
The Potential Use of Endophyte-Free inebrians as Sheep Feed Evaluated with In Vitro Fermentation



Abstract
:1. Introduction
2. Materials and Methods
2.1. Origin and Preparation of EF
2.2. Rumen Fluid Donors and Management
2.3. In Vitro Fermentation Assay
2.4. DNA Extraction and Sequencing
2.5. Calculations and Statistical Analyses
3. Results
3.1. Digestibility of DM, GP, and Fermentation Characteristics
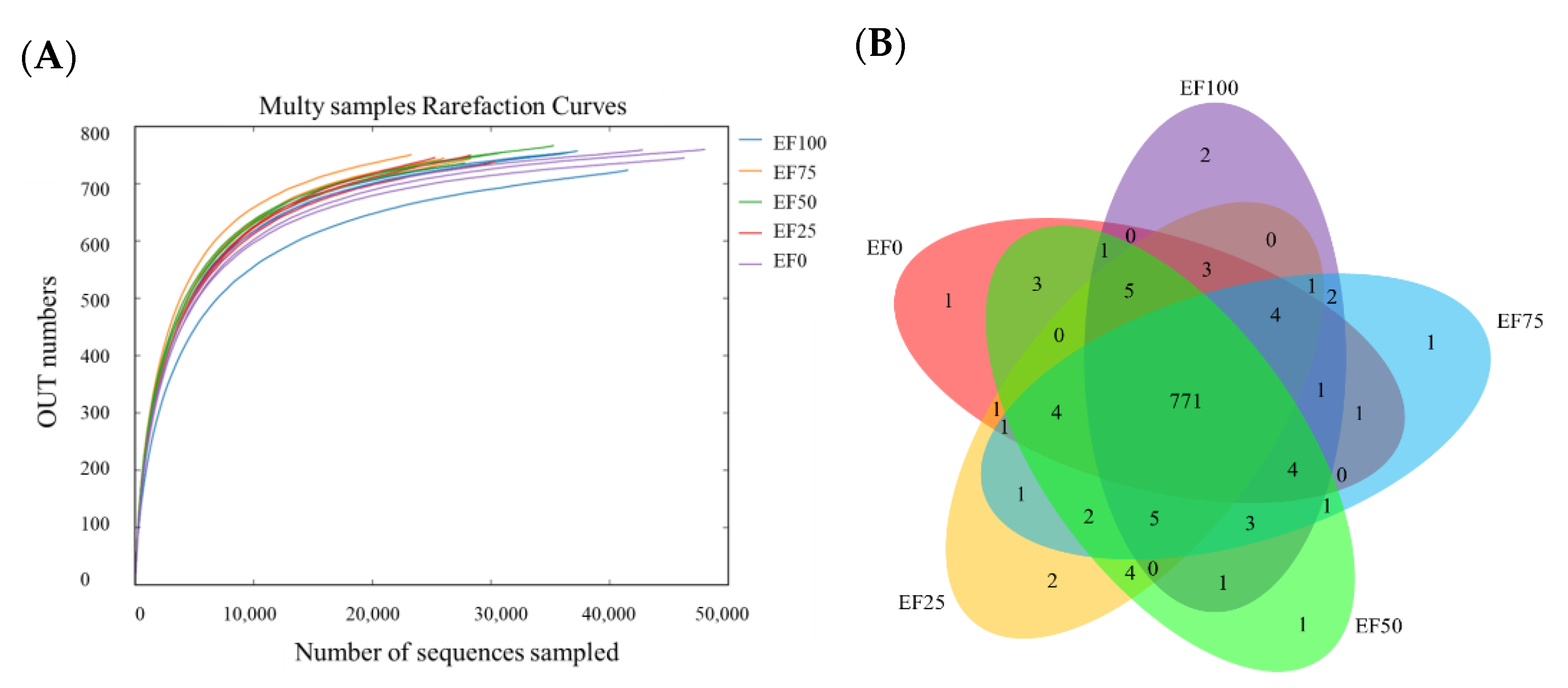
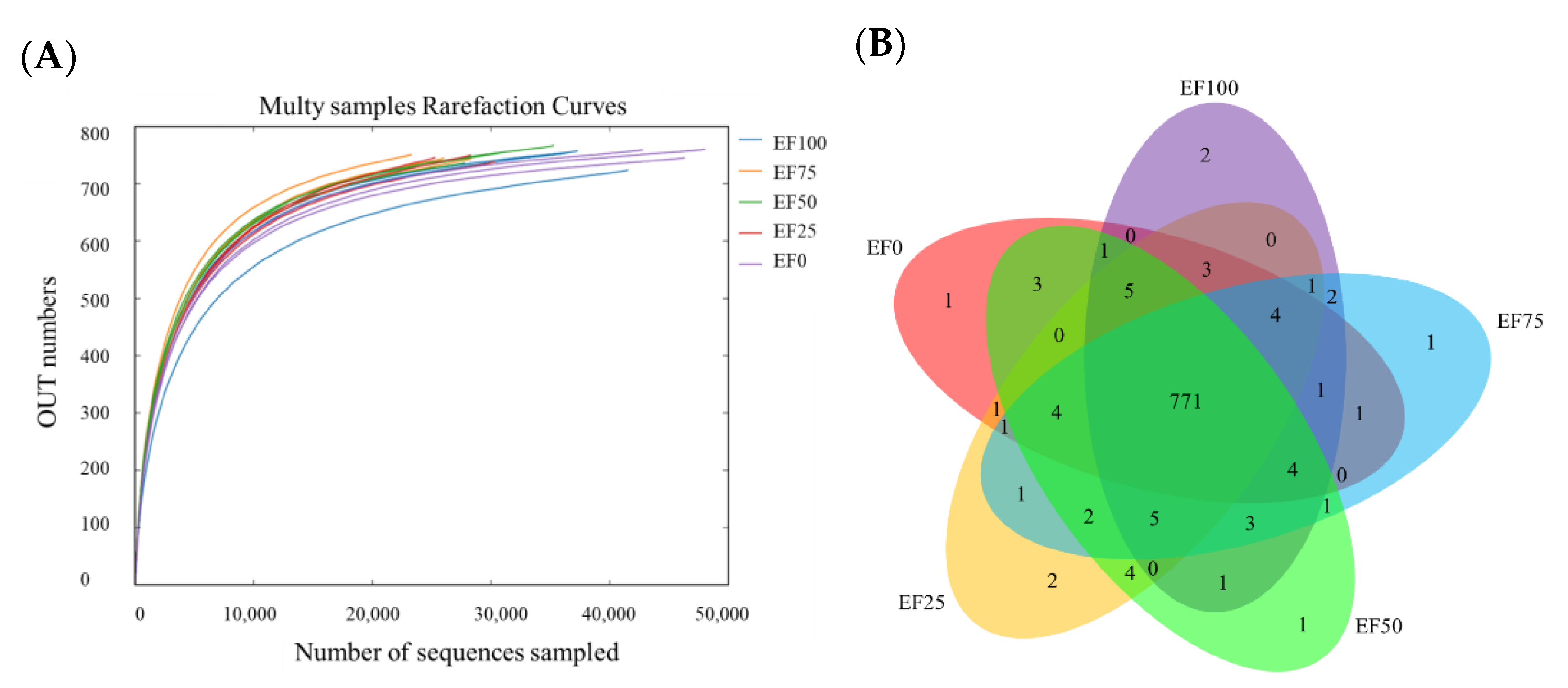
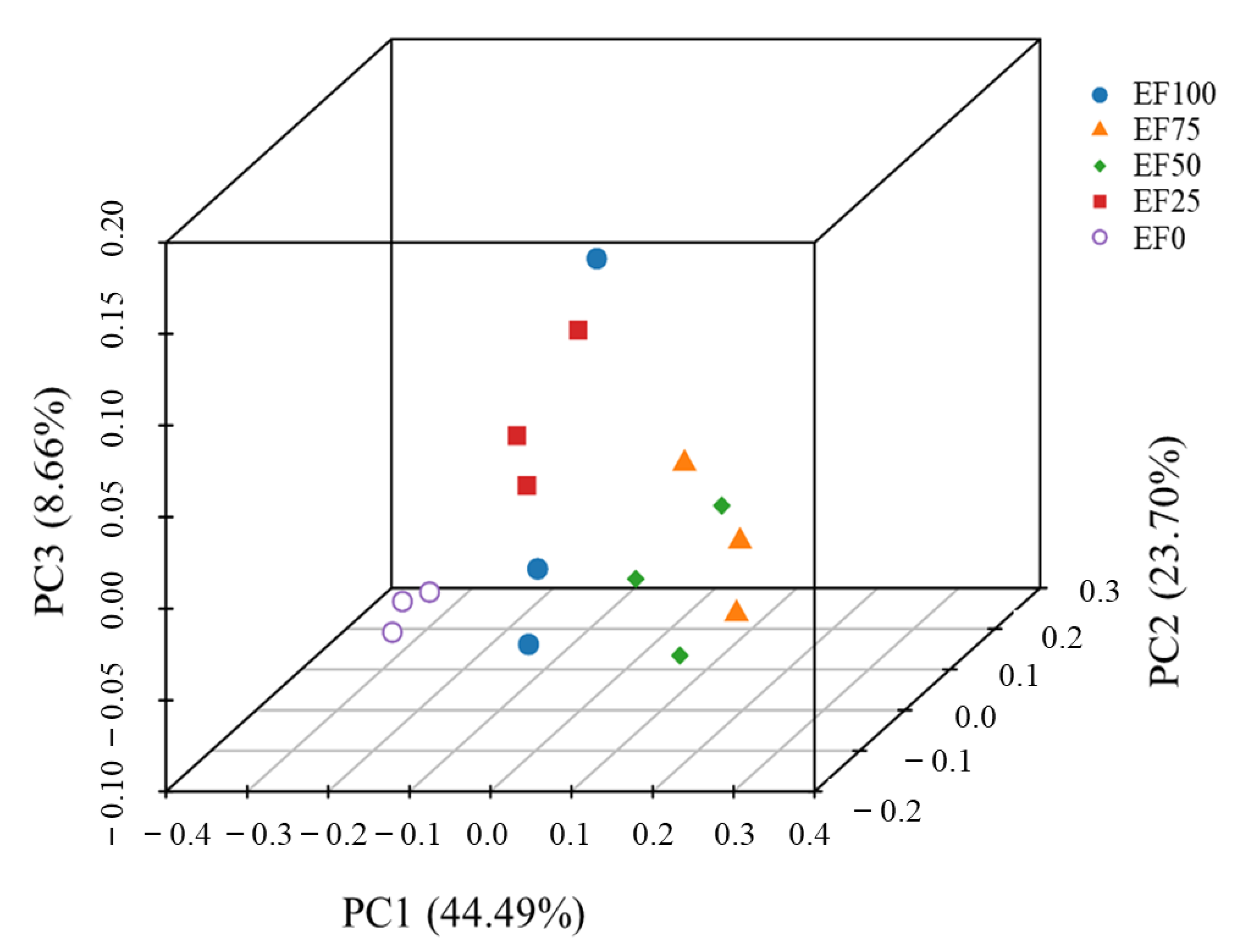
3.2. Diversity of Rumen Microbiota Community in the Sheep
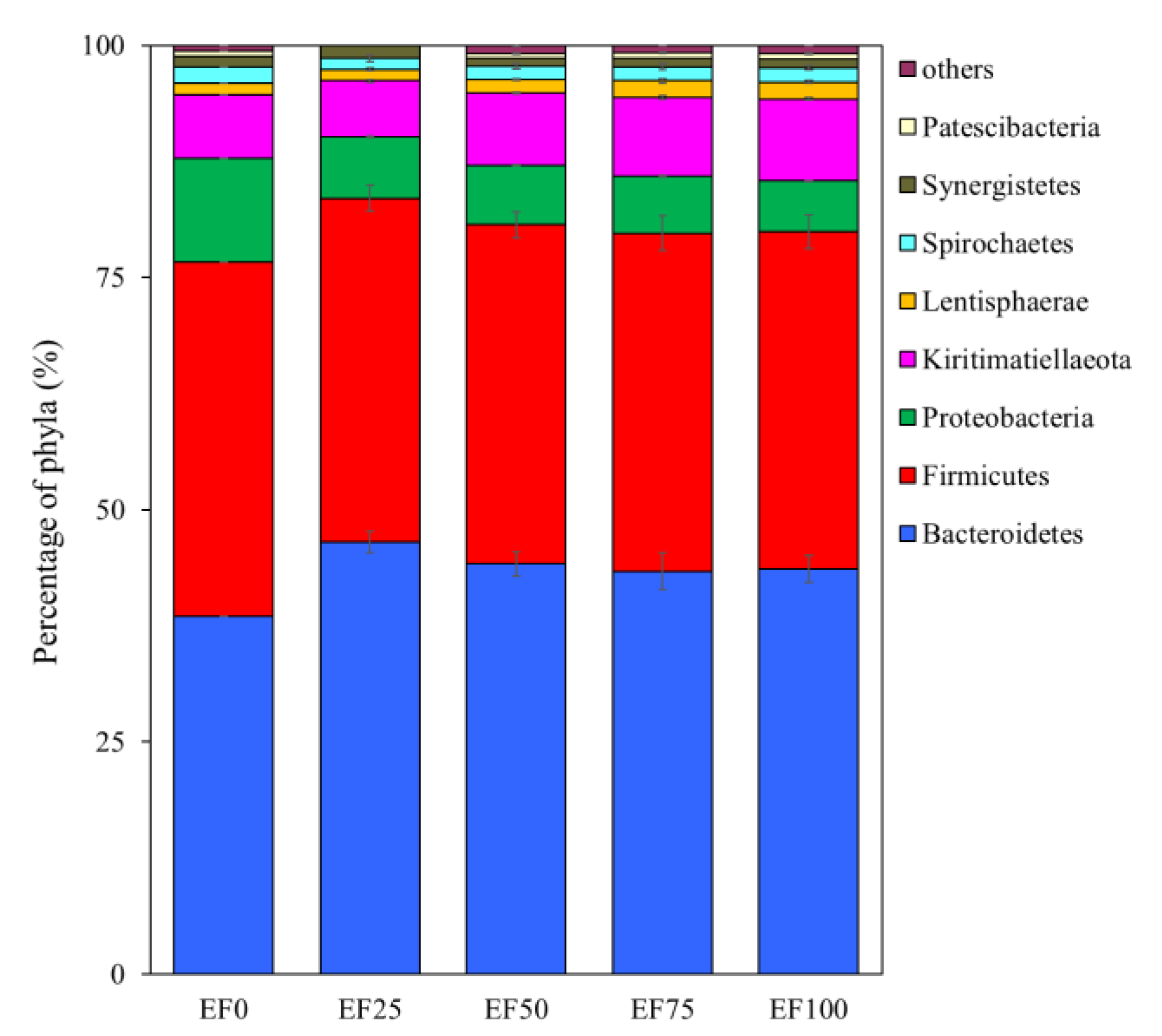
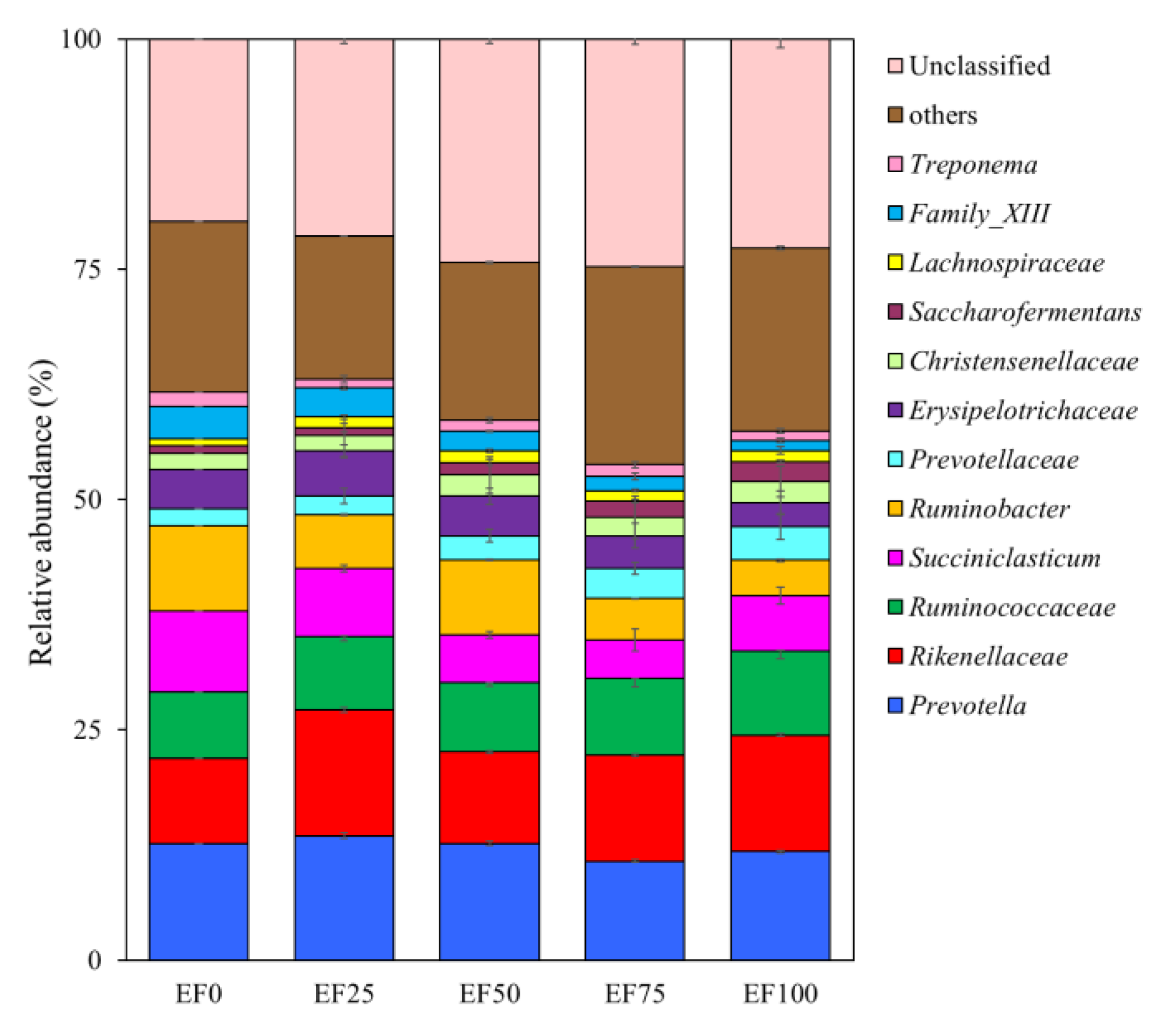
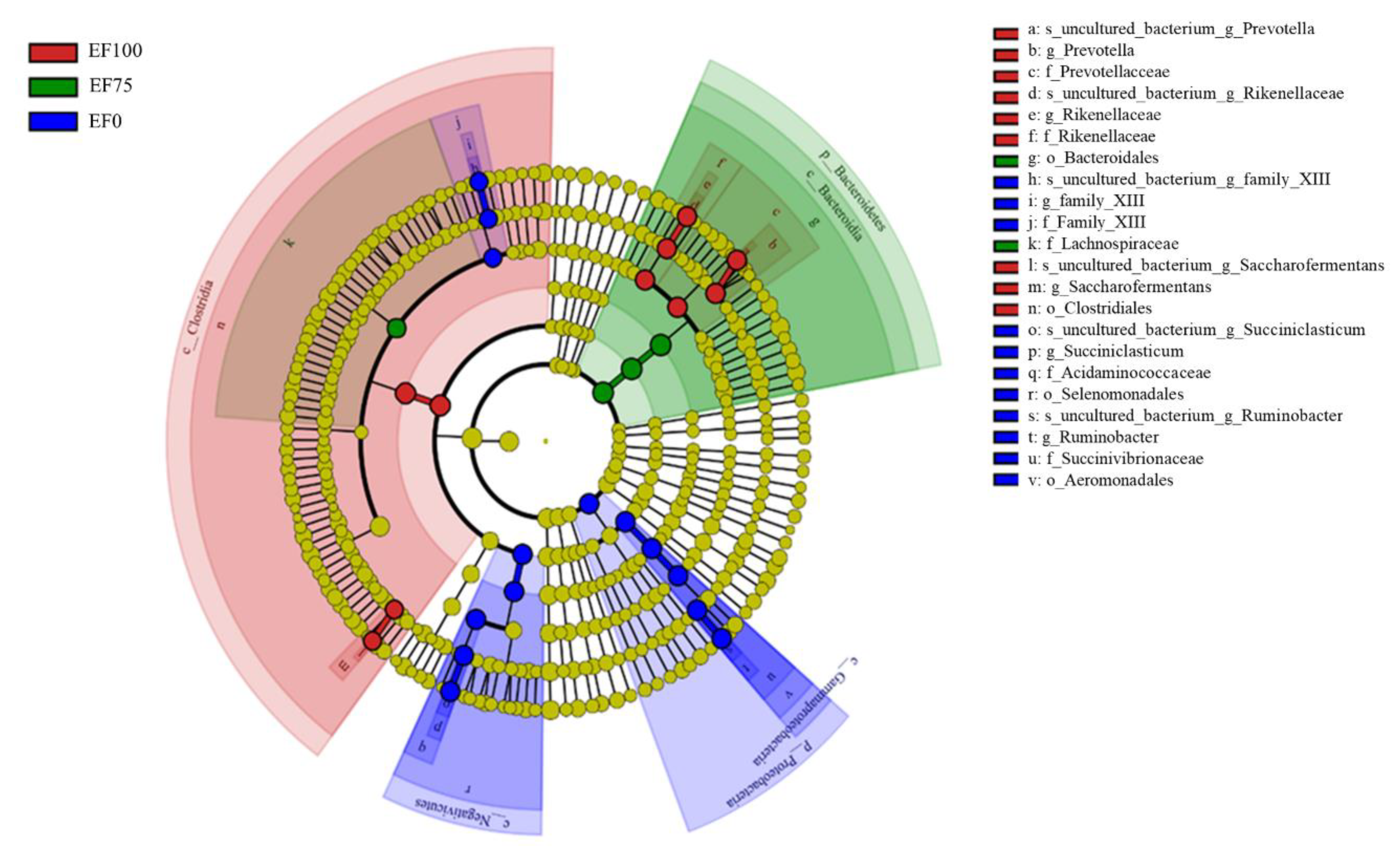
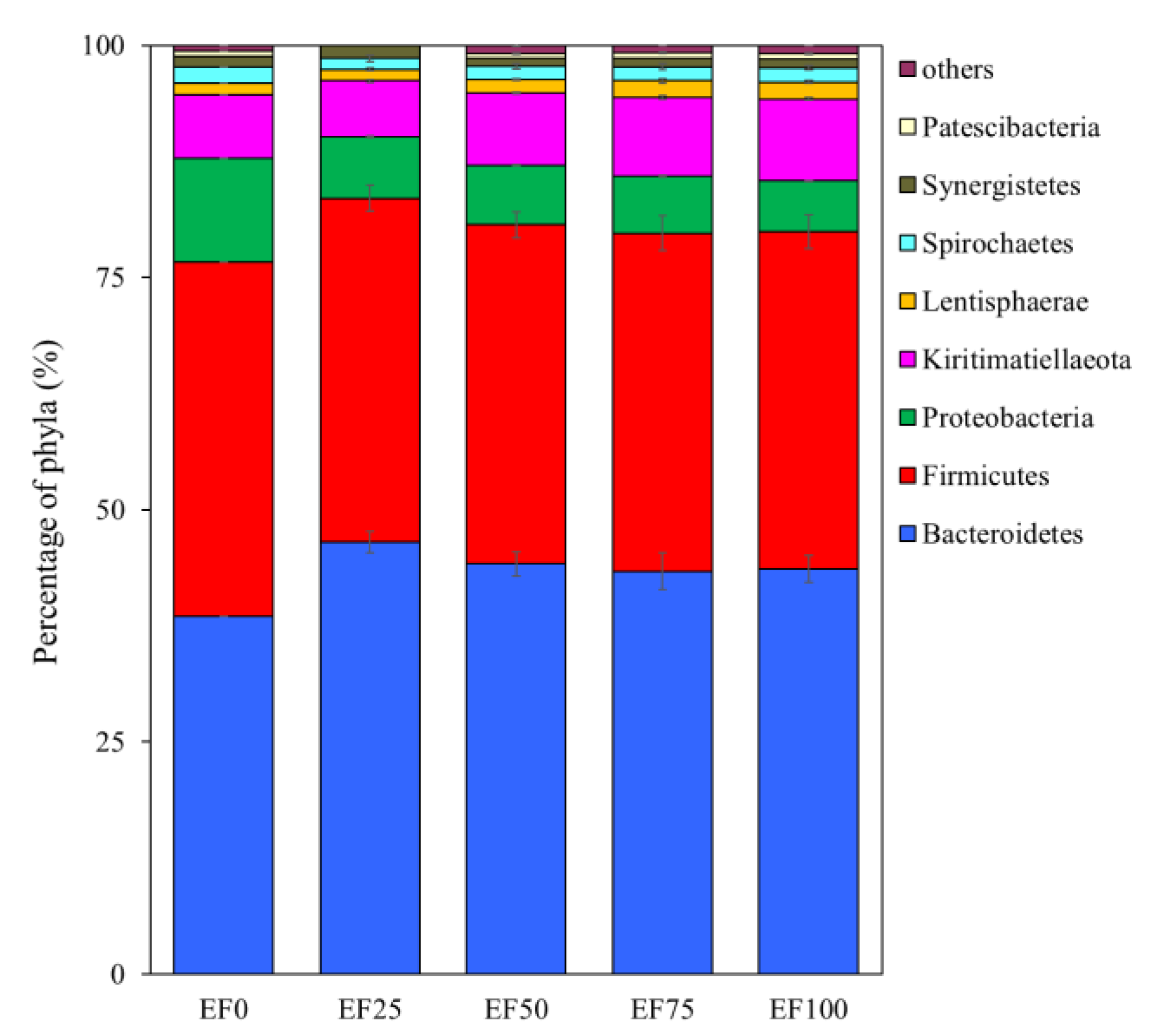
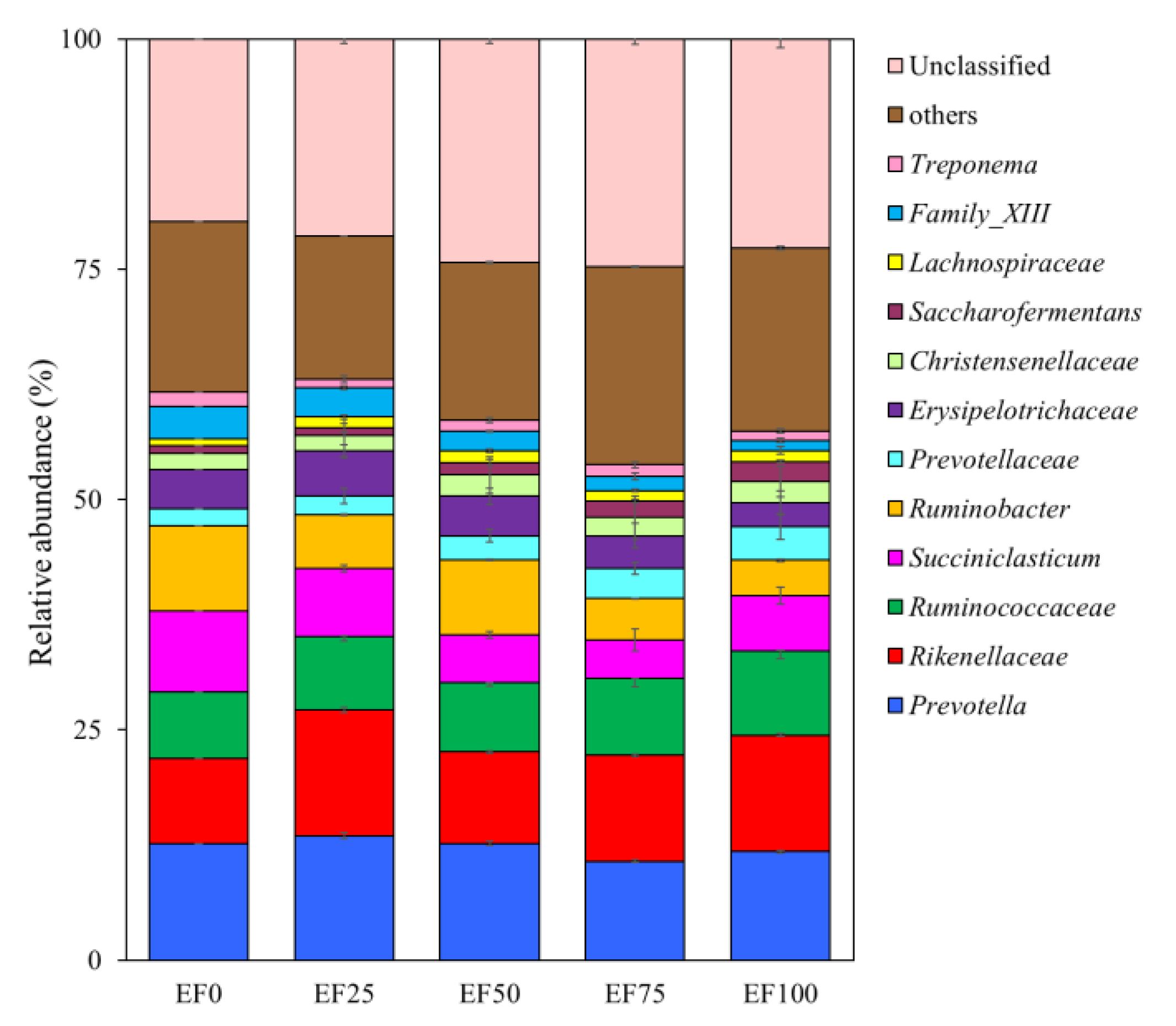
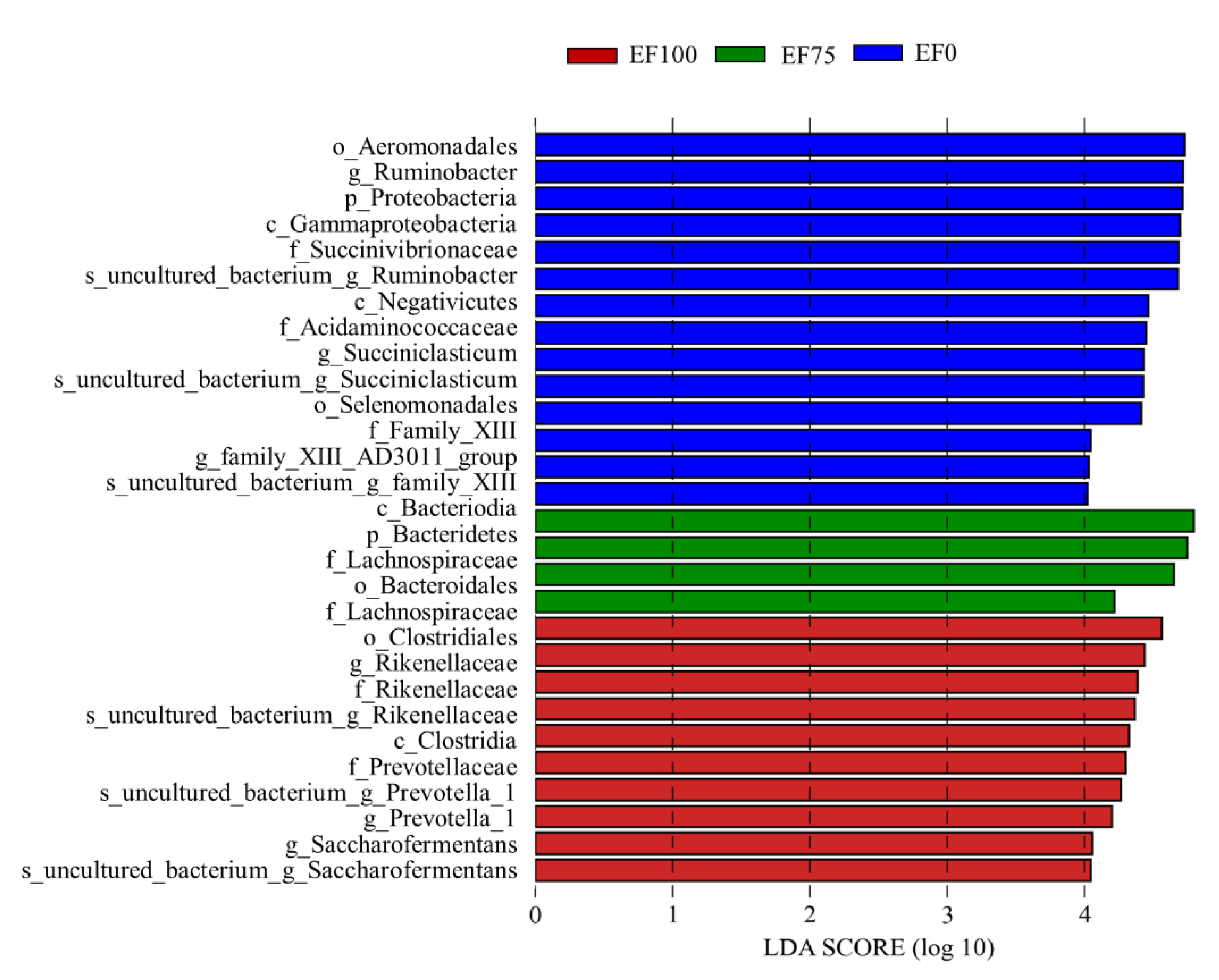
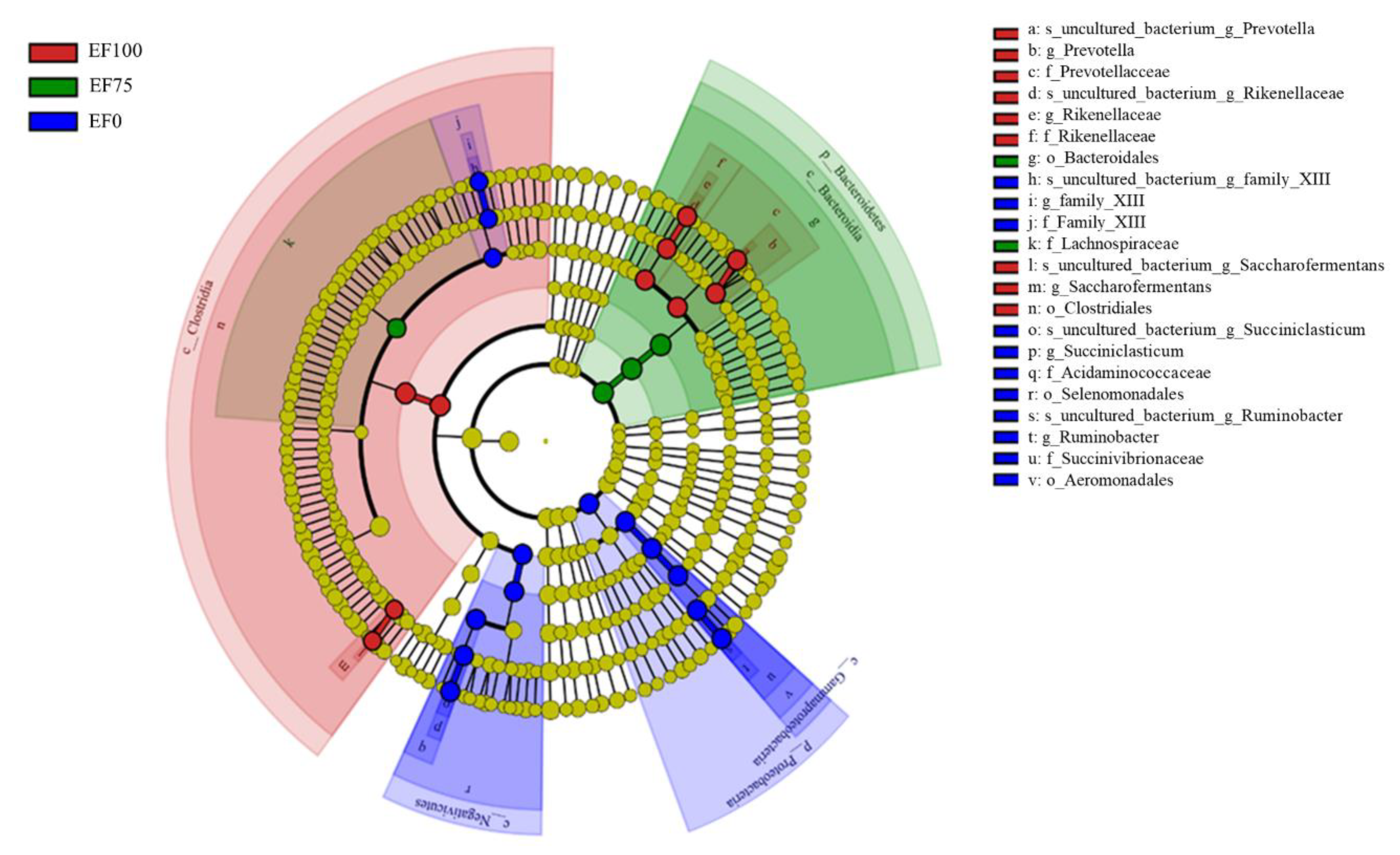
3.3. Composition of the Rumen Microbiota Community in the Sheep
3.4. Rumen Microbiome Metabolic Capacity
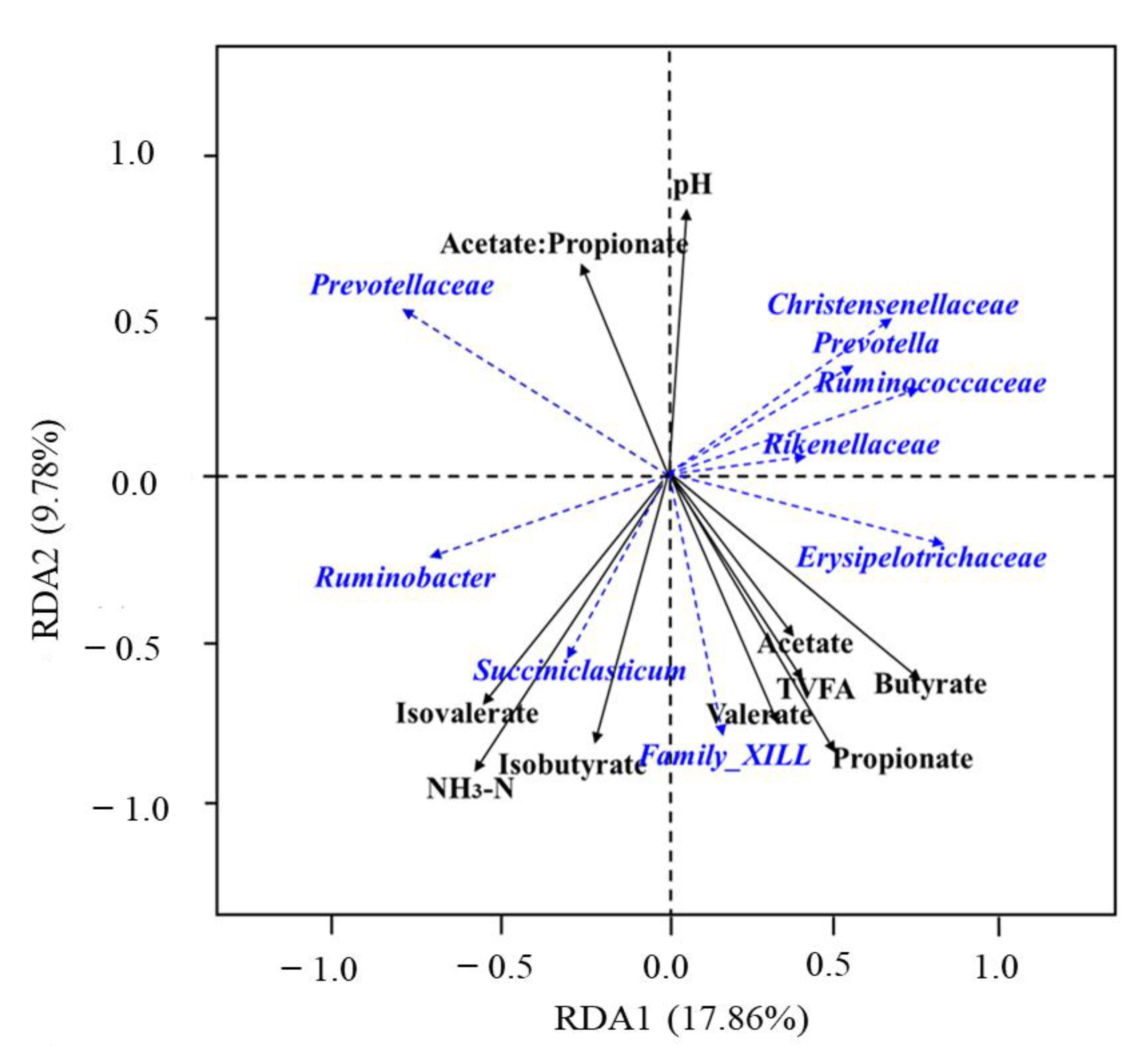
3.5. Relationship among the Sheep Rumen Microbial and Rumen Fermentation Parameters
4. Discussion
4.1. GP Performance
4.2. Ruminal pH, TVFA and NH3-N
4.3. Ruminal Microorganisms
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Shi, Z.C. Important Poisonous Plants in Chinese Grassland, in Study on the Toxicological Mechanism of Phytotoxin, 3rd ed.; China Agriculture Press: Beijing, China, 1997; pp. 166–176. [Google Scholar]
- Li, C.J.; Nan, Z.B.; Zhang, C.J.; Zhang, C.Y.; Zhang, Y.H. Effects of drunken horse grass infected with endophyte on chinese rabbit. J. Agric. Sci. Technol. 2009, 11, 84–90. [Google Scholar]
- Miles, C.O.; Lane, G.A.; diMenna, M.E.; Garthwaite, I.; Piper, E.L.; Ball, O.J.P.; Garrick, C.M.; Latch, G.C.M.; Allen, J.M.; Hunt, M.B.; et al. High levels of ergonovine and lysergic acid amide in toxic Achnatherum Inebrians accompany infection by an Acremonium-like endophytic fungus. J. Agric. Food Chem. 1996, 44, 1285–1290. [Google Scholar] [CrossRef]
- Ren, J.Z. Northwest grasslands several common poisonous herbs. Anim. Hus. Veter. Med. 1954, 2, 56–60. [Google Scholar]
- Wang, K.; Dang, X.P. Toxicity test of drunken horse grass to the sheep. China Veter. Sci. Technol. 1991, 21, 32–33. [Google Scholar]
- Abegunde, T.O.; Akinsoyinu, A.O. Replacement Effects of panicum maximum with ficus polita on performance of west african dwarf goats. J. Anim. Physiol. Anim. Nutr. 2011, 95, 192–197. [Google Scholar] [CrossRef] [PubMed]
