

In Vitro Micropropagation of Kale (Brassica oleracea var. sabellica L.)


Abstract

1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Seed Sterilisation Experiment
2.3. Micropropagation
2.3.1. Culture Media
2.3.2. Culture Conditions of Explants
2.3.3. Temperature and Light Conditions
2.3.4. Regeneration and Phenotypical Anomalies
2.4. Statistical Analysis
3. Results
3.1. Examination of Disinfection Methods
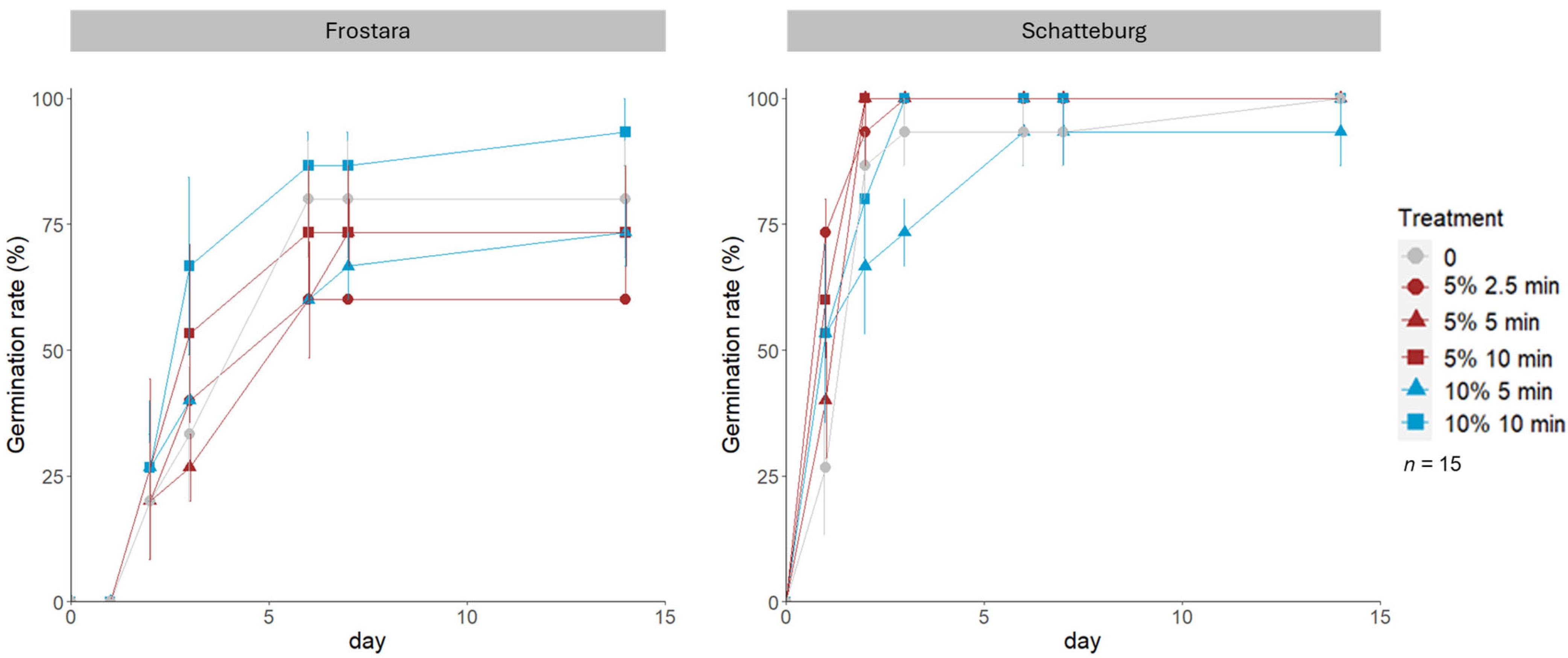
3.1.1. Germination Rate and Speed
3.1.2. Infection Rate and Speed
3.2. Examination of Micropropagation Protocols
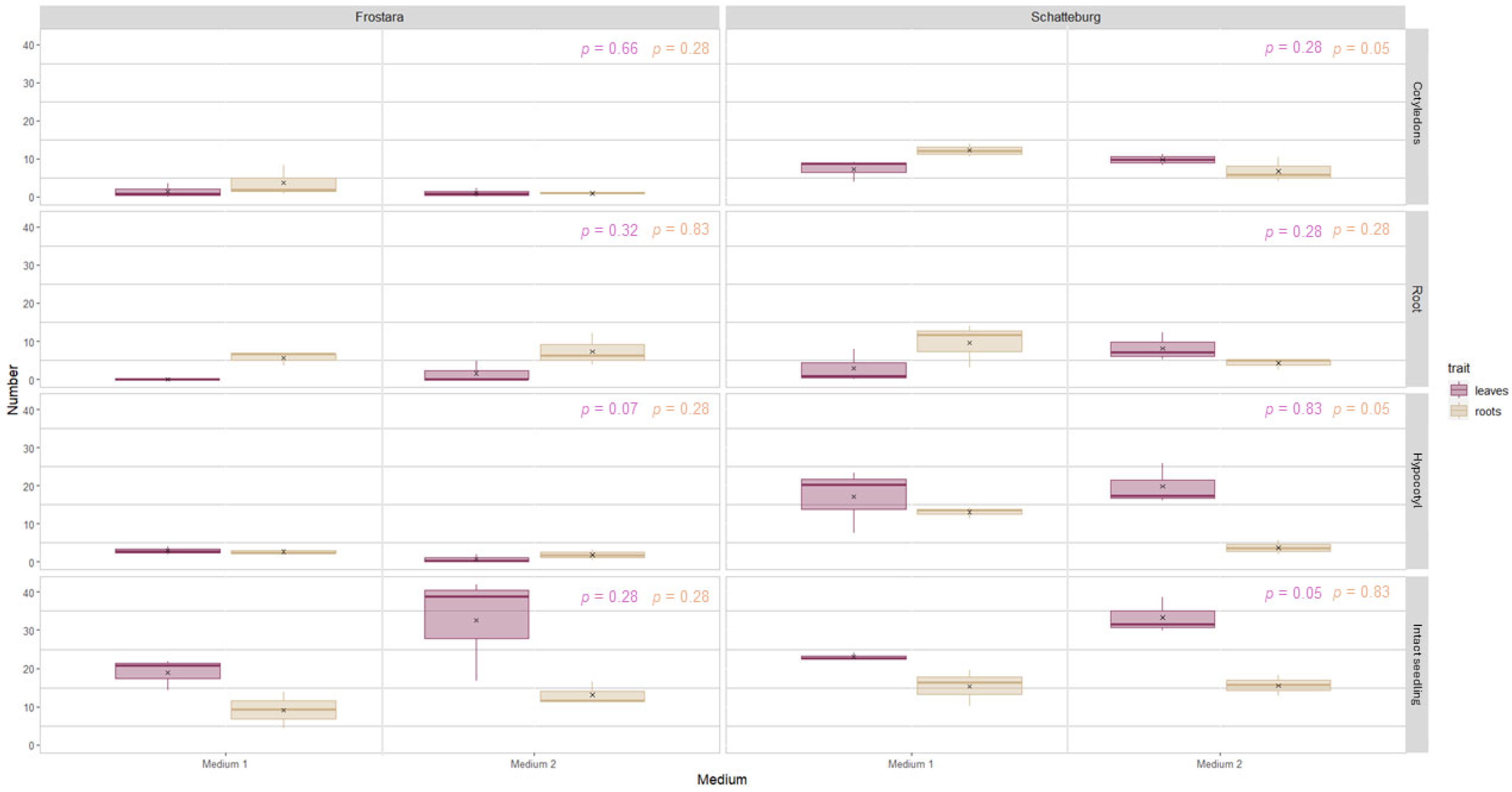
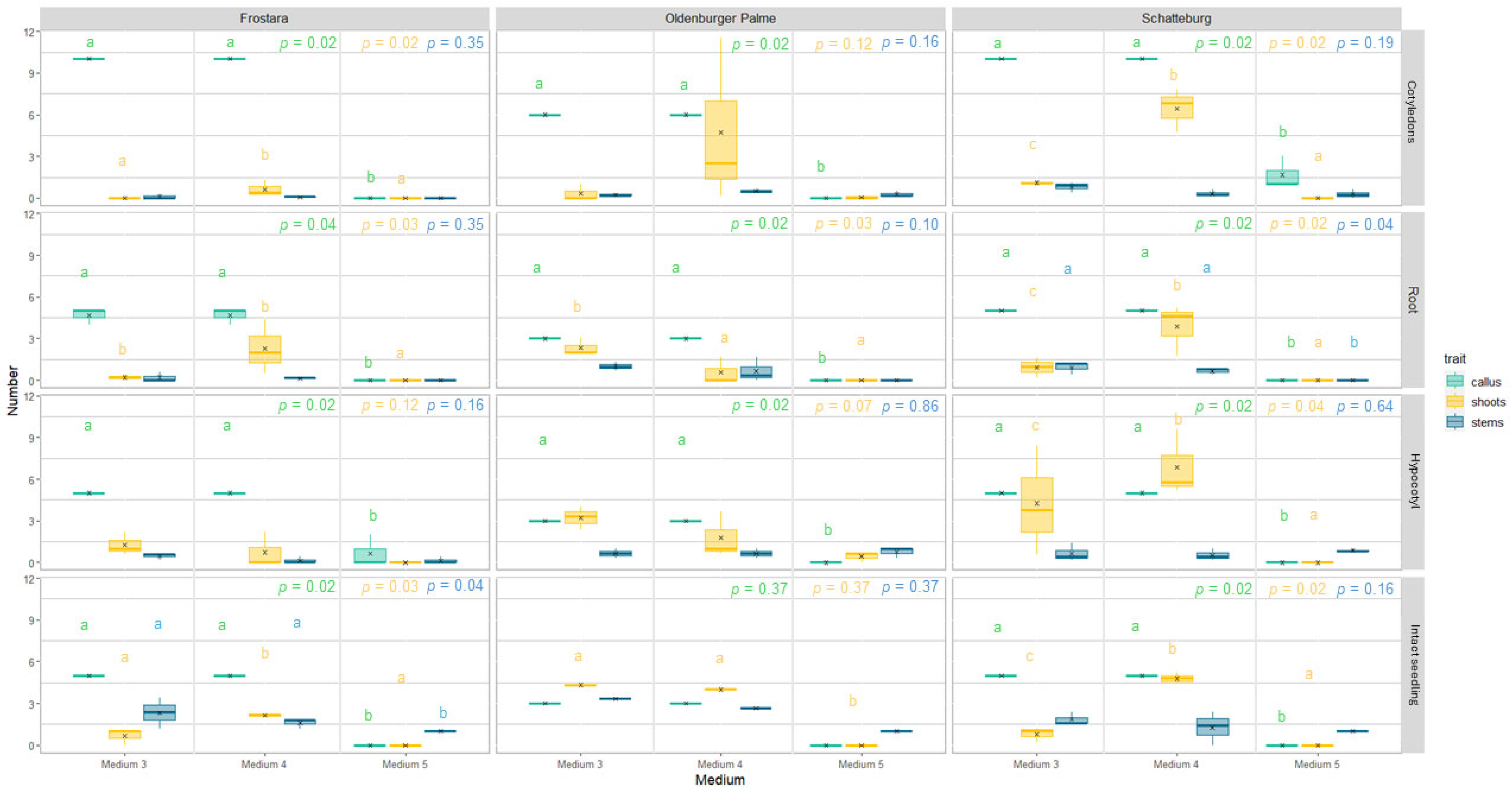
3.2.1. Regeneration of Leaves, Roots, Callus, Shoots, and Stems
3.2.2. Abnormal Morphological Development
4. Discussion
4.1. Disinfection Experiment
4.2. Micropropagation Experiment
4.2.1. Auxin to Cytokinin Ratio and Media Compositions
4.2.2. Impact of AgNO3
4.2.3. Performance of Different Explant Types
4.2.4. Phenotypic Anomalies
4.2.5. Implications of Our Findings
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| BAP | 6-benzylaminopurine |
| Co | cotyledon |
| D | days |
| GR | germination rate |
| GS | germination speed |
| Hy | hypocotyl |
| IR | infection rate |
| IS | infection speed |
| M1 | medium 1 |
| M2 | medium 2 |
| M3 | medium 3 |
| M4 | medium 4 |
| M5 | medium 5 |
| MS | Murashige and Skoog media |
| NA | not available |
| NAA | α-naphthaleneacetic acid |
| NGS | number of germinated seeds |
| NIS | number of infected seeds |
| NREx | number of explants showing regeneration |
| RE | regeneration efficiency |
| Ro | root |
| Se | intact seedling |
| TNEx | total number of explants |
| TS | total number of seeds |
References
- Cohen, J.E. Population and climate change. Proc. Am. Philos. Soc. 2010, 154, 158–182. [Google Scholar] [PubMed]
