
Fungal Bioprocessing to Improve Quality of Pennycress Meal as a Potential Feeding Ingredient for Monogastric Animals

Abstract
1. Introduction
2. Materials and Methods
2.1. Fungal Strains Preparation
2.2. Solid-State Fermentation
2.3. Analytical Methods
2.4. Statistical Analysis
3. Results and Discussion
3.1. PM as Monogastric Animal Feed Ingredient and Its Limitations
3.2. Change in Structural Carbohydrates in PM by Fermentation
3.3. Change in Protein and Amino Acids Profile in PM by Fermentation
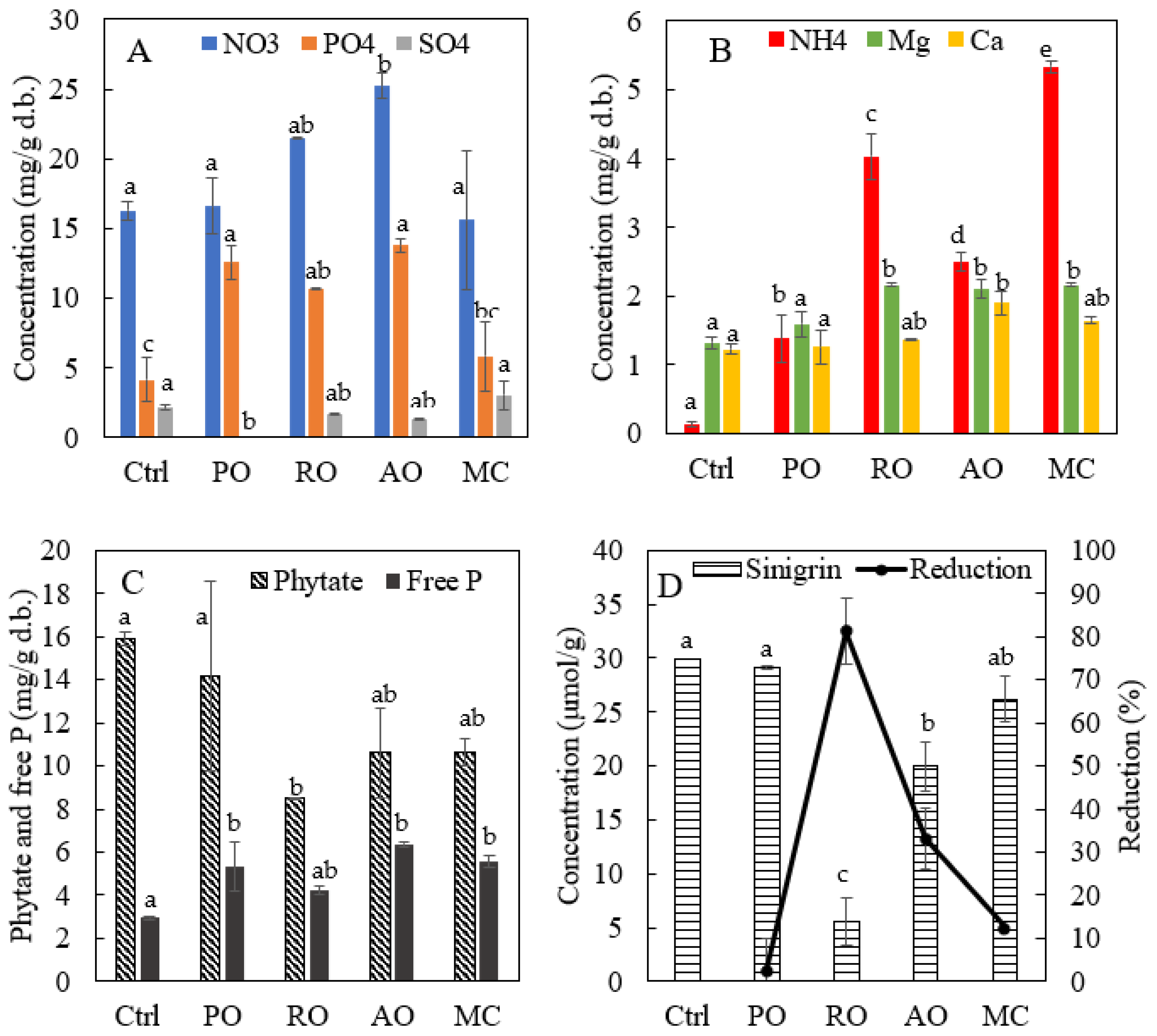
3.4. Change in Minerals, Phytate and Sinigrin in PM by Fermentation
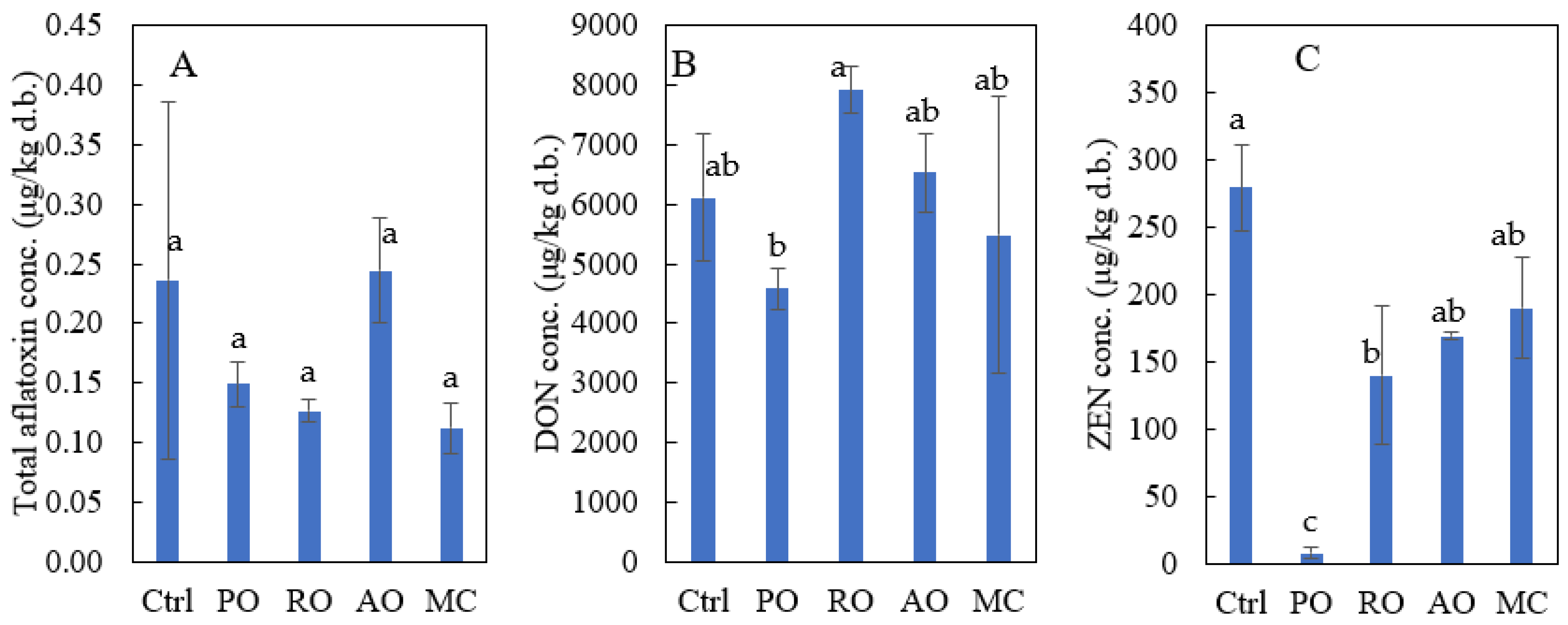
3.5. Change in Mycotoxins in PM by Fermentation
3.6. In Vitro Digestibility of Dry Matter and Amino Acids
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Dou, Z.; Toth, J.D.; Westendorf, M.L. Food waste for livestock feeding: Feasibility, safety, and sustainability implications. Glob. Food Sec. 2018, 17, 154–161. [Google Scholar] [CrossRef]
- Marks, M.D.; Chopra, R.; Sedbrook, J.C. Technologies enabling rapid crop improvements for sustainable agriculture: Example pennycress (Thlaspi arvense L.). Emerg. Top. Life Sci. 2021, 5, 325–335. [Google Scholar] [CrossRef]
- Marks, M.D.; Sedbrook, J.C.; Wyse, D.L.; Kevin, D. Plants Having Increased Oil Quality. U.S. Patent US11396657B2, 26 July 2022. [Google Scholar]
- Moser, B.R. Biodiesel from alternative oilseed feedstocks: Camelina and field pennycress. Biofuels 2012, 3, 193–209. [Google Scholar] [CrossRef]
- Mousavi-Avval, S.H.; Shah, A. Techno-economic analysis of pennycress production, harvest and post-harvest logistics for renewable jet fuel. Renew. Sust. Energy Rev. 2020, 123, 109764. [Google Scholar] [CrossRef]
- Sedbrook, J.C.; Phippen, W.B.; Marks, M.D. New approaches to facilitate rapid domestication of a wild plant to an oilseed crop: Example pennycress (Thlaspi arvense L.). Plant Sci. 2014, 227, 122–132. [Google Scholar] [CrossRef]
- Alhotan, R.A.; Wang, R.L.; Holser, R.A.; Pesti, G.M. Nutritive value and the maximum inclusion level of pennycress meal for broiler chickens. Poult. Sci. 2017, 96, 2281–2293. [Google Scholar] [CrossRef] [PubMed]
