
Green Synthesis of Zinc and Iron Nanoparticles Using Psidium guajava Leaf Extract Stimulates Cowpea Growth, Yield, and Tolerance to Saline Water Irrigation

 , , , , and
, , , , and
Abstract
1. Introduction
- Psidium guajava leaf extract (GLE) can be used as a stabilizer and reducing agent for zinc and iron oxide nanoparticle green synthesis.
- Foliar application of ZnNPs and FeNPs may enhance cowpea growth, yield, and yield parameters under saline conditions.
- Promotion of polyphenol and osmoprotectant levels in stressed plants as affected by ZnNP and FeNP foliar application.
2. Materials and Methods
2.1. Experimental Site, Plant Material, and Experimental Design
2.2. Saline Water Treatments
2.3. Collection and Preparation of Plant Extract
2.4. Phytochemical Screening and Estimation
2.5. Green Synthesis of Nanoparticles
2.6. Characterization of ZnO and Fe Nanoparticles
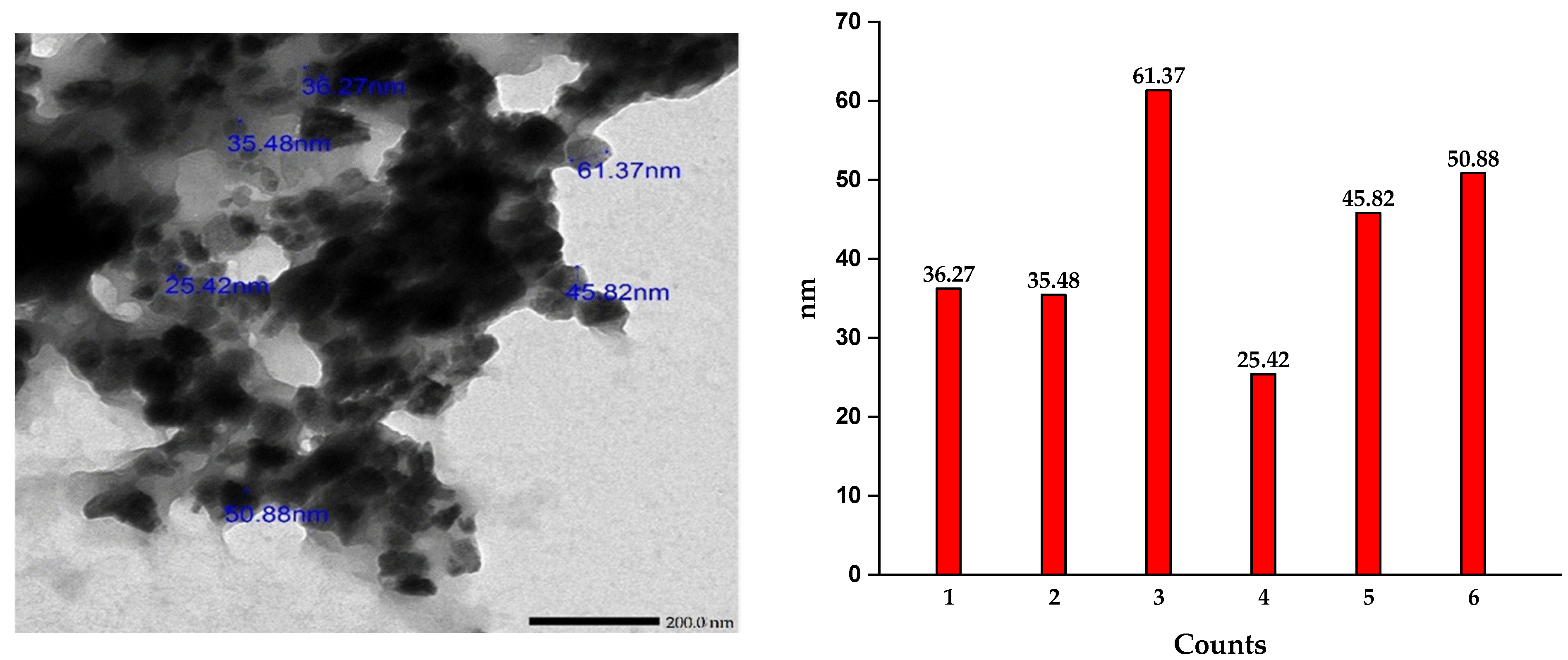
2.6.1. Transmission Electron Microscope Analysis (TEM)
2.6.2. Zeta Potential
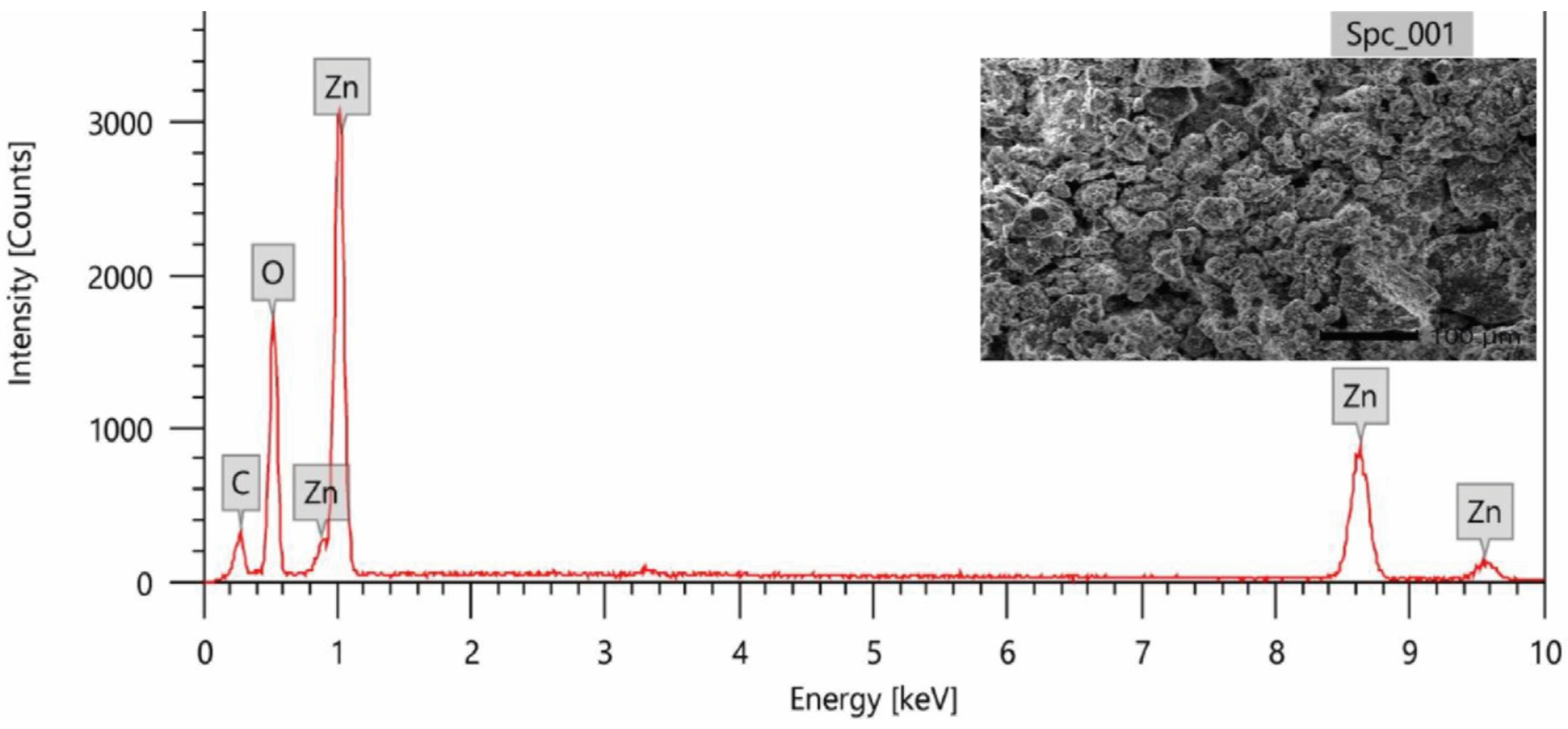
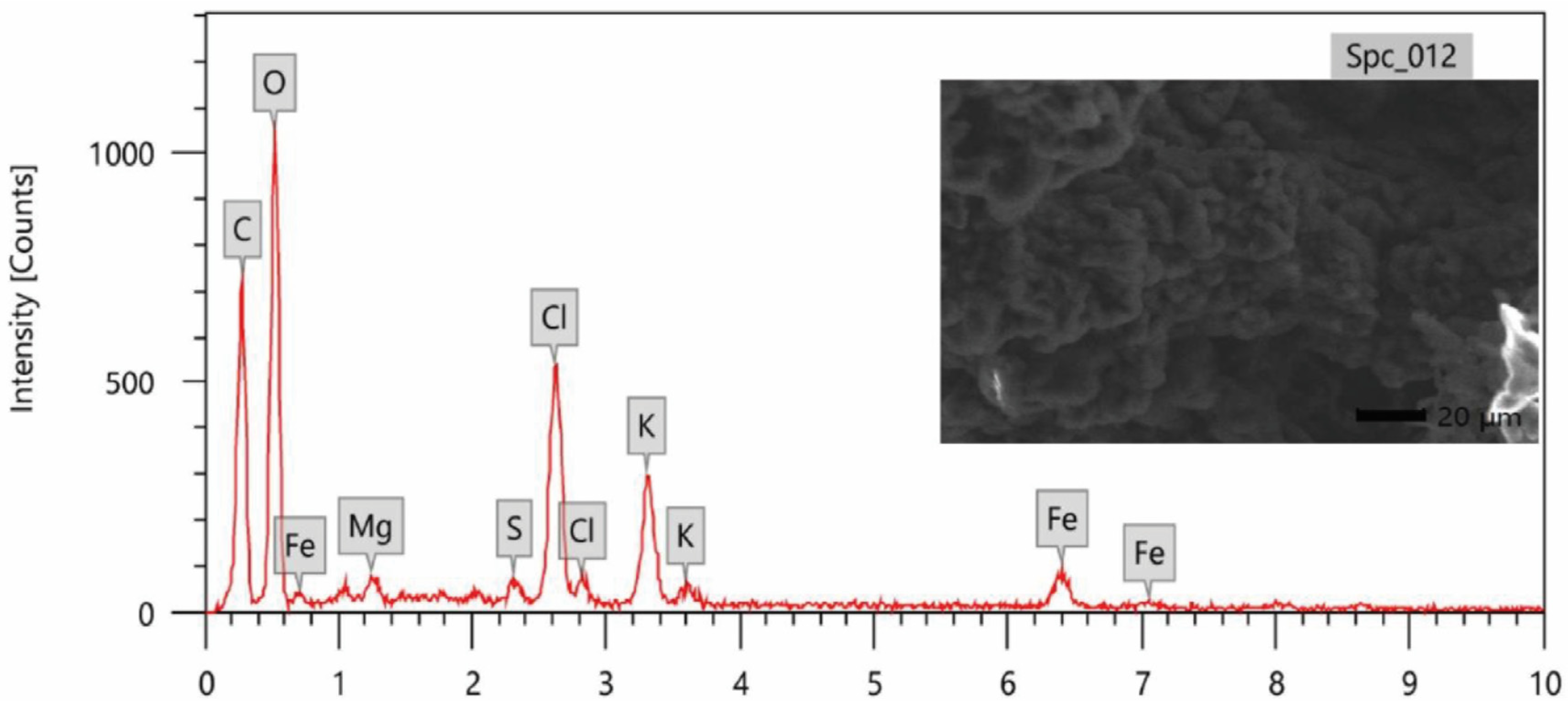
2.6.3. Energy Dispersive Analysis of X-rays (EDX)
2.7. ZnNP and FeNP Treatments
2.8. Leaf Traits and Yield Performance
2.9. Physiological and Biochemical Analysis
2.9.1. Photosynthetic Pigments
2.9.2. Proline Content
2.9.3. Peroxidase Activity Assay (POD)
2.9.4. Total Phenolic Content
2.9.5. Seed Analysis
2.10. Statistical Analysis
3. Results
3.1. Qualitative Detection of GLE
3.2. Characterization of ZnNPs and FeNPs
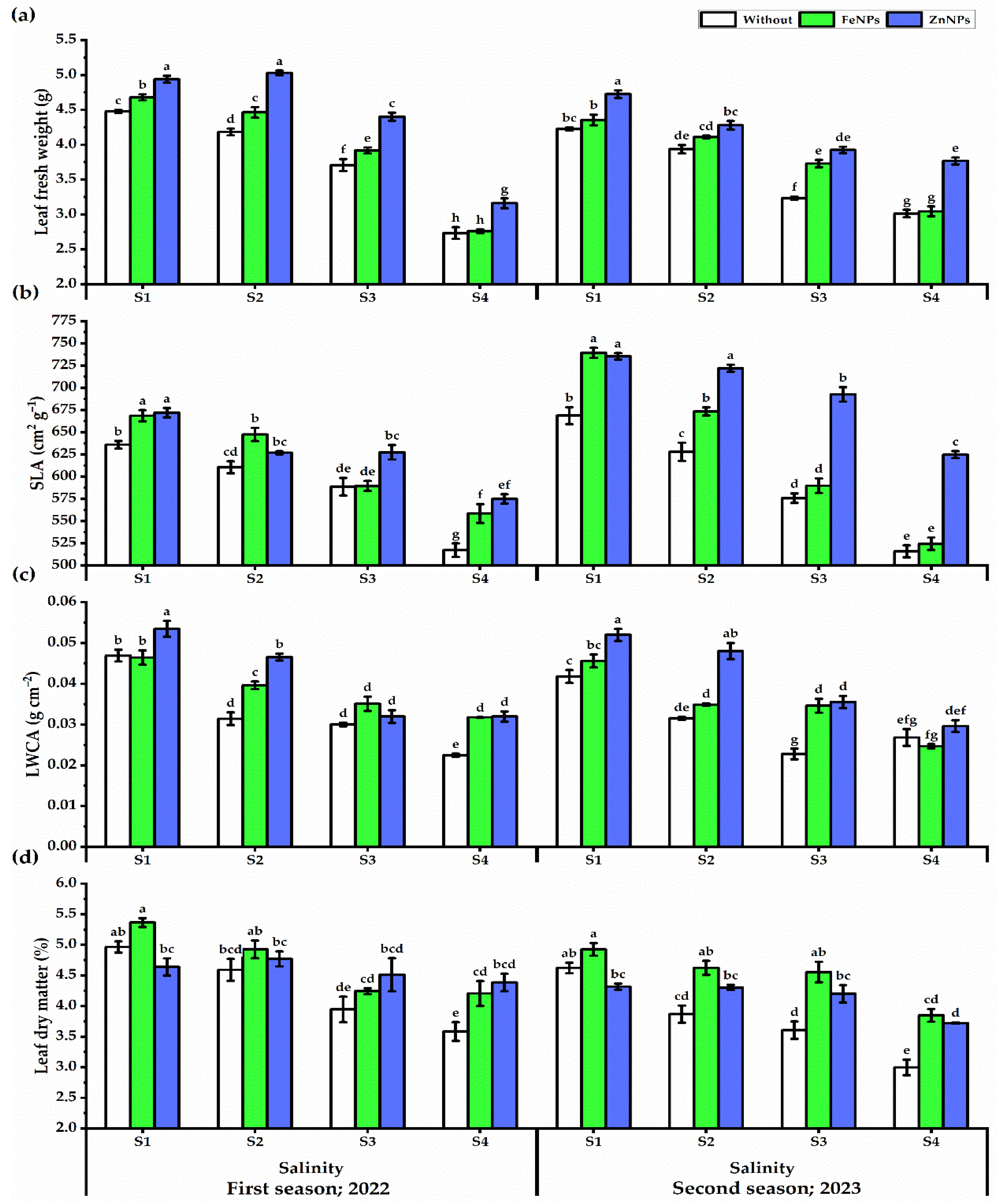
3.3. Leaf Traits
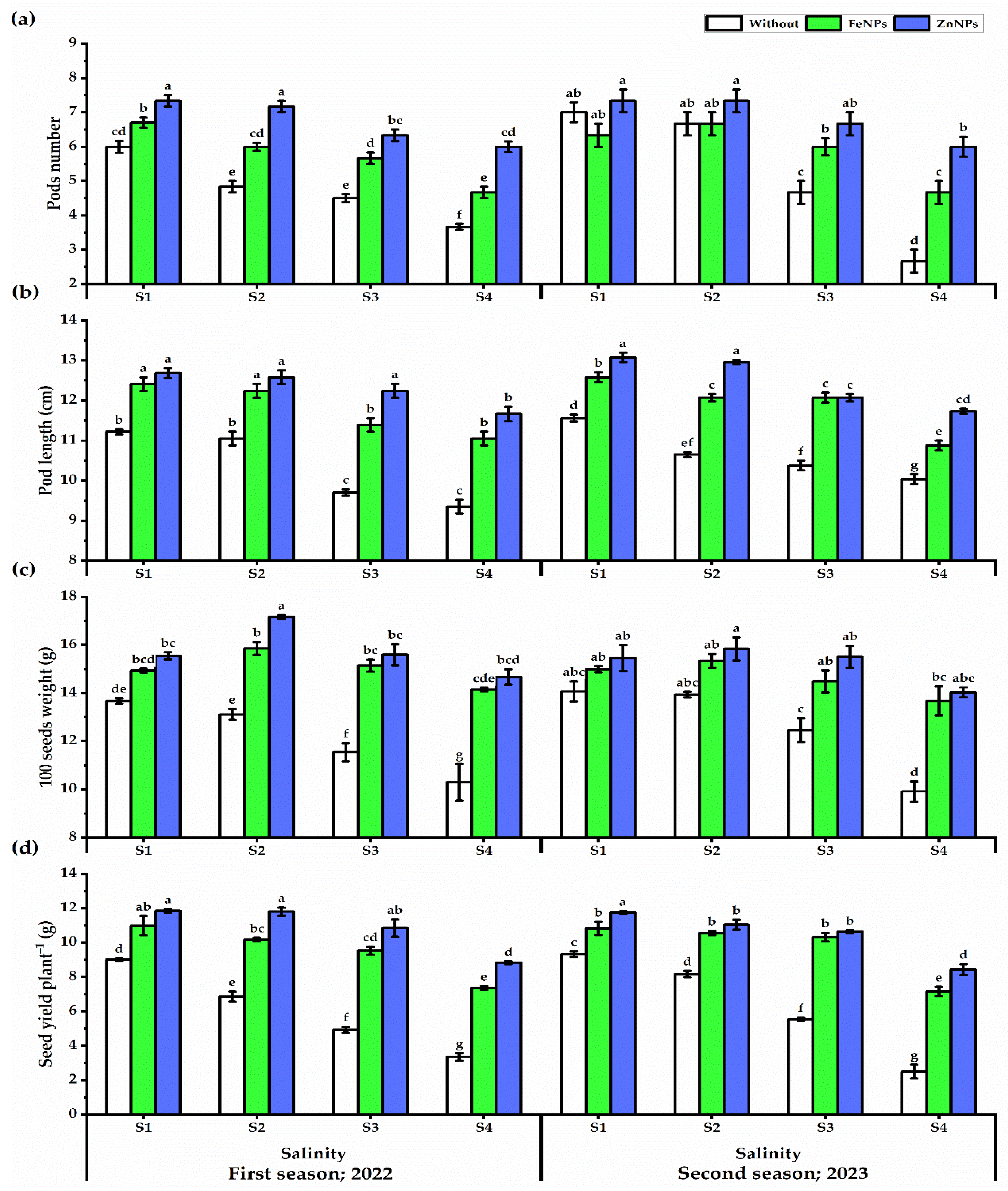
3.4. Yield Traits
3.5. Physiological and Biochemical Analysis
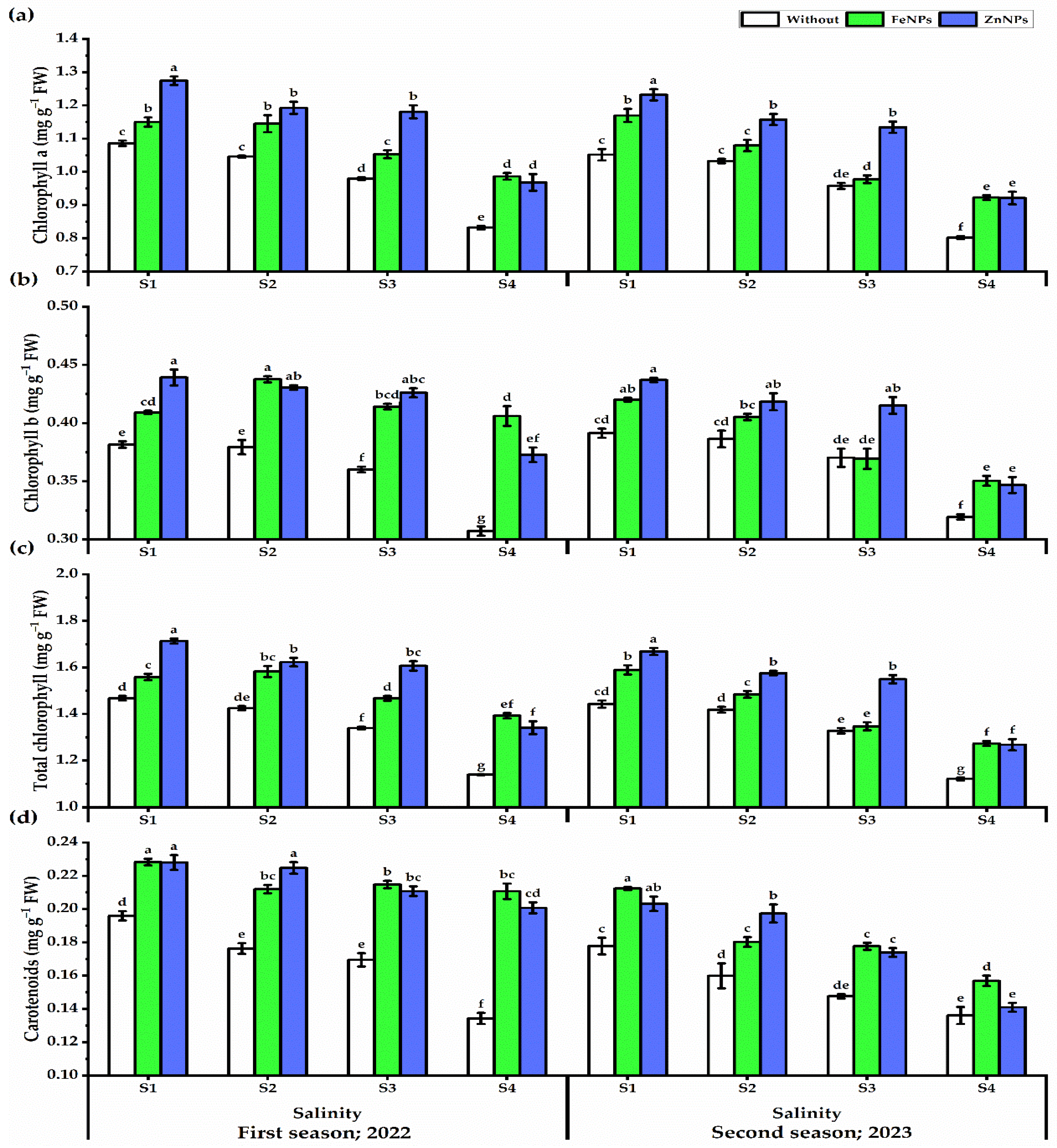
3.5.1. Photosynthetic Pigments
3.5.2. Proline Content
3.5.3. Peroxidase Activity
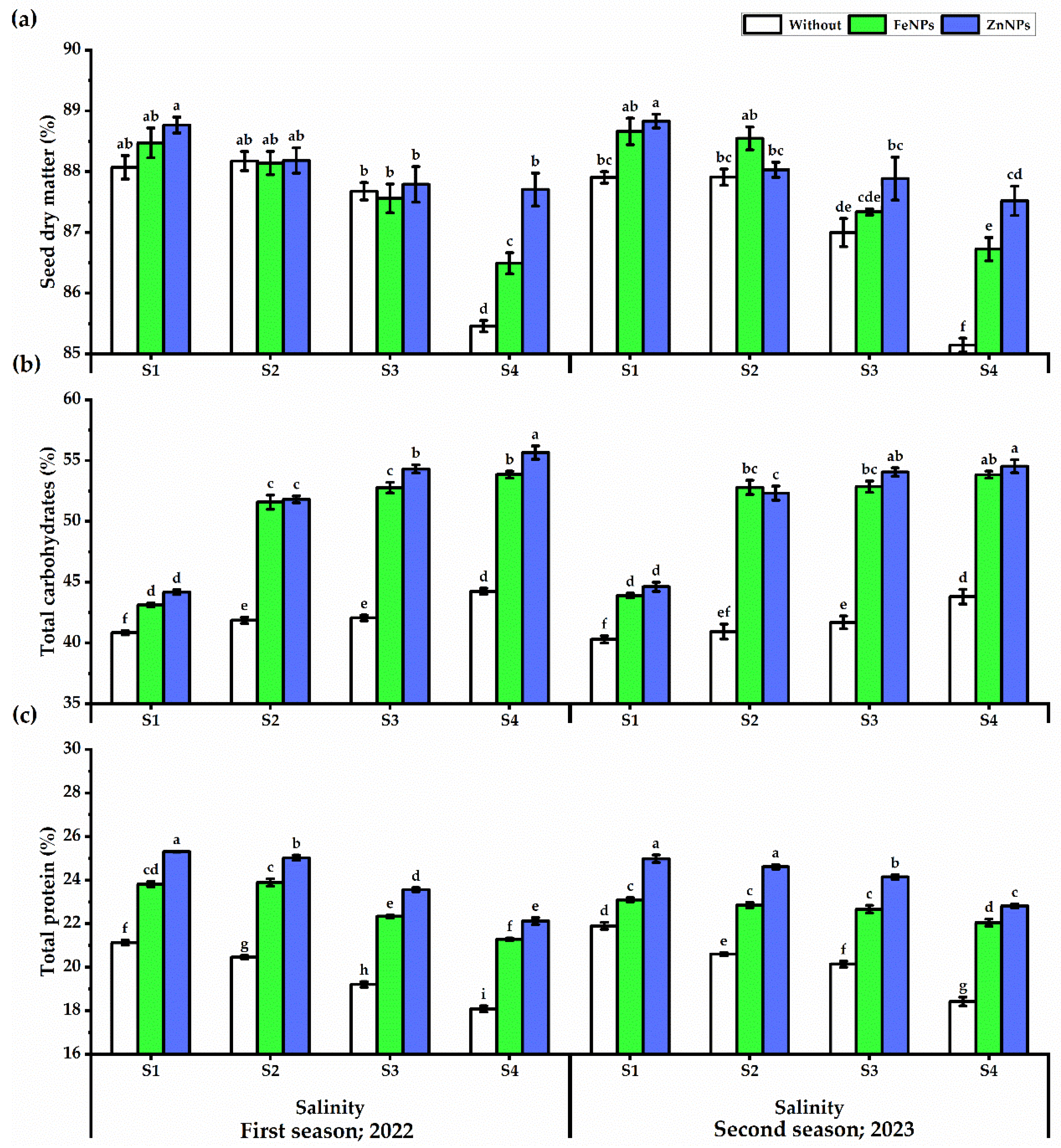
3.5.4. Total Phenols
3.5.5. Seed Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Brink, M.; Belay, G. Plant Resources of Tropical Africa: Cereals and Pulses; PROTA Foundation: Wageningen, The Netherlands; Blackhuys Publishers: Leiden, The Netherlands; CTA: Wageningen, The Netherlands, 2006. [Google Scholar]
- Affrifah, N.S.; Phillips, R.D.; Saalia, F.K. Cowpeas: Ntritional profile, processing methods and products—A review. Legume Sci. 2022, 4, e131. [Google Scholar] [CrossRef]
- Maas, E.V.; Hoffman, G.J. Crop salt tolerance-current assessment. J. Irrig. Drain. Div. 1977, 103, 115–134. [Google Scholar] [CrossRef]
- Hall, A.E.; Frate, C.A. Blackeye Bean Production in California; University of California, Division of Agriculture and Natural Resources: Los Angeles, CA, USA, 1996; 23p. [Google Scholar]
- Al-Hayany, E.A. Effect of salts stress and vitamin c on some growth characteristics and yield of cowpea Vigna sinensis. Plant Arch. 2020, 20, 2569–2572. [Google Scholar]
- Sheta, M.H.; Hasham, M.M.A.; Ghanem, K.Z.; Bayomy, H.M.; El-Sheshtawy, A.-N.A.; El-Serafy, R.S.; Naif, E. Screening of wheat genotypes for water stress tolerance using soil–water relationships and multivariate statistical approaches. Agronomy 2024, 14, 1029. [Google Scholar] [CrossRef]