- Zhou, S.; Hu, Z.; Zhu, M.; Zhang, B.; Deng, L.; Pan, Y.; Chen, G. Biochemical and molecular analysis of a temperature-sensitive albino mutant in kale named “White Dove”. Plant Growth Regul. 2013, 71, 281–294. [Google Scholar] [CrossRef]
- Körber-Grohne, U. Nutzpflanzen in Deutschland: Kulturgeschichte und Biologie; Konrad Theiss Verlag GmbH: Stuttgart, Germany, 1987. [Google Scholar]
- Hagen, S.F.; Borge, G.I.A.; Solhaug, K.A.; Bengtsson, G.B. Effect of cold storage and harvest date on bioactive compounds in curly kale (Brassica oleracea L. var. acephala). Postharvest Biol. Technol. 2009, 51, 36–42. [Google Scholar] [CrossRef]
- Šamec, D.; Urlić, B.; Salopek-Sondi, B. Kale (Brassica oleracea var. acephala) as a superfood: Review of the scientific evidence behind the statement. Crit. Rev. Food Sci. Nutr. 2019, 59, 2411–2422. [Google Scholar] [CrossRef] [PubMed]
- Vinterhalter, D.; Sretenović-Rajičić, T.; Vinterhalter, B.; Ninković, S. Genetic transformation of Brassica oleracea vegetables. Transgenic Plant J. 2007, 1, 340–355. [Google Scholar]
- Zou, J.; Zou, X.; Gong, Z.; Song, G.; Ren, J.; Feng, H. Thidiazuron promoted microspore embryogenesis and plant regeneration in curly kale (Brassica oleracea L. convar. acephala var. sabellica). Horticulturae 2023, 9, 327. [Google Scholar] [CrossRef]
- Bhalla, P.L.; Singh, M.B. Agrobacterium-mediated transformation of Brassica napus and Brassica oleracea. Nat. Protoc. 2008, 3, 181–189. [Google Scholar] [CrossRef]
- Kumar, N.; Reddy, M. In vitro plant propagation: A review. J. For. Environ. Sci. 2011, 27, 61–72. [Google Scholar]
- George, E.F.; Hall, M.A.; De Klerk, G.-J. Plant Propagation by Tissue Culture: Volume 1. the Background; Springer Science & Business Media: Dordrecht, The Netherlands, 2007; Volume 1. [Google Scholar]
- Abdalla, N.; El-Ramady, H.; Seliem, M.K.; El-Mahrouk, M.E.; Taha, N.; Bayoumi, Y.; Shalaby, T.A.; Dobránszki, J. An academic and technical overview on plant micropropagation challenges. Horticulturae 2022, 8, 677. [Google Scholar] [CrossRef]
