]
- Zhu, J.K. Regulation of ion homeostasis under salt stress. Curr. Opin. Plant Biol. 2003, 6, 441–445. [Google Scholar] [CrossRef]
- Yang, Y.; Guo, Y. Unraveling salt stress signaling in plants. J. Integr. Plant Biol. 2018, 60, 58–66. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hasegawa, P.M. Plant cellular and molecular responses to high salinity. Annu. Rev. Plant Physiol. Plant Mol. Biol. 2000, 51, 463–499. [Google Scholar] [CrossRef] [Green Version]
- Wu, G.Q.; Shui, Q.Z.; Wang, C.M.; Zhang, J.L.; Yuan, H.J.; Li, S.J.; Liu, Z.J. Characteristics of Na+ uptake in sugar beet (Beta vulgaris L.) seedlings under mild salt conditions. Acta Physiol. Plant. 2015, 37, 70. [Google Scholar] [CrossRef]
- Zhang, J.L.; Wetson, A.M.; Wang, S.M.; Gurmani, A.R.; Bao, A.K.; Wang, C.M. Factors associated with determination of root 22Na+ influx in the salt accumulation halophyte Suaeda maritima. Biol. Trace Elem. Res. 2011, 139, 108–117. [Google Scholar] [CrossRef] [PubMed]
- Assaha, D.V.M.; Ueda, A.; Saneoka, H.; Al-Yahyai, R.; Yaish, M.W. The role of Na+ and K+ transporters in salt stress adaptation in glycophytes. Front. Physiol. 2017, 8, 509. [Google Scholar] [CrossRef] [PubMed]
- Isayenkov, S.V.; Maathuis, F.J.M. Plant salinity stress: Many unanswered questions remain. Front. Plant Sci. 2019, 10, 80. [Google Scholar] [CrossRef] [Green Version]
- Lv, X.; Chen, S.; Wang, Y. Advances in understanding the physiological and molecular responses of sugar beet to salt stress. Front. Plant Sci. 2019, 10, 1431. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.; Liu, J.; Wang, G.; Cha, J.Y.; Li, G.; Chen, S.; Li, Z.; Guo, J.; Zhang, C.; Yang, Y.; et al. A chaperone function of NO CATALASE ACTIVITY1 is required to maintain catalase activity and for multiple stress responses in Arabidopsis. Plant Cell 2015, 27, 908–925. [Google Scholar] [CrossRef] [Green Version]
- Del, L.A.D.; López-Huertas, E. ROS generation in peroxisomes and its role in cell signaling. Plant Cell Physiol. 2016, 57, 1364–1376. [Google Scholar] [CrossRef]
- Zhao, J.; Pan, L.; Zhou, M.; Yang, Z.; Meng, Y.; Zhang, X. Comparative physiological and transcriptomic analyses reveal mechanisms of improved osmotic stress tolerance in annual ryegrass by exogenous chitosan. Genes 2019, 10, 853. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Betchem, G.; Johnson, N.; Wang, Y. The application of chitosan in the control of post-harvest diseases: A review. J. Plant Dis. Protect. 2019, 126, 495–507. [Google Scholar] [CrossRef]
- Pongprayoon, W.; Roytrakul, S.; Pichayangkura, R.; Chadchawan, S. The role of hydrogen peroxide in chitosan-induced resistance to osmotic stress in rice (Oryza sativa L.). Plant Growth Regul. 2013, 70, 159–173. [Google Scholar] [CrossRef]
- Morin-Crini, N.; Lichtfouse, E.; Torri, G.; Crini, G. Applications of chitosan in food, pharmaceuticals, medicine, cosmetics, agriculture, textiles, pulp and paper, biotechnology, and environmental chemistry. Environ. Chem. Lett. 2019, 17, 1667–1692. [Google Scholar] [CrossRef] [Green Version]
- Malerba, M.; Cerana, R. Chitosan effects on plant systems. Int. J. Mol. Sci. 2016, 17, 996. [Google Scholar] [CrossRef]
- Guan, Y.J.; Hu, J.; Wang, X.J.; Shao, C.X. Seed priming with chitosan improves maize germination and seedling growth in relation to physiological changes under low temperature stress. J. Zhejiang Univ. Sci. B 2009, 10, 427–433. [Google Scholar] [CrossRef] [Green Version]