- Barnharst, T.; Sun, X.; Rajendran, A.; Urriola, P.; Shurson, G.; Hu, B. Enhanced protein and amino acids of corn–ethanol co-product by Mucor indicus and Rhizopus oryzae. Bioprocess Biosyst. Eng. 2021, 44, 1989–2000. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Tiffany, D.G.; Urriola, P.E.; Shurson, G.G.; Hu, B. Nutrition upgrading of corn-ethanol co-product by fungal fermentation: Amino acids enrichment and anti-nutritional factors degradation. Food Bioprod. Proc. 2021, 130, 1–13. [Google Scholar] [CrossRef]
- Sun, X.; Devi, D.; Urriola, P.; Tiffany, D.; Jang, J.-C.; Shurson, G.; Hu, B. Feeding value improvement of corn-ethanol co-product and soybean hull by fungal fermentation: Fiber degradation and digestibility improvement. Food Bioprod. Proc. 2021, 130, 143–153. [Google Scholar] [CrossRef]
- Heidari, F.; Øverland, M.; Hansen, J.Ø.; Mydland, L.T.; Urriola, P.E.; Chen, C.; Shurson, G.C.; Hu, B. Solid-state fermentation of Pleurotus ostreatus to improve the nutritional profile of mechanically-fractionated canola meal. Biochem. Eng. J. 2022, 187, 108591. [Google Scholar] [CrossRef]
- Ji, C.; Fan, Y.; Zhao, L. Review on biological degradation of mycotoxins. Anim. Nutr. 2016, 2, 127–133. [Google Scholar] [CrossRef]
- Pandey, A. Recent process developments in solid-state fermentation. Process Biochem. 1992, 27, 109–117. [Google Scholar] [CrossRef]
- Mukherjee, R.; Chakraborty, R.; Dutta, A. Role of fermentation in improving nutritional quality of soybean meal—A Review. Asian-Australas. J. Anim. Sci. 2016, 29, 1523–1529. [Google Scholar] [CrossRef] [PubMed]
- Feng, J.; Liu, X.; Xu, Z.R.; Lu, Y.P.; Liu, Y.Y. The effect of Aspergillus oryzae fermented soybean meal on growth performance, digestibility of dietary components and activities of intestinal enzymes in weaned piglets. Anim. Feed Sci. Technol. 2007, 134, 295–303. [Google Scholar] [CrossRef]
- Aljuobori, A.; Abdullah, N.; Zulkifli, I.; Soleimani, A.; Liang, J.; Oskoueian, E. Lactobacillus salivarius fermentation reduced glucosinolate and fibre in canola meal. J. Food Res. 2014, 3, 95. [Google Scholar]