- Sivanandhan, G.; Moon, J.; Sung, C.; Bae, S.; Yang, Z.H.; Jeong, S.Y.; Choi, S.R.; Kim, S.-G.; Lim, Y.P. L-Cysteine increases the transformation efficiency of Chinese cabbage (Brassica rapa ssp. pekinensis). Front. Plant Sci. 2021, 12, 767140. [Google Scholar] [CrossRef]
- Zhao, Y.; Huang, S.; Zhang, Y.; Shi, F.; Liu, X.; Du, S.; Feng, H. Establishment of an efficient shoot regeneration system in vitro in Brassica rapa. Vitr. Cell. Dev. Biol.-Plant 2021, 57, 977–986. [Google Scholar] [CrossRef]
- Cardoza, V.; Stewart, C.N. Brassica biotechnology: Progress in cellular and molecular biology. Vitr. Cell. Dev. Biol.-Plant 2004, 40, 542–551. [Google Scholar] [CrossRef]
- Liu, W.; Yang, Y.; Liu, Q. Establishment of an efficient regeneration system using heading leaves of Chinese cabbage (Brassica rapa L.) and its application in genetic transformation. Hortic. Environ. Biotechnol. 2018, 59, 583–596. [Google Scholar] [CrossRef]
- Sparrow, P.; Townsend, T.; Morgan, C.; Dale, P.; Arthur, A.; Irwin, J. Genetic analysis of in vitro shoot regeneration from cotyledonary petioles of Brassica oleracea. Theor. Appl. Genet. 2004, 108, 1249–1255. [Google Scholar] [CrossRef] [PubMed]
- da Silva, J.A.T.; Winarto, B.; Dobránszki, J.; Zeng, S. Disinfection procedures for propagation of purple basil Ocimum basilicum L. ‘red rubin’ at different levels of salts, charcoal, sucrose and potassium iodine. Folia Hortic. 2015, 27, 3–14. [Google Scholar]
- Williams, J.; Pink, D.; Biddington, N. Effect of silver nitrate on long-term culture and regeneration of callus from Brassica oleracea var. gemmifera. Plant Cell Tissue Organ Cult. 1990, 21, 61–66. [Google Scholar] [CrossRef]
- Wright, K.M.; Marshall, J.; Wright, P.J.; Holden, N.J. Vacuolar localisation of anthocyanin pigmentation in microgreen cotyledons of basil, cabbage and mustard greens does not impact on colonisation by Shiga-toxigenic Escherichia coli O157:H7. Food Microbiol. 2023, 116, 104367. [Google Scholar] [CrossRef]
