1. Introduction
Forage is an indispensable component in feeds and serves a nutritional and behavioral function for the maintenance and production of ruminants. One important function of dietary forage is to promote chewing activity to stimulate saliva secretion, which is critical in improving rumen buffering capacity and maintaining rumen homeostasis [
1]. The decomposition of plant forage into volatile fatty acids (VFA) by ruminal cellulolytic bacteria generates large amounts of hydrogen (H
2) [
2], which is used by methanogenic archaea as an energy source, resulting in methane (CH
4) production. Ruminant-derived CH
4 emissions account for around one quarter of all anthropogenic CH
4 emissions and are implicated as a driver of global climate change [
3,
4]. Feeding with a high proportion of dietary forage always causes an increase in enteric CH
4 emissions in ruminants. Due to the current ruminant production and climate issues, contemporary work exploring natural plant forage is needed to alter feeding behavior and reduce enteric CH
4 emissions [
5].
Ramie (
Boehmeria nivea), a hardy perennial herbaceous plant with high quality of bast fiber, has been traditionally grown as a plant textile raw material [
6]. Ramie also has high crude protein content (179–211 g/kg, dry matter (DM) basis), and can be cut annually with high biological yields (90.1–94.6 t/hm
2 a) [
7,
8]. Replacing less than 67% of alfalfa hay with ramie silage has been reported to have no adverse effects on milk production and blood biochemical parameters in dairy cows [
8]. Replacing less than 35% of alfalfa hay with ramie had no negative effects on growth performance and improved the meat quality in goats [
9]. However, no studies have been conducted to investigate the effect of ramie supplementation on chewing activity and enteric CH
4 emissions, although ramie is rich in bast fiber and plant secondary metabolites such as tannins, polyphenol, and flavonoids [
10,
11].
We hypothesized that ramie silage had distinct plant fiber and rumen fermentation characteristics compared to corn stover (CS) silage, which might affect the feed behavior and methanogenesis in ruminants. Goats were employed as experimental animals, while the study was designed to investigate the nutritional value of ramie silage, and its consequences for chewing activity, rumen fermentation, and enteric CH4 emissions, by comparing it with CS silage. The first experiment aimed to compare fiber degradation, gas production, and rumen fermentation between ramie and CS silage through in vitro batch culture. Then, an in vivo study was conducted to evaluate the effects of replacing 60% of CS silage with ramie silage on fiber digestibility, chewing activity, rumen fermentation, CH4 emissions, and microbiota in goats.
2. Materials and Methods
All procedures involving animals were approved (No. ISA-W-202001) by the Animal Care Committee, Institute of Subtropical Agriculture, the Chinese Academy of Sciences, Changsha, China.
2.1. Preparation of Silages
Ramie silage (
B. Nivea “Zhongzhu No.1”) and CS silage were obtained from Hunan Deren Animal Husbandry Technology Co., Ltd., Changsha, China The fresh ramie was collected at 1.2 m height, while corn stover after fresh corn harvesting was collected with the stem cut at around 15 cm above the ground level. Both forages were chopped to 3–4 cm length with a straw cutter (RC500 Chaffcutter, Qufu Ruicheng Agricultural Machinery Co., Ltd., Qufu, China), and then packed by an automatic silage wrapping machine (model TSW2020C, Shanghai Shidaer Modern Agricultural Machinery Co., Ltd., Shanghai, China) to prepare the silage (>60 d). When the experiment started, three fresh samples of around 300 g were collected after opening per silage for in vitro incubation and for detection of DM content and chemical composition (
Table 1).
2.2. In Vitro Ruminal Batch Incubation
As rumen fluid donor animals, 3 healthy adult male fistulated Xiangdong black goats with comparable body weight (average BW 25 ± 2.0 kg) were chosen. The goats were fed a mixed diet of rice straw and concentrate (1:1) containing 137 and 380 g/kg DM of crude protein and neutral detergent fiber, respectively. Rumen contents were randomly collected from 2 fistula sheep before morning feeding (08:00 h), filtered through 4 layers of sterile gauze into preheated thermos flasks, and mixed artificial rumen cultures were created by mixing rumen fluid with artificial saliva [
12] at a ratio of 1:4 (
v/
v). Approximately 1.0 g of the substrate was weighed into a 135 mL fermentation bottle and incubated with 60 mL of buffered rumen fluid under a stream of carbon dioxide (CO
2) at 39.5 °C. Bottles were immediately placed into the fully automatic in vitro batch culture system, as described by Wang et al. [
13], with venting pressure set at 10.0 kPa. In vitro rumen fermentation was stopped at 48 h. The gas production was calculated using the method described by Wang et al. [
14].