- Chatterjee, P.N.; Kamra, D.N.; Agarwal, N.; Patra, A. Influence of supplementation of tropical plant feed additives on in vitro rumen fermentation and methanogenesis. Anim. Prod. Sci. 2014, 54, 1770–1774. [Google Scholar] [CrossRef]
- Khameneh, B.; Iranshahy, M.; Soheili, V.; Fazly Bazzaz, B.S. Review on plant antimicrobials: A mechanistic viewpoint. Antimicrob. Resist. Infect. Control 2019, 8, 118. [Google Scholar] [CrossRef]
- Liang, Y.; Wang, H.C.; Li, C.J.; Nan, Z.B.; Li, F.D. Effects of feeding drunken horse grass infected with epichloë gansuensis endophyte on animal performance, clinical symptoms and physiological parameters in sheep. BMC Veter. Res. 2017, 13, 223. [Google Scholar] [CrossRef]
- Wina, E.; Muetzel, S.; Becker, K. The dynamics of major fibrolytic microbes and enzyme activity in the rumen in response to short- and long-term feeding of Sapindus rarak saponins. J. Appl. Microbiol. 2006, 100, 114–122. [Google Scholar] [CrossRef]
- Patra, A.K.; Saxena, J. The effect and mode of action of saponins on the microbial populations and fermentation in the rumen and ruminant production. Nutr. Res. Rev. 2009, 22, 204–219. [Google Scholar] [CrossRef]
- Morgavi, D.P.; Kelly, W.J.; Janssen, P.H.; Attwood, G.T. Rumen microbial (meta) genomics and its application to ruminant production. Animal. 2012, 7, 184–201. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Wang, H.C.; Guo, X.S. Comparative analysis of rumen fermentation parameters and bacterial profiles during adaption to different fattening stages in beef cattle fed TMR with various forage silage. Anim. Feed Sci. Technol. 2021, 278, 115006. [Google Scholar] [CrossRef]
- Belanche, A.; Doreau, M.; Edwards, J.E.; Moorby, J.M.; Pinloche, E.; Newbold, C.J. Shifts in the rumen microbiota due to the type of carbohydrate and level of protein ingested by dairy cattle are associated with changes in rumen fermentation. J. Nutr. 2012, 142, 1684–1692. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, Y.L.; Wang, H.C.; Li, C.J. Response of sheep rumen fermentation and microbial communities to feed infected with the endophyte Epichloë gansuensis as evaluated with rumen-simulating technology. J. Microbiol. 2021, 59, 719–728. [Google Scholar] [CrossRef]
