
Volatile Flavor Improvement and Spoilage Microorganism Inhibition in Low-Salt Fish Sauce (Yulu) by Salt-Tolerant Bacillus subtilis




{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Microbial Starter and Pre-Culture
2.2. Yulu Processing
2.3. Volatile Compound Analysis
2.4. Microbial Community Analysis
2.5. Statistical Analysis
3. Results and Discussion
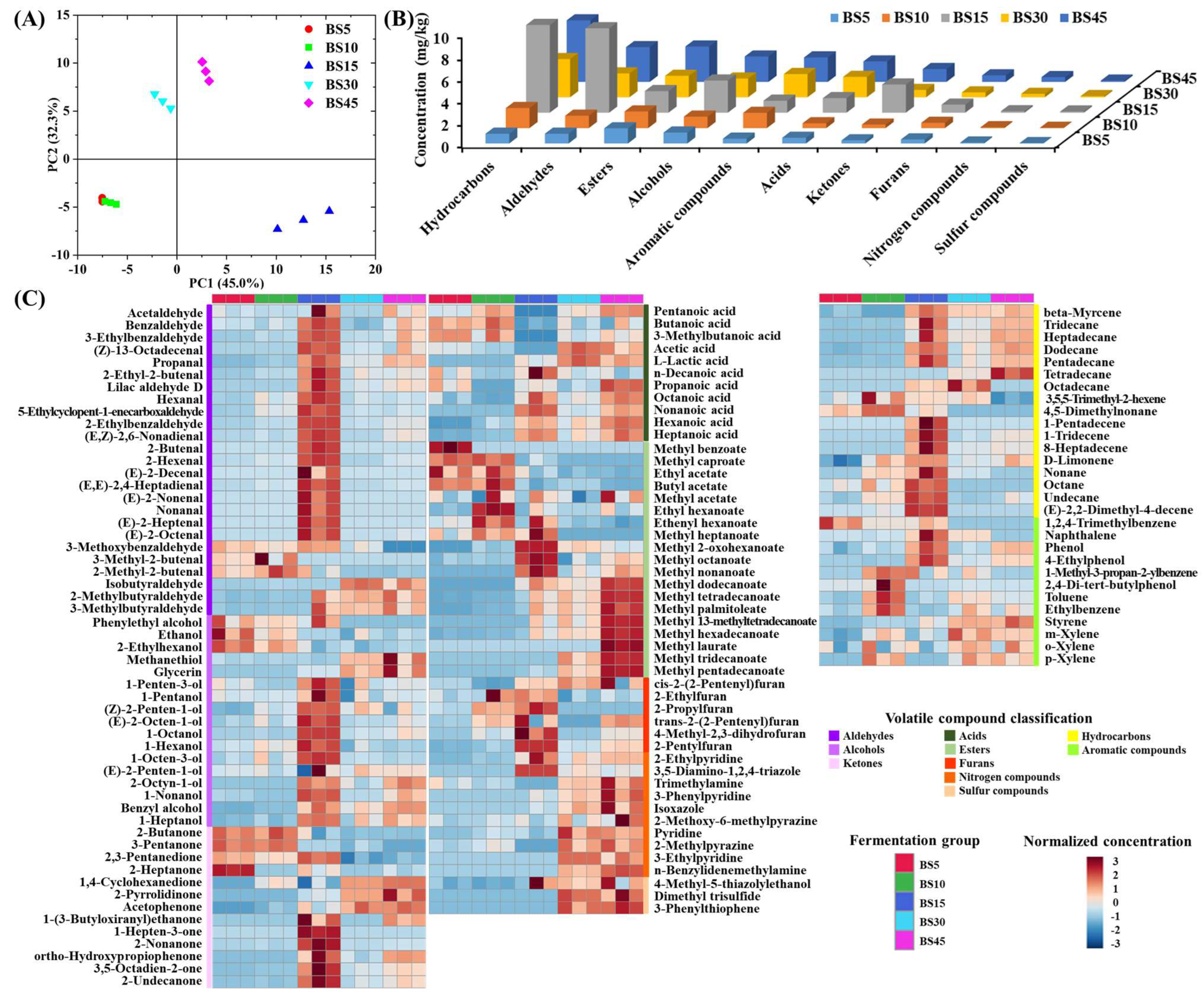
3.1. Change in Volatile Compounds during Yulu Fermentation
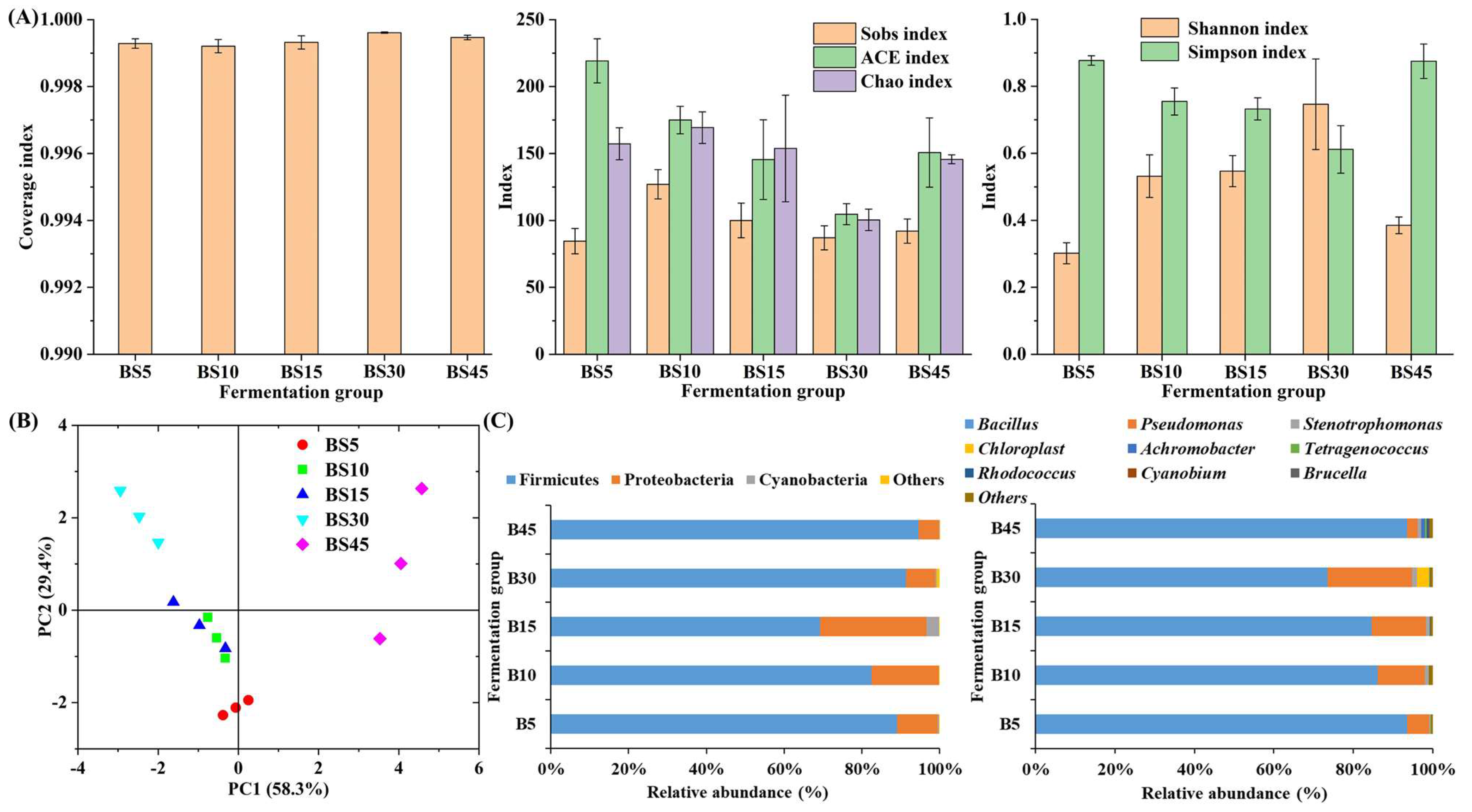
3.2. Change in Microbial Community during Yulu Fermentation
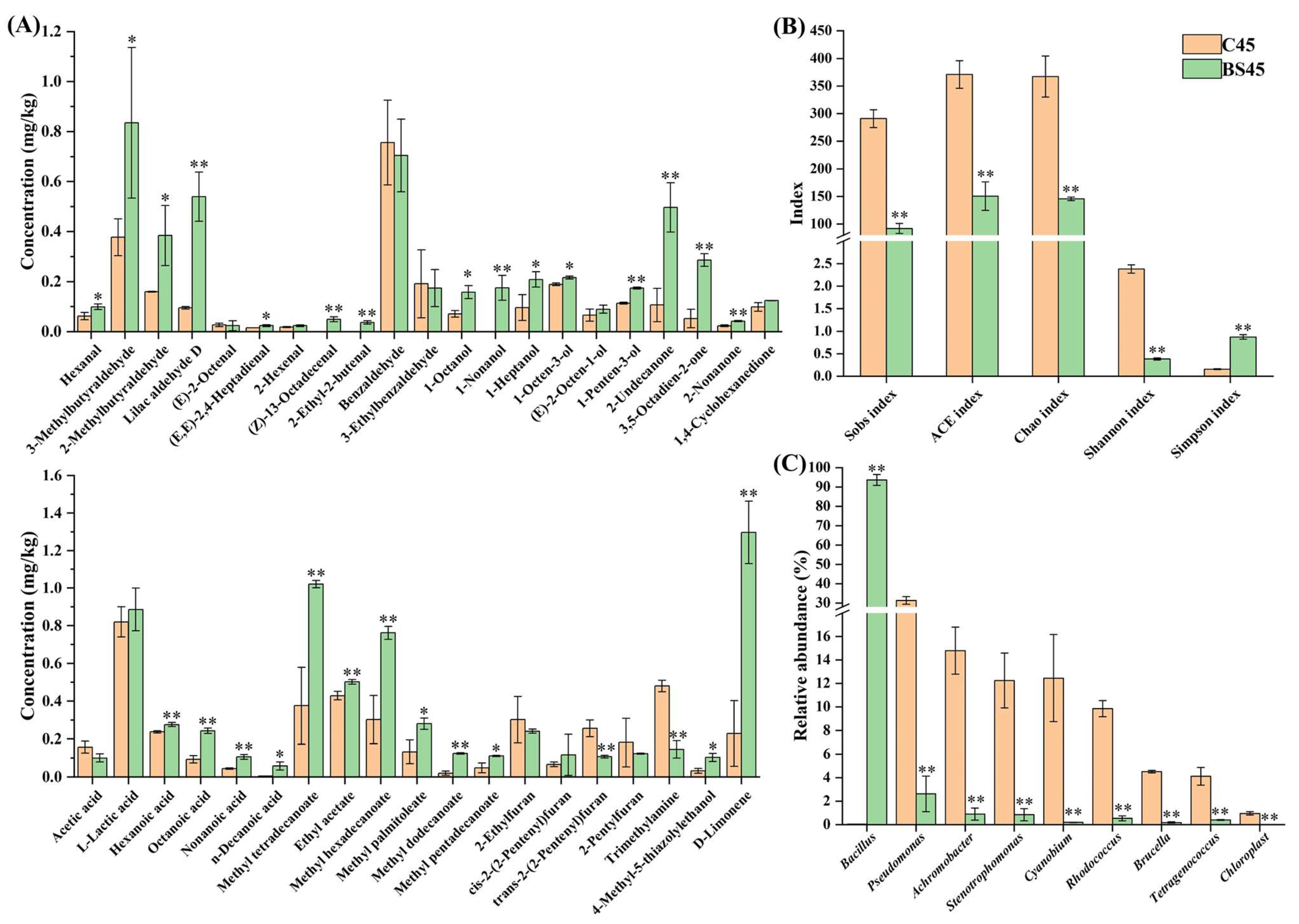
3.3. Improvement in Volatile Flavor of Yulu after B. subtilis Addition
3.4. Inhibition of Spoilage Microorganisms in Yulu after B. subtilis Addition
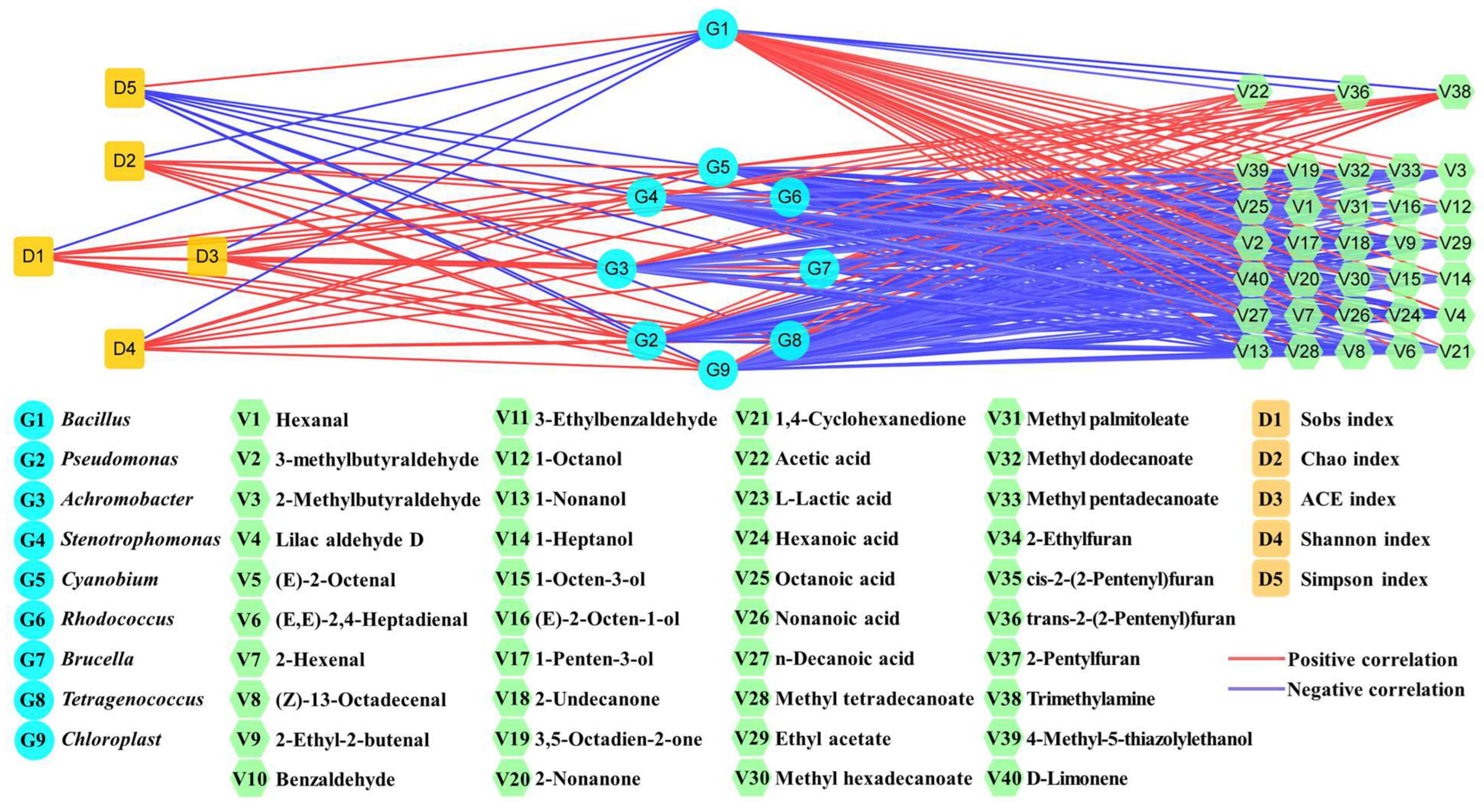
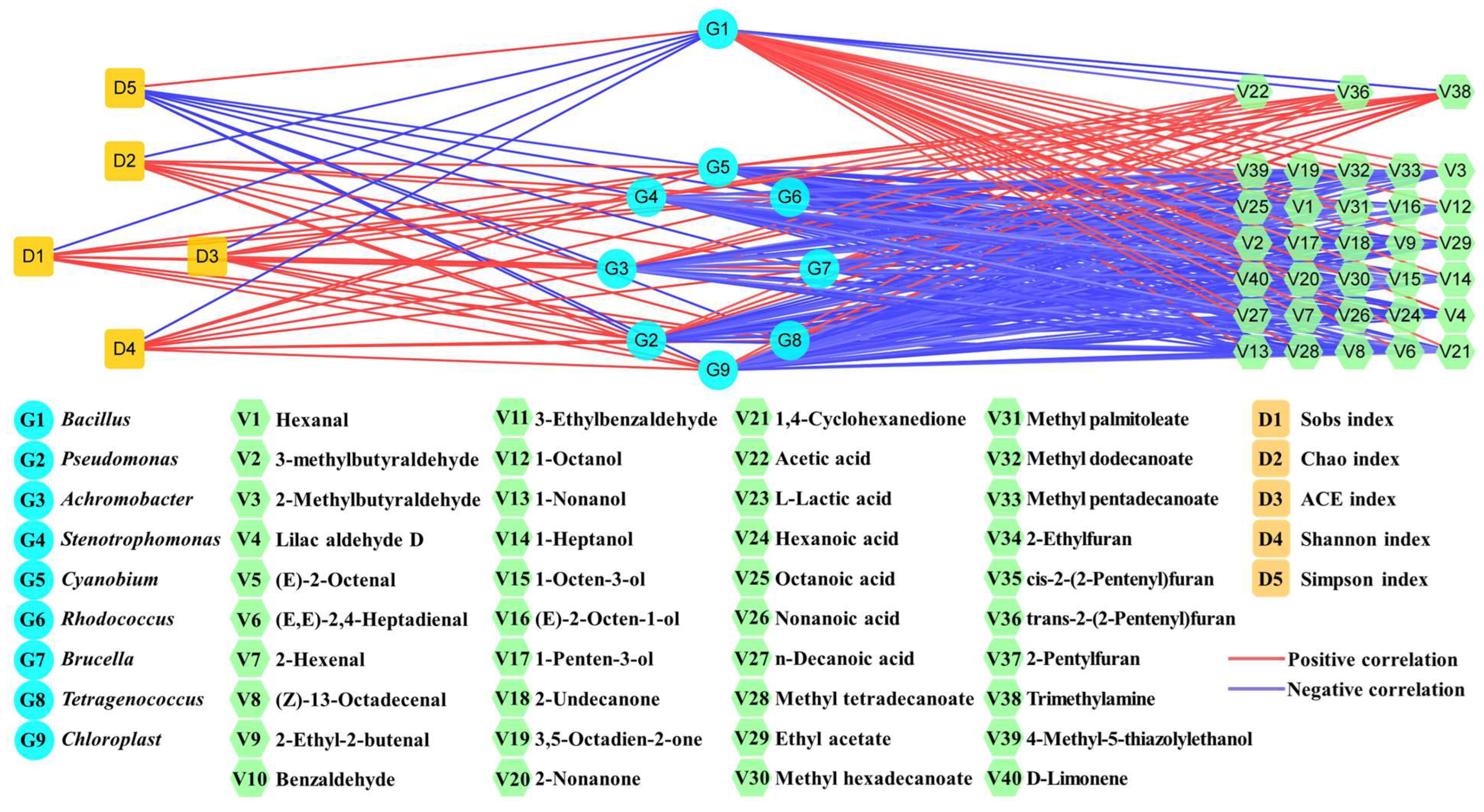
3.5. Change Mechanism of Volatile Flavors and Spoilage Microorganisms in Yulu after B. subtilis Addition
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kanjan, P.; Sakpetch, P.; Masniyom, P.; Hongpattarakere, T. Quality characteristics of high salt fermented fish sauce (budu) produced using autochthonous Virgibacillus halodenitrificans PS21 and Staphylococcus simulans PMRS35. Int. J. Food Sci. Technol. 2021, 56, 4098–4107. [Google Scholar] [CrossRef]
