
Effects of Short-Term Exposure to Low Temperatures on Proline, Pigments, and Phytochemicals Level in Kale (Brassica oleracea var. acephala)





Abstract
:1. Introduction
2. Materials and Methods
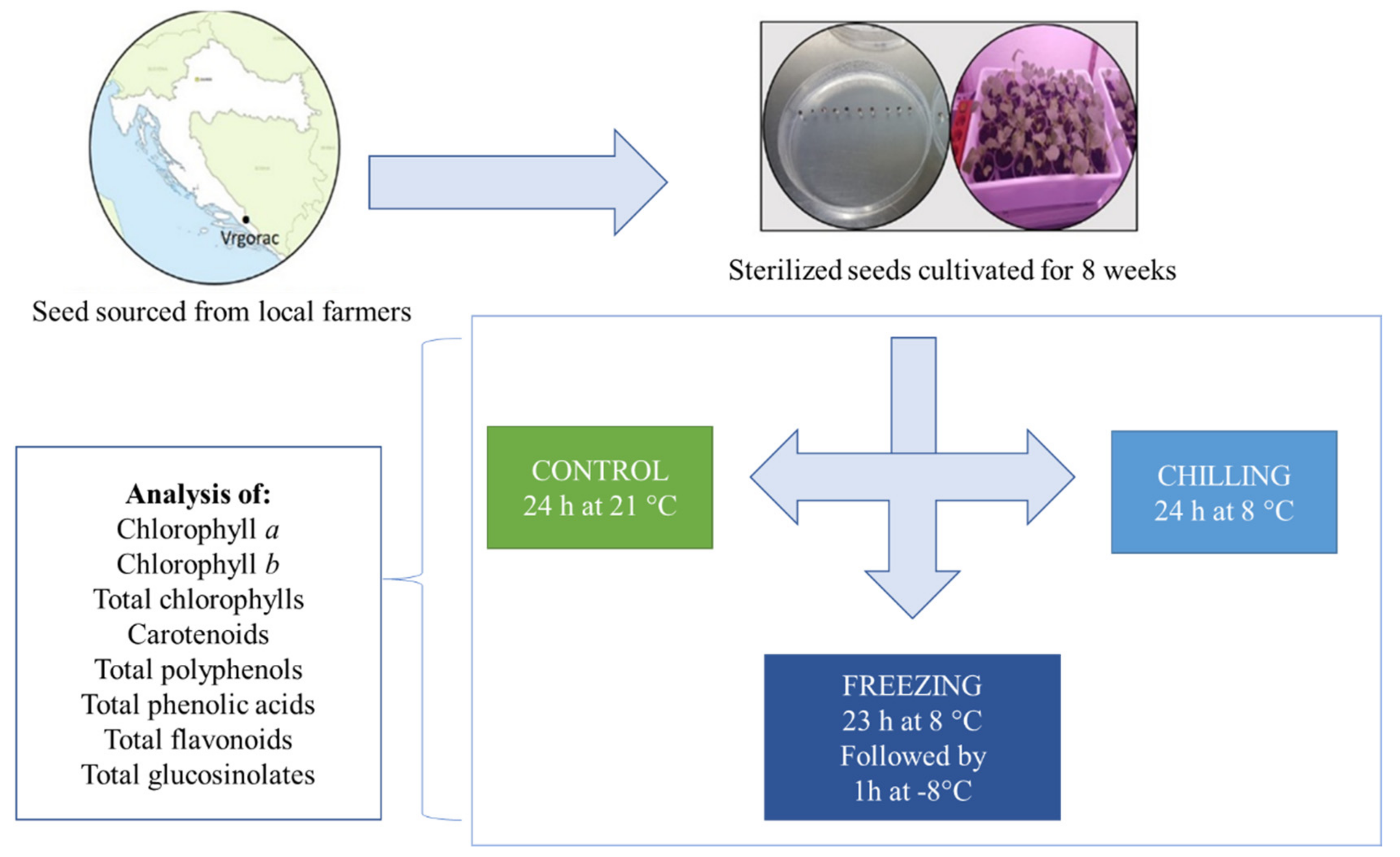
2.1. Plant Growing and Stress Experiments
2.2. Determination of Proline Content
2.3. Determination of Chlorophylls and Carotenoids Content
2.4. Determination of Polyphenolic Compounds
2.5. Determination of Total Glucosinolates
2.6. Statistical Analysis
3. Results
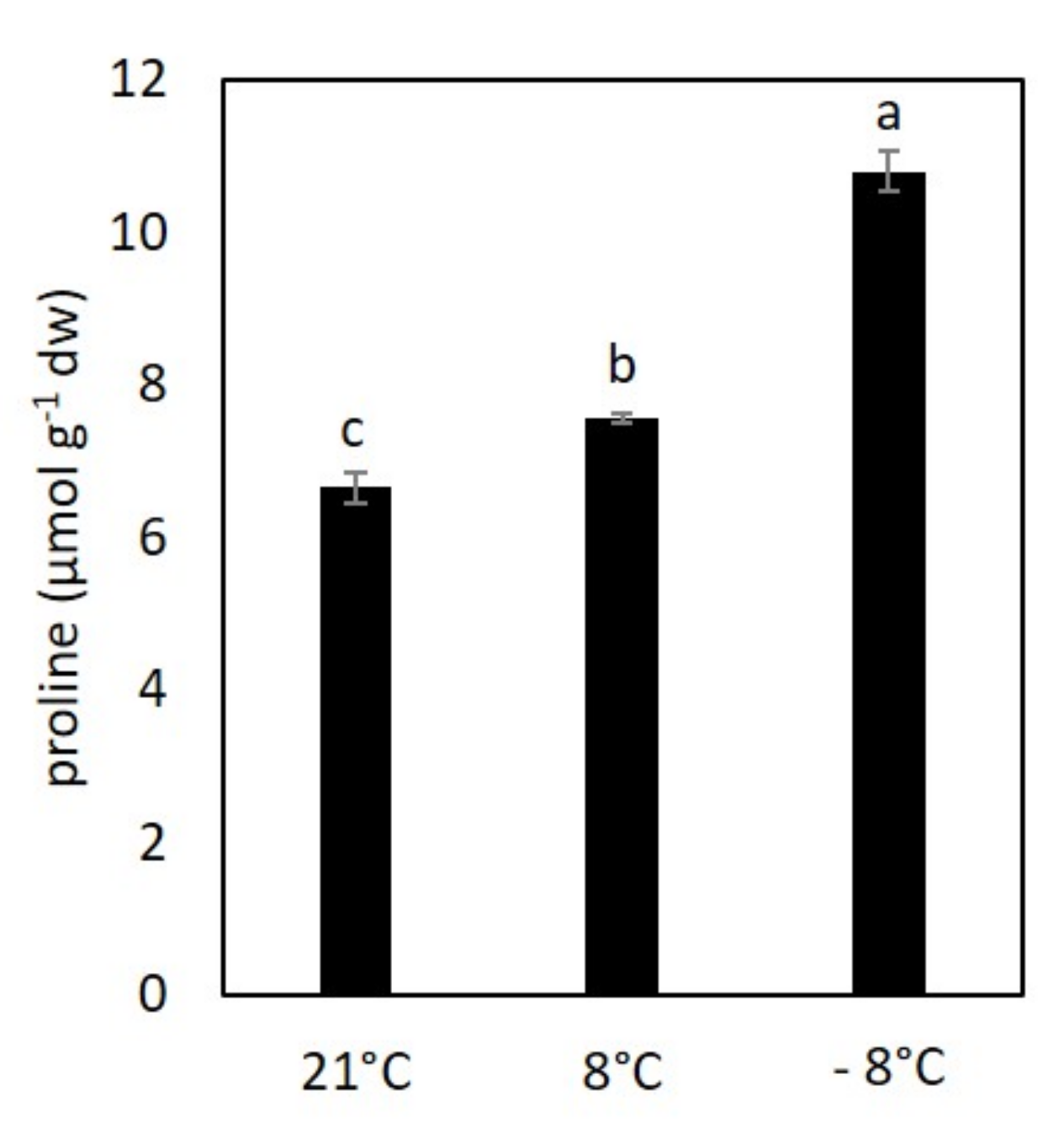
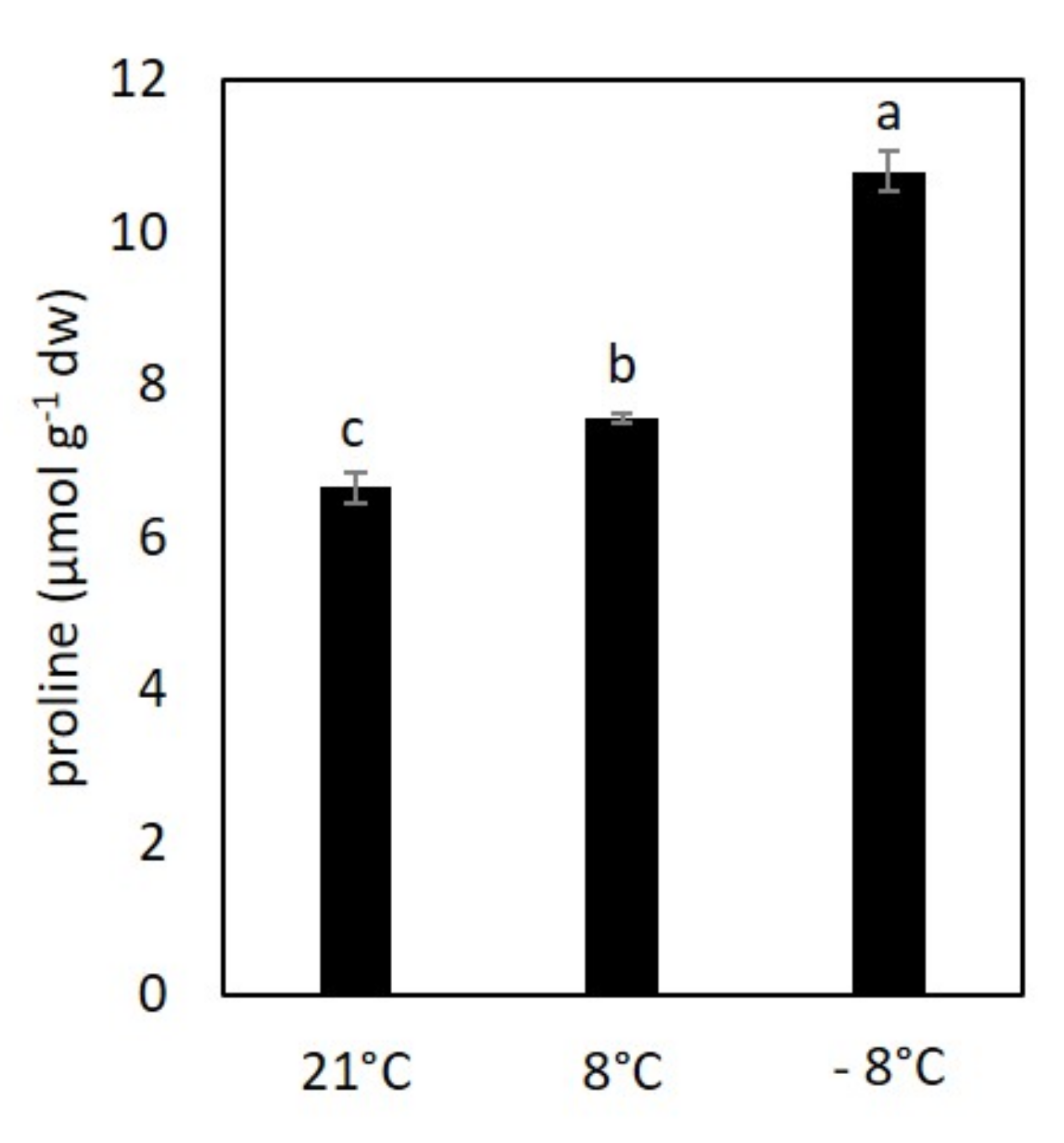
3.1. Effect of Cold Stress on Proline Content in Kale
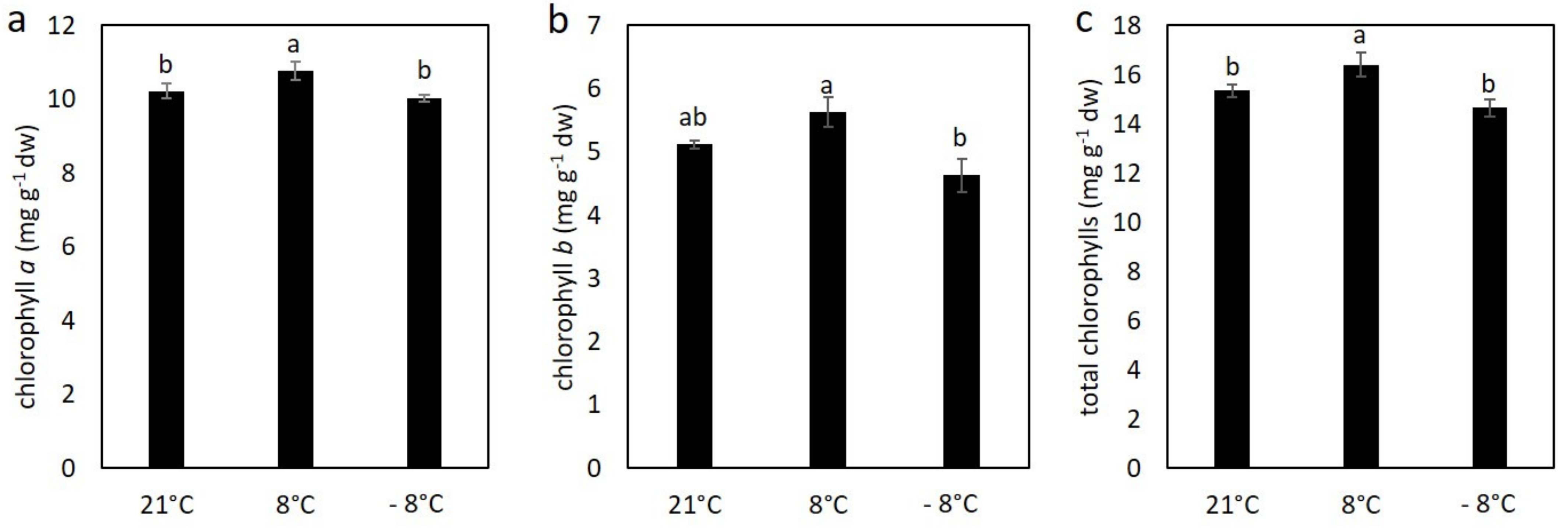
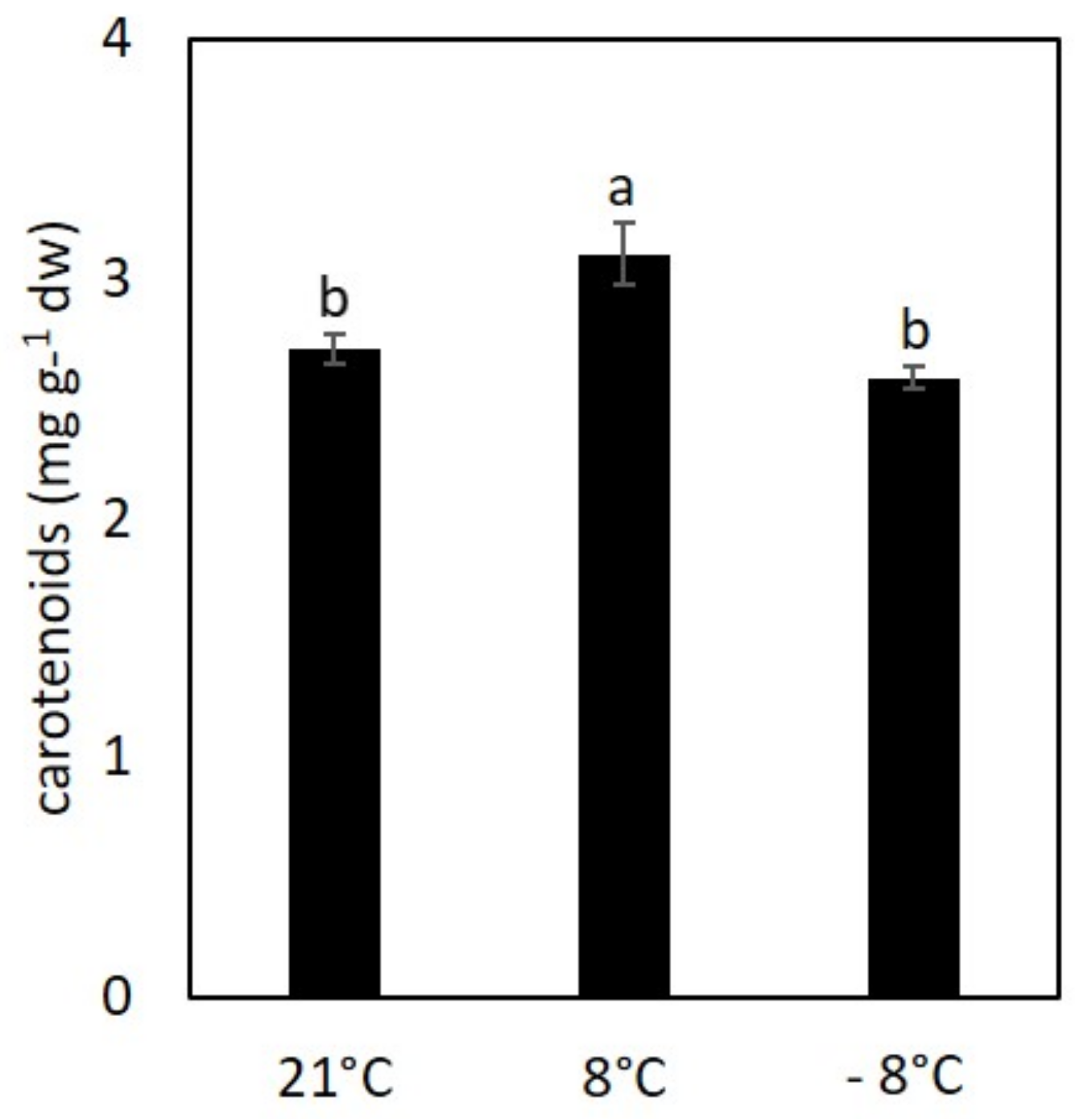
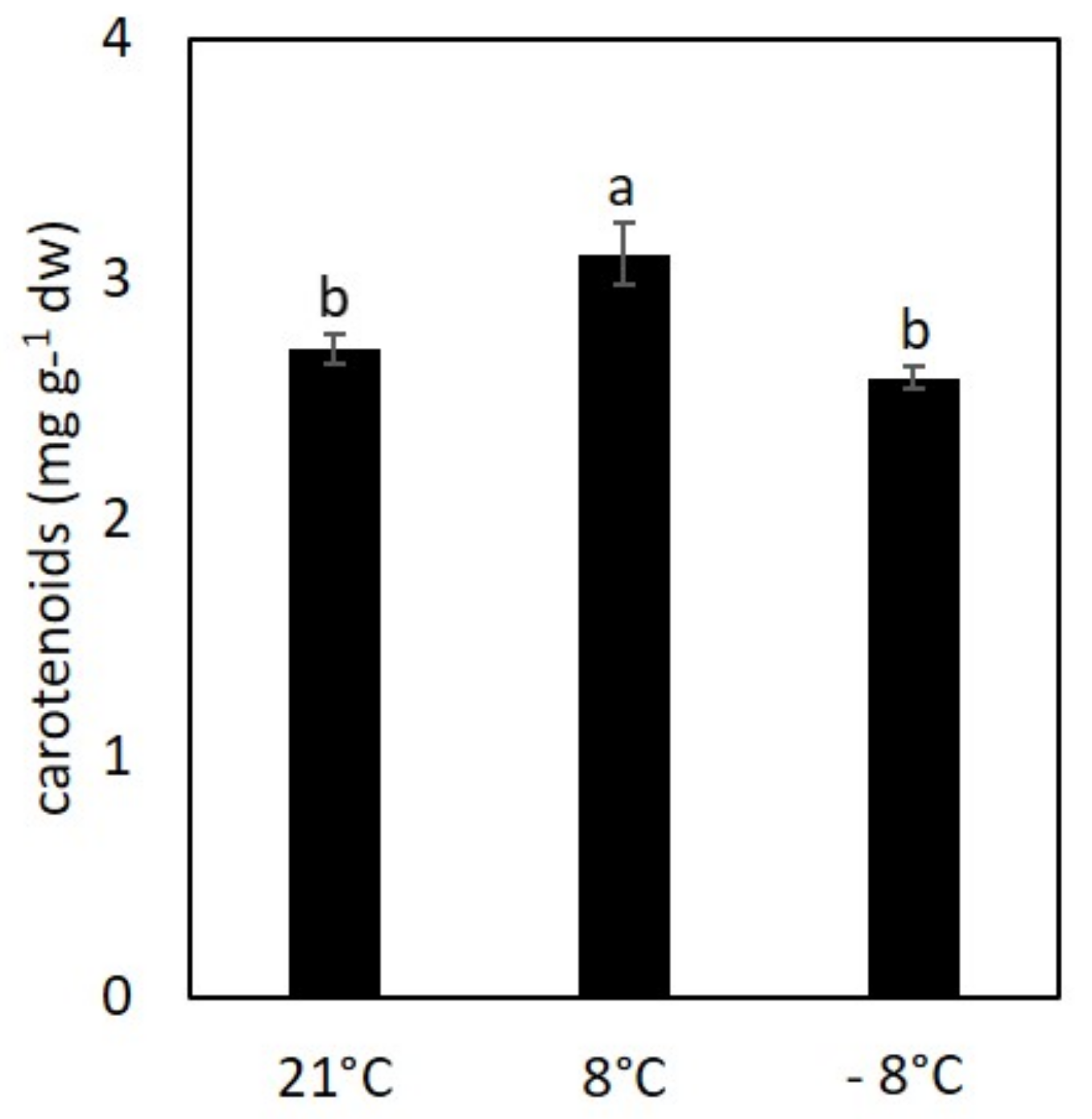
3.2. Effect of Cold Stress on Content of Chlorophylls and Carotenoids in Kale
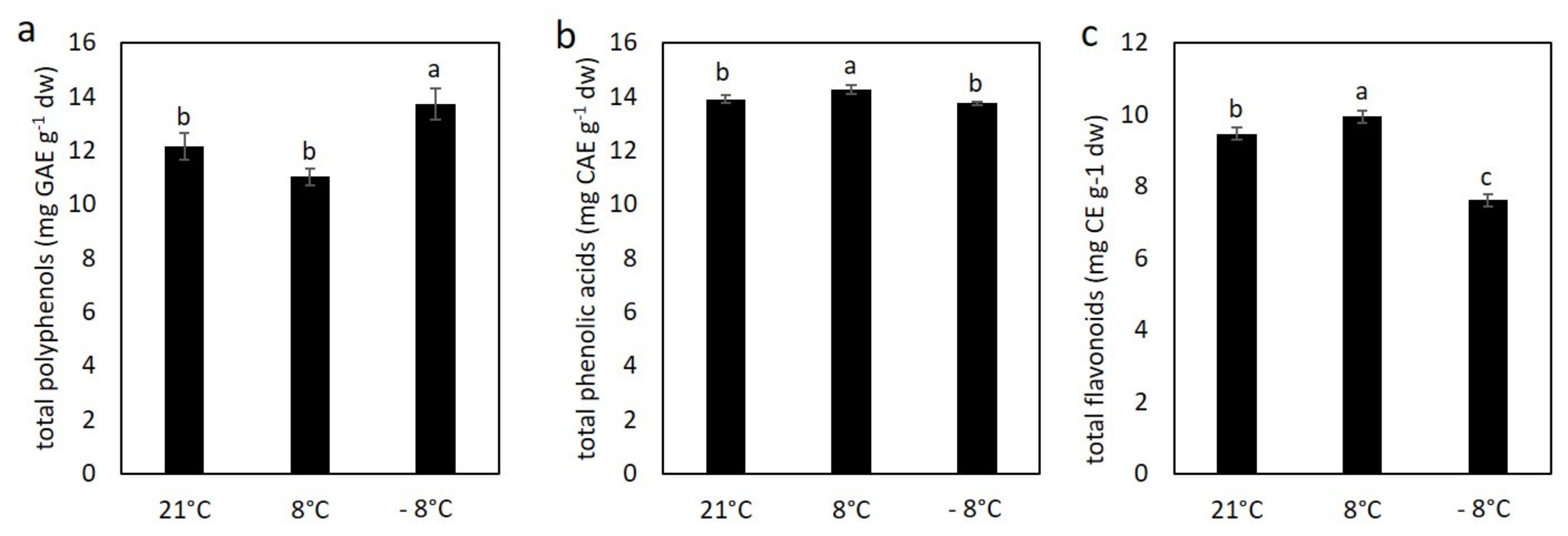
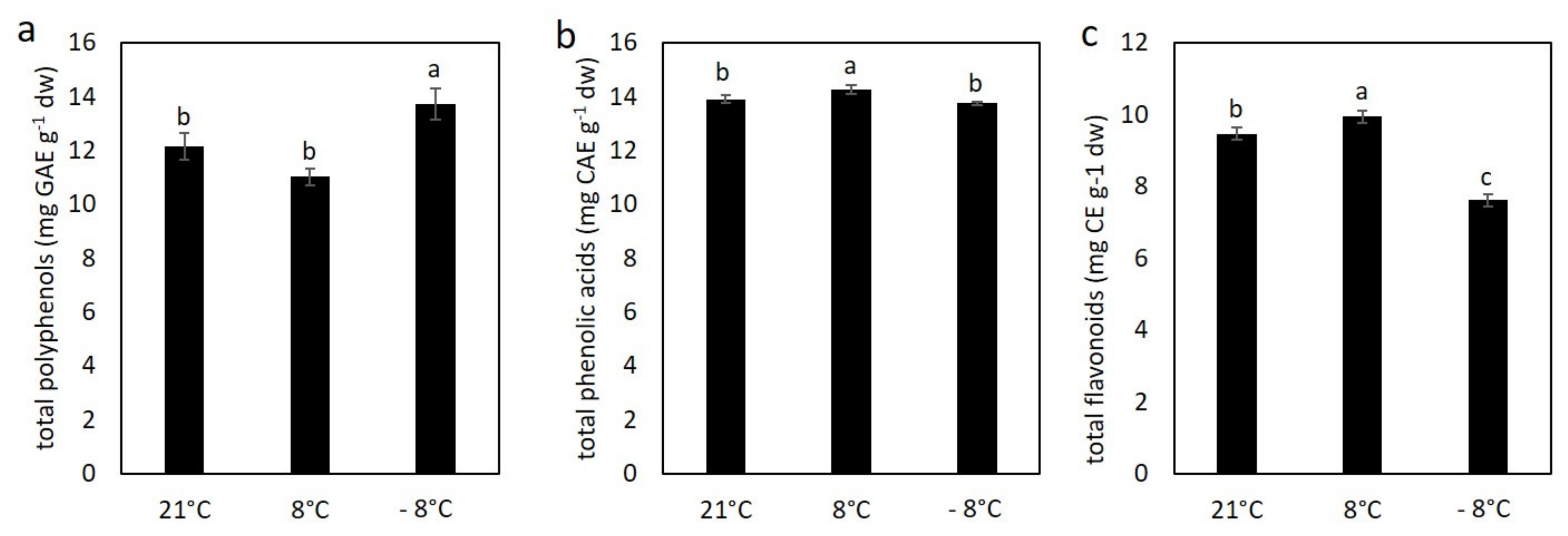
3.3. Effect of Cold Stress on Content of Polyphenolic Compounds in Kale
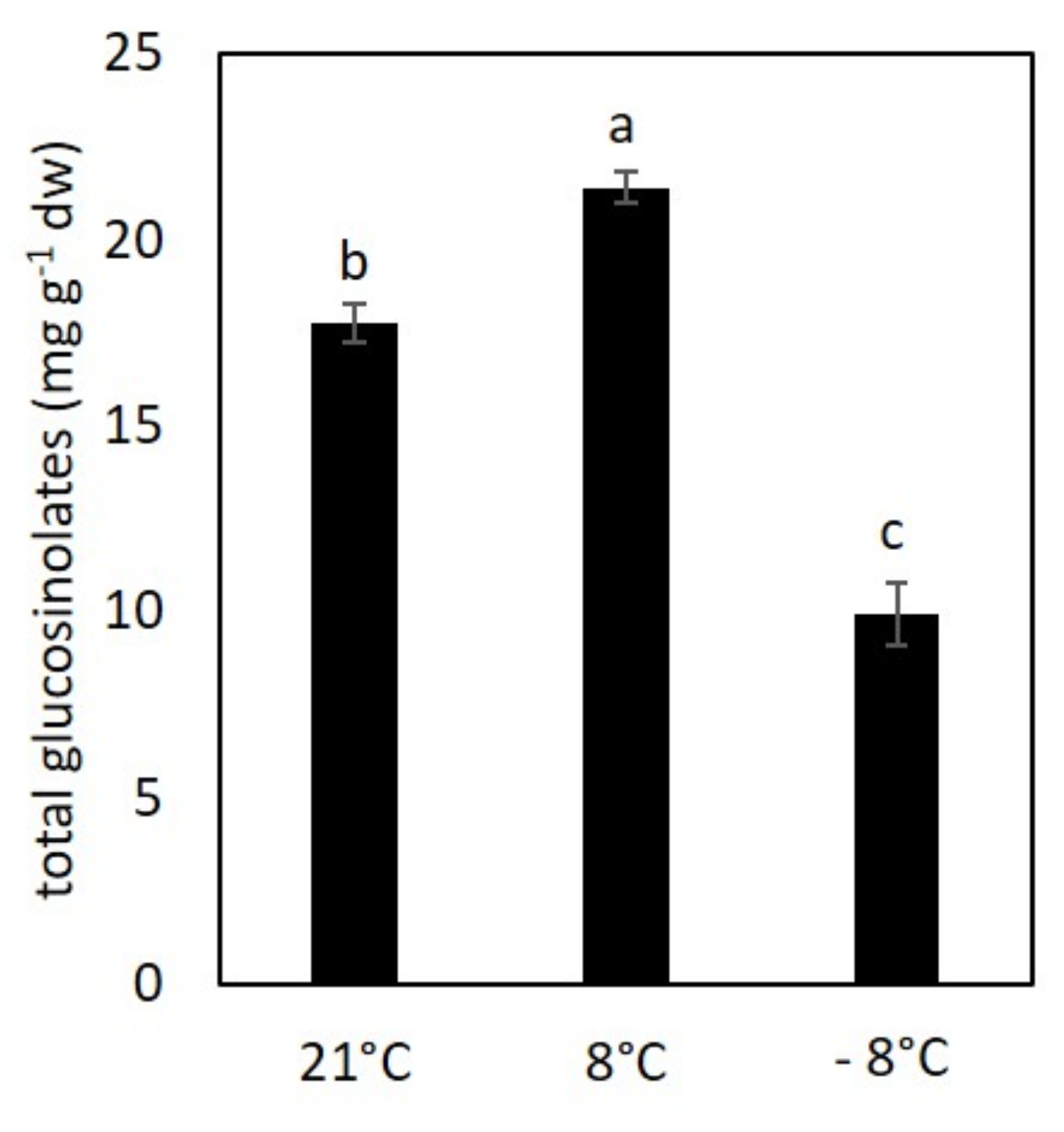
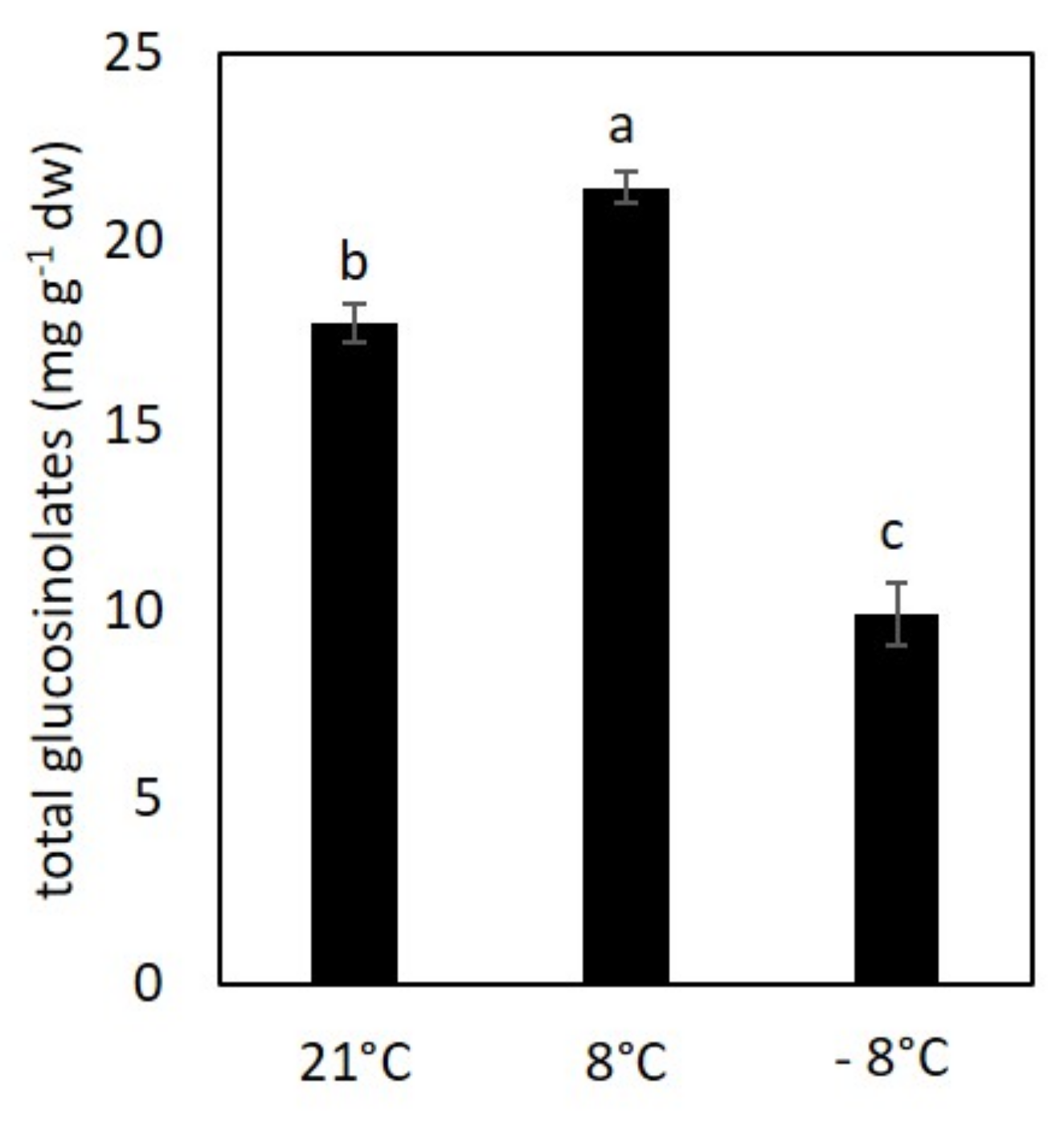
3.4. Content of Glucosinolates under Low Temperature
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ritonga, F.N.; Chen, S. Physiological and Molecular Mechanism Involved in Cold Stress Tolerance in Plants. Plants 2020, 9, 560. [Google Scholar] [CrossRef]