- Jabeen, N.; Ahmad, R. The activity of antioxidant enzymes in response to salt stress in safflower (Carthamus tinctorius L.) and sunflower (Helianthus annuus L.) seedlings raised from seed treated with chitosan. J. Sci. Food Agric. 2012, 93, 1699–1705. [Google Scholar] [CrossRef]
- Geng, W.; Li, Z.; Hassan, M.; Peng, Y. Chitosan regulates metabolic balance, polyamine accumulation, and Na+ transport contributing to salt tolerance in creeping bentgrass. BMC Plant Biol. 2020, 20, 506. [Google Scholar] [CrossRef] [PubMed]
- Cho, M.H.; No, H.K.; Prinyawiwatkul, W. Chitosan treatments affect growth and selected quality of sunflower sprouts. J. Food Sci. 2010, 73, S70–S77. [Google Scholar] [CrossRef] [PubMed]
- Bittelli, M.; Flury, M.; Campbell, G.S.; Nichols, E.J. Reduction of transpiration through foliar application of chitosan. Agric. For. Meteorol. 2001, 107, 167–175. [Google Scholar] [CrossRef]
- Dzung, N.A. Chitosan and their derivatives as prospective biosubstances for developing sustainable eco-agriculture. Adv. Chitin Sci. 2007, 10, 453–459. [Google Scholar]
- Dzung, N.A.; Thang, N.T. Effect of oligoglucosamine on the growth and development of peanut (Arachis hypogea L.). In Proceedings of the 6th Asia-Pacific on Chitin, Chitosan Symposium, Singapore, 23–26 May 2004; Khor, E., Hutmacher, D., Yong, L.L., Eds.; ISBN 981-05r-r0904-9. [Google Scholar]
- Li, Z.; Zhang, Y.; Zhang, X.; Merewitz, E.; Peng, Y.; Ma, X.; Huang, L.; Yan, Y. Metabolic pathways regulated by chitosan contributing to drought resistance in white clover. J. Proteome Res. 2017, 16, 3039–3052. [Google Scholar] [CrossRef]
- Yang, F.; Hu, J.; Li, J.; Wu, X.; Qian, Y. Chitosan enhances leaf membrane stability and antioxidant enzyme activities in apple seedlings under drought stress. Plant Growth Regul. 2009, 58, 131–136. [Google Scholar] [CrossRef]
- Hadwiger, L.A. Anatomy of a nonhost disease resistance response of pea to Fusarium solani: PR gene elicitation via DNase, chitosan and chromatin alterations. Front. Plant Sci. 2015, 6, 373. [Google Scholar] [CrossRef]
- Aleksandrowicz-Trzcińska, M.; Bogusiewicz, A.; Szkop, M.; Drozdowski, S. Effect of chitosan on disease control and growth of scots pine (Pinus sylvestris L.) in a forest nursery. Forests 2015, 6, 3165–3176. [Google Scholar] [CrossRef] [Green Version]
- Viacava, G.E.; Gonzalez-Aguilar, G.G.; Roura, S.I. Determination of phytochemicals and antioxidant activity in butterhead lettuce related to leaf age and position. J. Food Biochem. 2014, 38, 352–362. [Google Scholar] [CrossRef]
- Aksakal, O.; Tabay, D.; Esringu, A.; Icoglu, A.F.; Esim, N. Effect of proline on biochemical and molecular mechanisms in lettuce (Lactuca sativa L.) exposed to UV-B radiation. Photochem. Photobiol. Sci. 2017, 16, 246–254. [Google Scholar] [CrossRef] [PubMed]
- Yildirim, E.; Ekinci, M.; Turan, M.; Dursun, A.; Kul, R.; Parlakova, F. Roles of glycine betaine in mitigating deleterious effect of salt stress on lettuce (Lactuca sativa L.). Arch. Agron. Soil Sci. 2015, 61, 1673–1689. [Google Scholar] [CrossRef]
- Fernández, J.A.; Niirola, D.; Ochoa, J.; Orsini, F.; Pennisi, G.; Gianquinto, G.; Egea-Gilabert, C. Root adaptation and ion selectivity affects the nutritional value of salt-stressed hydroponically grown baby-leaf Nasturtium officinale and Lactuca sativa. Agr. Food Sci. 2016, 25, 230–239. [Google Scholar] [CrossRef]