- Ghanem, K.Z.; Hasham, M.M.A.; El-Sheshtawy, A.-N.A.; El-Serafy, R.S.; Sheta, M.H. Biochar stimulated actual evapotranspiration and wheat productivity under water deficit conditions in sandy soil based on non-weighing lysimeter. Plants 2022, 11, 3346. [Google Scholar] [CrossRef] [PubMed]
- Latif, M.; Bukhari, S.A.H.; Alrajhi, A.A.; Alotaibi, F.S.; Ahmad, M.; Shahzad, A.N.; Dewidar, A.Z.; Mattar, M.A. Inducing Drought Tolerance in Wheat through Exopolysaccharide-Producing Rhizobacteria. Agronomy 2022, 12, 1140. [Google Scholar] [CrossRef]
- Attia, M.A.E.H.; El-Enin, M.M.A.; Tahoun, A.M.A.; Abdelghany, F.I.M.; El-Serafy, R.S. Productivity of some barley cultivars asaffected by supplemental irrigation under rainfed conditions. Aust. J. Crop Sci. 2022, 16, 665–675. [Google Scholar] [CrossRef]
- Atteya, A.K.G.; El-Serafy, R.S.; El-Zabalawy, K.M.; Elhakem, A.; Genaidy, E.A.E. Exogenously supplemented proline and phenylalanine improve growth, productivity, and oil composition of salted moringa by up-regulating osmoprotectants and stimulating antioxidant machinery. Plants 2022, 11, 1553. [Google Scholar] [CrossRef]
- Youssef, S.M.; El-Serafy, R.S.; Ghanem, K.Z.; Elhakem, A.; Abdel Aal, A.A. Foliar spray or soil drench: Microalgae applicationimpacts on soil microbiology, morpho-physiological and biochemical responses, oil and fatty acid profiles of chia plants underalkaline stress. Biology 2022, 11, 1844. [Google Scholar] [CrossRef]
- Raza, A.; Tabassum, J.; Fakhar, A.Z.; Sharif, R.; Chen, H.; Zhang, C.; Ju, L.; Fotopoulos, V.; Siddique, K.; Singh, R.; et al. Smart reprograming of plants against salinity stress using modern biotechnological tools. Crit. Rev. Biotechnol. 2023, 43, 1035–1062. [Google Scholar] [CrossRef]
- Etesami, H.; Fatemi, H.; Muhammad Rizwan, M. Interactions of nanoparticles and salinity stress at physiological, biochemical and molecular levels in plants: A review. Ecotox. Environ. Safe. 2021, 225, 112769. [Google Scholar] [CrossRef] [PubMed]
- Powers, K.W.; Brown, S.C.; Krishna, V.B.; Wasdo, S.C.; Moudgil, B.M.; Roberts, S.M. Research strategies for safety evaluation of nanomaterials. Part 6. Characterization of nano scale particles for toxicological evaluation. Toxicol. Sci. 2006, 90, 296–303. [Google Scholar] [CrossRef] [PubMed]
- El-Enin, M.M.A.; Sheha, A.M.; El-Serafy, R.S.; Ali, O.A.M.; Saudy, H.S.; Shaaban, A. Foliage-sprayed nano-chitosan-loaded nitrogen boosts yield potentials, competitive ability, and profitability of intercropped maize-soybean. Int. J. Plant Prod. 2023, 17, 517–542. [Google Scholar] [CrossRef]
- Saleem, M.H.; Usman, K.; Rizwan, M.; Al Jabri, H.; Alsafran, M. Functions and strategies for enhancing zinc availability in plants for sustainable agriculture. Front. Plant Sci. 2022, 13, 1033092. [Google Scholar]
- Vojodi, M.L.; Kamran, R.V.; Hassanpouraghdam, M.B.; Pessarakli, M. Zinc sulfate foliar application affects some physiological characteristics and phenolic and essential oil contents of Lavandula stoechas L. under sodium chloride (NaCl) salinity conditions. Commun. Soil. Sci. Plant Anal. 2017, 48, 1860–1867. [Google Scholar] [CrossRef]