Approximately 2 mL of subsample was centrifuged at 12,000×
g for 10 min at 4 °C, and then the supernatants (1.5 mL) were transferred into tubes, acidified with 0.15 mL of 25% (
w/
v) metaphosphoric acid, and stored at −20 °C overnight for analysis of volatile fatty acids. The remaining solid residues were separated to dry at 105 °C to constant weight. The degradation of the incubated substrates was measured based on the difference in DM and NDF weights (dried at 105 °C) before and after incubation [
15].
Three runs of incubation were conducted. Each run contained six culture bottles as a technical repetition per each treatment, conducted with rumen fluid from 1 of 3 donor goats, and repeated 3 times on different days and donor goats so that each treatment had three biological replicates.
2.3. Animals and Experimental Design
Eight female Xiangdong black goats (a local breed in Southern China, 1 to 1.2 years of age) with BW of 21.0 ± 1.05 kg (mean ± standard deviation) were used for this experiment and were randomly assigned to either one of the two dietary treatments in a cross-over design. The diet was formulated to meet appropriately 1.2 to 1.3 times the energy and protein requirements of growing goats by referring to Zhang and Zhang [
16] (
Table 2). The ramie diet was formulated by replacing 60% of CS silage and urea with ramie silage (DM basis) to obtain similar CP content between CS and ramie silage diets (
Table 2). All goats were kept in individual pens with free access to fresh water and fed individually equal meals at 08:30 and 17:30 h.
Each experimental period lasted 41 d, with the first 28 d for diet adaptation and the last 13 d for sampling and data collection. During the initial 10 d of adaptation to diets, the feed was offered ad libitum, targeting 5% refusals. The amount of feed allocated daily during the next 18 d of adaptation was adjusted to 100% of the DM intake previously measured to minimize feed selection. The refusals, when present, were collected and analyzed to determine the actual nutrient intakes. The 15 d sampling period included 4 d for measurements of chewing activity, 5 d for total feces collection, 2 d for rumen fluid sampling, and 4 d for measurements of CH4 and CO2 emissions (2 d for each goat in respiratory chambers).
2.4. Chewing Activity
A high-definition infrared camera (Haier, WSC-580W) was installed above each goat’s cage to continuously monitor the chewing activity from 27 to 30 d. At the same time, the daily feed intake and leftover feed were recorded, and the dry matter intake was calculated. After the test, the eating activity, ruminating activity, non-nutritive oral activity, and idle activity of goats were observed and recorded on a computer. The above behavior definition, observation methods, and recording standards referred to Muhammed et al. [
17].
2.5. Nutrient Digestibility
Total-tract nutrient digestibility was measured daily from d 31 to 35. Total feces of each goat were collected, weighed, and mixed daily. Two subsamples (~10% each) were collected for nutrient analysis. A fecal subsample (~1%) was frozen immediately at −20 °C, and another fecal subsample (~1%) was acidified using 10% (
w/
w) H
2SO
4 to prevent N loss and then frozen immediately at −20 °C. The samples of the diets, feces, and orts were dried at 65 °C for 48 h in a forced-air oven, ground through a 1-mm screen, and then the acidified oven-dried samples were used for total N analysis, whereas non-acidified oven-dried samples were used for the rest of the chemical analyses. All samples were processed for chemical analysis according to the procedures of Wang et al. [
18].
2.6. Rumen Sampling
Rumen sampling was performed over a consecutive 2 d period from d 36 to 37 after measuring total-tract nutrient digestibility. Rumen contents were collected by oral stomach tubing (Anscitech Co., Ltd., Wuhan, China) at 0, 2.5, and 6 h after the morning feeding, according to the method described by Wang et al. [
18]. In order to avoid saliva contamination, the first 100 mL of rumen content was discarded, and the second approximately 150 mL of rumen content was rapidly collected for subsequent measurements.