- Ahmed, S.; Ahmed, S.; Roy, S.K.; Woo, S.H.; Sonawane, K.D.; Shohael, A.M. Effect of salinity on the morphological, physiological and biochemical properties of lettuce (Lactuca sativa L.) in Bangladesh. Open Agric. 2019, 4, 361–373. [Google Scholar] [CrossRef]
- Garrido, Y.; Tudela, J.A.; Marín, A.; Mestre, T.; Martínez, V.; Gil, M.I. Physiological, phytochemical and structural changes of multi-leaf lettuce caused by salt stress. J. Sci. Food Agric. 2014, 94, 1592–1599. [Google Scholar] [CrossRef] [PubMed]
- Joshi, J.; Zhang, G.; Shen, S.; Supaibulwatana, K.; Watanabe, C.K.A.; Yamori, W. A combination of downward lighting and supplemental upward lighting improves plant growth in a closed plant factory with artificial lighting. HortScience 2017, 52, 831–835. [Google Scholar] [CrossRef] [Green Version]
- Porra, R.J.; Thompson, W.A.; Kriedemann, P.E. Determination of accurate extinction coefficients and simultaneous equations for assaying chlorophylls a and b extracted with four different solvents: Verification of the concentration of chlorophyll standards by atomic absorption spectroscopy. BBA Bioenerg. 1989, 975, 384–394. [Google Scholar] [CrossRef]
- Ohtake, N.; Ishikura, M.; Suzuki, H.; Yamori, W.; Goto, E. Continuous irradiation with alternating red and blue light enhances plant growth while keeping nutritional quality in lettuce. HortScience 2018, 53, 1804–1809. [Google Scholar] [CrossRef] [Green Version]
- Yamasaki, S.; Dillenburg, L.R. Measurements of leaf relative water content in Araucaria angustifolia. Rev. Bras. Fisiol. Veg. 1999, 11, 69–75. [Google Scholar]
- Ahmad, P.; Latef, A.A.A.; Hashem, A.; Abd_Allah, E.F.; Gucel, S.; Tran, L.P. Nitric oxide mitigates salt stress by regulating levels of osmolytes and antioxidant enzymes in chickpea. Front. Plant Sci. 2016, 7, 347. [Google Scholar] [CrossRef] [Green Version]
- Zhang, G.; Yan, Z.; Wang, Y.; Feng, Y.; Yuan, Q. Exogenous proline improve the growth and yield of lettuce with low potassium content. Sci. Hortic. 2020, 271, 109469. [Google Scholar] [CrossRef]
- Bates, L.S.; Waldren, R.P.; Teare, I.D. Rapid determination of free proline for water-stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Yemm, E.W.; Willis, A.J. The estimation of carbohydrates in plant extracts by anthrone. Biochem. J. 1954, 57, 508–514. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kampfenkel, K.; Van Montagu, M.; Inzé, D. Extraction and determination of ascorbate and dehydroascorbate from plant tissue. Anal. Biochem. 1995, 225, 165–167. [Google Scholar] [CrossRef]
- Patterson, B.D.; MacRae, E.A.; Ferguson, I.B. Estimation of hydrogen peroxide in plant extracts using titanium (IV). Anal. Biochem. 1984, 139, 487–492. [Google Scholar] [CrossRef]
- Averina, N.G.; Nedved, E.L.; Shcherbakov, R.A.; Vershilovskaya, I.V.; Yaronskaya, E.B. Role of 5-aminolevulinic acid in the formation of winter rape resistance to sulfonylurea herbicides. Russ. J. Plant Physiol. 2014, 61, 679–687. [Google Scholar] [CrossRef]
- Heath, R.L.; Packer, L. Photoperoxidation in isolated chloroplasts: I. Kinetics and stoichiometry of fatty acid peroxidation. Arch. Biophys. Biophys. 1968, 125, 189–198. [Google Scholar] [CrossRef]