- McFarlane, Z.D.; Myer, P.P.; Cope, E.R.; Evans, N.D.; Carson Bone, T.; Biss, B.E.; Travis Mulliniks, J. Effect of biochar type and size in vitro on rumen fermentation of Orchard grass hay. Agr. Sci. 2017, 8, 316–325. [Google Scholar]
- Egbert, S.; Karen, S.; Christoph, C.T. Diversity responses of rumen microbial communities to Fusarium-contaminated feed, evaluated with rumen simulating technology. Environ. Microbiol. 2008, 10, 483–496. [Google Scholar]
- AOAC. Official Methods of Analysis, 15th ed.; Association of Official Analytical Chemists: Arlington, VA, USA, 1990. [Google Scholar]
- Goering, H.K.; van Soest, P.J. Forage fibre analysis (Apparatus, Reagent, Procedures and Some Application). In Agricultural Handbook No. 379; Agricultural Research Service, United States Dpartment of Agriculture: Washington, DC, USA, 1970. [Google Scholar]
- Ishii, S.; Kosaka, T.; Hori, K.; Hotta, Y.; Watanabe, K. Coaggregation facilitates interspecies hydrogen transfer between Pelotomaculum thermopropionicum and Methanothermobacter thermautotrophicus. Appl. Environ. Microbiol. 2005, 71, 7838–7845. [Google Scholar] [CrossRef]
- Broderick, G.A.; Kang, J.H. Automated simultaneous determination of ammonia and amino acids in ruminal fluids and in vitro media. J. Dairy Sci. 1980, 63, 65–75. [Google Scholar] [CrossRef]
- Mao, S.; Zhang, R.; Wang, D.; Zhu, W. The diversity of the fecal bacterial community and its relationship with the concentration of volatile fatty acids in the feces during subacute rumen acidosis in dairy cows. BMC Veter. Res. 2012, 8, 237. [Google Scholar] [CrossRef]
- Van, G.M.; Busschaert, P.; Honnay, O.; Lievens, B. Evaluation of six primer pairs targeting the nuclear rRNA operon for characterization of arbuscular mycorrhizal fungal (AMF) communities using 454 pyrosequencing. J. Microbiol. Methods. 2014, 106, 93–100. [Google Scholar]
- Zhang, R.; Zhu, W.; Zhu, W.; Liu, J.; Mao, S. Effect of dietary forage sources on rumen microbiota, rumen fermentation and biogenic amines in dairy cows. J. Sci. Food Agric. 2014, 94, 1886–1895. [Google Scholar] [CrossRef] [PubMed]
- Seddik, H.; Xu, L.; Wang, Y.; Mao, S.Y. A rapid shift to high-grain diet results in dynamic changes in rumen epimural microbiome in sheep. Animal 2019, 13, 1614–1622. [Google Scholar] [CrossRef] [PubMed]
- Øskov, E.R.; McDonald, L. The estimation of protein degradability in the rumen fromincubation measurements weighed according to the rate of passage. J. Agric. Sci. 1979, 92, 499–504. [Google Scholar] [CrossRef]