- Nguyen, A.Q.; Sekar, A.; Kim, M.; Nguyen, L.P.; Le, N.T.; Uh, S.; Hong, S.; Kim, K. Fish sauce fermentation using Marinococcus halotolerans SPQ isolate as a starter culture. Food Sci. Nutr. 2021, 9, 651–661. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Li, C.; Li, L.; Yang, X.; Chen, S.; Wu, Y.; Zhao, Y.; Wang, J.; Wei, Y.; Yang, D. Application of UHPLC-Q/TOF-MS-based metabolomics in the evaluation of metabolites and taste quality of Chinese fish sauce (Yu-lu) during fermentation. Food Chem. 2019, 296, 132–141. [Google Scholar] [CrossRef]
- Li, Y.; Li, W.; Li, C.; Li, L.; Yang, D.; Wang, Y.; Chen, S.; Wang, D.; Wu, Y. Novel insight into flavor and quality formation in naturally fermented low-salt fish sauce based on microbial metabolism. Food Res. Int. 2023, 166, 112586. [Google Scholar] [CrossRef]
- Li, C.; Li, W.; Li, L.; Chen, S.; Wu, Y.; Qi, B. Microbial community changes induced by a newly isolated salt-tolerant Tetragenococcus muriaticus improve the volatile flavor formation in low-salt fish sauce. Food Res. Int. 2022, 156, 111153. [Google Scholar] [CrossRef] [PubMed]
- Noma, S.; Koyanagi, L.; Kawano, S.; Hayashi, N. Application of pressurized carbon dioxide during salt-reduced sardine fish sauce production. Food Sci. Technol. Res. 2020, 26, 195–204. [Google Scholar] [CrossRef]
- Gao, R.; Zhou, J.; Leng, W.; Shi, T.; Jin, W.; Yuan, L. Screening of a Planococcus bacterium producing a cold-adapted protease and its application in low-salt fish sauce fermentation. J. Food Process Preserv. 2020, 44, e14625. [Google Scholar] [CrossRef]
- Akolkar, A.V.; Durai, D.; Desai, A.J. Halobacterium sp SP1(1) as a starter culture for accelerating fish sauce fermentation. J. Appl. Microbiol. 2010, 109, 44–53. [Google Scholar] [CrossRef]
