
The Sensorial and Chemical Changes in Beer Brewed with Yeast Genetically Modified to Release Polyfunctional Thiols from Malt and Hops


Abstract
:1. Introduction
2. Materials and Methodology
2.1. Yeast
2.2. Yeast Propagation
2.3. Brewing
2.4. Hops
2.5. Fermentation and Finished Beer
2.6. Chemistry
2.7. Sensory Evaluation
2.8. Statistical Analysis
3. Results and Discussion
3.1. Beer Chemistry and Fermentation Performance
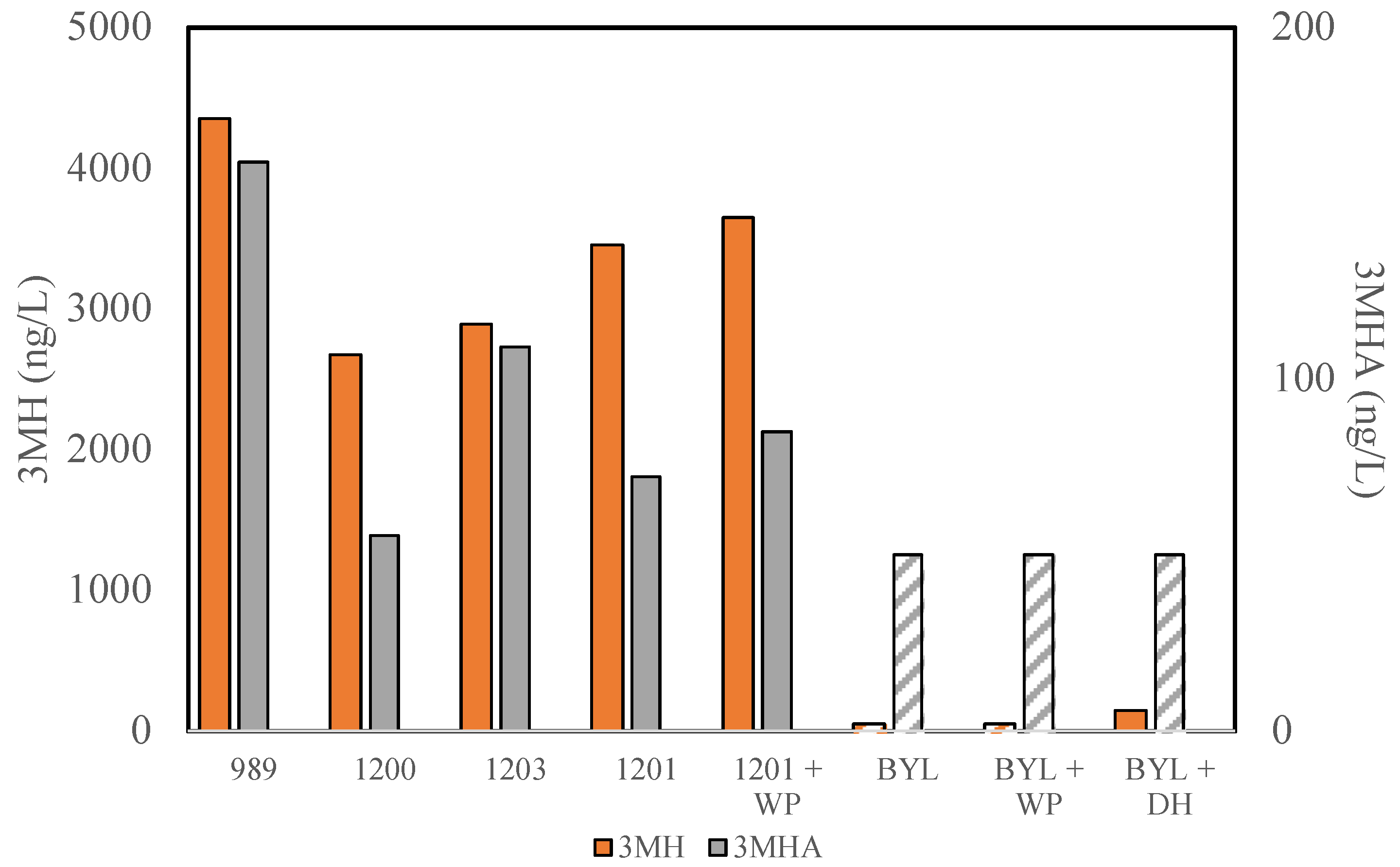
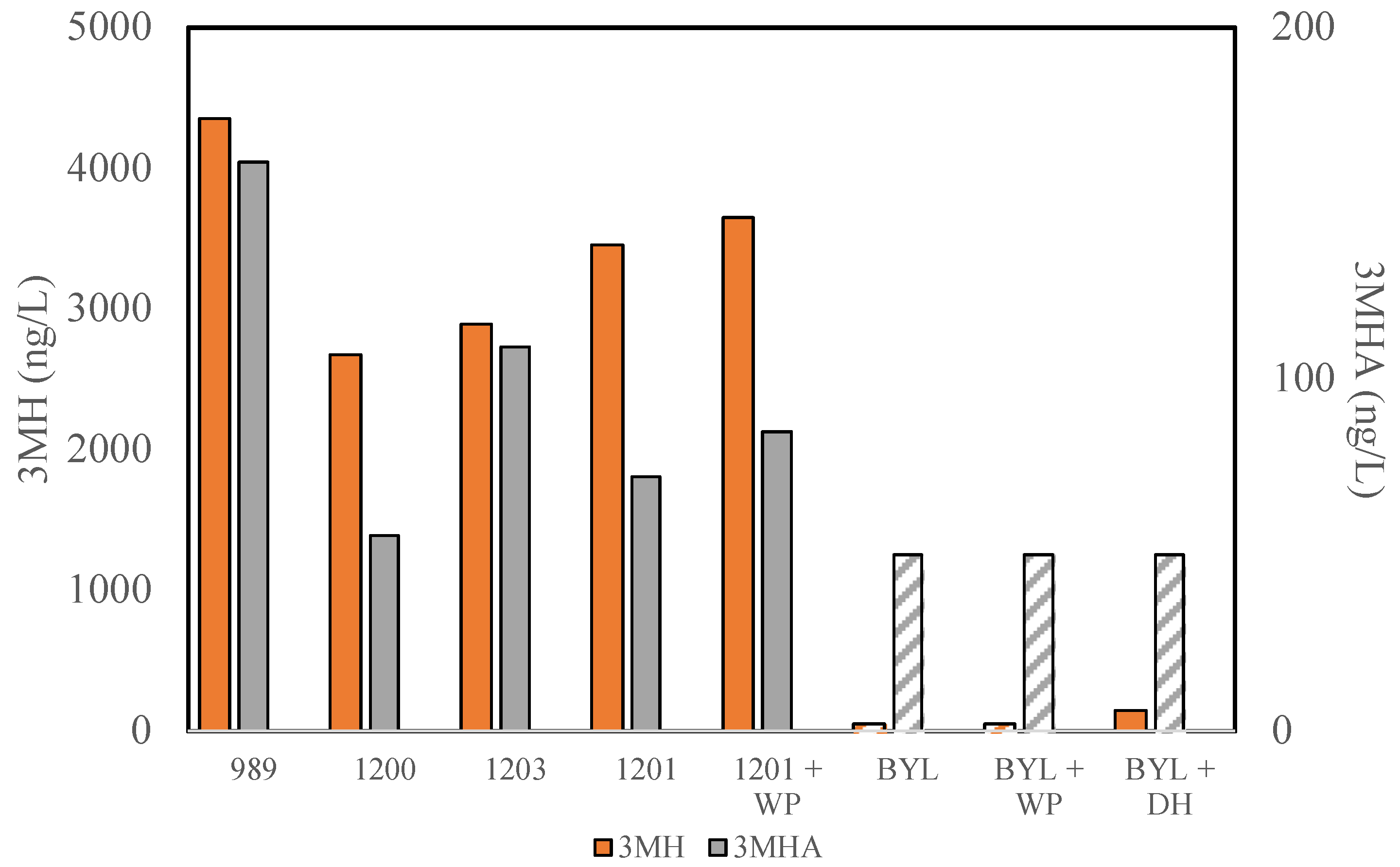
3.2. Thiol Production
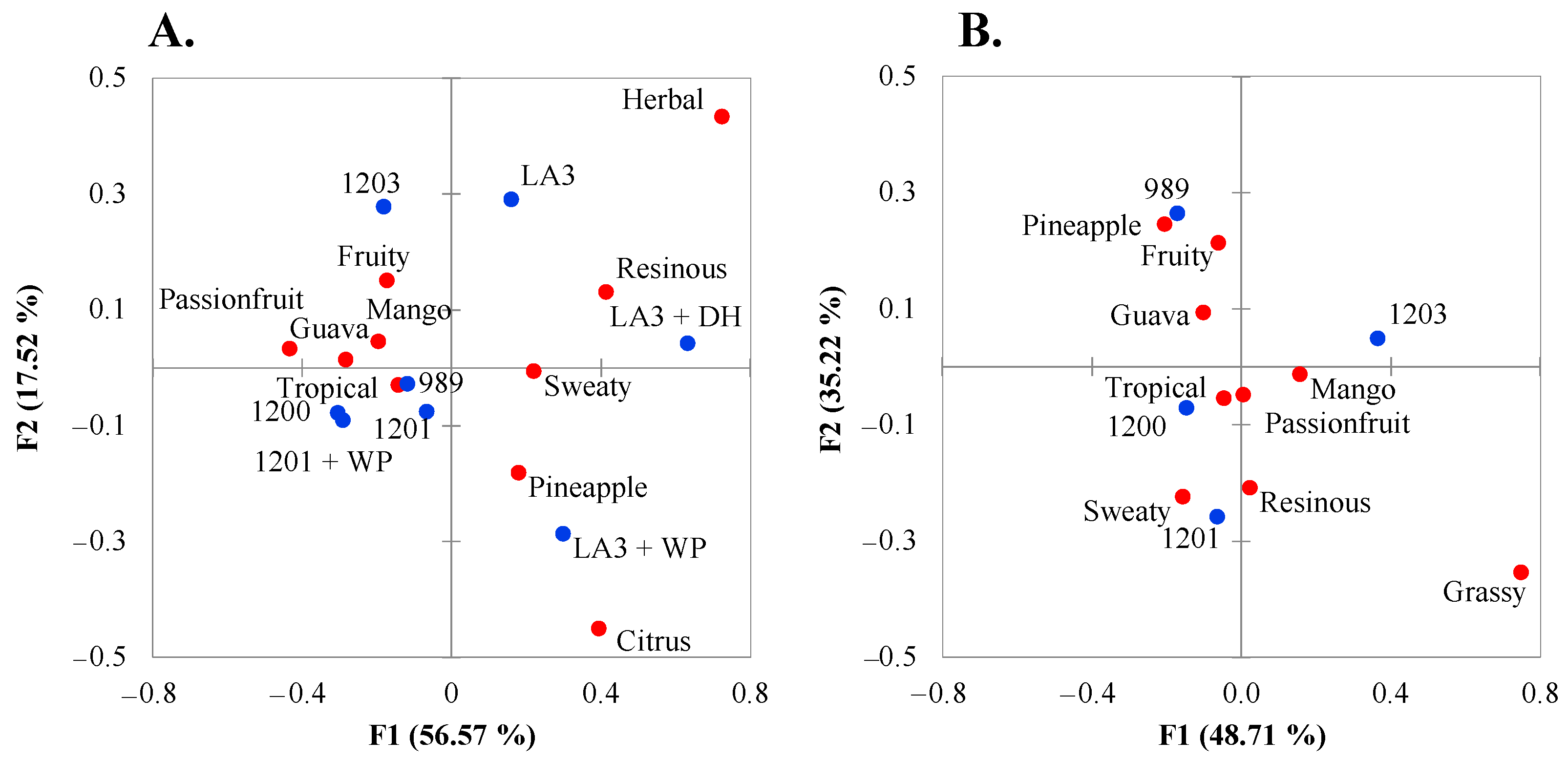
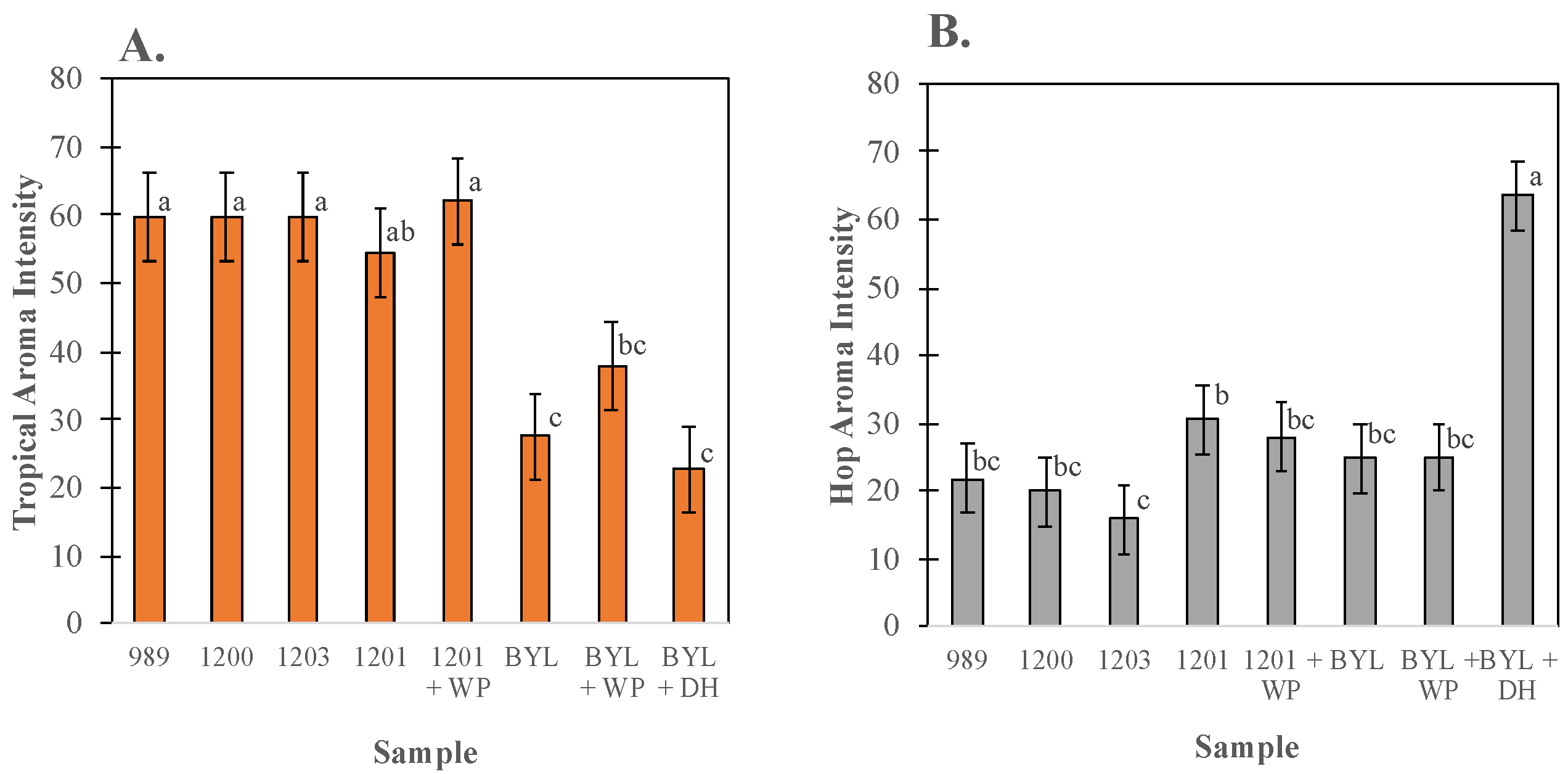
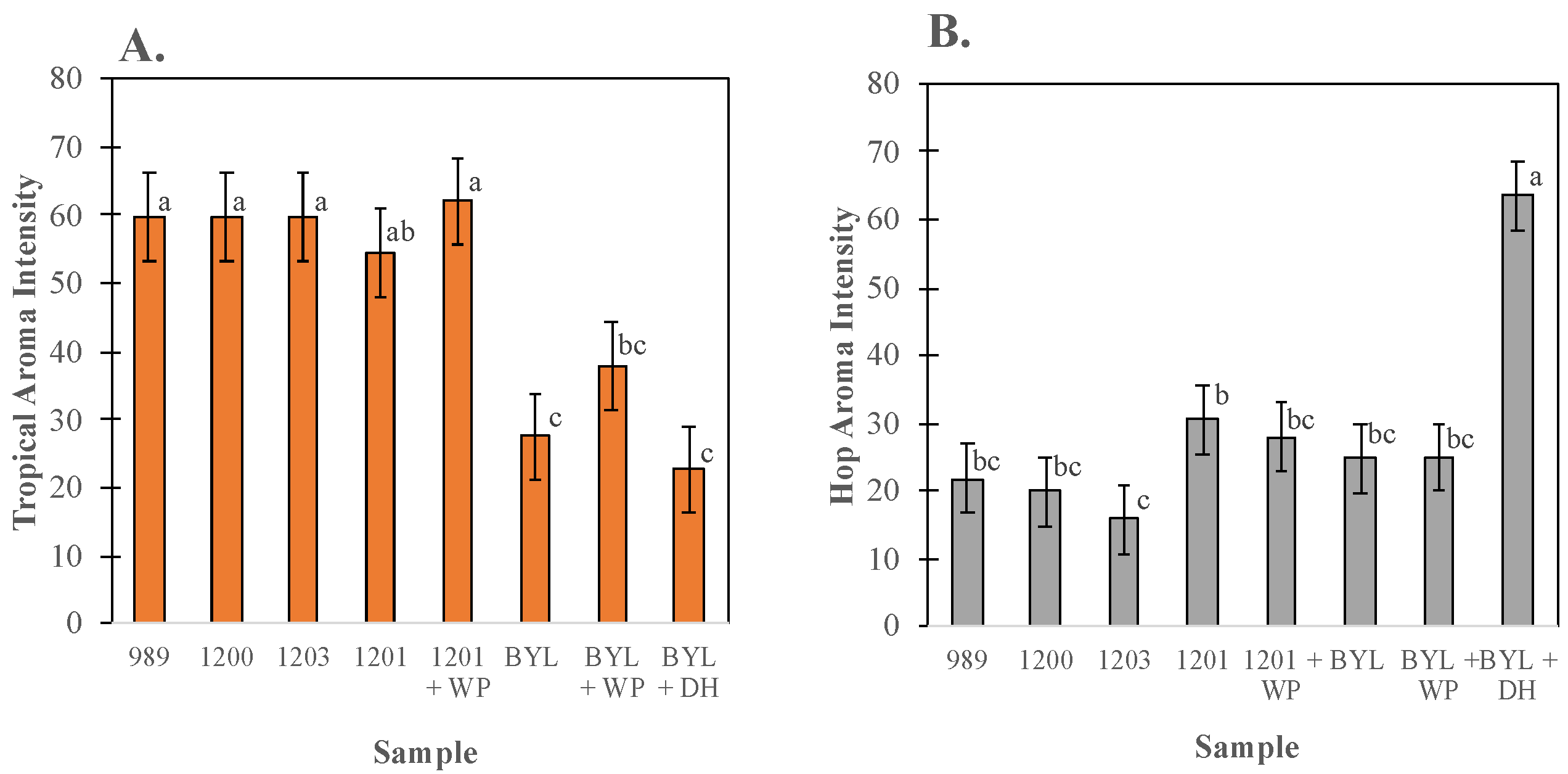
3.3. Sensory Characterization
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Biendl, M.; Engelhard, B.; Forster, A.; Gahr, A.; Lutz, A.; Mitter, W.; Schmidt, R.; Schönberger, C. Hops—Their Cultivation, Composition and Usage; Fachverlag Hans Carl: Nuremberg, Germany, 2014. [Google Scholar]
- Rettberg, N.; Biendl, M.; Garbe, L.-A. Hop Aroma and Hoppy Beer Flavor: Chemical Backgrounds and Analytical Tools—A Review. J. Am. Soc. Brew. Chem. 2018, 76, 1–20. [Google Scholar] [CrossRef]
- Eyres, G.; Dufour, J.-P. 22—Hop Essential Oil: Analysis, Chemical Composition and Odor Characteristics. In Beer in Health and Disease Prevention; Preedy, V.R., Ed.; Academic Press: Dunedin, New Zealand, 2009; pp. 239–254. [Google Scholar]
