Fruit Stem-End Rot



Abstract
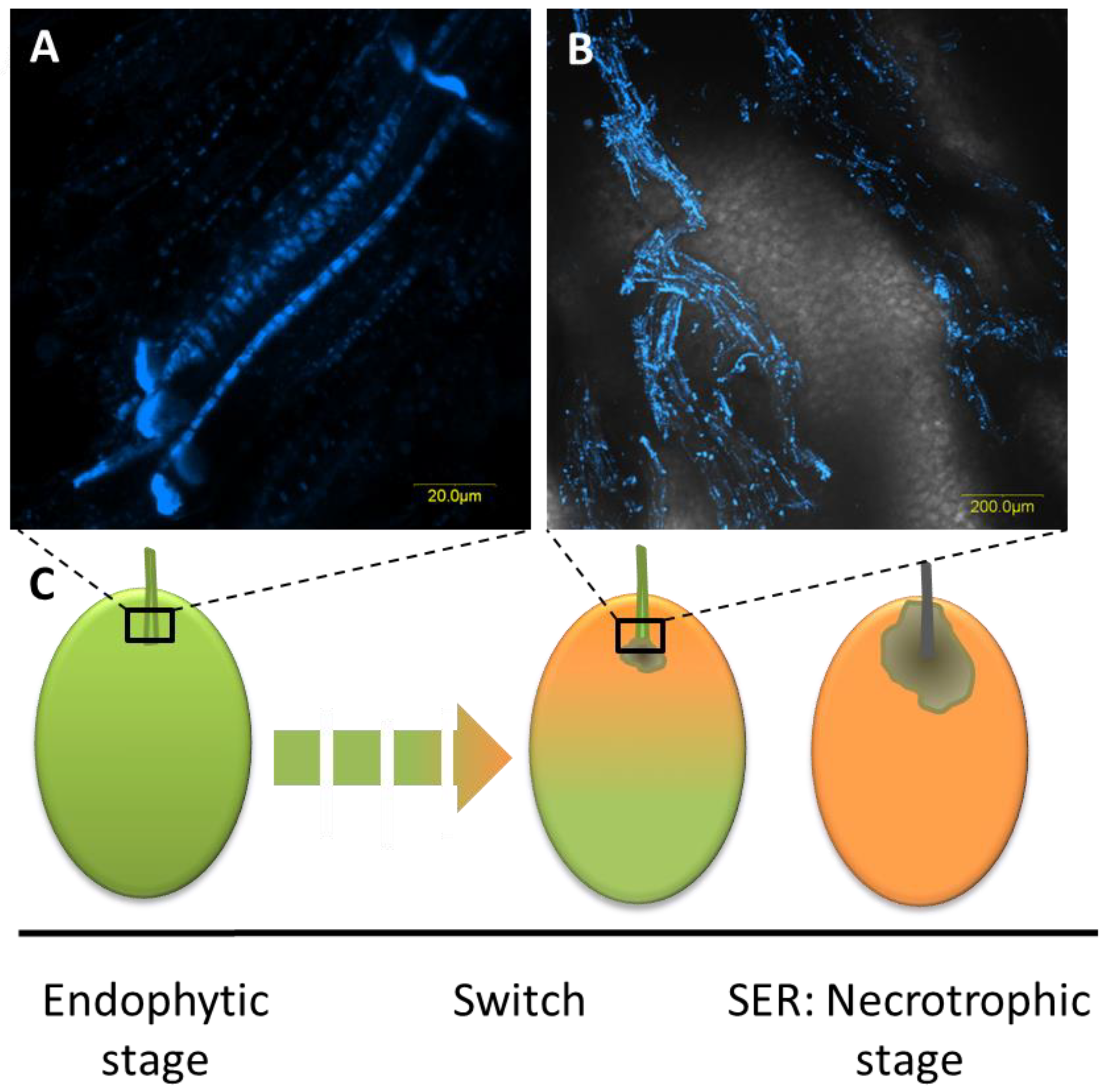
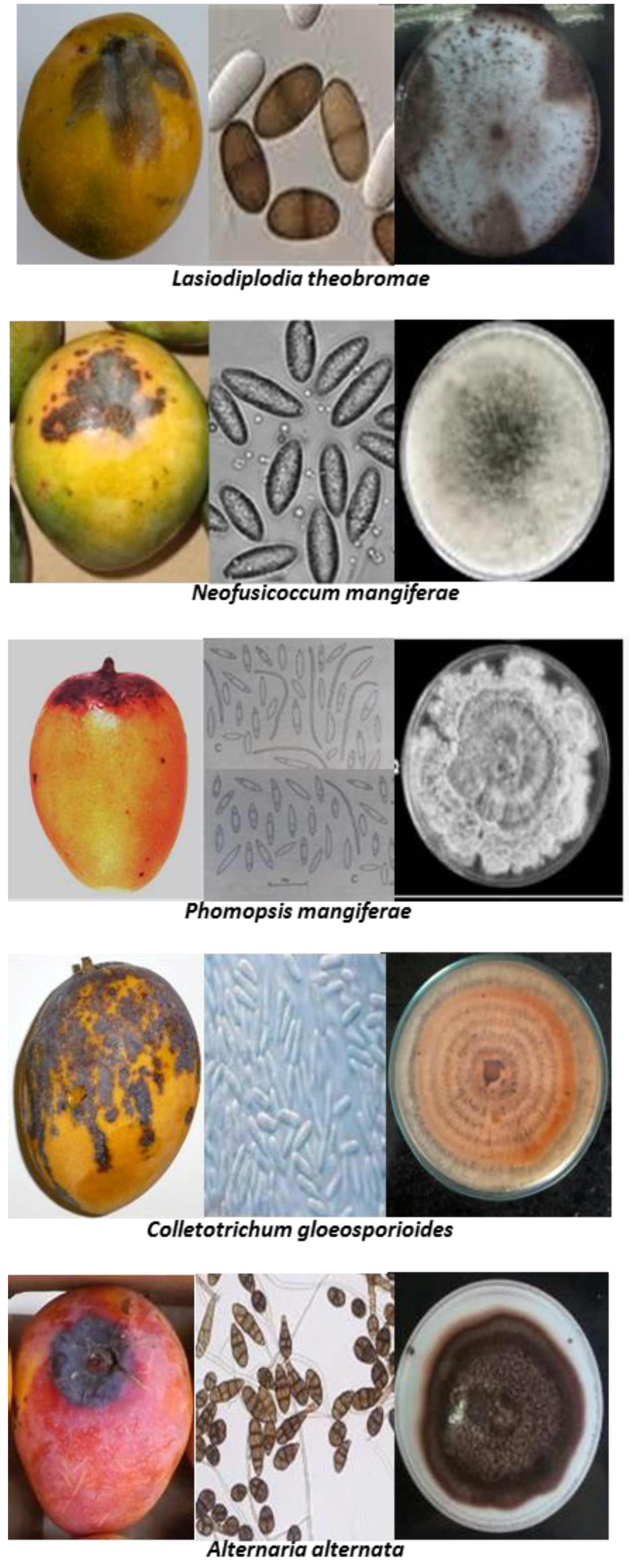
1. Characterization of Stem-End Rot Causing Pathogens and Their Lifestyle
2. Endophytic Community in Fruit Stem-End
3. Factors and Treatments Affecting SER
3.1. Inhibition of Fruit Ripening
3.2. Harvesting with Stem
3.3. Chemical Treatments
3.3.1. Preharvest Chemical Control
3.3.2. Postharvest Chemical Control
3.4. Biological Control
3.4.1. Preharvest Biological Control
3.4.2. Postharvest Biological Control
3.5. Plant Extracts
3.6. Physical Control
3.7. Combined Treatments
4. Fruit Stem-End Microbiome and Modern Molecular Tools
5. Summary and Future Directions
Author Contributions
Funding
Conflicts of Interest
References
- Alkan, N.; Fortes, A.M. Insights into molecular and metabolic events associated with fruit response to post-harvest fungal pathogens. Front. Plant Sci. 2015, 6, 889. [Google Scholar] [CrossRef] [PubMed]
- Prusky, D.; Kobiler, I.; Miyara, I.; Alkan, N. Fruit diseasess. In The Mango, Botany, Production and Uses, 2nd ed.; Litz, R., Ed.; CABI International: Wallingford, UK, 2009; pp. 210–231. [Google Scholar]
- Alkan, N.; Kumar, P. Postharvest storage management of mango fruit. In Achieving Sustainable Cultivation of Mango; Galán Saúco, V., Lu, P., Eds.; Burleigh Dodds Science Publishing: Cambridge, UK, 2018; pp. 377–402. [Google Scholar]
- Diskin, S.; Feygenberg, O.; Maurer, D.; Alkan, N. Biological and chemical application during flowering increase fruit count, yield and reduce postharvest decay in mango fruit. Front. Plant Sci. submitted.