- Xie, L.; Xiao, Y.; Gao, X. Purification and characterization of a halotolerant alkaline serine protease from Penicillium citrinum YL-1 isolated from traditional Chinese fish sauce. Food Biotechnol. 2016, 30, 137–153. [Google Scholar] [CrossRef]
- Kamala, K.; Sivaperumal, P.; Paray, B.A.; Al-Sadoon, M.K. Identification of haloarchaea during fermentation of Sardinella longiceps for being the starter culture to accelerate fish sauce production. Int. J. Food Sci. Technol. 2021, 56, 5717–5725. [Google Scholar] [CrossRef]
- Wakinaka, T.; Iwata, S.; Takeishi, Y.; Watanabe, J.; Mogi, Y.; Tsukioka, Y.; Shibata, Y. Isolation of halophilic lactic acid bacteria possessing aspartate decarboxylase and application to fish sauce fermentation starter. Int. J. Food Microbiol. 2019, 292, 137–143. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Li, C.; Li, L.; Wang, Y.; Chen, S.; Yang, S. Isolation, identification and enzyme-producing condition of a salt-tolerant strain with high protease production. Food Ferment. Ind. 2021, 47, 134–142. [Google Scholar]
- Li, C.; Liu, D.; Yang, X.; Chen, S.; Deng, J.; Rong, H.; Li, C. Efficient biodegradation of malachite green by a newly isolated Klebsiella pneumoniae strain WA-1. Environ. Prog. Sustain. Energy 2020, 39, e13346. [Google Scholar] [CrossRef]