- Menke, K.H.; Steingawss, H. Estimation of the energetic feed value obtained from chemical analysis and gas production using rumen fluid. Anim. Res. 1988, 28, 7–55. [Google Scholar]
- Zhang, J.K.; Lu, D.X.; Liu, J.X.; Bao, S.N.; Zhou, Q.H.; Liu, Q.H. Research status and development of crude feed quality evaluation index. Prata. Sci. 2004, 21, 55–61. [Google Scholar]
- Chai, J.M.; Alrashedi, S.; Coffey, K.; Burke, J.M.; Feye, K.; Ricke, S.C.; Park, S.H.; Edwards, J.L.; Zhao, A.C. Endophyte-infected tall fescue affects rumen microbiota in grazing ewes at gestation and lactation. Front. Veter. Sci. 2020, 7, 544707. [Google Scholar] [CrossRef]
- Rohweder, D.A.; Barnes, R.F.; Neal, J. Proposed hay grading standards based on laboratory analyses for evaluating quality. J. Anim. Sci. 1978, 47, 747–759. [Google Scholar] [CrossRef]
- Yang, Z.L.; Li, Q.F.; Cao, Y.F. Study on the combined effect of whole plant corn silage and cereal grass by in vitro gas production method. Chinese Anim. Hus. Veter. Med. 2017, 44, 698–707. [Google Scholar]
- Liu, Q.; Wang, C.; Li, H.Q.; Guo, G.; Huo, W.J.; Pei, C.X.; Zhang, S.L.; Wang, H. Effects of dietary protein levels and rumenprotected pantothenate on ruminal fermentation, microbial enzyme activity and bacteria population in Blonde d’Aquitaine × Simmental beef steers. Anim. Feed Sci. Technol. 2017, 232, 31–39. [Google Scholar] [CrossRef]
- Bull, L.S.; Bush, L.J.; Friend, J.D.; Harris, B., Jr.; Jones, E.W. Incidence of ruminal parakeratosis in calves fed different rations and its relation to volatile fatty acid absorption. J. Daily Sci. 1965, 48, 1459–1466. [Google Scholar] [CrossRef]
- Wang, S.; Shi, L.F.; Zhang, Z.B. Effects of the ratio of ramie and ramie on the rumen microbial fermentation parameters in vitro. Chinese J. Anim. Eco. 2018, 39, 39–43. [Google Scholar]
- Trabi, E.B.; Seddik, H.E.; Xie, F.; Lin, L.M.; Mao, S.Y. Comparison of the rumen bacterial community, rumen fermentation and growth performance of fattening lambs fed low grain, pelleted or non-pelleted high grain total mixed ration. Anim. Feed Sci. Technol. 2019, 253, 1–12. [Google Scholar] [CrossRef]