- Borm, P.; Klaessig, F.C.; Landry, T.D.; Moudgil, B.; Pauluhn, J.; Thomas, K.; Trottier, R.; Wood, S. Research strategies for safety evaluation of nanomaterials, part V: Role of dissolution in biological fate and effects of nanoscale particles. Toxicol. Sci. 2006, 90, 23–32. [Google Scholar] [CrossRef]
- Adil, M.; Bashir, S.; Bashir, S.; Aslam, Z.; Ahmad, N.; Younas, T.; Asghar, R.M.A.; Alkahtani, J.; Dwiningsih, Y.; Elshikh, M.S. Zinc oxide nanoparticles improved chlorophyll contents, physical parameters, and wheat yield under salt stress. Front. Plant Sci. 2022, 13, 932861. [Google Scholar] [CrossRef]
- Afshar, R.M.; Hadi, H.; Pirzad, A. Effect of nano-iron on the yield and yield component of cowpea (Vigna unguiculata) under end season water deficit. Intl. J. Agric. Res. Rev. 2013, 3, 27–34. [Google Scholar]
- Havlin, J.L.; Beaton, J.D.; Tisdale, S.L.; Nelson, W.L. Soil Fertility and Fertilizers-An Introduction to Nutrient Management, 6th ed.; Prentice Hall: Hoboken, NJ, USA, 1999. [Google Scholar]
- Singh, D.; Sillu, D.; Kumar, A.; Agnihotri, S. Dual nanozyme characteristics of iron oxide nanoparticles alleviate salinity stress and promote the growth of an agroforestry tree, Eucalyptus tereticornis Sm. Environ. Sci. Nano 2021, 8, 1308–1325. [Google Scholar] [CrossRef]
- Ashraf, M.A.; Rasheed, R.; Rizwan, M.; Hussain, I.; Aslam, R.; Qureshi, F.F.; Hafiza, B.S.; Bashir, R.; Ali, S. Effect of exogenous taurine on pea (Pisum sativum L.) plants under salinity and iron deficiency stress. Environ. Res. 2023, 223, 115448. [Google Scholar] [CrossRef]
- Bhardwaj, B.; Singh, P.; Kumar, A.; Kumar, S.; Budhwar, V. Eco-friendly greener synthesis of nanoparticles. Adv. Pharm. Bull. 2020, 10, 566–576. [Google Scholar] [CrossRef]
- Patil, S.P.; Rane, P.M. Psidium guajava leaves assisted green synthesis of metallic nanoparticles: A review. Beni-Suef Univ. J. Basic Appl. Sci. 2020, 9, 60. [Google Scholar] [CrossRef]
- Alharbi, F.N.; Abaker, Z.M.; Makawi, S.Z.A. Phytochemical substances—Mediated synthesis of zinc oxide nanoparticles (ZnO NPS). Inorganics 2023, 11, 328. [Google Scholar] [CrossRef]
- Abomuti, M.A.; Danish, E.Y.; Firoz, A.; Hasan, N.; Malik, M.A. Green synthesis of zinc oxide nanoparticles using Salvia officinalis leaf extract and their photocatalytic and antifungal activities. Biology 2021, 10, 1075. [Google Scholar] [CrossRef] [PubMed]
- El-Saadony, M.T.; Saad, A.M.; Soliman, S.M.; Salem, H.M.; Desoky, E.M.; Babalghith, A.O.; El-Tahan, A.M.; Ibrahim, O.M.; Ebrahim, A.A.M.; Abd El-Mageed, T.A.; et al. Role of nanoparticles in enhancing crop tolerance to abiotic stress: A Comprehensive Review. Front. Plant Sci. 2022, 13, 946717. [Google Scholar] [CrossRef]
- Ragab, S.M.; Turoop, L.; Runo, S.; Nyanjom, S. Nanoparticle treatments based on zinc oxide and Moringa oleifera leaf extracts alleviate salinity stress in Faba bean (Vicia faba L.). J. Agric. Chem. Environ. 2022, 11, 42–65. [Google Scholar]
- Rehman, M.Z.; Anayatullah, S.; Irfan, E.; Hussain, S.M.; Rizwan, M.; Sohail, M.I.; Jafir, M.; Ahmad, T.; Usman, M.; Alharby, F. Nanoparticles assisted regulation of oxidative stress and antioxidant enzyme system in plants under salt stress: A review. Chemosphere 2023, 314, 137649. [Google Scholar] [CrossRef] [PubMed]