- Sharpe, F.R.; Laws, D.R.J. The essential oil of hops a review. J. Inst. Brew. 1981, 87, 96–107. [Google Scholar] [CrossRef]
- Takoi, K.; Degueil, M.; Shinkaruk, S.; Thibon, C.; Maeda, K.; Ito, K.; Bennetau, B.; Dubourdieu, D.; Tominaga, T. Identification and Characteristics of New Volatile Thiols Derived from the Hop (Humulus luplus L.) Cultivar Nelson Sauvin. J. Agric. Food Chem. 2009, 57, 2493–2502. [Google Scholar] [CrossRef]
- Lafontaine, S.R.; Shellhammer, T.H. How Hoppy Beer Production Has Redefined Hop Quality and a Discussion of Agricultural and Processing Strategies to Promote It. MBAA TQ 2019, 56, 1–12. [Google Scholar]
- Kishimoto, T.; Wanikawa, A.; Kono, K.; Shibata, K. Comparison of the Odor-Active Compounds in Unhopped Beer and Beers Hopped with Different Hop Varieties. J. Agric. Food Chem. 2006, 54, 8855–8861. [Google Scholar] [CrossRef]
- Kishimoto, T.; Morimoto, M.; Kobayashi, M.; Yako, N.; Wanikawa, A. Behaviors of 3-Mercaptohexan-1-ol and 3-Mercaptohexyl Acetate during Brewing Processes. J. Am. Soc. Brew. Chem. 2008, 66, 192–196. [Google Scholar] [CrossRef]