- Wang, Y.; Li, C.; Zhao, Y.; Li, L.; Yang, X.; Wu, Y.; Chen, S.; Cen, J.; Yang, S.; Yang, D. Novel insight into the formation mechanism of volatile flavor in Chinese fish sauce (Yu-lu) based on molecular sensory and metagenomics analyses. Food Chem. 2020, 323, 126839. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Wang, Y.; Li, C.; Li, L.; Yang, X.; Wu, Y.; Chen, S.; Zhao, Y. Novel insight into physicochemical and flavor formation in naturally fermented tilapia sausage based on microbial metabolic network. Food Res. Int. 2021, 141, 110122. [Google Scholar] [CrossRef]
- Li, C.; Zhao, Y.; Wang, Y.; Li, L.; Yang, X.; Chen, S.; Zhao, Y.; Zhou, W. Microbial community changes induced by Pediococcus pentosaceus improve the physicochemical properties and safety in fermented tilapia sausage. Food Res. Int. 2021, 147, 110476. [Google Scholar] [CrossRef]
- Yang, D.; Li, C.; Li, L.; Wang, Y.; Wu, Y.; Chen, S.; Zhao, Y.; Wei, Y.; Wang, D. Novel insight into the formation mechanism of umami peptides based on microbial metabolism in Chouguiyu, a traditional Chinese fermented fish. Food Res. Int. 2022, 157, 111211. [Google Scholar] [CrossRef]
- Feng, Y.; Su, G.; Zhao, H.; Cai, Y.; Cui, C.; Sun-Waterhouse, D.; Zhao, M. Characterisation of aroma profiles of commercial soy sauce by odour activity value and omission test. Food Chem. 2015, 167, 220–228. [Google Scholar] [CrossRef]
- Shen, Y.; Wu, Y.; Wang, Y.; Li, L.; Li, C.; Zhao, Y.; Yang, S. Contribution of autochthonous microbiota succession to flavor formation during Chinese fermented mandarin fish (Siniperca chuatsi). Food Chem. 2021, 348, 129107. [Google Scholar] [CrossRef]