- Hussain, H.A.; Hussain, S.; Khaliq, A.; Ashraf, U.; Anjum, S.A.; Men, S.; Wang, L. Chilling and Drought Stresses in Crop Plants: Implications, Cross Talk, and Potential Management Opportunities. Front. Plant Sci. 2018, 9, 393. [Google Scholar] [CrossRef]
- Sanghera, G.S.; Wani, S.H.; Hussain, W.; Singh, N.B. Engineering Cold Stress Tolerance in Crop Plants. Curr. Genom. 2011, 12, 30–43. [Google Scholar] [CrossRef] [Green Version]
- Carvajal, F.; Rosales, R.; Palma, F.; Manzano, S.; Cañizares, J.; Jamilena, M.; Garrido, D. Transcriptomic changes in Cucurbita pepo fruit after cold storage: Differential response between two cultivars contrasting in chilling sensitivity. BMC Genom. 2018, 19, 125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, L.; Wen, K.-S.; Ruan, X.; Zhao, Y.-X.; Wei, F.; Wang, Q. Response of Plant Secondary Metabolites to Environmental Factors. Molecules 2018, 23, 762. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Franzke, A.; Lysak, M.A.; Al-Shehbaz, I.A.; Koch, M.A.; Mummenhoff, K. Cabbage family affairs: The evolutionary history of Brassicaceae. Trend. Plant Sci. 2011, 16, 108–116. [Google Scholar] [CrossRef]
- Šamec, D.; Salopek-Sondi, B. Cruciferous (Brassicaceae) vegetables. In Nonvitamin and Nonmineral Nutritional Supplements; Nabavi, S.M., Sanches Silva, T., Eds.; Academic Press: Cambridge, MA, USA, 2019; pp. 195–202. [Google Scholar]
- Šamec, D.; Urlić, B.; Salopek-Sondi, B. Kale (Brassica oleracea var. acephala) as a superfood: Review of the scientific evidence behind the statement. Crit. Rev. Food Sci. Nutr. 2019, 59, 2411–2422. [Google Scholar] [CrossRef] [PubMed]