Two subsamples (15 mL each) were immediately frozen at −80 °C in liquid nitrogen for DNA extraction and subsequent quantification of microbial groups. Two other subsamples (35 mL each) were immediately transferred into 50-mL plastic syringes for measuring dissolved hydrogen (dH2) and dissolved methane (dCH4) concentrations. Approximately 20 mL of rumen content was used for the immediate measurement of ruminal pH using a portable pH meter (Starter 300; Ohaus Instruments Co., Ltd., Shanghai, China). An aliquot of 2.5 mL of rumen content was centrifuged at 15,000× g for 10 min at 4 °C, and aliquots of the supernatants (1.5 mL) were transferred into tubes containing 0.15 mL of 25% (w/v) metaphosphoric acid, vigorously hand-shaken and stored at −20 °C for subsequent determination of VFA and ammonia concentration.
2.7. Measurement of Enteric CH4 and CO2 Emissions
Enteric CH
4 and CO
2 emissions were measured over the 48-h period after the collection of rumen contents, according to Wang et al. [
19]. As only 4 respiration chambers were available, it took a total of 4 d to measure the CH
4 and CO
2 emissions of the 8 goats. The net volume of each respiration chamber was around 2.0 m
3 (i.e., 1.7 m length × 0.9 m width × 1.3 m height). The goats were restricted but free to eat feed and drink water. The plexiglass structure of the five respiration chambers permitted goats to see each other, minimizing stress. The airflow was controlled by the air pump and maintained at negative pressure (flow rate = 40 m
3/h) to prevent the losses of CH
4 and CO
2 produced by goats. Gas discharged from the respiration chamber and ambient gas were connected to a multiport inlet unit of the gas analyzer (GGA-30P, Los Gatos Institute, Los Gatos, CA, USA) to measure CH
4 and CO
2 concentrations. To minimize the chamber effect, correction factors accounting for respiration chamber differences were estimated by using the methodology described by McGinn et al. [
20]. The respiration chambers were opened twice a day at 07:30 and 17:30 to deliver the diets and cleaned before morning feeding.
2.8. Sample Analyses
2.8.1. Chemical Composition
All samples of feeds, feed refusals, and feces were dried at 65 °C for 48 h and ground through a 1-mm screen for chemical analysis. The DM (method 945.15), OM (method 942.05), CP (method 954.01), and EE (method 920.39) were determined according to AOAC methodologies [
21]. The GE content was measured using an isothermal automatic calorimeter (5EAC8018; Changsha Kaiyuan Instruments Co., Ltd., Changsha, China). Neutral detergent fiber (NDF) and acid detergent fiber (ADF) were assayed and expressed inclusive of residual ash using the methods of Van Soest et al. [
22], and NDF was assayed with the addition of a heat-stable amylase and sodium sulfite. Hemicellulose was calculated according to NDF and ADF content. Starch was analyzed following pre-extraction with ethanol (80%), according to the procedure of Kartchner and Theurer [
23]. Dissolved CH
4 and dH
2 in rumen fluid were measured using the procedures described by Wang et al. [
24].
2.8.2. Fermentation End Products
Frozen rumen samples were thawed and centrifuged at 12,000×
g for 10 min at 4 °C, and the supernatants were used for subsequent analysis of VFA and ammonia concentration. Individual VFA concentrations were analyzed by gas chromatography (GC, Agilent 7890A, and Agilent Inc., PaloAlto, CA, USA) according to Wang et al. [
25], assuming equal fractional rates of individual VFA absorption.
2.8.3. DNA Extraction and Microbial Analysis
The rumen contents obtained at 0, 2.5, and 6 h were mixed and freeze-dried for microbial analysis. The microbial DNA was extracted by using a modified RBB + C methodology [
26] with sand beating, according to Ma et al. [
27]. The quantity of the DNA extracts was assessed based on absorbance at 260 and 280 nm using a ND-2000 spectrophotometer (NanoDrop Technologies, Wilmington, DE, USA). Total bacteria, protozoa, fungi, and methanogens, and selected groups of bacteria (
Ruminococcus albus,
Ruminococcus flavefaciens,
Fibrobacter succinogenes,
Selenomonas ruminantium,
Prevotella ruminicola,
Prevotella,
Ruminobacter amylophilus, and
Butyrivibrio fibrisolven), were quantified by qPCR using primers listed in
Table S1. Quantitative PCR was performed according to Jiao et al. [
28]. Final absolute amounts of the target group or species were estimated by relating the cycle threshold (C
T) value to the standard curves and expressed as log10 copies/g DM rumen content. The abundance of these microorganisms was calculated as copy number per g DM rumen content and was further converted to log10 for further statistical analysis.