- Jin, P.; Wang, H.; Zhang, Y.; Huang, Y.; Wang, L.; Zheng, Y. UV-C enhances resistance against gray mold decay caused by Botrytis cinerea in strawberry fruit. Sci. Hortic. 2017, 225, 106–111. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principaldye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Latef, A.A.H.A.; Chaoxing, H. Effect of arbuscular mycorrhizal fungi on growth, mineral nutrition, antioxidant enzymes activity and fruit yield of tomato grown under salinity stress. Sci. Hortic. 2011, 127, 228–233. [Google Scholar] [CrossRef]
- Latef, A.A.H.A.; Chaoxing, H. Does the inoculation with Glomus mosseae improve salt tolerance in pepper plants? J. Plant Growth Regul. 2014, 33, 644–653. [Google Scholar] [CrossRef]
- Hemantaranjan, A.; Deepmala, K.; Bharti, S.; Nishant, B.A. A future perspective in crop protection: Chitosan and its oligosaccharides. Adv. Plants Agric. Res. 2014, 1, 00006. [Google Scholar] [CrossRef] [Green Version]
- Mahdavi, B.; Rahimi, A. Seed priming with chitosan improves the germination and growth performance of ajowan (Carum copticum) under salt stress. Eurasia. J. Biosci. 2013, 7, 69–76. [Google Scholar] [CrossRef]
- Turk, H. Chitosan-induced enhanced expression and activation of alternative oxidase confer tolerance to salt stress in maize seedlings. Plant Physiol. Bioch. 2019, 141, 415–422. [Google Scholar] [CrossRef]
- Zou, P.; Li, K.C.; Liu, S.; Xing, R.G.; Qin, Y.K.; Yu, H.H.; Zhou, M.M.; Li, P.C. Effect of chitooligosaccharides with different degrees of acetylation on wheat seedlings under salt stress. Carbohydr. Polym. 2015, 126, 62–69. [Google Scholar] [CrossRef]
- Hoffmann, W.A.; Poorter, H. Avoiding bias in calculations of relative growth rate. Ann. Bot. 2002, 80, 37–42. [Google Scholar] [CrossRef] [Green Version]
- Rady, M.M.; El-Shewy, A.A.; El-Yazal, M.A.; Abdelaal, K.E.S. Response of salt-stressed common bean plant performances to foliar application of phosphorus (MAP). Int. Lett. Nat. Sci. 2018, 72, 7–20. [Google Scholar] [CrossRef] [Green Version]
- Qi, H.; Wu, Z.; Zhang, L.; Li, J.; Zhou, J.; Jun, Z.; Zhu, B. Monitoring of peanut leaves chlorophyll content based on drone-based multispectral image feature extraction. Comput. Electron. Agr. 2021, 187, 106292. [Google Scholar] [CrossRef]
- Zou, P.; Lu, X.; Jing, C.; Yuan, Y.; Lu, Y.; Zhang, C.; Meng, L.; Zhao, H.; Li, Y. Low-molecular-weightt polysaccharides from Pyropia yezoensis enhance tolerance of wheat seedlings (Triticum aestivum L.) to Salt Stress. Front. Plant Sci. 2018, 9, 427. [Google Scholar] [CrossRef] [Green Version]
- Sen, S.; Chouhan, D.; Das, D.; Ghosh, R.; Mandal, P. Improvisation of salinity stress response in mung bean through solid matrix priming with normal and nano-sized chitosan. Int. J. Biol. Macromol. 2020, 145, 108–123. [Google Scholar] [CrossRef] [PubMed]
- Rasool, S.; Ahmad, A.; Siddiqi, T.O.; Ahmad, P. Changes in growth, lipid peroxidation and some key antioxidant enzymes in chickpea genotypes under salt stress. Acta Physiol. Plant. 2013, 35, 1039–1050. [Google Scholar] [CrossRef]
- Negrão, S.; Schmöckel, S.M.; Tester, M. Evaluating physiological responses of plants to salinity stress. Ann. Bot. 2017, 119, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eckardt, N.A. A new chlorophyll degradation pathway. Plant Cell 2009, 21, 700. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suriya-arunroj, D.; Supapoj, N.; Toojinda, T.; Vanavichit, A. Relative leaf water content as an efficient method for evaluating rice cultivars for tolerance to salt stress. ScienceAsia 2004, 30, 411–415. [Google Scholar] [CrossRef]