- Johnson, G.I.; Mead, A.J.; Cooke, A.W.; Dean, J.R. Mango stem end rot pathogens—Fruit infection by endophytic colonisation of the inflorescence and pedicel. Ann. Appl. Biol. 1992, 120, 225–234. [Google Scholar] [CrossRef]
- Diskin, S.; Feygenberg, O.; Maurer, D.; Droby, S.; Prusky, D.; Alkan, N. Microbiome alterations are correlated with occurrence of postharvest stem-end rot in mango fruit. Phytobiomes 2017, 1, 117–127. [Google Scholar] [CrossRef]
- Prusky, D.; Alkan, N.; Mengiste, T.; Fluhr, R. Quiescent and necrotrophic lifestyle choice during postharvest disease development. Annu. Rev. Phytopathol. 2013, 51, 155–176. [Google Scholar] [CrossRef] [PubMed]
- Ploetz, R.C. Compendium of Tropical Fruit Diseases; APS Press: St. Paul, MN, USA, 1994. [Google Scholar]
- Pérez-Jiménez, R.M. Significant avocado diseases caused by fungi and oomycetes. Eur. J. Plant Sci. Biotechnol. 2008, 2, 1–24. [Google Scholar]
- Darvas, J.; Kotze, J.; Wehner, F. Effect of treatment after picking on the incidence of post-harvest fruit diseases of avocado. Phytophylactica 1990, 22, 93–96. [Google Scholar]
- Hopkirk, G.; White, A.; Beever, D.; Forbes, S. Influence of postharvest temperatures and the rate of fruit ripening on internal postharvest rots and disorders of New Zealand ‘Hass’ avocado fruit. N. Z. J. Crop Hortic. Sci. 1994, 22, 305–311. [Google Scholar] [CrossRef]
- Arpaia, M.L.; Collin, S.; Sievert, J.; Obenland, D. Influence of cold storage prior to and after ripening on quality factors and sensory attributes of ‘Hass’ avocados. Postharvest Biol. Technol. 2015, 110, 149–157. [Google Scholar] [CrossRef]
- Johnson, G.I.; Mead, A.J.; Cooke, A.W.; Dean, J.R. Mango stem end rot pathogens—Infection levels between flowering and harvest. Ann. Appl. Biol. 1991, 119, 465–473. [Google Scholar] [CrossRef]
- Hartill, W.; Everett, K. Inoculum sources and infection pathways of pathogens causing stem-end rots of ‘Hass’ avocado (Persea americana). N. Z. J. Crop Hortic. Sci. 2002, 30, 249–260. [Google Scholar] [CrossRef]
- Coutinho, I.; Freire, F.; Lima, C.; Lima, J.; Gonçalves, F.; Machado, A.; Silva, A.; Cardoso, J. Diversity of genus Lasiodiplodia associated with perennial tropical fruit plants in northeastern Brazil. Plant Pathol. 2017, 66, 90–104. [Google Scholar] [CrossRef]
- Zhang, H.; Wei, Y.; Qi, Y.; Pu, J.; Liu, X. First report of Lasiodiplodia Brasiliense associated with stem-end rot of mango in China. Plant Dis. 2018, 102, 679. [Google Scholar] [CrossRef]
- Rodríguez-Gálvez, E.; Guerrero, P.; Barradas, C.; Crous, P.W.; Alves, A. Phylogeny and pathogenicity of Lasiodiplodia species associated with dieback of mango in Peru. Fungal Biol. 2017, 121, 452–465. [Google Scholar] [CrossRef] [PubMed]
- Sathya, K.; Parthasarathy, S.; Thiribhuvanamala, G.; Prabakar, K. Morphological and molecular variability of Lasiodiplodia theobromae causing stem end rot of mango in Tamil Nadu, India. Int. J. Pure Appl. Biosci. 2017, 5, 1024–1031. [Google Scholar] [CrossRef]
- Liu, J.; Abdelfattah, A.; Norelli, J.; Burchard, E.; Schena, L.; Droby, S.; Wisniewski, M. Apple endophytic microbiota of different rootstock/scion combinations suggests a genotype-specific influence. Microbiome 2018, 6, 18. [Google Scholar] [CrossRef] [PubMed]
- Wilson, D. Endophyte: The evolution of a term, and clarification of its use and definition. Oikos 1995, 73, 274–276. [Google Scholar] [CrossRef]
- Porras-Alfaro, A.; Bayman, P. Hidden fungi, emergent properties: Endophytes and microbiomes. Annu. Rev. Phytopathol. 2011, 49, 291–315. [Google Scholar] [CrossRef] [PubMed]
- Arnold, A.E. Understanding the diversity of foliar endophytic fungi: Progress, challenges, and frontiers. Fungal Biol. Rev. 2007, 21, 51–66. [Google Scholar] [CrossRef]
- Gond, S.; Verma, V.; Kumar, A.; Kumar, V.; Kharwar, R. Study of endophytic fungal community from different parts of Aegle marmelos Correae (Rutaceae) from Varanasi (India). World J. Microbiol. Biotechnol. 2007, 23, 1371–1375. [Google Scholar] [CrossRef]
- Abarenkov, K.; Henrik Nilsson, R.; Larsson, K.H.; Alexander, I.J.; Eberhardt, U.; Erland, S.; Høiland, K.; Kjøller, R.; Larsson, E.; Pennanen, T. The unite database for molecular identification of fungi–recent updates and future perspectives. New Phytol. 2010, 186, 281–285. [Google Scholar] [CrossRef] [PubMed]