- Roland, A.; Schneider, R.; Razungles, A.; Cavelier, F. Varietal Thiols in Wine: Discovery, Analysis and Applications. Chem. Rev. 2011, 111, 7355–7376. [Google Scholar] [CrossRef]
- Bonnaffoux, H.; Roland, A.; Schneider, R.; Cavelier, F. Spotlight on release mechanisms of volatile thiols in beverages. Food Chem. 2021, 339, 27628. [Google Scholar] [CrossRef]
- Chenot, C.; Collin, S. Ability of the Mandarina Bavaria hop variety to release free odorant polyfunctional thiols in late-hopped beers. J. Inst. Brew. 2021, 127, 140–148. [Google Scholar] [CrossRef]
- Lafontaine, S.; Varnum, S.; Roland, A.; Delpech, S.; Dagan, L.; Vollmer, D.; Kishimoto, T.; Shellhammer, T. Impact of harvest maturity on the aroma characteristics and chemistry of Cascade hops used for dry-hopping. Food Chem. 2019, 278, 228–239. [Google Scholar] [CrossRef]
- Van Holle, A.; Van Landschoot, A.; Roldán-Ruiz, I.; Naudts, D.; De Keukeleire, D. The brewing value of Amarillo hops (Humulus lupulus L.) grown in northwestern USA: A preliminary study of terroir significance. J. Inst. Brew. 2017, 123, 312–318. [Google Scholar] [CrossRef] [Green Version]
- Staff, C.E. Results from the 2019 Australia Hop Harvest. Craft Brewing Business. Available online: https://www.craftbrewingbusiness.com/news/results-from-the-2018-australia-hop-harvest/ (accessed on 6 February 2022).