- Croat, J.R.; Berhow, M.; Karki, B.; Muthukumarappan, K.; Gibbons, W.R. Conversion of canola meal into a high-protein feed additive via solid-state fungal incubation process. J. Am. Oil Chem. Soc. 2016, 93, 499–507. [Google Scholar] [CrossRef]
- Alhomodi, A.F.; Zavadil, A.; Berhow, M.; Gibbons, W.R.; Karki, B. Application of cocultures of fungal mycelium during solid-state fermentation of canola meal for potential feed application. J. Am. Oil Chem. Soc. 2021, 98, 509–517. [Google Scholar] [CrossRef]
- Sluiter, A.; Hames, B.; Ruiz, R.; Scarlata, C.; Sluiter, J.; Templeton, D.; Crocker, D. Determination of structural carbohydrates and lignin in biomass. Lab. Anal. Proceed. 2008, 1617, 1–16. [Google Scholar]
- AOAC. AOAC Official Method 994.12: Amino Acids in Feed; AOAC International: Rockville, MD, USA, 2005. [Google Scholar]
- Henderson, J.W.; Brooks, A. Improved Amino Acid Methods Using Agilent ZORBAX Eclipse Plus C18 Columns for a Variety of Agilent LC Instrumentation and Separation Goals; Agilent Technologies: Santa Clara, CA, USA, 2010. [Google Scholar]
- Megazyme. Phytic Acid (Phytate)/Total Phosphorous Assay Procedure; Megazyme: Wicklow, Ireland, 2019. [Google Scholar]
- Jang, J.-C.; Zeng, Z.; Shurson, G.C.; Urriola, P.E. Effects of gas production recording system and pig fecal inoculum volume on kinetics and variation of in vitro fermentation using corn distiller’s dried grains with solubles and soybean hulls. Animals 2019, 9, 773. [Google Scholar] [CrossRef]
- Alhomodi, A.F.; Kasiga, T.; Berhow, M.; Brown, M.L.; Gibbons, W.R.; Karki, B. Combined effect of mild pretreatment and fungal fermentation on nutritional characteristics of canola meal and nutrient digestibility of processed canola meal in rainbow trout. Food Bioprod. Proc. 2022, 133, 57–66. [Google Scholar] [CrossRef]
- Mejicanos, G.; Sanjayan, N.; Kim, I.H.; Nyachoti, C.M. Recent advances in canola meal utilization in swine nutrition. J. Anim. Sci. Technol. 2016, 58, 7. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Chen, Y.; Luo, L.; Heidari, F.; Tiffany, D.G.; Urriola, P.E.; Shurson, G.G.; Hu, B. Feeding value improvement by co-fermentation of corn-ethanol co-product and agro-industrial residues with Rhizopus oryzae. Process Biochem. 2021, 111, 144–153. [Google Scholar] [CrossRef]