- Page, A.L.; Miller, R.H.; Keeney, D.R. Methods of Soil Analysis. Part 2. Chemical and Microbiological Properties, 2nd ed.; American Society of Agronomy Inc. Publisher: Madison, WI, USA, 1982. [Google Scholar]
- Klute, A. Methods of Soil Analysis. Part 1. Physical and Mineralogical Methods, 2nd ed.; Monograph No. 9; American Society of Agronomy: Madison, WI, USA, 1986. [Google Scholar]
- Ramya, V.; Kalaiselvi, V.; Kannan, S.K.; Shkir, M.; Ghramh, H.A.; Ahmad, Z.; Vidhya, N. Facile synthesis and characterization of zinc oxide nanoparticles using Psidium guajava leaf extract and their antibacterial applications. Arab. J. Sci. Eng. 2022, 47, 909–918. [Google Scholar] [CrossRef]
- Harborne, A.J. Phytochemical Methods a Guide to Modern Techniques of Plant Analysis; Springer Science & Business Media: Berlin/Heidelberg, Germany, 1998. [Google Scholar]
- Pasdaran, A.; Hamedi, A. Natural products as source of new antimicrobial compounds for skin infections. In The Microbiology of Skin, Soft Tissue, Bone and Joint Infections; Kon, K., Rai, M., Eds.; Elsevier Inc.: Amsterdam, The Netherlands, 2017; pp. 223–253. [Google Scholar]
- Kupina, S.; Fields, C.; Roman, M.C.; Brunelle, S.L. Determination of total phenolic content using the Folin-C assay: Single-laboratory validation, first action 2017.13. J. AOAC Int. 2019, 102, 320–321. [Google Scholar] [CrossRef]
- Pękal, A.; Pyrzynska, K. Evaluation of aluminium complexation reaction for flavonoid content assay. Food Anal. Methods 2014, 7, 1776–1782. [Google Scholar] [CrossRef]
- Faisal, S.; Jan, H.; Shah, S.A.; Shah, S.; Khan, A.; Akbar, M.T.; Syed, S. Green synthesis of zinc oxide (ZnO) nanoparticles using aqueous fruit extracts of Myristica fragrans: Their characterizations and biological and environmental applications. ACS Omega 2021, 6, 9709–9722. [Google Scholar] [CrossRef]
- Syarifah, S.; Imawan, C.; Handayani, W.; Djuhana, D. Biosynthesis of ferric oxide nanoparticles using Pometia pinnata J.R.Frost. & G.Forst. leaves water extract. AIP Conf. Proc. 2018, 2023, 020054. [Google Scholar]
- Jain, N.; Bhargava, A.; Majumdar, S.; Tarafdar, J.C.; Panwar, J. Extracellular biosynthesis and characterization of silver nanoparticles using Aspergillus flavus NJP08: A mechanism perspective. Nanoscale 2011, 3, 635–641. [Google Scholar] [CrossRef] [PubMed]
- Wypij, M.; Jędrzejewski, T.; Trzcińska-Wencel, J.; Ostrowski, M.; Rai, M.; Golińska, P. Green synthesized silver nanoparticles: Antibacterial and anticancer activities, biocompatibility, and analyses of surface-attached proteins. Front. Microbiol. 2021, 12, 632505. [Google Scholar] [CrossRef] [PubMed]
- Devi, L.S.; Joshi, S.R. Antimicrobial and synergistic effects of silver nanoparticles synthesized using soil fungi of high altitudes of eastern Himalaya. Mycobiology 2012, 40, 27–34. [Google Scholar] [CrossRef] [PubMed]
- Sepehri, A.; Golparvar, A.R. The effect of drought stress on water relations, chlorophyll content and leaf area in canola cultivars (Brassica napus L.). Electron. J. Biol. 2011, 7, 49–53. [Google Scholar]