- Bao, R.; Liu, S.; Ji, C.; Liang, H.; Yang, S.; Yan, X.; Zhou, Y.; Lin, X.; Zhu, B. Shortening fermentation period and quality improvement of fermented fish, Chouguiyu, by co-inoculation of Lactococcus lactis M10 and Weissella cibaria M3. Front. Microbiol. 2018, 9, 3003. [Google Scholar] [CrossRef]
- Yang, J.; Jiang, C.; Bao, R.; Liu, M.; Lv, J.; Yang, Z.; Xu, W.; Liang, H.; Ji, C.; Li, S.; et al. Effects of flavourzyme addition on physicochemical properties, volatile compound components and microbial community succession of Suanzhayu. Int. J. Food Microbiol. 2020, 334, 108839. [Google Scholar] [CrossRef] [PubMed]
- Zeng, X.; Xia, W.; Jiang, Q.; Xu, Y.; Fan, J. Contribution of mixed starter cultures to flavor profile of Suanyu—A traditional Chinese low-salt fermented whole fish. J. Food Process Preserv. 2017, 41, e13131. [Google Scholar] [CrossRef]
- Zhang, Q.; Ding, Y.; Gu, S.; Zhu, S.; Zhou, X.; Ding, Y. Identification of changes in volatile compounds in dry-cured fish during storage using HS-GC-IMS. Food Res. Int. 2020, 137, 109339. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Zhang, L.; Liu, Q.; Wang, Y.; Chen, Q.; Kong, B. The potential correlation between bacterial diversity and the characteristic volatile flavour of traditional dry sausages from Northeast China. Food Microbiol. 2020, 91, 103505. [Google Scholar] [CrossRef]
- Zhu, W.; Luan, H.; Bu, Y.; Li, X.; Li, J.; Ji, G. Flavor characteristics of shrimp sauces with different fermentation and storage time. LWT-Food Sci. Technol. 2019, 110, 142–151. [Google Scholar] [CrossRef]