The DNA sequencing of rumen fluid was performed according to our previous study [
29]. Briefly, the primer pair of 341F: 5′-CCTAYGGGRBGCASCAG-3′ and 806R: 5′-GGACTACNNGGGTATCTAAT-3′ was used to amplify the V3–V4 region [
30]. All amplicon libraries’ preparation and sequencing were performed on the MiSeq platform (Illumina, San Diego, CA, USA) at Shanghai Biozeron Biological Technology Co., Ltd, Shanghai, China. A total of 611,382 high-quality reads were generated, with an average of 38,211 ± 574 reads per sample, for the assignment of amplicon sequence variants (ASVs) by DADA2 [
31]. Taxonomy was annotated using Silva (release 138.1,
http://www.arb-silva.de (accessed on 4 August 2022)). Alpha diversity of samples, mainly including Chao1, Shannon, Evenness, and Coverage indices, was evaluated using MOTHUR v.1.39. [
32]. The principal coordinate analysis (PCoA) of the ruminal bacterial community was performed based on the unweighted UniFrac dissimilarity matrix at the taxon (ASV) level. For linear discriminant analysis (LDA), the statistical differences between different treatments with LDA scores greater than 3 were performed.
2.9. Statistical Analysis
Data were subjected to analysis using the SPSS 22.0 software (SPSS Inc., Chicago, IL, USA). The in vitro data were analyzed using the procedure of the generalized linear model (GLM), which included types of silage (n = 2) as fixed effects, and incubation run (j = 3) as random effects. The in vivo data were analyzed using the linear mixed model, which included treatment as a fixed effect (n = 2), and goat (n = 4) and period (n = 2) as random effects. When sampling time was included, the model included treatment (n = 2) and the interaction between treatment and sampling time as a fixed effect, sampling time (n = 3) as a repeated measurement, and goat (n = 4) and period (n = 2) as random effects. The covariance structure was heterogeneous autoregressive (ARH1), which had low Akaike’s information criterion values to optimize the statistical analysis. Relative abundances of bacteria were analyzed using the Wilcoxon rank sum tests in the JMP Pro software (JMP Pro version 13.2.1, SAS Institute Inc. SAS Institute, Cary, NC, USA). Differences at p < 0.05 were considered significant.
3. Results
The CS silage had higher in vitro digestibility of OM (
p < 0.01) and NDF (
p < 0.01), total gas (
p < 0.01) and CH
4 volume (
p < 0.01), and lower H
2 volume (
p < 0.01), in comparison with ramie silage (
Table 3). The ramie silage had greater (
p < 0.01) molar percentages of acetate, isobutyrate, valerate, and isovalerate, and a lower (
p < 0.01) total VFA concentration and molar percentage of propionate than CS silage (
Table 3). The fermentation kinetics during the incubation is represented in
Figure S1.
Goats fed the ramie silage diet had lower OM (−7.66%;
p = 0.02) and GE (−7.20%;
p = 0.03) intake and total-tract digestibility of DM (−10.1%;
p < 0.01), OM (−7.72%;
p < 0.01), CP (−12.2%;
p < 0.01), NDF (−12.4%;
p < 0.01), starch (−1.6%;
p = < 0.01), and GE (−4.1%;
p < 0.01) than those fed the CS silage diet (
Table 4). Feeding with the ramie silage diet decreased enteric CH
4 emissions expressed as g/day (−24.1%;
p < 0.01), g/kg DM intake (−18.7%;
p = 0.01), and g/kg OM digested (−18.5%;
p = 0.01) (
Table 5), and enteric CO
2 emissions expressed as g/day (−17.3%;
p < 0.01) and g/kg DM intake (−11.3%;
p = 0.03) (
Table 4).
Goats fed the ramie silage diet had greater (
p < 0.05) rumination activity expressed as min/day, number/day, and s/g DM intake, greater chewing activity expressed as min/day and s/g DM intake, and lower (
p < 0.05) eating activity expressed as min/day and s/g DM intake and idle activity as min/day, in comparison with those fed the CS silage diet (
Table 5).