- Sommer, N.F. Role of controlled environments in suppression of postharvest diseases. Can. J. Plant Pathol. 1985, 7, 331–339. [Google Scholar] [CrossRef]
- Kumar, R.; Khurana, A.; Sharma, A.K. Role of plant hormones and their interplay in development and ripening of fleshy fruits. J. Exp. Bot. 2014, 65, 4561–4575. [Google Scholar] [CrossRef] [PubMed]
- Coggins, C.W., Jr. Use of growth regulators to delay maturity and prolong shelf life of citrus. In Proceedings of the Symposium on Growth Regulators in Fruit Production, St. Paul, MN, USA, 11–15 November 1972; pp. 469–472. [Google Scholar]
- Kobiler, I.; Shalom, Y.; Roth, I.; Akerman, M.; Vinokur, Y.; Fuchs, Y.; Prusky, D. Effect of 2,4-dichlorophenoxyacetic acid on the incidence of side and stem end rots in mango fruits. Postharvest Biol. Technol. 2001, 23, 23–32. [Google Scholar] [CrossRef]
- Watkins, C.B.; Nock, J.F. Effects of delayed controlled atmosphere storage of apples after rapid 1-MCP treatment. HortScience 2008, 43, 1087. [Google Scholar]
- Watkins, C.B. The use of 1-methylcyclopropene (1-MCP) on fruits and vegetables. Biotechnol. Adv. 2006, 24, 389–409. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Tian, S.; Zhu, Z.; Xu, Y.; Qin, G. Effects of 1-methylcyclopropene (1-MCP) on ripening and resistance of jujube (Zizyphus jujuba cv. Huping) fruit against postharvest disease. LWT-Food Sci. Technol. 2012, 45, 13–19. [Google Scholar] [CrossRef]
- Zhong, Q.; Xia, W. Effect of 1-methylcyclopropene and/or chitosan coating treatments on storage life and quality maintenance of Indian jujube fruit. LWT-Food Sci. Technol. 2007, 40, 404–411. [Google Scholar]
- Diskin, S.; Feygenberg, O.; Maurer, D.; Prusky, D.; Droby, S.; Alkan, N. Postharvest treatments for stem-end rot in fruits. Alon Hanotea 2015, 70, 36–38. [Google Scholar]
- Porat, R.; Weiss, B.; Cohen, L.; Daus, A.; Goren, R.; Droby, S. Effects of ethylene and 1-methylcyclopropene on the postharvest qualities of ‘Shamouti’ oranges. Postharvest Biol. Technol. 1999, 15, 155–163. [Google Scholar] [CrossRef]
- Hofman, P.J.; Jobin-Decor, M.; Meiburg, G.F.; Macnish, A.J.; Joyce, D.C. Ripening and quality responses of avocado, custard apple, mango and papaya fruit to 1-methylcyclopropene. Aust. J. Exp. Agric. 2001, 41, 567–572. [Google Scholar] [CrossRef]
- Woolf, A.B.; Requejo-Tapia, C.; Cox, K.A.; Jackman, R.C.; Gunson, A.; Arpaia, M.L.; White, A. 1-MCP reduces physiological storage disorders of ‘Hass’ avocados. Postharvest Biol. Technol. 2005, 35, 43–60. [Google Scholar] [CrossRef]
- Dou, H.; Jones, S.; Ritenour, M. Influence of 1-MCP application and concentration on post-harvest peel disorders and incidence of decay in citrus fruit. J. Hortic. Sci. Biotechnol. 2005, 80, 786–792. [Google Scholar] [CrossRef]
- Ku, V.V.V.; Wills, R.B.H.; Ben-Yehoshua, S. 1-methylcyclopropene can differentially affect the postharvest life of strawberries exposed to ethylene. HortScience 1999, 34, 119–120. [Google Scholar]
- Blanco-Ulate, B.; Vincenti, E.; Powell, A.L.T.; Cantu, D. Tomato transcriptome and mutant analyses suggest a role for plant stress hormones in the interaction between fruit and Botrytis cinerea. Front. Plant Sci. 2013, 4, 142. [Google Scholar] [CrossRef] [PubMed]
- Diskin, S.; Lurie, S.; Feygenberg, O.; Maurer, D.; Droby, S.; Prusky, D.; Alkan, N. Postharvest Microbiota Dynamics of Mango Fruit Stem-End. In Proceedings of the IV International Symposium on Postharvest Pathology, Skukuza, South Africa, 28 May–3 June 2017. [Google Scholar]
- Sangchote, S. Botryodiplodia Stem End Rot of Mango and Its Control. In Proceedings of the III International Mango Symposium, Darwin, Australia, 25–29 September 1989; pp. 296–303. [Google Scholar]
- Droby, S.; Prusky, D.; Jacoby, B.; Goldman, A. Induction of antifungal resorcinols in flesh of unripe mango fruits and its relation to latent infection by Alternaria alternata. Physiol. Mol. Plant Pathol. 1987, 30, 285–292. [Google Scholar] [CrossRef]
- Negi, P.; John, S.K.; Rao, P.U. Antimicrobial activity of mango sap. Eur. Food Res. Technol. 2002, 214, 327–330. [Google Scholar]
- Hassan, M.; Dann, E.; Irving, D.; Coates, L. Concentrations of constitutive alk(en)ylresorcinols in peel of commercial mango varieties and resistance to postharvest anthracnose. Physiol. Mol. Plant Pathol. 2007, 71, 158–165. [Google Scholar] [CrossRef]