- Lichtenther, H.K. Cholorophylls and carotenoids: Pigments of photosynthetic biomembranes. Met. Enzy. 1987, 148, 350–382. [Google Scholar]
- Bates, L.S.; Waldren, R.P.; Teare, I.D. Rapid determination of free proline for water-stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- El-Argawy, E.; Adss, I.A. Quantitative gene expression of peroxidase, polyphenoloxidase and catalase as molecular markers for resistance against Ralstonia solanacearum. Am. J. Mol. Biol. 2016, 6, 88–100. [Google Scholar] [CrossRef]
- Singleton, V.L.; Rossi, J.A. Colorimetry of total phenolics with phosphomolybdic-phosphotungstic acid reagents. Am. J. Enol. Vitic. 1965, 16, 144–158. [Google Scholar] [CrossRef]
- Association of Official Analytical Chemists. Official Methods of Analysis, 18th ed.; Association of Official Analytical Chemists: Washington, DC, USA, 1995. [Google Scholar]
- Swilam, N.; Nematallah, K.A. Polyphenols profile of pomegranate leaves and their role in green synthesis of silver nanoparticles. Sci. Rep. 2020, 10, 14851. [Google Scholar] [CrossRef] [PubMed]
- Somchaidee, P.; Tedsree, K. Green synthesis of high dispersion and narrow size distribution of zero-valent iron nanoparticles using guava leaf (Psidium guajava L.) extract. Adv. Nat. Sci. Nanosci. Nanotechnol. 2018, 9, 035006. [Google Scholar] [CrossRef]
- Saha, R.; Subramani, K.; Petchi Muthu Raju, S.A.K.; Rangaraj, S.; Venkatachalam, R. Psidium guajava leaf extract-mediated synthesis of ZnO nanoparticles under different processing parameters for hydrophobic and antibacterial finishing over cotton fabrics. Prog. Org. Coat. 2018, 124, 80–91. [Google Scholar] [CrossRef]
- Sornapudi, S.D.; Srivastava, M. Quantitative estimation of phytochemicals in different leaf extracts. Pharma Innov. J. 2022, 11, 125–128. [Google Scholar]
- Jiang, M.; Song, Y.; Kanwar, M.K.; Ahammed, G.J.; Shao, S.; Zhou, J. Phytonanotechnology applications in modern agriculture. J. Nanobiotechnol. 2021, 19, 430. [Google Scholar] [CrossRef]
- El-Serafy, R.S. Silica nanoparticles enhances physio-biochemical characters and postharvest quality of Rosa hybrida L. cut flowers. J. Hortic. Res. 2019, 27, 47–54. [Google Scholar] [CrossRef]
- Mozafari, A.A.; Ghadakchi asl, A.; Ghaderi, N. Grape response to salinity stress and role of iron nanoparticle and potassium silicate to mitigate salt induced damage under in vitro conditions. Physiol. Mol. Biol. Plants 2018, 24, 25–35. [Google Scholar] [CrossRef]
- Merinero, M.; Alcudia, A.; Begines, B.; Martínez, G.; Martín-Valero, M.J.; Pérez-Romero, J.A.; Mateos-Naranjo, E.; Redondo-Gómez, S.; Navarro-Torre, S.; Torres, Y.; et al. Assessing the biofortification of wheat plants by combining a plant growth-promoting rhizobacterium (PGPR) and Polymeric Fe-nanoparticles: Allies or Enemies? Agronomy 2022, 12, 228. [Google Scholar] [CrossRef]
- Bahgat, A.R.; Dahab, A.A.; Elhakem, A.; Gururani, M.A.; El-Serafy, R.S. Integrated action of rhizobacteria with Aloe vera and moringa leaf extracts improves defense mechanisms in Hibiscus sabdariffa L. cultivated in saline Soil. Plants 2023, 12, 3684. [Google Scholar] [CrossRef] [PubMed]