- Mekawy, A.M.; Assaha, D.V.; Yahagi, H.; Tada, Y.; Ueda, A.; Saneoka, H. Growth, physiological adaptation, and gene expression analysis of two Egyptian rice cultivars under salt stress. Plant Physiol. Biochem. 2015, 87, 17–25. [Google Scholar] [CrossRef]
- Cuin, T.A.; Betts, S.A.; Chalmandrier, R.; Shabala, S. A root’s ability to retain K+ correlates with salt tolerance in wheat. J. Exp. Bot. 2008, 59, 2697–2706. [Google Scholar] [CrossRef] [Green Version]
- Rocha, I.M.A.; Vitorello, V.A.; Silva, J.S.; Ferreira-Silva, S.L.; Viegas, R.A.; Silva, E.N.; Silveira, J.A.G. Exogenous ornithine is an effective precursor and the delta-ornithine amino transferase pathway contributes to proline accumulation under high N recycling in salt-stressed cashew leaves. J. Plant Physiol. 2012, 169, 41–49. [Google Scholar] [CrossRef]
- Zhang, X.Q.; Li, K.C.; Xing, R.G.; Liu, S.; Li, P.C. Metabolite profiling of wheat seedlings induced by chitosan: Revelation of the enhanced carbon and nitrogen metabolism. Front. Plant Sci. 2017, 8, 13. [Google Scholar] [CrossRef] [Green Version]
- Gupta, A.K.; Kaur, N. Sugar signalling and gene expression in relation to carbohydrate metabolism under abiotic stresses in plants. J. Biosci. 2005, 30, 761–776. [Google Scholar] [CrossRef]
- Bolouri-Moghaddam, M.R.; Le, R.K.; Xiang, L.; Rolland, F.; Van den Ende, W. Sugar signalling and antioxidant network connections in plant cells. FEBS J. 2010, 277, 2022–2037. [Google Scholar] [CrossRef]
- Hayat, S.; Yadav, S.; Wani, A.S.; Irfan, M.; Alyemini, M.N.; Ahmad, A. Impact of sodium nitroprusside on nitrate reductase, proline and antioxidant systemin Solanum lycopersicum under salinity stress. Hort. Environ. Biotechnol. 2012, 53, 362–367. [Google Scholar] [CrossRef]
- Butt, M.; Ayyub, C.M.; Amjad, M.; Ah-mad, R. Proline application enhances growth of chilli by improving physiological and biochemical attributes under salt stress. Pak. J. Agr. Sci. 2016, 53, 43–49. [Google Scholar]
- Miller, G.; Suzuki, N.; Ciftci-Yilmaz, S.; Mittler, R. Reactive oxygen species homeostasis and signalling during drought and salinity stresses. Plant Cell Environ. 2010, 33, 453–467. [Google Scholar] [CrossRef] [PubMed]
- Li, J.T.; Qiu, Z.B.; Zhang, X.W.; Wang, L.S. Exogenous hydrogen peroxide can enhance tolerance of wheat seedlings to salt stress. Acta Physiol. Plant. 2011, 33, 835–842. [Google Scholar] [CrossRef]
- Wu, G.Q. Exogenous application of proline alleviates salt-induced toxicity in sainfoin seedlings. J. Anim. Plant Sci. 2017, 27, 246–251. [Google Scholar]
- Asada, K. Ascorbate peroxidase—A hydrogen peroxide-scavenging enzyme in plants. Physiol. Plant. 1992, 85, 235–241. [Google Scholar] [CrossRef]
- Alscher, R.G.; Donahue, J.L.; Cramer, C.L. Reactive oxygen species and antioxidants: Relationships in green cells. Physiol. Plant. 1997, 100, 224–233. [Google Scholar] [CrossRef]
- Orsini, F.; Pennisi, G.; Mancarella, S.; Nayef, M.A.; Sanoubar, R.; Nicola, S.; Gianquinto, G. Hydroponic lettuce yields are improved under salt stress by utilizing white plastic film and exogenous applications of proline. Sci. Hortic. 2018, 233, 283–293. [Google Scholar] [CrossRef]
- Apel, K.; Hirt, H. Reactive oxygen species: Metabolism, oxidative stress, and signal transduction. Annu. Rev. Plant Biol. 2004, 55, 373–399. [Google Scholar] [CrossRef] [Green Version]