- Cannon, R.J.; Ho, C.T. Volatile sulfur compounds in tropical fruits. Food Drug Anal. 2018, 26, 445–468. [Google Scholar] [CrossRef] [PubMed]
- Holt, S.; Miks, M.H.; De Carvalho, B.T.; Foulquié-Moreno, M.R.; Thevelein, J.M. The molecular biology of fruity and floral aromas in beer and other alcoholic beverages. FEMS Microbiol. Rev. 2019, 43, 193–222. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kishimoto, T.; Kobayashi, M.; Yako, N.; Iida, A.; Wanikawa, A. Comparison of 4-Mercapto-4-methylpentan-2-one Contents in Hop Cultivars from Different Growing Regions. J. Agric. Food Chem. 2008, 56, 1051–1057. [Google Scholar] [CrossRef]
- Cordente, A.G.; Capone, D.L.; Curtin, C.D. Unravelling glutathione conjugate catabolism in Saccharomyces cerevisiae: The role of glutathione/dipeptide transporters and vacuolar function in the release of volatile sulfur compounds 3-mercaptohexan-1-ol and 4-mercapto-4-methylpentan-2-one. Appl. Microbiol. Biotechnol. 2015, 99, 9709–9722. [Google Scholar] [CrossRef] [PubMed]
- Santiago, M.; Gardner, R.C. Yeast genes required for conversion of grape precursors to varietal thiols in wine. FEMS Yeast Res. 2015, 15, fov034. [Google Scholar] [CrossRef] [Green Version]
