


Effect of Microbial Reinforcement on Polyphenols in the Acetic Acid Fermentation of Shanxi-Aged Vinegar
 , ,
, ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Microculture
2.1.1. Strains
2.1.2. Media
2.1.3. Strain Culture
2.2. Sample Collection
2.3. Analysis of PPs Contents and Composition during AAF
2.4. Macrotranscriptome Sequencing of Microbial Communities during AAF
2.4.1. Extraction of RNA
2.4.2. Library Quality Checks and Sorting
2.5. Effects of Microorganisms on PPs
2.6. Determination of Physical and Chemical Indices during AAF
2.7. Statistical Analysis
3. Results
3.1. Correlation Analysis between PPs and Microorganisms during AAF of SAV
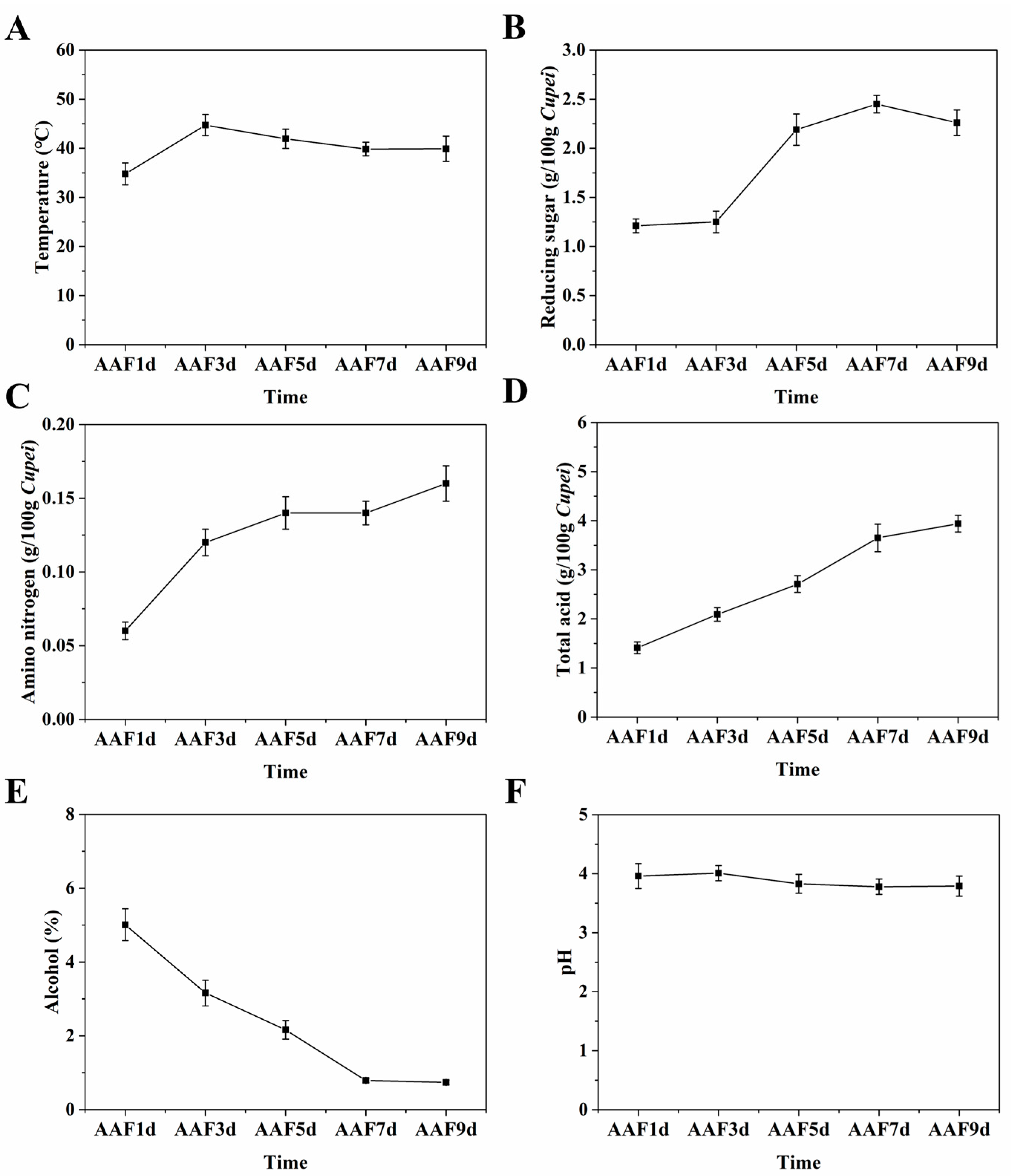
3.1.1. Change in the PPs Contents during AAF
3.1.2. Changes in the Physical and Chemical Indices during AAF
3.1.3. Succession Law of the Microbial Community during AAF
3.1.4. Correlation Analysis between PPs and Microorganisms during AAF
3.2. Single Factor Verification of Microorganisms Affecting PPs Contents
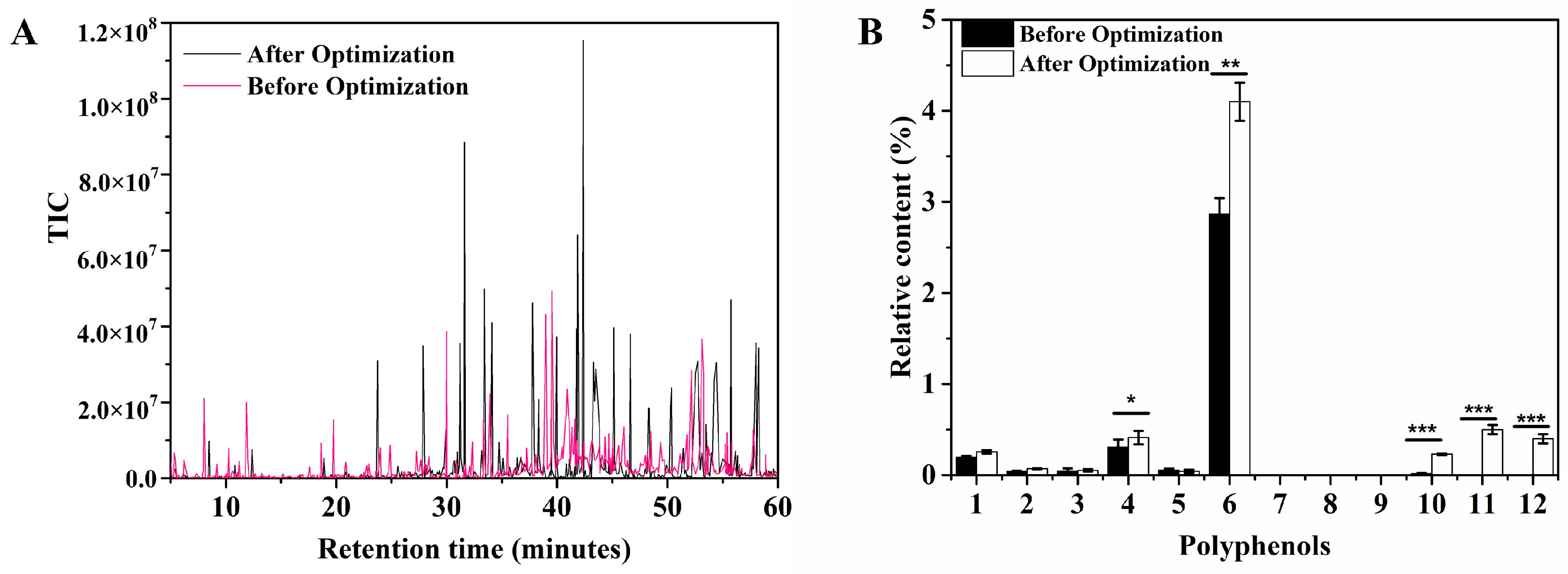
3.3. In Situ Regulation of PPs during AAF
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nie, Z.; Zheng, Y.; Xie, S.; Zhang, X.; Song, J.; Xia, M.; Wang, M. Unraveling the correlation between microbiota succession and metabolite changes in traditional Shanxi aged vinegar. Sci. Rep. 2017, 7, 9240. [Google Scholar] [CrossRef] [PubMed]
- Xie, X.; Zheng, Y.; Liu, X.; Cheng, C.; Zhang, X.; Xia, T.; Yu, S.; Wang, M. Antioxidant Activity of Chinese Shanxi Aged Vinegar and Its Correlation with Polyphenols and Flavonoids During the Brewing Process. J. Food Sci. 2017, 82, 2479–2486. [Google Scholar] [CrossRef] [PubMed]
- Del Rio, D.; Rodriguez-Mateos, A.; Spencer, J.P.E.; Tognolini, M.; Borges, G.; Crozier, A. Dietary (Poly)phenolics in Human Health: Structures, Bioavailability, and Evidence of Protective Effects Against Chronic Diseases. Antioxid. Redox Signal. 2013, 18, 1818–1892. [Google Scholar] [CrossRef] [PubMed]
- Zhu, M.J. Dietary polyphenols, gut microbiota, and intestinal epithelial health. In Nutritional and Therapeutic Interventions for Diabetes and Metabolic Syndrome; Academic Press: Cambridge, MA, USA, 2018; pp. 295–314. [Google Scholar]