- Huo, W.; Zhu, W.; Mao, S. Impact of subacute ruminal acidosis on the diversity of liquid and solid-associated bacteria in the rumen of goats. World J. Microbiol. Biotechnol. 2014, 30, 669–680. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.S.; Kim, D.H.; Paradhipta, D.H.V.; Lee, H.J.; Yoon, H.; Joo1, Y.H.; Adesogan, A.T.; Kim, S.C. Effects of wormwood (Artemisia montana) essential oils on digestibility, fermentation indices, and microbial diversity in the rumen. Microorganisms 2020, 8, 1605. [Google Scholar] [CrossRef]
- Gharechahi, J.; Vahidi, M.F.; Ding, X.Z.; Han, J.L.; Salekdeh, G.H. Temporal changes in microbial communities attached to forages with different lignocellulosic compositions in cattle rumen. FEMS Microb. Ecol. 2020, 96, fiaa069. [Google Scholar] [CrossRef]
- Lapébie, P.; Lombard, V.; Drula, E.; Terrapon, N.; Henrissat, B. Bacteroidetes use thousands of enzyme combinations to break down glycans. Nat. Commun. 2019, 10, 2043. [Google Scholar] [CrossRef]
- Ransom, J.E.; Jones, D.L.; McCarthy, A.J.; McDonald, J.E. The fibrobacteres: An important phylum of cellulose-degrading bacteria. Microb. Ecol. 2012, 63, 267–281. [Google Scholar] [CrossRef]
- Suen, G.; Weimer, P.J.; Stevenson, D.M.; Aylward, F.O.; Boyum, J.; Deneke, J.; Drinkwater, C.; Ivanova, N.N.; Mikhailova, N.; Chertkov, O. The complete genome sequence of fibrobacter succinogenes S85 reveals a cellulolytic and metabolic specialist. PLoS ONE 2011, 6, e18814. [Google Scholar] [CrossRef]
- McCrackena, B.A.; Garcia, M.N. Phylum synergistetes in the oral cavity: A possible contributor to periodontal disease. Anaerobe 2021, 68, 102250. [Google Scholar] [CrossRef]
- Strobel, H.J. Vitamin B12-dependent propionate production by the ruminal bacterium prevotella ruminicola. Environ. Microb. 1992, 23, 2331–2333. [Google Scholar] [CrossRef]
- Gharechahi, J.; Salekdeh, G.H. A metagenomic analysis of the camel rumen’s microbiome identifies the aajor microbes responsible for lignocellulose degradation and fermentation. Biotechnol. Biofuels 2018, 11, 216. [Google Scholar] [CrossRef] [PubMed]
- Stackebrandt, E.; Hippe, H. Transfer of bacteroidesamylophilus to a new genus ruminobacter gen. nov., nom. rev. as ruminobacter amylophilus comb. nov. Syst. Appl. Microbiol. 1986, 8, 204–207. [Google Scholar] [CrossRef]
- Lamendella, R.; Santo Domingo, J.W.; Ghosh, S.; Martinson, J.; Oerther, D.B. Comparative fecal metagenomics unveils unique functional capacity of the swine gut. BMC Microbiol. 2011, 11, 103. [Google Scholar] [CrossRef] [PubMed]