- Myung, K.; Hamilton-Kemp, T.R.; Archbold, D.D. Biosythesis of trans-2-Hexenal in Response to Wounding in Strawberry Fruit. J. Agric. Food Chem. 2006, 54, 1442–1448. [Google Scholar] [CrossRef]
- Hirst, M.B.; Richter, C.L. Review of Aroma Formation through Metabolic Pathways of Saccharomyces cerevisiae in Beverage Fermentations. Am. J. Enol. Vitic. 2016, 67, 361–370. [Google Scholar] [CrossRef] [Green Version]
- Roland, A.L.; Schneider, R.M.; Razungles, A.; Le Guernevé, C.; Cavelier, F. Straightforward Synthesis of Deuterated Precursors to Demonstrate the Biogenesis of Aromatic Thiols in Wine. J. Agric. Food Chem. 2010, 58, 10684–10689. [Google Scholar] [CrossRef]
- Roncoroni, M.; Santiago, M.; Hooks, D.O.; Moroney, S.; Harsch, M.J.; Lee, S.A.; Richards, K.D.; Nicolau, L.; Gardner, R.C. The yeast IRC7 gene encodes a β-lyase responsible for production of the varietal thiol 4-mercapto-4-methylpentan-2-one in wine. Food Microbiol. 2011, 28, 926–935. [Google Scholar] [CrossRef]
- Swiegers, J.H.; Willmott, R.; Hill-Ling, A.; Capone, D.L.; Pardon, K.H.; Elsey, G.M.; Howell, K.S.; de Barros Lopes, M.A.; Sefton, M.A.; Lilly, M.; et al. Modulation of volatile thiol and ester aromas by modified wine yeast. Dev. Food Sci. 2006, 43, 113–116. [Google Scholar]