- Fan, J.; Zhang, Y.; Chang, X.; Zhang, B.; Jiang, D.; Saito, M.; Li, Z. Antithrombotic and fibrinolytic activities of methanolic extract of aged sorghum vinegar. J. Agric. Food Chem. 2009, 57, 8683–8687. [Google Scholar] [CrossRef] [PubMed]
- Gu, X.; Zhao, H.-L.; Sui, Y.; Guan, J.; Chan, J.C.N.; Tong, P.C.Y. White rice vinegar improves pancreatic beta-cell function and fatty liver in streptozotocin-induced diabetic rats. Acta Diabetol. 2010, 49, 185–191. [Google Scholar] [CrossRef] [PubMed]
- Kahraman, N.K. The effect of vinegar on postprandial glycemia: Does the amount matter? Acta Endocrinol. 2011, 7, 577–584. [Google Scholar] [CrossRef]
- Kondo, S.; Tayama, K.; Tsukamoto, Y.; Ikeda, K.; Yamori, Y. Antihypertensive effects of acetic acid and vinegar on spontaneously hypertensive rats. Biosci. Biotechnol. Biochem. 2001, 65, 2690–2694. [Google Scholar] [CrossRef]
- Sugiyama, A.; Saitoh, M.; Takahara, A.; Satoh, Y.; Hashimoto, K. Acute cardiovascular effects of a new beverage made of wine vinegar and grape juice, assessed using an in vivo rat. Nutr. Res. 2003, 23, 1291–1296. [Google Scholar] [CrossRef]
- Tsao, R. Chemistry and biochemistry of dietary polyphenols. Nutrients 2010, 2, 1231–1246. [Google Scholar] [CrossRef]
- Pastor-Villaescusa, B.; Sanchez Rodriguez, E.; Rangel-Huerta, O.D. Polyphenols in obesity and metabolic syndrome. In Obesity; Academic Press: Cambridge, MA, USA, 2018; pp. 213–239. [Google Scholar] [CrossRef]
- Rosazza, J.P.N.; Huang, Z.; Dostal, L.; Volm, T.; Rousseau, B. Review: Biocatalytic transformations of ferulic acid: An abundant aromatic natural product. J. Ind. Microbiol. Biotechnol. 1995, 15, 457–471. [Google Scholar] [CrossRef]
- Hansen, E.H.; Møller, B.L.; Kock, G.R.; Bünner, C.M.; Kristensen, C.; Jensen, O.R.; Okkels, F.T.; Olsen, C.E.; Motawia, M.S.; Hansen, J. De novo biosynthesis of vanillin in fission yeast (Schizosaccharomyces pombe) and baker’s yeast (Saccharomyces cerevisiae). Appl. Environ. Microbiol. 2009, 75, 2765–2774. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.J.; Ma, Y.K.; Zhang, F.F.; Chen, F.S. Biodiversity of yeasts, lactic acid bacteria and acetic acid bacteria in the fermentation of “Shanxi aged vinegar”, a traditional Chinese vinegar. Food Microbiol. 2012, 30, 289–297. [Google Scholar] [CrossRef]