Goats fed the ramie silage diet had lower dCH
4 (−22.5%,
p = 0.01), in comparison with those fed the CS diet (
Table 6). Feeding with the ramie silage diet caused a greater total VFA concentration (+24.8%,
p < 0.01), together with a greater molar percentage of acetate (
p < 0.01) and isobutyrate (
p < 0.01) and acetate to propionate ratio (
p < 0.01), and lower molar percentage of propionate (
p < 0.01) and isovalerate (
p < 0.01) (
Table 6). The time of sampling had a significant effect (
p < 0.01) on rumen parameters, except dCH
4 and molar percentage of butyrate (
p > 0.05). Significant diet × time interactions (
p < 0.01) were observed for the molar percentage of acetate and propionate. Goats fed the ramie silage diet had a greater (
p < 0.05) molar percentage of acetate at 2.5 and 6 h after feeding and a lower (
p < 0.05) molar percentage of propionate at 2.5 and 6 h after feeding, in comparison with those fed with the CS diet (
Figure S2).
Goats fed the ramie silage diet had a greater (
p = 0.01) 16S rRNA gene copy number of
Ruminococcus flavefaciens than those fed the CS silage diet (
Table 7). Although feeding with ramie silage did not alter (
p > 0.05) alpha diversity (
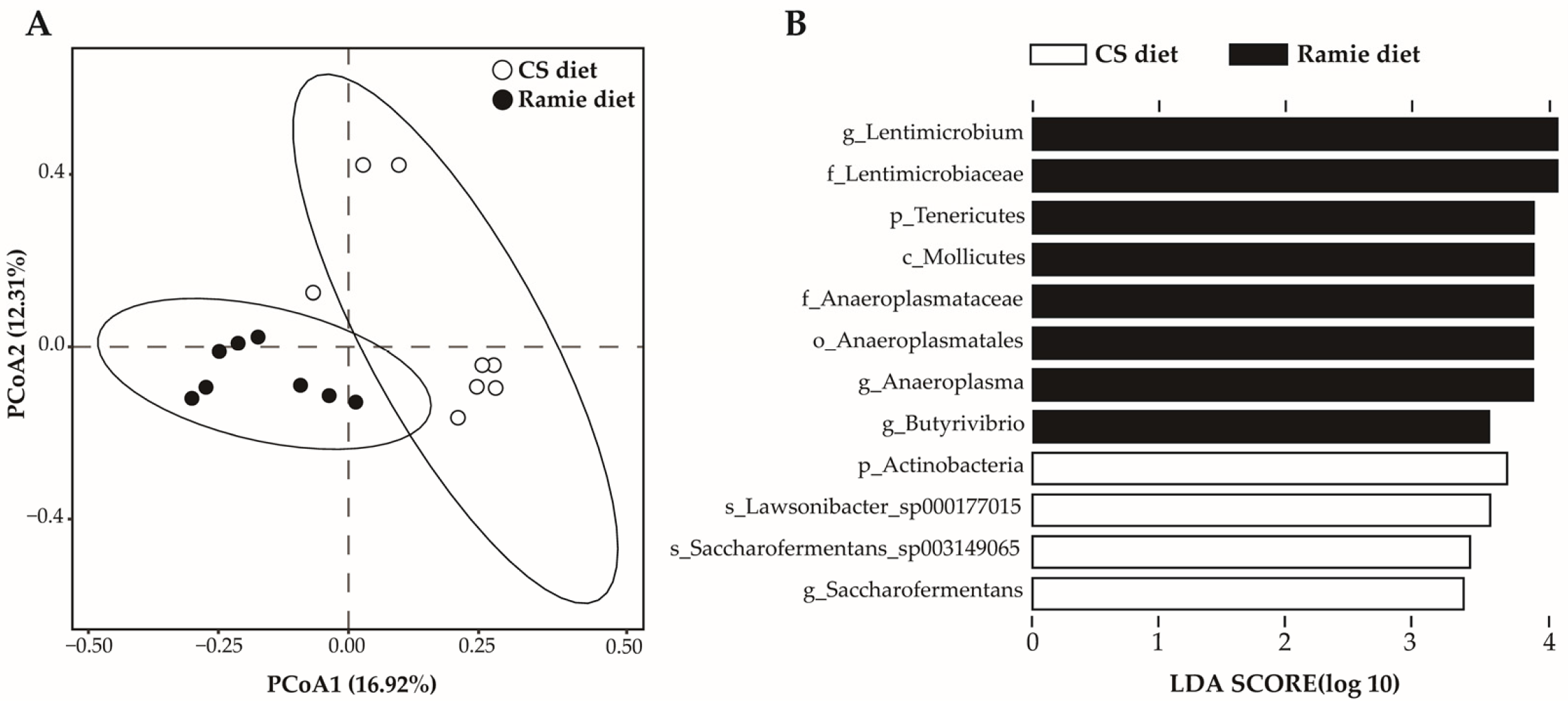
Table 8), it led to a distinct composition of the bacterial community (R
2 = 0.11,
p < 0.01) in those fed the CS silage diet based on unweighted UniFrac dissimilarity (
Figure 1). At the phylum level, goats fed the ramie silage diet had a lower abundance of genera
Firmicutes,
Actinobacteria,
Candidatus Saccharibacteria, and
Tenericutes and higher abundance of genera
Tenericutes than those fed the CS silage diet (
p < 0.05) (
Table 8). At the genus level, feeding with the ramie silage diet led to a greater abundance of genera
Lawsonibacter,
Sedimentibacter,
Saccharofermentans,
Sediminibacterium, and
Bifidobacterium and lower abundance of genus
Anaeroplasma (
p < 0.05) (
Table 8). The line difference discriminant analysis (LDA) indicated that the ramie silage treatment enriched phylum
Tenericutes and genera
Lentimicrobium,
Anaeroplasma, and
Butyrivibrio (LDA > 3). The CS silage treatment enriched phylum
Actinobacteria and genus
Saccharofermentans (LDA > 3).