- Humer, E.; Schwarz, C.; Schedle, K. Phytate in pig and poultry nutrition. J. Anim. Physiol. Anim. Nutr. 2015, 99, 605–625. [Google Scholar] [CrossRef]
- Almeida, F.N.; Stein, H.H. Effects of graded levels of microbial phytase on the standardized total tract digestibility of phosphorus in corn and corn coproducts fed to pigs. J. Anim. Sci. 2012, 90, 1262–1269. [Google Scholar] [CrossRef] [PubMed]
- Valmaseda, M.; Martínez, M.J.; Martínez, A.T. Kinetics of wheat straw solid-state fermentation with Trametes versicolor and Pleurotus ostreatus—Lignin and polysaccharide alteration and production of related enzymatic activities. Appl. Microbiol. Biotechnol. 1991, 35, 817–823. [Google Scholar] [CrossRef]
- Rodrigues Reis, C.E.; Bento, H.B.S.; Carvalho, A.K.F.; Rajendran, A.; Hu, B.; De Castro, H.F. Critical applications of Mucor circinelloides within a biorefinery context. Critic. Rev. Biotechnol. 2019, 39, 555–570. [Google Scholar] [CrossRef]
- Jha, R.; Fouhse, J.M.; Tiwari, U.P.; Li, L.; Willing, B.P. Dietary fiber and intestinal health of monogastric animals. Front. Vet. Sci. 2019, 6, 48. [Google Scholar] [CrossRef]
- Zhao, G.; Ding, L.-L.; Pan, Z.-H.; Kong, D.-H.; Hadiatullah, H.; Fan, Z.-C. Proteinase and glycoside hydrolase production is enhanced in solid-state fermentation by manipulating the carbon and nitrogen fluxes in Aspergillus oryzae. Food Chem. 2019, 271, 606–613. [Google Scholar] [CrossRef] [PubMed]
- Blanco, A.; Blanco, G. Chapter 16—Amino Acid Metabolism. In Medical Biochemistry; Blanco, A., Blanco, G., Eds.; Academic Press: Cambridge, MA, USA, 2017; pp. 367–399. [Google Scholar]
- Al-Hussainy, K.S.J.; Al-Fadhly, N.K.Z. Comparison between protein and amino acids of mushroom Agarieus bispours with some kinds of meat and meat’s products. IOP Conf. Ser. Earth Environ. Sci. 2019, 388, 012059. [Google Scholar] [CrossRef]
- Rani, R.; Ghosh, S. Production of phytase under solid-state fermentation using Rhizopus oryzae: Novel strain improvement approach and studies on purification and characterization. Bioresour. Technol. 2011, 102, 10641–10649. [Google Scholar] [CrossRef]