- Pavlović, I.; Petrík, I.; Tarkowska, D.; Lepeduš, H.; Vujčić Bok, V.; Radić Brkanac, S.; Novák, O.; Salopek Sondi, B. Correlations between Phytohormones and Drought Tolerance in Selected Brassica Crops: Chinese Cabbage, White Cabbage and Kale. Int. J. Mol. Sci. 2018, 19, 2866. [Google Scholar] [CrossRef] [Green Version]
- Linić, I.; Šamec, D.; Grúz, J.; Vujčić Bok, V.; Strnad, M.; Salopek Sondi, B. Involvement of Phenolic Acids in Short-Term Adaptation to Salinity Stress is Species-Specific among Brassicaceae. Plants 2019, 8, 155. [Google Scholar] [CrossRef] [Green Version]
- Šamec, D.; Linić, I.; Salopek-Sondi, B. Salinity Stress as an Elicitor for Phytochemicals and Minerals Accumulation in Selected Leafy Vegetables of Brassicaceae. Agronomy 2021, 11, 361. [Google Scholar] [CrossRef]
- Ljubej, V.; Radojčić Redovniković, I.; Salopek-Sondi, B.; Smolko, A.; Roje, S.; Šamec, D. Chilling and Freezing Temperature Stress Differently Influence Glucosinolates Content in Brassica oleracea var. acephala. Plants 2021, 10, 1305. [Google Scholar] [CrossRef]
- Šamec, D.; Pavlović, I.; Radojčić Redovniković, I.; Salopek-Sondi, B. Comparative analysis of phytochemicals and activity of endogenous enzymes associated with their stability, bioavailability and food quality in five Brassicaceae sprouts. Food Chem. 2018, 269, 96–102. [Google Scholar] [CrossRef]