- Hundleby, P.; Chhetry, M. Brassica oleracea transformation. In Genetic Transformation in Crops; IntechOpen: London, UK, 2020. [Google Scholar]
- Kumar, P.; Srivastava, D. High frequency organogenesis in hypocotyl, cotyledon, leaf and petiole explants of broccoli (Brassica oleracea L. var. italica), an important vegetable crop. Physiol. Mol. Biol. Plants 2015, 21, 279–285. [Google Scholar] [CrossRef]
- Smith, N.A.; Bhalla, P.L. Comparison of shoot regeneration potential from seedling explants of Australian cauliflower (Brassica oleracea var. botrytis) varieties. Aust. J. Agric. Res. 1998, 49, 1261–1266. [Google Scholar] [CrossRef]
- Dai, X.; Shi, X.; Ye, Y.; Fu, Q.; Bao, M. High frequency plant regeneration from cotyledon and hypocotyl explants of ornamental kale. Biol. Plant. 2009, 53, 769–773. [Google Scholar] [CrossRef]
- Jaiswal, S.; Hammatt, N.; Bhojwani, S.; Cocking, E.; Davey, M. Plant regeneration from cotyledon protoplasts of Brassica carinata. Plant Cell Tissue Organ Cult. 1990, 22, 159–165. [Google Scholar] [CrossRef]
- Wright, K.M.; Holden, N.J. Quantification and colonisation dynamics of Escherichia coli O157:H7 inoculation of microgreens species and plant growth substrates. Int. J. Food Microbiol. 2018, 273, 1–10. [Google Scholar] [CrossRef]
- Bewley, J.D. Seed germination and dormancy. Plant Cell 1997, 9, 1055. [Google Scholar] [CrossRef] [PubMed]
- Timson, J. New method of recording germination data. Nature 1965, 207, 216–217. [Google Scholar] [CrossRef]
- Khan, M.A.; Ungar, I.A. The effect of salinity and temperature on the germination of polymorphic seeds and growth of Atriplex triangularis Willd. Am. J. Bot. 1984, 71, 481–489. [Google Scholar] [CrossRef]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bio assays with tobacco tissue cultures. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Farooq, N.; Nawaz, M.A.; Mukhtar, Z.; Ali, I.; Hundleby, P.; Ahmad, N. Investigating the in vitro regeneration potential of commercial cultivars of Brassica. Plants 2019, 8, 558. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2022. [Google Scholar]