- Hassan, A.A. Evaluation of some Basidiomycetes fruit bodies and cultivation conditions of most efficient fungus, Pleurotus ostreatus for phytase production Abdullah A. Hassan & Hawazin A. Al-Jobory. Tikrit J. Agric. Sci. 2016, 16, 18–29. [Google Scholar]
- Sapna; Singh, B. Phytase production by Aspergillus oryzae in solid-state fermentation and its applicability in dephytinization of wheat bran. Appl. Biochem. Biotechnol. 2014, 173, 1885–1895. [Google Scholar] [CrossRef] [PubMed]
- Venkataraman, S.; Vaidyanathan, V.K. Dephytinization of wheat and rice bran by cross-linked enzyme aggregates of Mucor indicus phytase: A viable prospect for food and feed industries. J. Sci. Food Agric. 2023, 103, 1935–1945. [Google Scholar] [CrossRef]
- Ulmasov, T.; Sedbrook, J.C.; Marks, M.D.; McGinn, M.G.; Chopra, R.; Jarvis, B.A. Low Glucosinolate Pennycress Meal and Methods of Making. U.S. Patent US10988772B2, 27 June 2023. [Google Scholar]
- Ulmasov, T.; Hartnell, G.; Sedbrook, J.C.; Marks, M.D.; Chopra, R.; Esfahanian, M. Low Fiber Pennycress Meal, Seeds, and Methods of Making. U.S. Patent US10709151B2, 16 May 2023. [Google Scholar]
- Smits, J.P.; Knol, W.; Bol, J. Glucosinolate degradation by Aspergillus clavatus and Fusarium oxysporum in liquid and solid-state fermentation. Appl. Microbiol. Biotechnol. 1993, 38, 696–701. [Google Scholar] [CrossRef]
- Cheng, D.L.; Hashimoto, K.; Uda, Y. In vitro digestion of sinigrin and glucotropaeolin by single strains of Bifidobacterium and identification of the digestive products. Food Chem. Toxicol. 2004, 42, 351–357. [Google Scholar] [CrossRef]
- Andini, S.; Dekker, P.; Gruppen, H.; Araya-Cloutier, C.; Vincken, J.-P. Modulation of glucosinolate composition in Brassicaceae seeds by germination and fungal elicitation. J. Agric. Food Chem. 2019, 67, 12770–12779. [Google Scholar] [CrossRef]
- Zain, M.E. Impact of mycotoxins on humans and animals. J. Saudi Chem. Soc. 2011, 15, 129–144. [Google Scholar] [CrossRef]
- Gruber-Dorninger, C.; Jenkins, T.; Schatzmayr, G. Global mycotoxin occurrence in feed: A ten-year survey. Toxins 2019, 11, 375. [Google Scholar] [CrossRef]
- FDA. FDA Mycotoxin Regulatory Guidance; National Grain and Feed Association: Arlington, VA, USA, 2011.