- Wang, Z.-M.; Lu, Z.-M.; Shi, J.-S.; Xu, Z.-H. Exploring flavour-producing core microbiota in multispecies solid-state fermentation of traditional Chinese vinegar. Sci. Rep. 2016, 6, 26818. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.-M.; Lu, Z.-M.; Yu, Y.-J.; Li, G.-Q.; Shi, J.-S.; Xu, Z.-H. Batch-to-batch uniformity of bacterial community succession and flavour formation in the fermentation of Zhenjiang aromatic vinegar. Food Microbiol. 2015, 50, 64–69. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Wang, X.; Zhang, C.; Zhou, X.; Xu, X.; Han, L.; Lv, X.; Liu, Y.; Liu, S.; Li, J.; et al. De novo biosynthesis of rubusoside and rebaudiosides in engineered yeasts. Nat. Commun. 2022, 13, 3040. [Google Scholar] [CrossRef] [PubMed]
- Zhou, R.; Bhuiya, M.W.; Cai, X.; Yu, X.; Eilerman, R.G. Methods of Making Vanillin via Microbial Fermentation Utilizing Ferulic Acid Provided by a Modified Caffeic Acid 3-o-Methyltransferase. Francia. Patent WO 106189, 3 July 2014. [Google Scholar]
- Sankuan, X.; Cuimei, Z.; Bingqian, F.; Yu, Z.; Menglei, X.; Linna, T.; Jia, S.; Xinyi, Z.; Min, W. Metabolic network of ammonium in cereal vinegar solid-state fermentation and its response to acid stress. Food Microbiol. 2020, 95, 103684. [Google Scholar] [CrossRef]
- Du, P.; Zhou, J.; Zhang, L.; Zhang, J.; Li, N.; Zhao, C.; Tu, L.; Zheng, Y.; Xia, T.; Luo, J.; et al. GC× GC-MS analysis and hypolipidemic effects of polyphenol extracts from Shanxi-aged vinegar in rats under a high-fat diet. Food Funct. 2020, 11, 7468–7480. [Google Scholar] [CrossRef]
- Martin, M. Cutadapt removes adapter sequences from high-throughput sequencing reads. EMBnet. J. 2011, 17, 10–12. [Google Scholar] [CrossRef]
- Kopylova, E.; Noé, L.; Touzet, H. SortMeRNA: Fast and accurate filtering of ribosomal RNAs in metatranscriptomic data. Bioinformatics 2012, 28, 3211–3217. [Google Scholar] [CrossRef]
- Robert, S.; Yan, L.; Robert, E. Identification and removal of ribosomal RNA sequences from metatranscriptomes. Bioinformatics 2012, 28, 433–435. [Google Scholar] [CrossRef]