- Krishnapillai, N.; Wijeratnam, W. Sap volatile components in relation to susceptibility of anthracnose and aspergillus rot of mangoes (Mangifera indica L.). J. Hortic. Sci. Biotechnol. 2017, 92, 206–213. [Google Scholar] [CrossRef]
- Singh, D.; Sharma, R. Postharvest diseases of fruit and vegetables and their management. In Sustainable Pest Management; Prasad, D., Ed.; Daya Publishing House: New Delhi, India, 2007; pp. 67–69. [Google Scholar]
- Brown, G.E.; McCornack, A. Benlate, an experimental preharvest fungicide for control of postharvest citrus fruit decay. Proc. Fla. State Hortic. Soc. 1969 1970, 82, 39–43. [Google Scholar]
- Darvas, J. Pre-harvest chemical control of post-harvest avocado diseases. S. Afr. Avocado Growers’ Assoc. Yearb. 1981, 7, 57–59. [Google Scholar]
- Lonsdale, J.; Kotze, J. Chemical control of mango blossom diseases and the effect on fruit set and yield. Plant Dis. (USA) 1993, 77, 558. [Google Scholar] [CrossRef]
- Dodd, J.C.; Prusky, D.; Jeffries, P. Fruit Diseases. In The Mango: Botany, Production and Uses; CAB International: Wallingford, UK, 1997; pp. 257–280. [Google Scholar]
- Muirhead, I.; Fitzell, R.; Davis, R.; Peterson, R. Post-harvest control of anthracnose and stem-end rots of fuerte avocados with prochloraz and other fungicides. Aust. J. Exp. Agric. 1982, 22, 441–446. [Google Scholar] [CrossRef]
- Everett, K.; Owen, S.; Cutting, J. Testing efficacy of fungicides against postharvest pathogens of avocado (Persea americana cv. Hass). N. Z. Plant Protect. 2005, 58, 89–95. [Google Scholar]
- Estrada, A.; Jeffries, P.; Dodd, J.C. Field evaluation of a predictive model to control anthracnose disease of mango in the Philippines. Plant Pathol. 1996, 45, 294–301. [Google Scholar] [CrossRef]
- Plan, M.; Joyce, D.; Ogle, H.; Johnson, G. Mango stem-end rot (Botryosphaeria dothidea) disease control by partial-pressure infiltration of fungicides. Aust. J. Exp. Agric. 2002, 42, 625–629. [Google Scholar] [CrossRef]
- Swart, S.; Serfontein, J.; Kalinnowski, J. Chemical control of postharvest diseases of mango-the effect of prochloraz, thiobendazole and fludioxonil on soft brown rot, stem-end rot and anthracnose. S. Afr. Mango Growers’ Assoc. Yearb. 2002, 22, 55–62. [Google Scholar]
- Swart, S.; Serfontein, J.; Swart, G.; Labuschagne, C. Chemical Control of Post-Harvest Diseases of Mango: The Effect of Fludioxonil and Prochloraz on Soft Brown Rot, Stem-End Rot and Anthracnose. In Proceedings of the VIII International Mango Symposium, Sun City, South Africa, 5–10 February 2006; pp. 503–510. [Google Scholar]
- Syed, R.N.; Mansha, N.; Khaskheli, M.A.; Khanzada, M.A.; Lodhi, A.M. Chemical control of stem end rot of mango caused by Lasiodiplodia theobromae. Pak. J. Phytopathol. 2014, 26, 201–206. [Google Scholar]
- Zhang, J. Comparative effects of postharvest fungicides for diplodia stem-end rot control of Florida citrus. Proc. Fla. State Hortic. Soc. 2012, 125, 243–247. [Google Scholar]
- Cerioni, L.; Sepulveda, M.; Rubio-Ames, Z.; Volentini, S.I.; Rodríguez-Montelongo, L.; Smilanick, J.; Ramallo, J.; Rapisarda, V.A. Control of lemon postharvest diseases by low-toxicity salts combined with hydrogen peroxide and heat. Postharvest Biol. Technol. 2013, 83, 17–21. [Google Scholar] [CrossRef]
- Cerioni, L.; Bennasar, P.B.; Lazarte, D.; Sepulveda, M.; Smilanick, J.L.; Ramallo, J.; Rapisarda, V.A. Conventional and reduced-risk fungicides to control postharvest Diplodia and Phomopsis stem-end rot on lemons. Sci. Hortic. 2017, 225, 783–787. [Google Scholar] [CrossRef]
- Wisniewski, M.; Droby, S.; Norelli, J.; Liu, J.; Schena, L. Alternative management technologies for postharvest disease control: The journey from simplicity to complexity. Postharvest Biol. Technol. 2016, 122, 3–10. [Google Scholar] [CrossRef]
- Droby, S.; Wisniewski, M.; Teixidó, N.; Spadaro, D.; Jijakli, M.H. The science, development, and commercialization of postharvest biocontrol products. Postharvest Biol. Technol. 2016, 122, 22–29. [Google Scholar] [CrossRef]
- Sharma, R.R.; Singh, D.; Singh, R. Biological control of postharvest diseases of fruits and vegetables by microbial antagonists: A review. Biol. Control 2009, 50, 205–221. [Google Scholar] [CrossRef]