- Sun, X.; Urriola, P.E.; Shurson, G.; Tiffany, D.; Hu, B. Enhancing feeding value of corn distiller’s grains with solubles via fungal co-cultured solid-state fermentation for monogastric animal nutrition. Anim. Feed Sci. Technol. 2023, 303, 115673. [Google Scholar] [CrossRef]
- Chutmanop, J.; Chuichulcherm, S.; Chisti, Y.; Srinophakun, P. Protease production by Aspergillus oryzae in solid-state fermentation using agroindustrial substrates. J. Chem. Technol. Biotechnol. 2008, 83, 1012–1018. [Google Scholar] [CrossRef]
- Tripathi, J.P.; Yadav, J.S. Optimisation of solid substrate fermentation of wheat straw into animal feed by Pleurotus ostreatus: A pilot effort. Anim. Feed Sci. Technol. 1992, 37, 59–72. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Parameter a | PM (This Study) |
|---|---|
| TS, % | 82.5 |
| MC, % | 17.5 |
| CP, % DM basis | 31.1 |
| Total SC, % DM basis | 15.1 |
| Total AA, % DM basis | 22.7 |
| Phytate, % DM basis | 1.6 |
| Glucosinolates, μmol/g, DM basis | 30 (sinigrin) |
| Ctrl | PO | RO | AO | MC | |
|---|---|---|---|---|---|
| Solid recovery, % | 100.00 | 98.56 | 91.24 | 92.71 | 92.82 |
| mg/g DM basis | |||||
| Cellulose | 82.70 ± 0.01 a | 111.40 ± 0.50 cd | 101.99 ± 0.24 bc | 120.99 ± 0.22 d | 92.94 ± 0.04 ab |
| Hemicellulose | 68.00 ± 0.37 ab | 41.71 ± 0.14 c | 49.71 ± 0.22 c | 54.11 ± 0.03 bc | 73.37 ± 0.71 a |
| Glucose | 7.15 ± 0.49 ab | 3.95 ± 0.49 c | 4.10 ± 0.28 bc | 8.15 ± 1.48 a | 6.75 ± 0.64 abc |
| Xylose | 0.95 ± 0.64 a | 0.75 ± 1.06 a | 0.00 ± 0.00 a | 0.50 ± 0.71 a | 1.00 ± 0.57 a |
| Galactose | 2.45 ± 0.92 a | 0.00 ± 0.00 a | 0.85 ± 0.21 a | 0.70 ± 0.99 a | 1.05 ± 0.35 a |
| Mannose | 0.55 ± 0.78 b | 1.05 ± 0.21 ab | 0.55 ± 0.07 b | 1.05 ± 0.07 ab | 2.15 ± 0.07 a |
| Ctrl | PO | RO | AO | MC | |
|---|---|---|---|---|---|
| EAA | mg/g d.b. | ||||
| Arginine | 19.20 ± 1.54 a | 20.38 ± 0.48 a | 14.94 ± 0.21 b | 13.12 ± 0.43 b | 12.60 ± 0.74 b |
| Histidine | 6.09 ± 0.39 a | 5.49 ± 0.06 ab | 5.29 ± 0.25 ab | 5.52 ± 0.08 ab | 4.92 ± 0.21 b |
| Isoleucine | 10.82 ± 0.43 b | 12.12 ± 0.03 a | 11.02 ± 0.33 b | 11.54 ± 0.19 ab | 10.68 ± 0.05 b |
| Leucine | 18.69 ± 0.85 a | 18.88 ± 0.09 a | 17.11 ± 0.50 a | 18.00 ± 0.35 a | 17.13 ± 0.65 a |
| Lysine | 9.41 ± 0.61 b | 10.85 ± 0.09 a | 10.21 ± 0.27 ab | 11.35 ± 0.15 a | 10.03 ± 0.35 ab |
| Methionine | 3.96 ± 0.06 a | 3.51 ± 0.10 ab | 3.26 ± 0.21 ab | 2.97 ± 0.25 b | 3.65 ± 0.24 ab |