- Pardo, J.M. Biotechnology of water and salinity stress tolerance. Curr. Opin. Biotechnol. 2010, 21, 185–196. [Google Scholar] [CrossRef]
- Munns, R.; Tester, M. Mechanisms of salinity tolerance. Annu. Rev. Plant Biol. 2008, 59, 651–681. [Google Scholar] [CrossRef]
- Almeida, D.M.; Oliveira, M.M.; Saibo, N.J.M. Regulation of Na+ and K+ homeostasis in plants: Towards improved salt stress tolerance in crop plants. Genet. Mol. Biol. 2017, 40, 326–345. [Google Scholar] [CrossRef]
- Roy, S.J.; Negrão, S.; Tester, M. Salt resistant crop plants. Curr. Opin. Biotechnol. 2014, 26, 115–124. [Google Scholar] [CrossRef]
- Krishnamurthy, L.; Serraj, R.; Hash, C.T.; Dakheel, A.J.; Reddy, B.V.S. Screening sorghum genotypes for salinity tolerant biomass production. Euphytica 2007, 156, 15–24. [Google Scholar] [CrossRef]
- Romero-Aranda, R.; Soria, T.; Cuartero, J. Tomato plant-water uptake and plant water relationships under saline growth conditions. Plant Sci. 2001, 160, 265–272. [Google Scholar] [CrossRef]
- AL-Mushhin, A.A. Interactive effect of potassium and spermidine protects growth, photosynthesis and chlorophyll biosynthesis in Vigna angularis from salinity induced damage by up-regulating the tolerance mechanisms. Not. Bot. Horti Agrobot. 2022, 50, 12607. [Google Scholar] [CrossRef]
- Hanafy-Ahmed, A.H.; Khalil, M.K.; Abd-EiRahman, A.M.; Nadia, A.M. Effect of zinc, tryptophan and indole acetic acid on growth, yield and chemical composition of Valencia orange trees. J. Appl. Sci. Res. 2012, 8, 901–914. [Google Scholar]
- Mehrabani, L.V.; Hassanpouraghdam, M.B.; Shamsi-Khotab, T. The effects of common and nano-zinc foliar application on the alleviation of salinity stress in Rosmarinus officinalis L. Acta Sci. Pol.-Hortoru. 2018, 17, 65–73. [Google Scholar] [CrossRef]
- Meng, Y.; Yin, Q.; Yan, Z.; Wang, Y.; Niu, J.; Zhang, J.; Fan, K. Exogenous silicon enhanced salt resistance by maintaining K+/Na+ homeostasis and antioxidant performance in Alfalfa leaves. Front. Plant Sci. 2020, 11, 1183. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, J.; Berni, R.; Hausman, J.-F.; Guerriero, G.A. Review on the beneficial role of silicon against salinity in non-accumulator crops: Tomato as a model. Biomolecules 2020, 10, 1284. [Google Scholar] [CrossRef] [PubMed]
- Alabdallah, N.M.; Alzahrani, H.S. Impact of ZnO nanoparticles on growth of cowpea and okra plants under salt stress conditions. Biosci. Biotech. Res. Asia 2020, 17, 329–340. [Google Scholar] [CrossRef]
- Nasar, J.; Wang, G.-Y.; Ahmad, S.; Muhammad, I.; Zeeshan, M.; Gitari, H.; Adnan, M.; Fahad, S.; Khalid, M.H.B.; Zhou, X.-B.; et al. Nitrogen fertilization coupled with iron foliar application improves the photosynthetic characteristics, photosynthetic nitrogen use efficiency, and the related enzymes of maize crops under different planting patterns. Front. Plant Sci. 2022, 13, 988055. [Google Scholar] [CrossRef]