- Schonberger, C. Development of Thiols and Thiol Precursors in Different Hop Varieties during Hop Harvest and Their Impact on Beer Flavor. Available online: https://www.brewersassociation.org/seminars/researcher-presentation-development-of-thiols-and-thiol-precursors-in-different-hop-varieties-during-hop-harvest-and-their-impact-on-beer-flavor/ (accessed on 7 July 2022).
- Roland, A.; Viel, C.; Reillon, F.; Delpech, S.; Boivin, P.; Schneider, R.; Dagan, L. First identification and quantification of glutathionylated and cysteinylated precursors of 3-mercaptohexan-1-ol and 4-methyl-4-mercaptopentan-2-one in hops (Humulus lupulus). Flavour Fragr. 2016, 31, 455–463. [Google Scholar] [CrossRef]
- Gros, J.; Tran, T.T.H.; Collin, S. Enzymatic release of odourant polyfunctional thiols from cysteine conjugates in hop. J. Inst. Brew. 2013, 119, 221–227. [Google Scholar] [CrossRef]
- Roland, A.; Delpech, S.; Viel, C.; Reillon, F.; Schneider, R.; Dagan, L. First evidence of cysteinylated and glutathionylated precursors of 3-mercaptohexan-1-ol in malts: Toward a better aromatic potential management? In Proceedings of the World Brewing Congress, Denver, CO, USA, 13–17 August 2016. [Google Scholar]
- Lewis, M.J.; Young, T.W. Brewing, 2nd ed.; Kluwer Academic/Plenum Publishers: New York, NY, USA, 2002. [Google Scholar]