| Phenylalanine | 11.61 ± 0.46 ab | 11.98 ± 0.09 a | 10.69 ± 0.32 b | 10.85 ± 0.24 ab | 10.46 ± 0.26 b |
| Threonine | 11.87 ± 0.44 a | 12.35 ± 0.06 a | 11.28 ± 0.33 a | 12.46 ± 0.19 a | 11.73 ± 0.39 a |
| Valine | 14.58 ± 0.66 ab | 15.09 ± 0.36 a | 13.04 ± 0.38 b | 14.11 ± 0.20 ab | 13.05 ± 0.43 b |
| Tryptophan | 1.42 ± 0.32 a | 1.46 ± 0.01 a | 1.50 ± 0.00 a | 1.56 ± 0.08 a | 1.58 ± 0.11 a |
| NEAA | mg/g d.b. | ||||
| Aspartate | 16.99 ± 0.78 a | 17.75 ± 0.09 a | 17.15 ± 0.68 a | 17.02 ± 0.52 a | 16.85 ± 0.38 a |
| Glutamate | 35.27 ± 1.46 a | 32.99 ± 0.94 a | 26.86 ± 0.95 b | 31.96 ± 0.47 a | 26.32 ± 0.77 b |
| Serine | 10.63 ± 0.39 c | 11.85 ± 0.08 ab | 10.57 ± 0.26 c | 12.22 ± 0.24 a | 10.92 ± 0.41 bc |
| Glycine | 15.95 ± 0.77 a | 15.56 ± 0.51 ab | 13.16 ± 0.31 c | 13.94 ± 0.22 bc | 12.96 ± 0.38 c |
| Alanine | 13.35 ± 0.61 b | 16.18 ± 1.57 ab | 16.09 ± 0.82 ab | 18.83 ± 0.28 a | 16.50 ± 0.63 ab |
| Tyrosine | 5.38 ± 0.30 a | 4.34 ± 0.08 a | 4.67 ± 0.78 a | 5.27 ± 0.38 a | 4.18 ± 0.30 a |
| Cystine | 0.40 ± 0.01 b | 0.48 ± 0.01 a | 0.27 ± 0.03 c | 0.44 ± 0.00 ab | 0.23 ± 0.01 c |
| Hydroxyproline | 21.33 ± 2.08 a | 24.56 ± 0.48 a | 25.96 ± 1.10 a | 26.40 ± 0.61 a | 23.31 ± 1.67 a |
| Glutamine | 0.45 ± 0.00 a | 0.42 ± 0.00 ab | 0.41 ± 0.00 abc | 0.37 ± 0.00 c | 0.39 ± 0.02 bc |
| Total AA (mg/g d.b.) | 227.39 ± 12.17 a | 236.24 ± 4.38 a | 213.50 ± 7.73 a | 227.92 ± 3.35 a | 207.47 ± 7.94 a |
| Total AA, with DM loss (mg/g) | 218.78 ± 12.17 ab | 232.89 ± 8.72 a | 194.81 ± 8.29 bc | 211.29 ± 2.73 abc | 192.59 ± 7.72 c |
| Total CP, with DM loss (mg/g) | 311.13 ± 2.56 b | 340.06 ± 8.25 a | 286.35 ± 7.10 c | 306.82 ± 0.47 b | 260.44 ± 0.39 d |
| Ctrl | PO | RO | AO | MC | |
|---|---|---|---|---|---|
| IVDMD, % | 55.39 ± 9.39 a | 45.36 ± 2.39 a | 52.93 ± 4.02 a | 61.09 ± 2.13 a | 53.33 ± 3.99 a |
| IVDAA | % | ||||
| EAA | |||||
| Arginine | 77.28 ± 5.46 a | 60.45 ± 2.17 b | 56.48 ± 5.01 b | 63.47 ± 2.25 ab | 54.00 ± 4.52 b |
| Histidine | 77.93 ± 5.77 a | 54.54 ± 0.83 c | 61.76 ± 4.14 bc | 73.23 ± 0.25 ab | 61.72 ± 4.00 bc |
| Isoleucine | 74.40 ± 8.92 a | 55.52 ± 0.63 a | 59.41 ± 4.03 a | 73.20 ± 0.63 a | 60.70 ± 7.48 a |
| Leucine | 76.49 ± 8.11 a | 54.06 ± 0.46 b | 58.21 ± 3.44 ab | 72.41 ± 0.46 ab | 60.40 ± 6.19 ab |
| Lysine | 74.27 ± 7.36 a | 61.27 ± 1.16 a | 63.25 ± 4.15 a | 77.14 ± 0.83 a | 65.24 ± 2.94 a |
| Methionine | 85.23 ± 5.31 a | 61.71 ± 0.64 b | 70.89 ± 3.23 ab | 81.84 ± 5.97 a | 70.64 ± 0.57 ab |