- Grabherr, M.G.; Haas, B.J.; Yassour, M.; Levin, J.Z.; Thompson, D.A.; Amit, I.; Adiconis, X.; Fan, L.; Raychowdhury, R.; Zeng, Q.D.; et al. Full-length transcriptome assembly from RNA-Seq data without a reference genome. Nat. Biotechnol. 2011, 29, 644–652. [Google Scholar] [CrossRef] [PubMed]
- Nie, Z.; Zheng, Y.; Wang, M.; Han, Y.; Wang, Y.; Luo, J.; Niu, D. Exploring microbial succession and diversity during solid-state fermentation of Tianjin duliu mature vinegar. Bioresour. Technol. 2013, 148, 325–333. [Google Scholar] [CrossRef] [PubMed]
- Song, J.; Jia, Y.-X.; Su, Y.; Zhang, X.-Y.; Tu, L.-N.; Nie, Z.-Q.; Zheng, Y.; Wang, M. Initial Analysis on the Characteristics and Synthesis of Exopolysaccharides from Sclerotium rolfsii with Different Sugars as Carbon Sources. Polymers 2020, 12, 348. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Li, S.; Cheng, L.; Luo, L. Analyzing the relation between the microbial diversity of DaQu and the turbidity spoilage of traditional Chinese vinegar. Appl. Microbiol. Biotechnol. 2014, 98, 6073–6084. [Google Scholar] [CrossRef] [PubMed]
- Xu, W.; Huang, Z.; Zhang, X.; Li, Q.; Lu, Z.; Shi, J.; Xu, Z.; Ma, Y. Monitoring the microbial community during solid-state acetic acid fermentation of Zhenjiang aromatic vinegar. Food Microbiol. 2011, 28, 1175–1181. [Google Scholar] [CrossRef] [PubMed]
- Priefert, H.; Rabenhorst, J.; Steinbüchel, A. Biotechnological production of vanillin. Appl. Microbiol. Biotechnol. 2001, 56, 296–314. [Google Scholar] [CrossRef] [PubMed]
- Fadda, S.; Oliver, G.; Vignolo, G. Protein Degradation by Lactobacillus plantarum and Lactobacillus casei in a Sausage Model System. J. Food Sci. 2002, 67, 1179–1183. [Google Scholar] [CrossRef]
- Pfeiler, E.A.; Klaenhammer, T.R. The genomics of lactic acid bacteria. Trends Microbiol. 2007, 15, 546–553. [Google Scholar] [CrossRef]
- Rao, B.V.; Ramanjaneyulu, K.; Rambabu, T.; Devi, C.H.B.T.S. Synthesis and antioxidant activity of galloyl tyrosine derivatives from young leaves of Inga laurina. Int. J. Pharma Bio Sci. 2011, 2, 39–44. [Google Scholar]