- Korsten, L.; De Villiers, E.; Wehner, F.; Kotzé, J. Field sprays of Bacillus subtilis and fungicides for control of preharvest fruit diseases of avocado in South Africa. Plant Dis. 1997, 81, 455–459. [Google Scholar] [CrossRef]
- Demoz, B.T.; Korsten, L. Bacillus subtilis attachment, colonization, and survival on avocado flowers and its mode of action on stem-end rot pathogens. Biol. Control 2006, 37, 68–74. [Google Scholar] [CrossRef]
- Govender, V.; Korsten, L.; Sivakumar, D. Semi-commercial evaluation of bacillus licheniformis to control mango postharvest diseases in South Africa. Postharvest Biol. Technol. 2005, 38, 57–65. [Google Scholar] [CrossRef]
- De Costa, D.; Erabadupitiya, H. An integrated method to control postharvest diseases of banana using a member of the Burkholderia cepacia complex. Postharvest Biol. Technol. 2005, 36, 31–39. [Google Scholar] [CrossRef]
- Lassois, L.; de Bellaire, L.D.L.; Jijakli, M.H. Biological control of crown rot of bananas with Pichia anomala strain K and Candida oleophila strain O. Biol. Control 2008, 45, 410–418. [Google Scholar] [CrossRef]
- Sivakumar, D.; Wijeratnam, R.W.; Wijesundera, R.; Marikar, F.; Abeyesekere, M. Antagonistic effect of Trichoderma harzianum on postharvest pathogens of rambutan (Nephelium lappaceum). Phytoparasitica 2000, 28, 240. [Google Scholar] [CrossRef]
- Kota, V.; Kulkarni, S.; Hegde, Y. Postharvest diseases of mango and their biological management. J. Plant Dis. Sci. 2006, 1, 186–188. [Google Scholar]
- Abbaszadeh, S.; Sharifzadeh, A.; Shokri, H.; Khosravi, A.; Abbaszadeh, A. Antifungal efficacy of thymol, carvacrol, eugenol and menthol as alternative agents to control the growth of food-relevant fungi. J. Mycol. Méd./J. Med. Mycol. 2014, 24, e51–e56. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.; Tao, N.; Jia, L. Antifungal activity of citral, octanal and α-terpineol against Geotrichum citri-aurantii. Food Control 2014, 37, 277–283. [Google Scholar] [CrossRef]
- Anusuya, P.; Nagaraj, R.; Janavi, G.J.; Subramanian, K.S.; Paliyath, G.; Subramanian, J. Pre-harvest sprays of hexanal formulation for extending retention and shelf-life of mango (Mangifera indica L.) fruits. Sci. Hortic. 2016, 211, 231–240. [Google Scholar] [CrossRef]
- Wuryatmo, E.; Able, A.; Ford, C.; Scott, E. Effect of volatile citral on the development of blue mould, green mould and sour rot on navel orange. Aust. Plant Pathol. 2014, 43, 403–411. [Google Scholar] [CrossRef]
- Fan, F.; Tao, N.; Jia, L.; He, X. Use of citral incorporated in postharvest wax of citrus fruit as a botanical fungicide against Penicillium digitatum. Postharvest Biol. Technol. 2014, 90, 52–55. [Google Scholar] [CrossRef]
- Sepiah, M. Effectiveness of hot water, hot benomyl and cooling on postharvest diseases of mango cv. Harumanis. ASEAN Food J. 1986, 2, 117–120. [Google Scholar]
- Patni, C.; Kolte, S. Effect of some botanicals in management of Alternaria blight of rapeseed-mustard. Ann. Plant Protect. Sci. 2006, 14, 151–156. [Google Scholar]
- Bhalodia, N.R.; Shukla, V. Antibacterial and antifungal activities from leaf extracts of Cassia fistula L.: An ethnomedicinal plant. J. Adv. Pharm. Technol. Res. 2011, 2, 104–109. [Google Scholar] [CrossRef] [PubMed]
- Alam, M.W.; Rehman, A.; Sahi, S.T.; Malik, A.U. Exploitation of natural products as an alternative strategy to control stem end rot disease of mango fruit in Pakistan. Pak. J. Agric. Sci. 2017, 54, 503–511. [Google Scholar]
- Ullah, S.F.; Hussain, Y.; Iram, S. Pathogenic characterization of Lasiodiplodia causing stem end rot of mango and its control using botanics. Pak. J. Bot. 2017, 49, 1605–1613. [Google Scholar]
- Perumal, A.B.; Sellamuthu, P.S.; Nambiar, R.B.; Sadiku, E.R. Antifungal activity of five different essential oils in vapour phase for the control of Colletotrichum gloeosporioides and Lasiodiplodia theobromae in vitro and on mango. Int. J. Food Sci. Technol. 2016, 51, 411–418. [Google Scholar] [CrossRef]
- Shao, X.; Cao, B.; Xu, F.; Xie, S.; Yu, D.; Wang, H. Effect of postharvest application of chitosan combined with clove oil against citrus green mold. Postharvest Biol. Technol. 2015, 99, 37–43. [Google Scholar] [CrossRef]