- Šamec, D.; Kruk, V.; Ivanišević, P. Influence of Seed Origin on Morphological Characteristics and Phytochemicals Levels in Brassica oleracea var. acephala. Agronomy 2019, 9, 502. [Google Scholar] [CrossRef] [Green Version]
- USDA, National Agricultural Statistics Service. 2012; Census of Agriculture. Available online: https://www.agcensus.usda.gov/Publications/2012/Full_Report/Volume_1,_Chapter_1_US/st99_1_038_038.pdf (accessed on 18 September 2021).
- Lee, J.-H.; Oh, M.-M. Short-term Low Temperature Increases Phenolic Antioxidant Levels in Kale. Hortic. Environ. Biotechnol. 2015, 56, 588–596. [Google Scholar] [CrossRef]
- Lee, J.-H.; Kwon, M.C.; Jung, E.S.; Lee, C.H.; Oh, M.-M. Physiological and Metabolomic Responses of Kale to Combined Chilling and UV-A Treatment. Int. J. Mol. Sci. 2019, 20, 4950. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steindal, A.L.H.; Rødven, R.; Hansen, E.; Mølmann, J. Effects of photoperiod, growth temperature and cold acclimatisation on glucosinolates, sugars and fatty acids in kale. Food Chem. 2015, 174, 44–51. [Google Scholar] [CrossRef] [PubMed]
- Lichtenthaler, H.K.; Buschmann, C. Chlorophylls and carotenoids: Measurement and characterization by UV–VIS spectroscopy. In Current Protocols in Food Analytical Chemistry; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2001; pp. F4.3.1–F4.3.8. [Google Scholar]