- Murat, M.; Masneuf, I.; Darriet, P.; Lavigne, V.; Tominaga, T.; Dubourdieu, D. Effect of Saccharomyces cerivisiae yeast strains on the liberation of volatile thiols in Sauvignon blanc wine. Am. J. Enol. Vitic. 2001, 53, 136–139. [Google Scholar]
- Michel, M.; Haslbeck, K.; Ampenberger, F.; Meier-Dörnberg, T.; Stretz, D.; Hutzler, M.; Coelhan, M.; Jacob, F.; Liu, Y. Screening of brewing yeast β-lyase activity and release of hop volatile thiols from precursors during fermentation. BrewingScience 2019, 72, 179–186. [Google Scholar]
- Lee, M.E.; Deloache, W.C.; Cervantes, B.; Dueber, J.E. A Highly Characterized Yeast Toolkit for Modular, Multipart Assembly. ACS Synth. Biol. 2015, 4, 975–986. [Google Scholar] [CrossRef]
- Gietz, R.D.; Woods, R.A. Transformation of yeast by lithium acetate/single-stranded carrier DNA/polyethylene glycol method. Methods Enzymol. 2002, 350, 87–96. [Google Scholar]
- Cachon, R.; Girardon, P.; Voilley, A. (Eds.) Chapter 7.5—Liquid Food Stuffs Gas Treatmends. In Gases in Agro-Food Processes; Academic Press: Dublin, Ireland, 2019; pp. 455–477. [Google Scholar]
- Beer 23. Bitterness Units (BU), Manual Isooctane Extraction (IM). Available online: https://www.asbcnet.org/Methods/BeerMethods/Pages/Beer-23-MasterMethod.aspx (accessed on 7 July 2022).