- Kaoutari, A.E.; Armougom, F.; Gordon, J.I.; Raoult, D.; Henrissat, B. The abundance and variety of carbohydrate-active enzymes in the human get microbiota. Nat. Rev. Microbiol. 2013, 11, 497–504. [Google Scholar] [CrossRef]
- Liu, H.J.; Hu, L.Y.; Han, X.P.; Zhao, N.; Xu, T.W.; Ma, L.; Wang, X.J.; Zhang, X.L.; Kang, S.P.; Zhao, X.Q.; et al. Tibetan sheep adapt to plant phenology in alpine meadows by changing rumen microbial community structure and function. Front. Microbiol. 2020, 10, 587558. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ferment Feed 2 | Item 3 | ||||||
|---|---|---|---|---|---|---|---|
| OM | CP | NDF | ADF | Ash | Ergonovine | Ergine | |
| EF | 906 | 105 | 633 | 317 | 94 | ND 4 | ND |
| PTMR | 875 | 163 | 418 | 215 | 112 | ND | ND |
| Item 1 | Treatment 2 | SEM 3 | p-Value | |||||
|---|---|---|---|---|---|---|---|---|
| EF0 | EF25 | EF50 | EF75 | EF100 | Linear | Quadratic | ||
| DDM (%) | 78.22 a | 63.46 b | 56.79 bc | 55.09 c | 45.70 d | 2.286 | <0.001 | 0.125 |
| GP (mL/g) | 82.24 a | 62.83 b | 55.30 b | 35.57 c | 24.73 c | 4.084 | <0.001 | <0.001 |
| a (mL/g) | 66.42 a | 51.49 b | 50.02 b | 35.17 c | 32.79 d | 3.276 | <0.001 | <0.001 |
| b (mL/g) | 30.06 a | 21.00 b | 13.82 c | 4.23 d | 0.27 e | 2.910 | <0.001 | 0.001 |
| a + b (mL/g) | 96.48 a | 73.49 b | 63.84 c | 37.02 d | 36.44 d | 6.099 | <0.001 | <0.001 |
| c (%h) | 0.03 a | 0.02 b | 0.01 c | 0.03 a | 0.02 b | 0.002 | <0.001 | <0.001 |
| Item 1 | Treatment 2 | SEM 3 | p-Value | |||||
|---|---|---|---|---|---|---|---|---|
| EF0 | EF25 | EF50 | EF75 | EF100 | Linear | Quadratic | ||
| pH | 6.84 d | 6.89 c | 6.90 c | 6.98 b | 7.02 a | 0.013 | <0.001 | 0.331 |
| NH3-N(mg/dL) | 49.33 a | 45.91 ab | 42.91 ab | 39.21 bc | 35.17 c | 1.349 | <0.001 | 0.826 |
| TVFA (mmol/L) | 86.52 a | 73.28 b | 70.74 b | 59.67 bc | 57.28 c | 2.564 | <0.001 | 0.389 |
| Acetate (mmol/L) | 52.21 a | 44.84 ab | 44.18 b | 37.53 bc | 36.44 c | 1.498 | <0.001 | 0.499 |
| Propionate (mmol/L) | 21.17 a | 17.05 b | 15.48 bc | 12.88 c | 11.76 c | 0.765 | <0.001 | 0.299 |
| Butyrate (mmol/L) | 8.40 a | 7.31 ab | 7.24 a | 5.98 b | 5.86 c | 0.267 | <0.001 | 0.670 |
| Valerate (mmol/L) | 1.38 a | 1.17 b | 1.10 bc | 0.91 c | 0.89 cd | 0.044 | <0.001 | 0.304 |
| Isobutyrate (mmol/L) | 1.20 a | 1.07 ab | 0.96 bc | 0.85 c | 0.82 c | 0.036 | <0.001 | 0.384 |
| Isovalerate (mmol/L) | 2.15 a | 1.85 ab | 1.78 ab | 1.52 b | 1.49 b | 0.069 | <0.001 | 0.470 |
| Acetate: Propionate | 2.48 c | 2.65 bc | 2.87 ab | 2.91 ab | 3.14 a | 0.064 | <0.001 | 0.910 |