- El-Serafy, R.S.; Dahab, A.A.; Ghanem, K.Z.; Elhakem, A.; Bahgat, A.R.; Venkatesh, J.; El-Sheshtawy, A.A.; Badawy, A. As a natural antioxidant: Sesbania Grandiflora Leaf extract enhanced growth and yield performance, active ingredients and tolerance of Hibiscus sabdariffa L. under salt-affected soil. J. Soil Sci. Plant Nutr. 2024, 24, 3406–3420. [Google Scholar] [CrossRef]
- Blum, A. Effective use of water (EUW) and not water-use efficiency (WUE) is the target of crop yield improvement under drought stress. Field Crops Res. 2009, 112, 119–123. [Google Scholar] [CrossRef]
- Kumar, D.; Dhankher, O.P.; Tripathi, R.D.; Seth, C.S. Titanium dioxide nanoparticles potentially regulate the mechanism (s) for photosynthetic attributes, genotoxicity, antioxidants defense machinery, and phytochelatins synthesis in relation to hexavalent chromium toxicity in Helianthus annuus L. J. Hazard. Mater. 2023, 454, 131418. [Google Scholar] [CrossRef]
- Vinocur, B.; Altman, A. Recent advances in engineering plant tolerance to abiotic stress: Achievements and limitations. Curr. Opin. Biotech. 2005, 16, 123–132. [Google Scholar] [CrossRef] [PubMed]
- El-Serafy, R.S.; El-Sheshtawy, A.N.A.; Dahab, A.A. Fruit peel soil supplementation induces physiological and biochemical tolerance in Schefflera arboricola L. grown under heat conditions. J. Soil. Sci. Plant Nutr. 2023, 23, 1046–1059. [Google Scholar] [CrossRef]
- Sturikova, H.; Krystofova, O.; Huska, D.; Adam, V. Zinc nanoparticles and plants. J. Hazard. Mater. 2018, 349, 101–110. [Google Scholar] [CrossRef]
- Etesami, H.; Maheshwari, D.K. Use of plant growth promoting rhizobacteria (PGPRs) with multiple plant growth promoting traits in stress agriculture: Action mechanisms and future prospects. Ecotoxicol. Environ. Saf. 2018, 156, 225–246. [Google Scholar] [CrossRef]
- Javad, S.; Maqsood, S.; Shah, A.A.; Singh, A.; Noor Shah, A.; Nawaz, M.; Bashir, M.A.; El Nashar, E.M.; Alghamdi, M.A.; El-kott, A.F.; et al. Iron nanoparticles mitigates cadmium toxicity in Triticum aestivum; modulation of antioxidative defense system and physiochemical characteristics. J. King Saud Univ. Sci. 2023, 35, 102498. [Google Scholar] [CrossRef]
- Konate, A.; He, X.; Zhang, Z.; Ma, Y.; Zhang, P.; Alugongo, G. Magnetic (Fe3O4) nanoparticles reduce heavy metals uptake and mitigate theirtoxicity in wheat seedling. Sustainability 2017, 9, 790. [Google Scholar] [CrossRef]
- Zaheer, I.E.; Ali, S.; Saleem, M.H.; Noor, I.; El-Esawi, M.A.; Hayat, K.; Rizwan, M.; Abbas, Z.; El-Sheikh, M.A.; Alyemeni, M.N. Iron–lysine mediated alleviation of chromium toxicity in spinach (Spinacia oleracea L.) plants in relation to mor-pho-physiological traits and iron uptake when irrigated with tannery wastewater. Sustainability 2020, 12, 6690. [Google Scholar] [CrossRef]
- Weisany, W.; Sohrabi, Y.; Heidari, G.; Siosemardeh, A.; Ghassemi-Golezani, K. Changes in antioxidant enzymes activity and plant performance by salinity stress and zinc application in soybean (Glycine max L.). Plant Omics J. 2012, 5, 60–67. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Soil Property | Value |
|---|---|
| Particle size distribution: | |
| Coarse sand (%) | 6.99 |
| Fine sand (%) | 78.71 |