- Yan, Y.; Kohli, A.; Koffas, M.A.G. Biosynthesis of natural flavanones in Saccharomyces cerevisiae. Appl. Environ. Microbiol. 2005, 71, 5610–5613. [Google Scholar] [CrossRef]
- Li, K.; Frost, J.W. Synthesis of Vanillin from Glucose. J. Am. Chem. Soc. 1998, 120, 10545–10546. [Google Scholar] [CrossRef]
- Nethaji, J.G.; Birger, L.M. Vanillin–Bioconversion and Bioengineering of the Most Popular Plant Flavor and Its De Novo Biosynthesis in the Vanilla Orchid. Mol. Plant 2015, 8, 40–57. [Google Scholar] [CrossRef]
- Lu, Z.-M.; Xu, W.; Yu, N.-H.; Zhou, T.; Li, G.-Q.; Shi, J.-S.; Xu, Z.-H. Recovery of aroma substances from Zhenjiang aromatic vinegar by supercritical fluid extraction. Int. J. Food Sci. Technol. 2011, 46, 1508–1514. [Google Scholar] [CrossRef]
- Chen, J.-C.; Chen, Q.-H.; Guo, Q.; Ruan, S.; Ruan, H.; He, G.-Q.; Gu, Q. Simultaneous determination of acetoin and tetramethylpyrazine in traditional vinegars by HPLC method. Food Chem. 2010, 122, 1247–1252. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound Identification | CAS No. | Relative Content (%) | ||||
|---|---|---|---|---|---|---|
| AAF 1d | AAF 3d | AAF 5d | AAF 7d | AAF 9d | ||
| p-Coumaric acid | 501-98-4 | 0.102 ± 0.068 ab | 0.053 ± 0.037 a | 0.07 ± 0.06 ab | 0.127 ± 0.053 ab | 0.193 ± 0.017 b |
| 4-Hydroxy-3-methoxyphenylethylene Glycol | 534-82-7 | 0.156 ± 0.156 a | 0.074 ± 0.106 a | 0.013 ± 0.008 a | 0.067 ± 0.153 a | 0.0425 ± 0.047 a |
| Vanillic acid | 121-34-6 | 0.127 ± 0.213 a | 0.013 ± 0.017 a | 0.05 ± 0.02 a | 0.037 ± 0.017 a | 0.047 ± 0.028 a |
| Ferulic Acid | 1135-24-6 | 0.157 ± 0.077 a | 0.077 ± 0.073 a | 0.3 ± 0.08 a | 0.263 ± 0.147 a | 0.307 ± 0.083 a |
| L-Epicatechin | 490-46-0 | 0.301 ± 0.419 a | 0 | 0.075 ± 0.015 a | 0.11 ± 0.05 a | 0.053 ± 0.017 a |
| Gallic acid | 149-91-7 | 0 | 1.727 ± 0.3347 a | 2.59 ± 0.69 a | 2.383 ± 0.483 a | 2.865 ± 2.775 a |
| 3-(3-Hydroxy-4-methoxyphenyl)propanoic acid | 1135-15-5 | 0.163 ± 0.137 a | 0.053 ± 0.037 a | 0 | 0.107 ± 0.067 a | 0 |
| Caffeic acid | 331-39-5 | 0.063 ± 0.097 a | 0 | 0.05 ± 0.05 a | 0 | 0 |
| 2-Propenoic acid, 3-(3-hydroxyphenyl)-, ethyl ester, (2E)- | 96251-92-2 | 0 | 0.018 ± 0.022 a | 0 | 0.14 ± 0.11 b | 0 |
| Vanillin | 121-33-5 | 0 | 0 | 0 | 0.007 ± 0.007 a | 0.0175 ± 0.017 a |