- Šamec, D.; Bogović, M.; Vincek, D.; Martinčić, J.; Salopek-Sondi, B. Assessing the authenticity of the white cabbage (Brassica oleracea var. capitata f. alba) cv. ‘Varaždinski’ by molecular and phytochemical markers. Food Res. Int. 2014, 60, 266–272. [Google Scholar] [CrossRef]
- Aghajanzadeh, T.; Hawkesford, M.J.; Kok, L.J. The significance of glucosinolates for sulfur storage in Brassicaceae seedlings. Front. Plant Sci. 2014, 5, 704. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hammer, O.; Harper, A.A.T.; David, A.T.; Ryan, P.D. PAST: Paleontological Statistics Software Package for Education and Data Analysis. Palaeontol. Electron 2001, 4, 1–9. [Google Scholar]
- Hayat, S.; Hayat, Q.; Alyemeni, M.N.; Wani, A.S.; Pichtel, J.; Ahmad, A. Role of proline under changing environments. Plant Signal. Behav. 2012, 7, 1456–1466. [Google Scholar] [CrossRef] [Green Version]
- Hadi, F.; Gilpin, M.; Fuller, M.P. Identification and expression analysis of CBF/DREB1 and COR15 genes in mutants of Brassica oleracea var. botrytis with enhanced proline production and frost resistance. Plant Physiol. Biochem. 2011, 49, 1323–1332. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fuller, M.P.; Metwali, E.M.R.; Eed, M.H.; Jellings, A.J. Evaluation of Abiotic Stress Resistance in Mutated Populations of Cauliflower (Brassica oleracea var. botrytis). Plant Cell Tissue Organ. Cult. 2006, 86, 239. [Google Scholar] [CrossRef]