4. Discussion
The main characteristic of ramie silage is its higher CP (169 g/kg) and ash content and lower hemicellulose content when compared with CS silage. Such results were consistent with previous studies, which reported that the content of CP and ash in ramie silage is 161–174 g/kg and 141–155 g/kg, respectively [
33,
34]. Furthermore, ramie silage exhibited lower in vitro OM and NDF degradation and greater molar percentages of acetate, indicating that ramie was more difficult to be degraded by the ruminal microbiota and facilitated acetate production, which agrees with our previous study [
35]. However, the ramie silage led to lower CH
4 production with greater H
2 accumulation, which indicated that ramie might have a function of inhibiting ruminal methanogenesis in ruminants.
Replacing 60% of CS silage with ramie silage did not alter the feed intake in goats, although feeding with the ramie silage diet led to lower OM intake. Such a reduction in OM intake could be caused by the higher ash content in the ramie silage diet. Wu et al. [
8] reported that using ramie to replace other forages, such as alfalfa, did not significantly alter the feed intake in dairy cows. However, feeding with ramie silage may cause a reduction in the nutrient digestibility due to its high ash content (141–155 g/kg) and distinct fiber structure [
34,
36]. Tang et al. [
37] reported that feeding a diet with greater than 25% of ramie decreases nutrient digestibility in goats. In our study, feeding a diet with 44% of ramie silage led to lower apparent OM, CP, and NDF digestibility than those fed the CS silage diet, indicating lower nutrient utilization in goats. Although feeding a diet with a higher proportion of ramie might not alter feed intake, it could cause the inhibition of nutrient digestibility.
Chewing activity has an important biological function in breaking down the particle size of ingested feed and promoting saliva secretion, which is critical to maintaining rumen health and growth performance in ruminants [
38]. It has been widely reported that ramie has a very distinct fiber structure compared to other forages [
11,
36]. There are many colloids between the ramie fibers, which are attached to the fiber bundle [
39], while fiber bundles on the surface of the ramie are unevenly distributed and have a grid structure [
40]. Such a unique structure of ramie fiber can alter the chewing activity in ruminant animals. In our study, goats fed the ramie silage diet increased the time spent on rumination activity and decreased the time spent on eating activity and idling activity. These results indicated that goats fed a ramie silage diet need more time to break down the intake of feed into smaller particles. However, promoting chewing activity can also have some benefits for maintaining rumen function in ruminant animals, as enhancing oral saliva secretion stimulated by chewing activity can regulate the rumen pH [
41]. Unfortunately, we did not find a difference in rumen pH between the two treatments in the current study, which could be due to the high forage content in the diets. Such results were in agreement with the literature, which reported that feeding with forage diets leads to an optimum ruminal pH with a range of 6.5 to 7.0 [
42,
43].