- Shi, J.; Nian, Y.; Da, D.; Xu, X.; Zhou, G.; Zhao, D.; Li, C. Characterization of flavor volatile compounds in sauce spareribs by gas chromatography-mass spectrometry and electronic nose. LWT-Food Sci. Technol. 2020, 124, 109182. [Google Scholar] [CrossRef]
- Sha, K.; Lang, Y.M.; Sun, B.Z.; Su, H.W.; Li, H.P.; Zhang, L.; Lei, Y.H.; Li, H.B.; Zhang, Y. Changes in lipid oxidation, fatty acid profile and volatile compounds of traditional Kazakh dry-cured beef during processing and storage. J. Food Process Preserv. 2017, 41, e13059. [Google Scholar] [CrossRef]
- Lee, S.H.; Jung, J.Y.; Jeon, C.O. Bacterial community dynamics and metabolite changes in myeolchi-aekjeot, a Korean traditional fermented fish sauce, during fermentation. Int. J. Food Microbiol. 2015, 203, 15–22. [Google Scholar] [CrossRef]
- Li, C.; Zhao, Y.; Wang, Y.; Li, L.; Huang, J.; Yang, X.; Chen, S.; Zhao, Y. Contribution of microbial community to flavor formation in tilapia sausage during fermentation with Pediococcus pentosaceus. LWT-Food Sci. Technol. 2022, 154, 112628. [Google Scholar] [CrossRef]
- Zhang, J.; Cao, J.; Pei, Z.; Wei, P.; Xiang, D.; Cao, X.; Shen, X.; Li, C. Volatile flavour components and the mechanisms underlying their production in golden pompano (Trachinotus blochii) fillets subjected to different drying methods: A comparative study using an electronic nose, an electronic tongue and SDE-GC-MS. Food Res. Int. 2019, 123, 217–225. [Google Scholar] [CrossRef]