| Item 1 | Treatment 2 | SEM 3 | p-Value | |||||
|---|---|---|---|---|---|---|---|---|
| EF0 | EF25 | EF50 | EF75 | EF100 | Linear | Quadratic | ||
| ACE | 771.66 | 779.18 | 769.04 | 774.16 | 773.14 | 2.669 | 0.924 | 0.637 |
| Chaol index | 779.37 | 787.71 | 776.22 | 776.32 | 777.74 | 3.365 | 0.592 | 0.777 |
| Shannon index | 4.63 b | 4.78 b | 4.64 b | 5.08 a | 5.07 a | 0.060 | <0.001 | 0.130 |
| KEGG | Relative Abundance (%) 1 | SEM 2 | p-Value | |||||
|---|---|---|---|---|---|---|---|---|
| EF0 | EF25 | EF50 | EF75 | EF100 | Linear | Quadratic | ||
| Carbohydrate metabolism | 14.41 b | 14.46 ab | 14.46 ab | 14.52 a | 14.49 ab | 0.014 | 0.029 | 0.314 |
| Global and overview maps | 14.38 a | 14.36 ab | 14.33 ab | 14.27 b | 14.29 ab | 0.015 | 0.017 | 0.757 |
| Amino acid metabolism | 11.45 | 11.58 | 11.50 | 11.54 | 11.54 | 0.014 | 0.051 | 0.093 |
| Metabolism of cofactors and vitamins | 6.98 | 6.97 | 7.02 | 6.98 | 6.99 | 0.008 | 0.508 | 0.567 |
| Energy metabolism | 6.81 b | 6.86 a | 6.87 a | 6.88 a | 6.88 a | 0.008 | <0.001 | 0.007 |
| Nucleotide metabolism | 5.91 | 5.91 | 5.93 | 5.91 | 5.92 | 0.002 | 0.252 | 0.179 |
| Translation | 5.84 a | 5.84 a | 5.84 a | 5.81 b | 5.82 ab | 0.004 | <0.001 | 0.205 |
| Replication and repair | 5.06 a | 5.00 c | 5.02 b | 4.98 d | 4.99 bd | 0.008 | <0.001 | 0.002 |
| Membrane transport | 4.38 a | 4.08 c | 4.15 b | 3.93 d | 3.93 d | 0.044 | <0.001 | <0.001 |
| Lipid metabolism | 3.38 c | 3.43 bc | 3.41 c | 3.49 a | 3.44 b | 0.010 | 0.001 | 0.090 |
| Glycan biosynthesis and metabolism | 2.76 b | 2.87 a | 2.85 a | 2.83 a | 2.84 a | 0.012 | 0.024 | 0.004 |
| Signal transduction | 2.48 | 2.45 | 2.44 | 2.47 | 2.47 | 0.004 | 0.966 | 0.002 |
| Folding, sorting and degradation | 2.28 a | 2.24 d | 2.27 b | 2.25 cd | 2.25 c | 0.004 | <0.001 | 0.001 |
| Metabolism of other amino acids | 2.17 | 2.19 | 2.19 | 2.18 | 2.19 | 0.003 | 0.150 | 0.173 |
| Metabolism of terpenoids and polyketides | 1.97 c | 2.00 a | 1.99 b | 2.01 a | 2.00 a | 0.003 | <0.001 | 0.002 |
| Xenobiotics biodegradation and metabolism | 1.64 a | 1.65 a | 1.61 b | 1.64 | 1.64 | 0.004 | 0.258 | 0.166 |
| Feed Nutrient and Fermentation Characteristics 1 | Treatment 2 | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| EF0 | EF25 | EF50 | EF75 | EF100 | ||||||
| Shannon | Chao1 | Shannon | Chao1 | Shannon | Chao1 | Shannon | Chao1 | Shannon | Chao1 | |