- Neto, N.B.M.; Custódio, C.C.; Gatti, A.B.; Priolli, M.R.; Cardoso, V.J.M. Proline: Use as an indicator of temperature stress in bean seeds. Crop Breed. Appl. Biotechnol. 2004, 4, 330–337. [Google Scholar] [CrossRef]
- Vera-Hernández, P.; Ramírez, M.A.O.; Núñez, M.M.; Ruiz-Rivas, M.; Rosas-Cárdenas, F.F. Proline as a probable biomarker of cold stress tolerance in Sorghum (Sorghum bicolor). Mex. J. Biotechnol. 2018, 3, 77–86. [Google Scholar] [CrossRef]
- de Freitas, G.M.; Thomas, J.; Liyanage, R.; Lay, J.O.; Basu, S.; Ramegowda, V.; do Amaral, M.N.; Benitez, L.C.; Braga, E.J.B.; Pereira, A. Cold tolerance response mechanisms revealed through comparative analysis of gene and protein expression in multiple rice genotypes. PLoS ONE 2019, 14, e0218019. [Google Scholar] [CrossRef]
- Ashraf, M.; Harris, P.J.C. Photosynthesis under stressful environments: An overview. Photosynthetica 2013, 51, 163–190. [Google Scholar] [CrossRef]
- Ensminger, I.; Busch, F.; Huner, N.P.A. Photostasis and cold acclimation: Sensing low temperature through photosynthesis. Phisiol. Plant. 2006, 126, 28–44. [Google Scholar] [CrossRef]
- Zhao, Y.; Han, Q.; Ding, C.; Huang, Y.; Liao, J.; Tao Chen, T.; Feng, S.; Zhou, L.; Zhang, Z.; Chen, Y.; et al. Effect of Low Temperature on Chlorophyll Biosynthesis and Chloroplast Biogenesis of Rice Seedlings during Greening. Int. J. Mol. Sci. 2020, 21, 1390. [Google Scholar] [CrossRef] [Green Version]
- Mageney, V.; Baldermann, S.; Albach, D.C. Intraspecific Variation in Carotenoids of Brassica oleracea var. sabellica. J. Agric. Food Chem. 2016, 64, 3251–3257. [Google Scholar] [CrossRef] [PubMed]