| Phenylalanine | 76.05 ± 7.55 a | 55.16 ± 0.84 b | 60.99 ± 3.23 ab | 72.45 ± 0.74 ab | 62.23 ± 6.50 ab |
| Threonine | 76.96 ± 7.26 a | 56.98 ± 0.60 b | 62.43 ± 3.36 ab | 75.05 ± 0.87 a | 65.45 ± 5.44 ab |
| Valine | 76.66 ± 8.00 a | 55.76 ± 1.32 b | 59.70 ± 3.26 ab | 73.49 ± 0.84 ab | 61.55 ± 5.73 ab |
| Tryptophan | 77.94 ± 8.65 a | 69.16 ± 0.51 a | 70.13 ± 0.99 a | 79.29 ± 0.44 a | 71.76 ± 5.14 a |
| NEAA | |||||
| Aspartate | 82.58 ± 5.27 a | 64.52 ± 1.92 b | 71.62 ± 4.74 ab | 80.27 ± 2.30 ab | 73.96 ± 4.58 ab |
| Glutamate | 81.48 ± 4.68 a | 61.26 ± 2.06 c | 65.95 ± 2.98 c | 78.99 ± 0.59 ab | 67.32 ± 3.88 bc |
| Serine | 75.23 ± 7.48 a | 56.69 ± 0.89 b | 59.97 ± 3.31 ab | 74.21 ± 0.98 ab | 63.66 ± 5.80 ab |
| Glycine | 73.43 ± 6.61 a | 49.14 ± 3.26 b | 51.48 ± 3.81 b | 66.00 ± 1.31 ab | 53.15 ± 4.42 b |
| Alanine | 76.77 ± 7.54 a | 56.51 ± 4.03 a | 58.70 ± 5.69 a | 76.86 ± 0.74 a | 63.55 ± 6.03 a |
| Tyrosine | 82.94 ± 6.35 a | 62.60 ± 2.98 a | 72.73 ± 8.53 a | 83.86 ± 1.41 a | 72.49 ± 5.65 a |
| Cystine | 79.37 ± 5.47 a | 55.15 ± 0.56 ab | 49.53 ± 7.69 b | 78.62 ± 0.64 a | 52.36 ± 10.14 b |
| Hydroxyproline | 72.43 ± 7.73 a | 61.23 ± 4.03 a | 65.71 ± 4.49 a | 76.56 ± 0.75 a | 63.45 ± 4.81 a |
| Glutamine | 86.38 ± 19.27 a | 48.47 ± 1.09 a | 61.63 ± 1.09 a | 63.08 ± 4.01 a | 83.15 ± 23.82 a |
| Total | 77.35 ± 6.63 a | 58.23 ± 1.81 b | 62.20 ± 4.11 ab | 74.90 ± 0.91 ab | 63.58 ± 4.99 ab |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sun, X.; Boardman, K.; Marks, D.; Wyse, D.L.; Hu, B. Fungal Bioprocessing to Improve Quality of Pennycress Meal as a Potential Feeding Ingredient for Monogastric Animals. Fermentation 2023, 9, 732. https://doi.org/10.3390/fermentation9080732
Sun X, Boardman K, Marks D, Wyse DL, Hu B. Fungal Bioprocessing to Improve Quality of Pennycress Meal as a Potential Feeding Ingredient for Monogastric Animals. Fermentation. 2023; 9(8):732. https://doi.org/10.3390/fermentation9080732
Chicago/Turabian StyleSun, Xiao, Kristin Boardman, David Marks, Donald L. Wyse, and Bo Hu. 2023. "Fungal Bioprocessing to Improve Quality of Pennycress Meal as a Potential Feeding Ingredient for Monogastric Animals" Fermentation 9, no. 8: 732. https://doi.org/10.3390/fermentation9080732
APA StyleSun, X., Boardman, K., Marks, D., Wyse, D. L., & Hu, B. (2023). Fungal Bioprocessing to Improve Quality of Pennycress Meal as a Potential Feeding Ingredient for Monogastric Animals. Fermentation, 9(8), 732. https://doi.org/10.3390/fermentation9080732