| Compound Identification | CAS No. | Relative Content (%) | ||
|---|---|---|---|---|
| Before Optimization | After Optimization | |||
| 1 | p-Coumaric acid | 501-98-4 | 0.193 ± 0.017 | 0.256 ± 0.019 |
| 2 | 4-Hydroxy-3-methoxyphenylethylene Glycol | 534-82-7 | 0.0425 ± 0.0047 | 0.07 ± 0.005 |
| 3 | Vanillic acid | 121-34-6 | 0.047 ± 0.028 | 0.052 ± 0.012 |
| 4 | Ferulic Acid | 1135-24-6 | 0.307 ± 0.083 | 0.412 ± 0.075 * |
| 5 | L-Epicatechin | 490-46-0 | 0.053 ± 0.017 | 0.043 ± 0.016 |
| 6 | Gallic acid | 149-91-7 | 2.865 ± 0.175 | 4.1 ± 0.21 ** |
| 7 | 3-(3-Hydroxy-4-methoxyphenyl)propanoic acid | 1135-15-5 | 0 | 0 |
| 8 | Caffeic acid | 331-39-5 | 0 | 0 |
| 9 | 2-Propenoic acid, 3-(3-hydroxyphenyl)-, ethyl ester, (2E)- | 96251-92-2 | 0 | 0 |
| 10 | Vanillin | 121-33-5 | 0.0175 ± 0.007 | 0.23 ± 0.009 *** |
| 11 | Isoferulic acid | 1135-16-6 | 0 | 0.5 ± 0.05 *** |
| 12 | Salicylic acid | 69-72-7 | 0 | 0.4 ± 0.05 *** |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Du, P.; Li, Y.; Zhen, C.; Song, J.; Hou, J.; Gou, J.; Li, X.; Xie, S.; Zhou, J.; Yan, Y.; et al. Effect of Microbial Reinforcement on Polyphenols in the Acetic Acid Fermentation of Shanxi-Aged Vinegar. Fermentation 2023, 9, 756. https://doi.org/10.3390/fermentation9080756
Du P, Li Y, Zhen C, Song J, Hou J, Gou J, Li X, Xie S, Zhou J, Yan Y, et al. Effect of Microbial Reinforcement on Polyphenols in the Acetic Acid Fermentation of Shanxi-Aged Vinegar. Fermentation. 2023; 9(8):756. https://doi.org/10.3390/fermentation9080756
Chicago/Turabian StyleDu, Peng, Yingqi Li, Chenrui Zhen, Jia Song, Jiayi Hou, Jia Gou, Xinyue Li, Sankuan Xie, Jingli Zhou, Yufeng Yan, and et al. 2023. "Effect of Microbial Reinforcement on Polyphenols in the Acetic Acid Fermentation of Shanxi-Aged Vinegar" Fermentation 9, no. 8: 756. https://doi.org/10.3390/fermentation9080756
APA StyleDu, P., Li, Y., Zhen, C., Song, J., Hou, J., Gou, J., Li, X., Xie, S., Zhou, J., Yan, Y., Zheng, Y., & Wang, M. (2023). Effect of Microbial Reinforcement on Polyphenols in the Acetic Acid Fermentation of Shanxi-Aged Vinegar. Fermentation, 9(8), 756. https://doi.org/10.3390/fermentation9080756