- Kruskal, W.H.; Wallis, W.A. Use of ranks in one-criterion variance analysis. J. Am. Stat. Assoc. 1952, 47, 583–621. [Google Scholar] [CrossRef]
- Wilcoxon, F. Individual comparisons by ranking methods. Biom. Bull. 1992, 1, 80–83. [Google Scholar] [CrossRef]
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis, 1st ed.; Springer: New York, NY, USA, 2016. [Google Scholar]
- Wickham, H.; Averick, M.; Bryan, J.; Chang, W.; McGowan, L.D.A.; François, R.; Grolemund, G.; Hayes, A.; Henry, L.; Hester, J. Welcome to the Tidyverse. J. Open Source Softw. 2019, 4, 1686. [Google Scholar] [CrossRef]
- FC, M.; Davis, T.; ggplot2 authors. ggpattern: ‘ggplot2’ Pattern Geoms. Available online: https://github.com/coolbutuseless/ggpattern (accessed on 1 September 2023).
- Luo, C.; Bai, S. Versatile Pie Charts, Ring Charts, Bar Charts and Box Plots using Patterns, Colors and Images; R Package Version 1.0.0. Available online: https://github.com/cran/patternplot (accessed on 1 September 2024).
- van den Brand, T. ggh4x: Hacks for ‘ggplot2’, R Package Version 0.2.3. Available online: https://github.com/teunbrand/ggh4x (accessed on 18 September 2023).
- Fox, J.; Weisberg, S. An R Companion to Applied Regression, 3rd ed.; Sage: Thousand Oaks, CA, USA, 2019. [Google Scholar]
- Revelle, W. Psych: Procedures for Psychological, Psychometric, and Personality Research; R Package Version 2.4.6; Northwestern University: Evanston, IL, USA, 2020; Volume 2. [Google Scholar]
- Kassambara, A. ggpubr: ‘ggplot2’ Based Publication Ready Plots; R Package Version 0.6.0. Available online: https://rpkgs.datanovia.com/ggpubr/ (accessed on 19 September 2024).