- Aregbe, A.Y.; Mu, T.; Sun, H. Effect of different pretreatment on the microbial diversity of fermented potato revealed by high-throughput sequencing. Food Chem. 2019, 290, 125–134. [Google Scholar] [CrossRef] [PubMed]
- Serra, J.L.; Moura, F.G.; de Melo Pereira, G.V.; Soccol, C.R.; Rogez, H.; Darnet, S. Determination of the microbial community in Amazonian cocoa bean fermentation by Illumina-based metagenomic sequencing. LWT-Food Sci. Technol. 2019, 106, 229–239. [Google Scholar] [CrossRef]
- Xu, Y.; He, L.; Xia, W.; Jiang, Q.; Yang, F.; Gao, P.; Wang, B. The impact of fermentation at elevated temperature on quality attributes and biogenic amines formation of low-salt fermented fish. Int. J. Food Sci. Technol. 2019, 54, 723–733. [Google Scholar] [CrossRef]
- Wang, Y.; Shen, Y.; Wu, Y.; Li, C.; Li, L.; Zhao, Y.; Hu, X.; Wei, Y.; Huang, H. Comparison of the microbial community and flavor compounds in fermented mandarin fish (Siniperca chuatsi): Three typical types of Chinese fermented mandarin fish products. Food Res. Int. 2021, 144, 110365. [Google Scholar] [CrossRef] [PubMed]
- Belleggia, L.; Aquilanti, L.; Ferrocino, I.; Milanović, V.; Garofalo, C.; Clementi, F.; Cocolin, L.; Mozzon, M.; Foligni, R.; Haouet, M.N.; et al. Discovering microbiota and volatile compounds of surströmming, the traditional Swedish sour herring. Food Microbiol. 2020, 91, 103503. [Google Scholar] [CrossRef]