- Wu, Y.; Duan, X.; Jing, G.; Ouyang, Q.; Tao, N. Cinnamaldehyde inhibits the mycelial growth of geotrichum citri-aurantii and induces defense responses against sour rot in citrus fruit. Postharvest Biol. Technol. 2017, 129, 23–28. [Google Scholar] [CrossRef]
- Obianom, C.; Sivakumar, D. Differential response to combined prochloraz and thyme oil drench treatment in avocados against the control of anthracnose and stem-end rot. Phytoparasitica 2018, 46, 273–281. [Google Scholar] [CrossRef]
- Seethapathy, P.; Gurudevan, T.; Subramanian, K.S.; Kuppusamy, P. Bacterial antagonists and hexanal-induced systemic resistance of mango fruits against Lasiodiplodia theobromae causing stem-end rot. J. Plant Interact. 2016, 11, 158–166. [Google Scholar] [CrossRef]
- Stone, J.K.; Polishook, J.D.; White, J.F. Endophytic fungi. In Biodiversity of Fungi; Elsevier Academic Press: Burlington, MA, USA, 2004; pp. 241–270. [Google Scholar]
- Ayerst, G. The effects of moisture and temperature on growth and spore germination in some fungi. J. Stored Prod. Res. 1969, 5, 127–141. [Google Scholar] [CrossRef]
- Pesis, E.; Ackerman, M.; Ben-Arie, R.; Feygenberg, O.; Feng, X.; Apelbaum, A.; Goren, R.; Prusky, D. Ethylene involvement in chilling injury symptoms of avocado during cold storage. Postharvest Biol. Technol. 2002, 24, 171–181. [Google Scholar] [CrossRef]
- Imlay, J.A.; Linn, S. DNA damage and oxygen radical toxicity. Science 1988, 240, 1302–1309. [Google Scholar] [CrossRef] [PubMed]
- Farkas, J. Irradiation for better foods. Trends Food Sci. Technol. 2006, 17, 148–152. [Google Scholar] [CrossRef]
- Ladaniya, M.; Singh, S.; Wadhawan, A. Response of ‘nagpur’ mandarin, ‘mosambi’ sweet orange and ‘kagzi’ acid lime to gamma radiation. Radiat. Phys. Chem. 2003, 67, 665–675. [Google Scholar] [CrossRef]
- Spalding, D.; Reeder, W. Decay and acceptability of mangos treated with combinations of hot water, imazalil, and gamma radiation. Plant Dis. 1986, 70, 1149–1151. [Google Scholar] [CrossRef]
- Rashid, M.; Grout, B.W.W.; Continella, A.; Mahmud, T. Low-dose gamma irradiation following hot water immersion of papaya (Carica papaya Linn.) fruits provides additional control of postharvest fungal infection to extend shelf life. Radiat. Phys. Chem. 2015, 110, 77–81. [Google Scholar] [CrossRef]
- Stevens, C.; Wilson, C.; Lu, J.; Khan, V.; Chalutz, E.; Droby, S.; Kabwe, M.; Haung, Z.; Adeyeye, O.; Pusey, L. Plant hormesis induced by ultraviolet light-c for controlling postharvest diseases of tree fruits. Crop Protect. 1996, 15, 129–134. [Google Scholar] [CrossRef]
- Stevens, C.; Khan, V.; Wilson, C.; Lu, J.; Chalutz, E.; Droby, S. The effect of fruit orientation of postharvest commodities following low dose ultraviolet light-c treatment on host induced resistance to decay. Crop Protect. 2005, 24, 756–759. [Google Scholar] [CrossRef]
- Romanazzi, G.; Sanzani, S.M.; Bi, Y.; Tian, S.; Martínez, P.G.; Alkan, N. Induced resistance to control postharvest decay of fruit and vegetables. Postharvest Biol. Technol. 2016, 122, 82–94. [Google Scholar] [CrossRef]
- Fallik, E. Prestorage hot water treatments (immersion, rinsing and brushing). Postharvest Biol. Technol. 2004, 32, 125–134. [Google Scholar] [CrossRef]
- Lurie, S. Postharvest heat treatments. Postharvest Biol. Technol. 1998, 14, 257–269. [Google Scholar] [CrossRef]
- Schirra, M.; D’hallewin, G.; Ben-Yehoshua, S.; Fallik, E. Host–pathogen interactions modulated by heat treatment. Postharvest Biol. Technol. 2000, 21, 71–85. [Google Scholar] [CrossRef]
- Couey, H.; Alvarez, A.; Nelson, M. Comparison of hot-water spray and immersion treatments for control of postharvest decay of papaya [stem-end rots and anthracnose]. Plant Dis. 1984, 68, 436–437. [Google Scholar] [CrossRef]
- Alvindia, D.G.; Acda, M.A. Revisiting the efficacy of hot water treatment in managing anthracnose and stem-end rot diseases of mango cv. ‘Carabao’. Crop Protect. 2015, 67, 96–101. [Google Scholar] [CrossRef]
- Coates, L.; Johnson, G.; Cooke, A. Postharvest disease control in mangoes using high humidity hot air and fungicide treatments. Ann. Appl. Biol. 1993, 123, 441–448. [Google Scholar] [CrossRef]
- Johnson, G.; Muirhead, I.; Rappel, L. Mango post-harvest disease control: A review of research in Australia, Malaysia and Thailand. ASEAN Food J. 1989, 4, 139–141. [Google Scholar]