| CP | −0.982 ** | −0.685 | 0.683 | 0.748 | 0.01 | −0.482 | 0.059 | 0.537 | 0.831 * | 0.209 |
| NDF | −0.537 | 0.178 | 0.797 | 0.821 * | −0.653 | −0.328 | −0.507 | 0.736 | −0.183 | −0.15 |
| ADF | 0.179 | 0.094 | 0.077 | −0.057 | 0.827 * | −0.217 | −0.703 | −0.482 | 0.830 * | −0.006 |
| pH | −0.458 | −0.612 | −0.734 | −0.436 | −0.126 | −0.044 | −0.255 | 0.62 | 0.414 | 0.316 |
| TVFA | 0.559 | 0.303 | −0.349 | 0.195 | 0.526 | 0.634 | 0.59 | −0.031 | −0.071 | 0.435 |
| Acetate | 0.389 | 0.131 | −0.271 | 0.283 | 0.5 | 0.614 | 0.595 | −0.027 | 0.078 | 0.11 |
| Propionate | 0.72 | 0.537 | −0.645 | −0.164 | 0.606 | 0.681 | 0.577 | 0.075 | 0.036 | 0.657 |
| Butyrate | 0.377 | 0.033 | −0.052 | 0.314 | 0.53 | 0.623 | 0.479 | −0.017 | −0.39 | 0.684 |
| Valerate | 0.517 | 0.64 | 0.272 | 0.656 | 0.433 | 0.526 | 0.661 | −0.28 | −0.529 | 0.33 |
| Isobutyrate | 0.231 | 0.361 | 0.204 | 0.629 | −0.278 | −0.088 | 0.486 | −0.586 | −0.338 | −0.105 |
| Isovalerate | −0.078 | 0.321 | 0.338 | 0.601 | −0.373 | −0.183 | 0.508 | −0.657 | −0.163 | −0.211 |
| Acetate: Propionate | −0.468 | −0.492 | 0.798 | 0.584 | −0.544 | −0.465 | 0.21 | −0.47 | 0.037 | −0.453 |
| GP | 0.056 | −0.08 | 0.023 | 0.315 | 0.201 | −0.323 | 0.07 | −0.579 | −0.707 | 0.232 |
| NH3−N | −0.836 * | −0.607 | −0.707 | −0.481 | −0.606 | −0.636 | −0.216 | −0.316 | −0.941 ** | 0.018 |
| DDM | 0.451 | 0.172 | −0.435 | −0.583 | 0.23 | −0.371 | 0.193 | 0.823 * | −0.326 | −0.398 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ma, Y.; Wang, H.; Li, C.; Malik, K. The Potential Use of Endophyte-Free inebrians as Sheep Feed Evaluated with In Vitro Fermentation. Fermentation 2022, 8, 419. https://doi.org/10.3390/fermentation8090419
Ma Y, Wang H, Li C, Malik K. The Potential Use of Endophyte-Free inebrians as Sheep Feed Evaluated with In Vitro Fermentation. Fermentation. 2022; 8(9):419. https://doi.org/10.3390/fermentation8090419
Chicago/Turabian StyleMa, Yaling, Hucheng Wang, Chunjie Li, and Kamran Malik. 2022. "The Potential Use of Endophyte-Free inebrians as Sheep Feed Evaluated with In Vitro Fermentation" Fermentation 8, no. 9: 419. https://doi.org/10.3390/fermentation8090419
APA StyleMa, Y., Wang, H., Li, C., & Malik, K. (2022). The Potential Use of Endophyte-Free inebrians as Sheep Feed Evaluated with In Vitro Fermentation. Fermentation, 8(9), 419. https://doi.org/10.3390/fermentation8090419