- Wickham, H. Reshaping data with the reshape package. J. Stat. Softw. 2007, 21, 1–20. [Google Scholar] [CrossRef]
- Rudis, B. Hrbrthemes: Additional Themes, Theme Components and Utilities for ‘ggplot2’; R Package Version 0.8.7; R Foundation for Statistical Computing: Vienna, Austria, 2024. [Google Scholar]
- Leopold, A.C.; Vertucci, C.W. Moisture as a regulator of physiological reaction in seeds. Seed Moisture 1989, 14, 51–67. [Google Scholar]
- Taylor, A.; Paine, D.; Paine, C. Sinapine leakage from Brassica seeds. J. Am. Soc. Hortic. Sci. 1993, 118, 546–550. [Google Scholar] [CrossRef]
- Duke, S.H.; Kakefuda, G.; Harvey, T.M. Differential leakage of intracellular substances from imbibing soybean seeds. Plant Physiol. 1983, 72, 919–924. [Google Scholar] [CrossRef]
- Mahajan, G.; Mutti, N.K.; Jha, P.; Walsh, M.; Chauhan, B.S. Evaluation of dormancy breaking methods for enhanced germination in four biotypes of Brassica tournefortii. Sci. Rep. 2018, 8, 17103. [Google Scholar] [CrossRef] [PubMed]
- Carmello, C.R.; Cardoso, J.C. Effects of plant extracts and sodium hypochlorite on lettuce germination and inhibition of Cercospora longissima in vitro. Sci. Hortic. 2018, 234, 245–249. [Google Scholar] [CrossRef]
- Shu, F.; Zhao-xia, L.; Jin-zhi, C.; Geng-xiao, S.; Cui-ying, S.; Jin, C.; Guang, Y. Optimization of Seed Sterilization and Rooting Medium for Regeneration of Brassica napus. Fujian J. Agric. Sci. 2019, 34, 1371–1378. [Google Scholar]
- Gilbert, G.S.; Diaz, A.; Bregoff, H.A. Seed disinfestation practices to control seed-borne fungi and bacteria in home production of sprouts. Foods 2023, 12, 747. [Google Scholar] [CrossRef] [PubMed]
- Azis, N.; Hasbullah, N.; Rasad, F.; Daud, N.; Amin, M.; Lassim, M. Organogenesis and growth response of Brassica oleracea var. italica through in vitro culture. In Proceedings of the International Conference on Agricultural, Ecological and Medical Sciences, Sabah, Malaysia, 15–17 April 2015; pp. 7–8. [Google Scholar]
- Cui, J.; Li, M.; Qiu, L.; Cao, J.; Huang, L. Stable expression of exogenous imported sporamin in transgenic Chinese cabbage enhances resistance against insects. Plant Growth Regul. 2017, 81, 543–552. [Google Scholar] [CrossRef]
- Zhao, J.; Liang, A.; Zhu, Z.; Tang, Y. Regeneration of Chinese cabbage transgenic plants expressing antibacterial peptide gene and cowpea trypsin inhibitor gene. Euphytica 2006, 150, 397–406. [Google Scholar] [CrossRef]
- Skoog, F. Chemical regulation of growth and organ formation in plant tissue cultured in vitro. Proc. Symp. Soc. Exp. Biol. 1957, 11, 118–131. [Google Scholar]
- Jain, S.; Nidhi, N.; Kale, S.; Rathod, M.; Dhurve, L.; Mehara, H. A comprehensive review on role of bio-regulators in the growth and development of fruit and vegetable crops. Int. J. Environ. Clim. Chang. 2023, 13, 2879–2892. [Google Scholar] [CrossRef]
- Tahoori, F.; Majd, A.; Nejadsattari, T.; Ofoghi, H.; Iranbakhsh, A. Effects of silver nitrate (AgNO3) on growth and anatomical structure of vegetative organs of liquorice (Glycyrrhiza glabra L.) under in vitro condition. Plant Omics 2018, 11, 153–160. [Google Scholar] [CrossRef]
- Tang, G.; Zhou, W.; Li, H.; Mao, B.; He, Z.; Yoneyama, K. Medium, explant and genotype factors influencing shoot regeneration in oilseed Brassica spp. J. Agron. Crop Sci. 2003, 189, 351–358. [Google Scholar] [CrossRef]
- Naaz, A.; Shahzad, A.; Anis, M. Effect of adenine sulphate interaction on growth and development of shoot regeneration and inhibition of shoot tip necrosis under in vitro condition in adult Syzygium cumini L.—A multipurpose tree. Appl. Biochem. Biotechnol. 2014, 173, 90–102. [Google Scholar] [CrossRef]