- Johnson, G.; Sangchote, S.; Cooke, A. Control of stem end rot (Dothiorella dominicana) and other postharvest diseases of mangoes (cv. Kensington pride) during short-and long-term storage. Trop. Agric. Trinidad Tobago 1990, 67, 183–187. [Google Scholar]
- Johnson, G.; Hofman, P. Postharvest Technology and Quarantine Treatments; CAB International: Wallingford, UK, 2009. [Google Scholar]
- Prusky, D.; Fuchs, Y.; Kobiler, I.; Roth, I.; Weksler, A.; Shalom, Y.; Fallik, E.; Zauberman, G.; Pesis, E.; Akerman, M. Effect of hot water brushing, prochloraz treatment and waxing on the incidence of black spot decay caused by Alternaria alternata in mango fruits. Postharvest Biol. Technol. 1999, 15, 165–174. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Treatment | SER-Causing Target Pathogen | Fruit Host | References | |
|---|---|---|---|---|
| Preharvest chemical control | Benlate | Lasiodiplodia theobromae | Citrus | [47] |
| Copper-based chemicals | Lasiodiplodia spp. | Mango | [49] | |
| Fluopyram + pyrimethanil or fludioxonil + cyprodinil | Lasiodiplodia theobromae | Mango | [4] | |
| Postharvest chemical control | Benzimidazole: benomyl or thiabendazole | Lasiodiplodia theobromae | Mango | [53] |
| Prochloraz and fludioxonil | Lasiodiplodia theobromae | Mango | [56] | |
| Tebuconazole + trifloxystrobin, azoxystrobin or carbendazim | Lasiodiplodia theobromae | Mango | [57] | |
| Thiabendazole, imazalil and fludioxonil | Diplodia natalensis | Mango | [58] | |
| Salts: potassium sorbate and potassium phosphite | Diaporthe citri, Lasiodiplodia theobromae | Lemon | [59] | |
| Thiabendazole | Lasiodiplodia theobromae | Lemon | [60] | |
| Preharvest biological control | Bacillus subtilis | Lasiodiplodia theobromae | Avocado | [64,65] |
| Bacillus subtilis | Lasiodiplodia theobromae | Mango | [4] | |
| Postharvest biological control | Bacillus licheniformis | Botryosphaeria spp. and L. theobromae | Mango | [66] |
| Trichoderma harzianum | Lasiodiplodia theobromae | Rambutan | [69] | |
| Trichoderma viride | Lasiodiplodia theobromae | Mango | [70] | |
| Plant extracts | Thyme oil vapors | Lasiodiplodia theobromae and Colletotrichum gloeosporioides | Mango | [81] |
| Combined prochloraz and thyme oil | Lasiodiplodia theobromae and Colletotrichum gloeosporioides | Avocado | [84] | |
| Moringa oleifera, Syzygium aromaticum and Cinnamomum zeylanicum | Lasiodiplodia theobromae, Phomopsis mangiferae, Colletotrichum gloeosporioides | Mango | [79] | |
| Fruit Ripening inhibition | 1-MCP | Lasiodiplodia theobromae | Jujube fruit | [31,32] |
| 1-MCP | Lasiodiplodia theobromae | Avocado | [33] | |
| 2,4-D | Phomopsis spp. and Lasiodiplodia spp. | Citrus and mango | [27,28] | |
| Agrotechnical methods | Harvesting with short stems (pedicel) | Lasiodiplodia theobromae | Mango | [40,41] |
| Pruning, exposure to sunlight | Lasiodiplodia theobromae | Mango | [6] | |
| Physical treatment | Hot-water or combined hot water and gamma irradiation | Lasiodiplodia theobromae | Papya and Mango | [92,93,101] |
| Combined | Benomyl dip in hot water | Dothiorella dominicana and Lasiodiplodia theobromae | Mango | [102,103,104] |
| Combined HWB along with prochloraz followed by 2,4-D | Phomopsis spp. and Lasiodiplodia spp. | Mango | [28] | |
| Hot-water treatment with benomyl followed by a prochloraz | Dothiorella dominicana | Mango | [104] | |
| Combined Bacillus subtilis and hexanal | Lasiodiplodia theobromae | Mango | [85] | |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Galsurker, O.; Diskin, S.; Maurer, D.; Feygenberg, O.; Alkan, N. Fruit Stem-End Rot. Horticulturae 2018, 4, 50. https://doi.org/10.3390/horticulturae4040050
Galsurker O, Diskin S, Maurer D, Feygenberg O, Alkan N. Fruit Stem-End Rot. Horticulturae. 2018; 4(4):50. https://doi.org/10.3390/horticulturae4040050
Chicago/Turabian StyleGalsurker, Ortal, Sonia Diskin, Dalia Maurer, Oleg Feygenberg, and Noam Alkan. 2018. "Fruit Stem-End Rot" Horticulturae 4, no. 4: 50. https://doi.org/10.3390/horticulturae4040050
APA StyleGalsurker, O., Diskin, S., Maurer, D., Feygenberg, O., & Alkan, N. (2018). Fruit Stem-End Rot. Horticulturae, 4(4), 50. https://doi.org/10.3390/horticulturae4040050