Volatile fatty acids are mainly produced during carbohydrate fermentation by the rumen microbiota and can be absorbed via the rumen wall to meet up to 70% of a ruminant’s energy requirements [
43,
44]. Ruminal VFA concentration can be influenced by many factors, which include nutrient digestibility, saliva secretion, rumen epithelial absorption, and emptying rate [
45]. Feeding the ramie silage diet increased the VFA concentration in vivo, although ramie silage has lower degradability and VFA concentration through in vitro ruminal incubation. Such inconsistent results are also observed in other studies and may be caused by the decreased rumen epithelial absorption [
29,
46]. A high-forage diet rich in cellulose, hemicellulose, and lignin favors acetate and butyrate production, whereas a high-grain diet rich in starch favors propionate production [
47,
48]. Soluble sugars tend to stimulate greater proportions of butyrate. Although ramie silage had lower hemicellulose and higher pectin content than CS silage, replacing CS silage with ramie silage resulted in a higher molar percentage of acetate and lower molar percentage of propionate in the rumen of goats. A previous study also indicates that increasing the proportion of dietary ramie linearly increases the acetate molar percentage and decreases the propionate molar percentage in vitro and in vivo [
49]. It seems that ramie has a distinct rumen fermentation profile when compared with other forages [
50,
51]. Such facilitation of acetate production could be partially caused by the slightly increased pectin content and is not consistent with the lower hemicellulose content in the ramie silage diet, which needs further investigation.
Hydrogen is produced during the fermentation of carbohydrates to VFA and is mainly consumed by methanogenic archaea for methanogenesis in the rumen [
52]. The formation of acetate and butyrate causes net H
2 production, while the formation of propionate on the other hand acts as net H
2 utilization and is accompanied by a reduction in methanogenesis [
20]. In our study, feeding with the ramie silage diet decreased the ruminal dCH
4 concentration and enteric CH
4 emissions, together with an increased acetate molar percentage and acetate to propionate ratio. Such results were inconsistent with the widely accepted positive relationship between CH
4 and acetate produced in the rumen [
53,
54]. Many studies report that reductive acetogenic bacteria widely exist in the rumen microbial ecosystem and may compete with methanogens for H
2, producing acetate as the end-product through “2CO
2 + 4H
2 → CH
3COOH + 2H
2O” [
55,
56]. We speculated that the increased acetate molar percentage in the ramie silage treatment might be caused by the promotion of the CO
2 assimilation pathway for acetate production, leading to a reduction in methanogenesis. Ramie can also increase tannins [
10,
57], which show a function in CH
4 inhibition [
5,
58]. However, this was not the case in our study, as tannin content was not enriched in our ramie silage.
The rumen microbial ecosystem comprises a diverse symbiotic population of anaerobic bacteria, ciliated protozoa, fungi, and archaeal methanogens [
59]. Rumen fungi and protozoa are thought to play a role in the initial degradation of large fragments, which is helpful for the subsequent anaerobic bacterial colonization and decomposition into plant cell-wall polysaccharides [
48,
60]. Although feeding the ramie silage diet did not alter the population of bacteria, protozoa, and fungi, it increased the 16S rRNA gene copies of
R. flavefaciens. It seems that feeding the ramie silage diet favored the colonization and growth of
R. flavefaciens, which could be primarily involved in the degradation of plant cell walls in the rumen [
61]. However, such an increase in the
R. flavefaciens population did not lead to an increase in fiber digestibility in goats fed the ramie silage diet. Furthermore, feeding the ramie silage diet did not alter 16S rRNA gene copies of methanogens, which is not consistent with the reduction in ruminal dCH
4 and enteric CH
4 emissions. Many published studies also report that the inhibition of CH
4 production may not be always associated with a reduction in the abundance of the methanogen population [
50,
62]. Such a reduction in CH
4 production might be caused by the decreased activities of methanogens.
The 16SrRNA gene sequencing was further employed to investigate changes in bacterial community composition. Although replacing 60% of CS silage with ramie silage did not alter the alpha diversity indexes, the PCoA results indicated that the dietary treatment drove marked changes in the microbial community composition, which was consistent with the results reported by Sun et al. [
63]. Feeding with the ramie silage diet decreased the abundance of the predominant phyla
Firmicutes and
Bacteroidetes, although it increased the relative abundance of phylum
Tenericutes. Among these, genera
Lawsonibacter,
Sedimentibacter,
Saccharofermentans,
Sediminibacterium, and
Bifidobacterium, having the function of degrading various soluble and insoluble polysaccharides, such as starch, pectin, and cellulose in the rumen [
64,
65,
66,
67,
68], were greatly decreased under the ramie silage treatment. Such results were consistent with the decreased fiber digestibility in goats.


 ,
, 

{kind=link}