- Maoka, T. Carotenoids as natural functional pigments. J. Nat. Med. 2020, 74, 1–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hwang, S.-J.; Chun, J.-H.; Kim, S.-J. Effect of Cold Stress on Carotenoids in Kale Leaves (Brassica oleracea). Korean J. Environ. Agric. 2017, 36, 106–112. [Google Scholar] [CrossRef] [Green Version]
- Šamec, D.; Karalija, E.; Šola, I.; Vujčić-Bok, V.; Salopek-Sondi, B. The Role of Polyphenols in Abiotic Stress Response: The Influence of Molecular Structure. Plants 2021, 10, 118. [Google Scholar] [CrossRef] [PubMed]
- Jurkow, R.; Wurst, A.; Kalisz, A.; Sękara, A.; Cebula, S. Cold stress modifies bioactive compounds of kale cultivars during fall–winter harvests. Acta Agrobot. 2019, 72, 1761. [Google Scholar] [CrossRef] [Green Version]
- Schmidt, S.; Zietz, M.; Schreiner, M.; Rohn, S.; Kroh, L.W.; Krumbein, A. Genotypic and climatic influences on the concentration and composition of flavonoids in kale (Brassica oleracea var. sabellica). Food. Chem. 2010, 119, 1293–1299. [Google Scholar] [CrossRef]
- Stefanowska, M.; Kuras, M.; Kacperska, A. Low Temperature-induced Modifications in Cell Ultrastructure and Localization of Phenolics in Winter Oilseed Rape (Brassica napus L. var. oleifera L.) Leaves. Ann Bot. 2002, 90, 637–645. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Control (21 °C) | Chilling 24 h (8 °C) | Freezing 1 h after Acclimatization 23 h (−8 °C) | |