- Doğru, S.; Murata, M.; Balkaya, A.; Kurtar, E.S. In vitro micropropagation of maintainer white head cabbage lines using cotyledon and hypocotyl explants. Black Sea J. Agric. 2022, 5, 7–8. [Google Scholar]
- Cristea, T.; Leonte, C.; Brezeanu, C.; Brezeanu, M.; Ambarus, S.; Calin, M.; Prisecaru, M. Effect of AgNO3 on androgenesis of Brassica oleracea L. anthers cultivated in vitro. Afr. J. Biotechnol. 2012, 11, 13788–13795. [Google Scholar]
- Zhang, P.; Phansiri, S.; Puonti-Kaerlas, J. Improvement of cassava shoot organogenesis by the use of silver nitrate in vitro. Plant Cell Tissue Organ Cult. 2001, 67, 47–54. [Google Scholar] [CrossRef]
- Asgher, M.; Khan, M.I.R.; Anjum, N.A.; Verma, S.; Vyas, D.; Per, T.S.; Masood, A.; Khan, N.A. Ethylene and polyamines in counteracting heavy metal phytotoxicity: A crosstalk perspective. J. Plant Growth Regul. 2018, 37, 1050–1065. [Google Scholar] [CrossRef]
- Kumar, P.G.; Sivakumar, S.; Siva, G.; Vigneswaran, M.; Senthil Kumar, T.; Jayabalan, N. Silver nitrate promotes high-frequency multiple shoot regeneration in cotton (Gossypium hirsutum L.) by inhibiting ethylene production and phenolic secretion. Vitr. Cell. Dev. Biol.-Plant 2016, 52, 408–418. [Google Scholar] [CrossRef]
- Shyamali, S.; Hattori, K. Effect of polyamines and silver nitrate on the high frequency regeneration from cotyledon explants of bottle gourd (Lagenaria siceraria; sp. asiatica). Pak. J. Biol. Sci. 2007, 10, 1288–1293. [Google Scholar] [CrossRef] [PubMed]
- Mostafa, H.H.; Wang, H.; Song, J.; Li, X. Effects of genotypes and explants on garlic callus production and endogenous hormones. Sci. Rep. 2020, 10, 4867. [Google Scholar] [CrossRef]
- Ċosiċ, T.; Motyka, V.; Raspor, M.; Savić, J.; Cingel, A.; Vinterhalter, B.; Vinterhalter, D.; Trávníčková, A.; Dobrev, P.I.; Bohanec, B.; et al. In vitro shoot organogenesis and comparative analysis of endogenous phytohormones in kohlrabi (Brassica oleracea var. gongylodes): Effects of genotype, explant type and applied cytokinins. Plant Cell Tissue Organ Cult. 2015, 121, 741–760. [Google Scholar] [CrossRef]
- Christianson, M.; Warnick, D. Organogenesis in vitro as a developmental process. HortScience 1988, 23, 515–519. [Google Scholar] [CrossRef]
- Hoang, N.N.; Kitaya, Y.; Shibuya, T.; Endo, R. Effects of supporting materials in in vitro acclimatization stage on ex vitro growth of wasabi plants. Sci. Hortic. 2020, 261, 109042. [Google Scholar] [CrossRef]
- Sparrow, P.; Goldsack, C.; Østergaard, L. Transformation Technology in the Brassicaceae. In Genetics and Genomics of the Brassicaecea; Schmidt, R., Brncroft, I., Eds.; Springer: New York, NY, USA, 2010. [Google Scholar]
- José, D.P.; De Campos, J.M.S.; Viccini, L.F.; Alkimim, E.R.; de Oliveira Santos, M. Micropropagation and ploidy stability of Lippia lacunosa Mart. & Schauer: An endangered brazilian medicinal plant. Rev. De Agric. Neotrop. 2019, 6, 1–7. [Google Scholar]
- Gupta, S.K.; Khanuja, S. In vitro micropropagation of Lippia alba. Curr. Sci. 2001, 81, 206–210. [Google Scholar]
- Canonge, J.; Roby, C.; Hamon, C.; Potin, P.; Pfannschmidt, T.; Philippot, M. Occurrence of albinism during wheat androgenesis is correlated with repression of the key genes required for proper chloroplast biogenesis. Planta 2021, 254, 1–16. [Google Scholar] [CrossRef]
- Sreelekshmi, R.; Siril, E.A.; Muthukrishnan, S. Role of Biogenic Silver Nanoparticles on Hyperhydricity Reversion in Dianthus chinensis L. an In Vitro Model Culture. J. Plant Growth Regul. 2022, 41, 23–39. [Google Scholar] [CrossRef]