- Foyer, C.H.; Noctor, G. Oxidant and antioxidant signalling in plants: A re-evaluation of the concept of oxidative stress in a physiological context. Plant Cell Environ. 2005, 28, 1056–1071. [Google Scholar] [CrossRef]
- Kusvuran, S.; Bayat, R.; Ustun, A.S.; Ellialtioglu, S.S. Exogenous proline improves osmoregulation, physiological and biochemical responses of eggplant under salt stress. Fresenius Environ. Bull. 2020, 29, 152–161. [Google Scholar]
- Thomas, C.E.; Mclean, L.R.; Parkar, R.A.; Ohleweiler, D.F. Ascorbate and phenolic antioxidant interactions in prevention of liposomal oxidation. Lipids 1992, 27, 543–550. [Google Scholar] [CrossRef]
- Iqbal, N.; Umar, S.; Khan, N.A.; Khan, M.I.R. A new perspective of phytohormones in salinity tolerance: Regulation of proline metabolism. Environ. Exp. Bot. 2014, 100, 34–42. [Google Scholar] [CrossRef]
- Piotr, S.; Monika, G.; Edward, M.; Marcin, S. Oligo-alginate with low molecular mass improves growth and physiological activity of eucomis autumnalis under salinity stress. Molecules 2018, 23, 812. [Google Scholar]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatments | Total Leaf Area (m2) | Shoot FW (g∙plant−1) | Shoot DW (g∙plant−1) | Root FW (g∙plant−1) | Root DW (g∙plant−1) | Chl a (μg∙ml−1) | Chl b (μg∙ml−1) | Total Chl (μg∙ml−1) |
|---|---|---|---|---|---|---|---|---|
| CK | 0.052 ± 0.004 a | 48.8 ± 0.9 a | 2.39 ± 0.21 a | 8.73 ± 0.63 a | 0.31 ± 0.02 a | 8.68 ± 0.15 b | 3.30 ± 0.23 a | 11.98 ± 0.13 a |
| CTS | 0.054 ± 0.010 a | 45.2 ± 1.9 a | 2.11 ± 0.19 ab | 9.80 ± 1.23 a | 0.28 ± 0.02 ab | 9.24 ± 0.14 a | 3.29 ± 0.11 a | 12.53 ± 0.24 a |
| NaCl | 0.017 ± 0.003 b | 12.8 ± 1.7 c | 0.80 ± 0.05 c | 3.47 ± 0.51 b | 0.15 ± 0.01 c | 7.43 ± 0.14 c | 2.62 ± 0.04 b | 10.05 ± 0.16 c |
| NaCl + CTS | 0.041 ± 0.007 a | 29.1 ± 4.5 b | 1.56 ± 0.19 b | 5.98 ± 0.47 b | 0.24 ± 0.03 b | 8.18 ± 0.20 b | 2.73 ± 0.15 b | 10.91 ± 0.35 b |
| Treatments | RWC (%) | EL (%) | Potassium (mg∙g−1 DW) | Sodium (mg∙g−1 DW) | K+/Na+ Ratio |
|---|---|---|---|---|---|
| CK | 74.8 ± 1.3 ab | 16.1 ± 3.0 c | 73.96 ± 1.30 a | 1.09 ± 0.04 c | 68.23 ± 3.73 a |
| CTS | 79.0 ± 2.9 a | 15.5 ± 2.5 c | 72.02 ± 2.25 a | 1.05 ± 0.07 c | 69.38 ± 6.97 a |
| NaCl | 62.9 ± 1.6 c | 42.0 ± 3.1 a | 52.92 ± 1.55 c | 22.61 ± 1.31 a | 2.35 ± 0.07 b |
| NaCl + CTS | 72.8 ± 0.4 b | 33.1 ± 0.5 b | 61.27 ± 1.23 b | 12.21 ± 1.36 b | 5.11 ± 0.42 b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, G.; Wang, Y.; Wu, K.; Zhang, Q.; Feng, Y.; Miao, Y.; Yan, Z. Exogenous Application of Chitosan Alleviate Salinity Stress in Lettuce (Lactuca sativa L.). Horticulturae 2021, 7, 342. https://doi.org/10.3390/horticulturae7100342
Zhang G, Wang Y, Wu K, Zhang Q, Feng Y, Miao Y, Yan Z. Exogenous Application of Chitosan Alleviate Salinity Stress in Lettuce (Lactuca sativa L.). Horticulturae. 2021; 7(10):342. https://doi.org/10.3390/horticulturae7100342
Chicago/Turabian StyleZhang, Geng, Yuanhua Wang, Kai Wu, Qing Zhang, Yingna Feng, Yu Miao, and Zhiming Yan. 2021. "Exogenous Application of Chitosan Alleviate Salinity Stress in Lettuce (Lactuca sativa L.)" Horticulturae 7, no. 10: 342. https://doi.org/10.3390/horticulturae7100342
APA StyleZhang, G., Wang, Y., Wu, K., Zhang, Q., Feng, Y., Miao, Y., & Yan, Z. (2021). Exogenous Application of Chitosan Alleviate Salinity Stress in Lettuce (Lactuca sativa L.). Horticulturae, 7(10), 342. https://doi.org/10.3390/horticulturae7100342