| Silt (%) | 4.43 |
| Clay (%) | 9.87 |
| Texture class | Loamy sand |
| Field capacity (%) | 13.02 |
| Permanent wilting point (%) | 5.36 |
| Available water (%) | 7.66 |
| Bulk density (Mg m−3) | 1.70 |
| Total porosity (%) | 35.85 |
| pH (1:2.5 soil water suspension) | 8.49 |
| ECe (soil paste extract, dS m−1) | 0.69 |
| CaCO3 content (g kg−1) | 43.37 |
| Available nutrients (mg kg−1) | |
| N | 18.95 |
| P | 4.58 |
| K | 67.33 |
| Fe | 2.60 |
| Zn | 0.62 |
| Mn | 1.18 |
| Cu | 0.47 |
| Treatments | pH | ECW (dS m−1) | Cations (mmolc L−1) | Anions (mmolc L−1) | SAR | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Ca2+ | Mg2+ | Na+ | K+ | CO32− | HCO3− | Cl− | SO42− | ||||
| S1 | 7.12 | 0.75 | 1.50 | 1.74 | 3.84 | 0.42 | 0.00 | 2.09 | 4.95 | 0.46 | 3.02 |
| S2 | 7.44 | 2.00 | 3.45 | 5.02 | 10.98 | 0.55 | 0.00 | 3.63 | 12.94 | 3.43 | 5.34 |
| S3 | 7.51 | 4.00 | 6.86 | 8.49 | 23.55 | 1.10 | 0.00 | 6.63 | 20.88 | 12.49 | 8.50 |
| S4 | 7.55 | 6.00 | 10.26 | 12.44 | 34.75 | 2.55 | 0.00 | 9.47 | 28.98 | 21.55 | 10.31 |
| Active Compounds | Phenols | Saponins | Flavonoids | Coumarins | Carbohydrates | Alkaloids | Terpenes | Tannins |
|---|---|---|---|---|---|---|---|---|
| Inference | + | + | + | + | + | + | + | + |
| Compounds | Concentration |
|---|---|
| Phenols (mg gallic acid equivalent mL−1 of extract) | 15.80 ± 0.48 |
| Flavonoids (mg of quercetin equivalent mL−1 of extract) | 10.30 ± 0.39 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sheta, M.H.; Abd El-Wahed, A.H.M.; Elshaer, M.A.; Bayomy, H.M.; Ozaybi, N.A.; Abd-Elraheem, M.A.M.; El-Sheshtawy, A.-N.A.; El-Serafy, R.S.; Moustafa, M.M.I. Green Synthesis of Zinc and Iron Nanoparticles Using Psidium guajava Leaf Extract Stimulates Cowpea Growth, Yield, and Tolerance to Saline Water Irrigation. Horticulturae 2024, 10, 915. https://doi.org/10.3390/horticulturae10090915
Sheta MH, Abd El-Wahed AHM, Elshaer MA, Bayomy HM, Ozaybi NA, Abd-Elraheem MAM, El-Sheshtawy A-NA, El-Serafy RS, Moustafa MMI. Green Synthesis of Zinc and Iron Nanoparticles Using Psidium guajava Leaf Extract Stimulates Cowpea Growth, Yield, and Tolerance to Saline Water Irrigation. Horticulturae. 2024; 10(9):915. https://doi.org/10.3390/horticulturae10090915
Chicago/Turabian StyleSheta, Mohamed H., Ahmed H. M. Abd El-Wahed, Mohammed A. Elshaer, Hala M. Bayomy, Nawal A. Ozaybi, Mohamed A. M. Abd-Elraheem, Abdel-Nasser A. El-Sheshtawy, Rasha S. El-Serafy, and Mahmoud M. I. Moustafa. 2024. "Green Synthesis of Zinc and Iron Nanoparticles Using Psidium guajava Leaf Extract Stimulates Cowpea Growth, Yield, and Tolerance to Saline Water Irrigation" Horticulturae 10, no. 9: 915. https://doi.org/10.3390/horticulturae10090915
APA StyleSheta, M. H., Abd El-Wahed, A. H. M., Elshaer, M. A., Bayomy, H. M., Ozaybi, N. A., Abd-Elraheem, M. A. M., El-Sheshtawy, A.-N. A., El-Serafy, R. S., & Moustafa, M. M. I. (2024). Green Synthesis of Zinc and Iron Nanoparticles Using Psidium guajava Leaf Extract Stimulates Cowpea Growth, Yield, and Tolerance to Saline Water Irrigation. Horticulturae, 10(9), 915. https://doi.org/10.3390/horticulturae10090915