- Ha, N.T.M.; Manh Do, C.; Hoang, T.T.; Ngo, N.D.; Van Bui, L.; Nhut, D.T. The effect of cobalt and silver nanoparticles on overcoming leaf abscission and enhanced growth of rose (Rosa hybrida L. ‘Baby Love’) plantlets cultured in vitro. Plant Cell Tissue Organ Cult. (PCTOC) 2020, 141, 393–405. [Google Scholar] [CrossRef]
- Mahmoud, L.M.; Grosser, J.W.; Dutt, M. Silver compounds regulate leaf drop and improve in vitro regeneration from mature tissues of australian finger lime (Citrus australasica). Plant Cell Tissue Organ Cult. (PCTOC) 2020, 141, 455–464. [Google Scholar] [CrossRef]
- Isah, T. Adjustments to in vitro culture conditions and associated anomalies in plants. Acta Biol. Cracoviensia. Ser. Bot. 2015, 57, 9–28. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | NaClO (v/v) | Application Time (min) |
|---|---|---|
| a | 5% | 2.5 |
| b | 5% | 5 |
| c | 5% | 10 |
| d | 10% | 5 |
| e | 10% | 10 |
| f | control (sterile water) | rinsed three times |
| Reagents | Medium 1 1 | Medium 2 (Combination of the Media 1a–1c, Own Composition) | Medium 3 2 | Medium 4 2, 3 | Medium 5 Control | |||
|---|---|---|---|---|---|---|---|---|
| Medium 1a Callus Induction Medium (CIM) | Medium 1b Shoot Induction Medium (SIM) | Medium 1c Shoot Outgrowth Medium (SOM) | ||||||
| Silver nitrate (mg/L) | AgNO3 | 5.00 | 5.00 | - | 3.30 | - | 3 | - |
| Potassium iodide (mg/L) | KI | 0.75 | 0.75 | 0.75 | 0.75 | - | - | - |
| Adenine hemisulfate (mg/L) | C5H5N5•1/2H2SO4 | - | - | 40.00 | 13.30 | - | - | - |
| 6-benzylaminopurine (mg/L) | BAP | 0.75 | 3.00 | 0.001 | 1.25 | 2.00 | 2.00 | - |
| α-naphthaleneacetic acid (mg/L) | NAA | 0.20 | 0.20 | - | 0.20 | 0.50 | 0.50 | - |
| Gibberellic acid (mg/L) | GA3 | 0.01 | 0.01 | - | 0.07 | - | - | - |
| Polyvinylpyrrolidon (mg/L) | PVP 40.000 | - | - | 500.00 | 133.70 | - | - | - |
| Sucrose (g/L) | - | 20.00 | 20.00 | 20.00 | 20.00 | 20.00 | 20.00 | 20.00 |
| Gelrite (g/L) | - | 4.00 | 4.00 | 4.00 | 4.00 | 4.00 | 4.00 | 4.00 |
| pH value | - | 5.8 | 5.8 | 5.8 | 5.8 | 5.8 | 5.8 | 5.8 |
| Kale Variety | Media Tested | Explant Types Tested | Number of Media Boxes perExplant Type/Control | Number of Explants per Media Box |
|---|---|---|---|---|
| Schatteburg | 1–5 | all 4 | 3 | 5 (10 for cotyledons) |
| Frostara | 1–5 | all 4 | 3 | 5 (10 for cotyledons) |
| Oldenburger Palme | 3–5 | all 4 | 3 (1 for intact seedlings) | 3 (6 for cotyledons) |
| Week | Medium 1 | Medium 2 | Medium 3 | Medium 4 | Medium 5 |
|---|---|---|---|---|---|
| 1 | x (1a) | x | x | x | x |
| 2 | x (1b) | - | - | - | - |
| 3 | - | x | x | x | x |
| 4 | x (1b) | - | - | - | - |
| 5 | - | x | x | x | x |
| 6 | x (1c) | - | - | - | - |
| 7 | - | x | x | x | x |
| 8 | x (1c) | - | - | - | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Beyeler, M.; Albach, D.C. In Vitro Micropropagation of Kale (Brassica oleracea var. sabellica L.). Horticulturae 2025, 11, 767. https://doi.org/10.3390/horticulturae11070767
Beyeler M, Albach DC. In Vitro Micropropagation of Kale (Brassica oleracea var. sabellica L.). Horticulturae. 2025; 11(7):767. https://doi.org/10.3390/horticulturae11070767
Chicago/Turabian StyleBeyeler, Maike, and Dirk Carl Albach. 2025. "In Vitro Micropropagation of Kale (Brassica oleracea var. sabellica L.)" Horticulturae 11, no. 7: 767. https://doi.org/10.3390/horticulturae11070767
APA StyleBeyeler, M., & Albach, D. C. (2025). In Vitro Micropropagation of Kale (Brassica oleracea var. sabellica L.). Horticulturae, 11(7), 767. https://doi.org/10.3390/horticulturae11070767