- Yang, D.; Li, C.; Li, L.; Yang, X.; Chen, S.; Wu, Y.; Feng, Y. Novel Insight into the formation and inhibition mechanism of dipeptidyl peptidase-Ⅳ inhibitory peptides from fermented mandarin fish (Chouguiyu). Food Sci. Hum. Wellness 2023, 12, 2408–2416. [Google Scholar] [CrossRef]
- Li, C.; Sun, W.; Liu, S.; Pan, C.; Wang, D.; Feng, Y.; Cen, J.; Chen, S. Efficient aluminum removal and microorganism inhibition in ready-to-eat jellyfish by slightly acidic electrolyzed water. Food Biosci. 2023, 53, 102599. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, C.; Li, L.; Chen, S.; Zhao, Y.; Wu, Y. Volatile Flavor Improvement and Spoilage Microorganism Inhibition in Low-Salt Fish Sauce (Yulu) by Salt-Tolerant Bacillus subtilis. Fermentation 2023, 9, 515. https://doi.org/10.3390/fermentation9060515
Li C, Li L, Chen S, Zhao Y, Wu Y. Volatile Flavor Improvement and Spoilage Microorganism Inhibition in Low-Salt Fish Sauce (Yulu) by Salt-Tolerant Bacillus subtilis. Fermentation. 2023; 9(6):515. https://doi.org/10.3390/fermentation9060515
Chicago/Turabian StyleLi, Chunsheng, Laihao Li, Shengjun Chen, Yongqiang Zhao, and Yanyan Wu. 2023. "Volatile Flavor Improvement and Spoilage Microorganism Inhibition in Low-Salt Fish Sauce (Yulu) by Salt-Tolerant Bacillus subtilis" Fermentation 9, no. 6: 515. https://doi.org/10.3390/fermentation9060515
APA StyleLi, C., Li, L., Chen, S., Zhao, Y., & Wu, Y. (2023). Volatile Flavor Improvement and Spoilage Microorganism Inhibition in Low-Salt Fish Sauce (Yulu) by Salt-Tolerant Bacillus subtilis. Fermentation, 9(6), 515. https://doi.org/10.3390/fermentation9060515