|---|---|---|---|
| Chlorophyll a/ chlorophyll b | 2.00 ± 0.02 b | 1.91 ± 0.04 b | 2.17 ± 0.10 a |
| Total chlorophylls/ total carotenoids | 5.67 ± 0.07 a | 5.29 ± 0.14 a | 5.67 ± 0.24 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ljubej, V.; Karalija, E.; Salopek-Sondi, B.; Šamec, D. Effects of Short-Term Exposure to Low Temperatures on Proline, Pigments, and Phytochemicals Level in Kale (Brassica oleracea var. acephala). Horticulturae 2021, 7, 341. https://doi.org/10.3390/horticulturae7100341
Ljubej V, Karalija E, Salopek-Sondi B, Šamec D. Effects of Short-Term Exposure to Low Temperatures on Proline, Pigments, and Phytochemicals Level in Kale (Brassica oleracea var. acephala). Horticulturae. 2021; 7(10):341. https://doi.org/10.3390/horticulturae7100341
Chicago/Turabian StyleLjubej, Valentina, Erna Karalija, Branka Salopek-Sondi, and Dunja Šamec. 2021. "Effects of Short-Term Exposure to Low Temperatures on Proline, Pigments, and Phytochemicals Level in Kale (Brassica oleracea var. acephala)" Horticulturae 7, no. 10: 341. https://doi.org/10.3390/horticulturae7100341
APA StyleLjubej, V., Karalija, E., Salopek-Sondi, B., & Šamec, D. (2021). Effects of Short-Term Exposure to Low Temperatures on Proline, Pigments, and Phytochemicals Level in Kale (Brassica oleracea var. acephala). Horticulturae, 7(10), 341. https://doi.org/10.3390/horticulturae7100341