- Beer 10 (A)-Spectrophotometric Color Method. Available online: https://www.asbcnet.org/Methods/BeerMethods/Pages/default.aspx (accessed on 7 July 2022).
- Capone, D.L.; Ristic, R.; Pardon, K.H.; Jeffery, D.W. Simple Quantitative Determination of Potent Thiols at Ultratrace Levels in Wine by Derivatization and High-Performance Liquid Chromatography–Tandem Mass Spectrometry (HPLC-MS/MS) Analysis. Anal. Chem. 2015, 87, 1226–1231. [Google Scholar] [CrossRef]
- Ares, G.; Jaeger, S.R. Check-all-that-apply questions: Influence of attribute order on sensory product characterization. Food Qual. Prefer. 2013, 28, 141–153. [Google Scholar] [CrossRef]
- Lawless, H.T.; Heymann, H. Sensory Evaluation of Food: Principles and Practices; Aspen Publishers: Gaithersburg, MD, USA, 1999. [Google Scholar]
- Brewers Association. 2021 Beer Style Guidelines. Available online: https://www.brewersassociation.org/edu/brewers-association-beer-style-guidelines/ (accessed on 7 July 2022).
- Humia, B.V.; Santos, K.S.; Barbosa, A.M.; Sawata, M.; Mendonça, M.D.C.; Padilha, F.F. Beer Molecules and Its Sensory and Biological Properties: A Review. Molecules 2019, 24, 1568. [Google Scholar] [CrossRef] [Green Version]
- Seo, H.; Lee, J.-W.; Giannone, R.J.; Dunlap, N.J.; Trinh, C.T. Engineering promiscuity of chloramphenicol acetyltransferase for microbial designer ester biosynthesis. Metab. Eng. 2021, 66, 179–190. [Google Scholar] [CrossRef] [PubMed]
- Lee, T.J.; Parikh, R.Y.; Weitz, J.S.; Kim, H.D. Suppression of Expression between Adjacent Genes within Heterologous Modules in Yeast. G3 Genes Genomes Genet. 2014, 4, 109–116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, R.; Helfant, L.J.; Wu, T.; Li, Z.; Brokaw, S.E.; Stock, A.M. A balancing act in transcription regulation by response regulators: Titration of transcription factor activity by decoy DNA binding sites. Nucleic Acids Res. 2021, 49, 11537–11549. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Guo, F.; Yan, W.; Dong, W.; Zhou, J.; Zhang, W.; Xin, F.; Jiang, M. Perspectives for the microbial production of ethyl acetate. Appl. Microbiol. Biotechnol. 2020, 104, 7239–7245. [Google Scholar] [CrossRef]
- Plata, C.; Millán, C.; Mauricio, J.C.; Ortega, J.M. Formation of ethyl acetate and isoamyl acetate by various species of wine yeasts. Food Microbiol. 2003, 20, 217–224. [Google Scholar] [CrossRef]
- Wilson, C.; Brand, J.; Du Toit, W.; Buica, A. Interaction Effects of 3-Mercaptohexan-1-ol (3MH), Linalool and Ethyl Hexanoate on the Aromatic Profile of South African Dry Chenin blanc Wine by Descriptive Analysis (DA). S. Afr. J. Enol. Vitic. 2018, 39, 271–283. [Google Scholar] [CrossRef]
- Brajkovich, M.; Tibbits, N.; Peron, G.; Lund, C.M.; Dykes, S.I.; Kilmartin, P.A.; Nicolau, L. Effect of Screwcap and Cork Closures on SO2 Levels and Aromas in a Sauvignon Blanc Wine. J. Agric. Food Chem. 2005, 53, 10006–10011. [Google Scholar] [CrossRef]
- King, E.S.; Osidacz, P.; Curtin, C.; Bastian, S.E.P.; Francis, I.L. Assessing desirable levels of sensory properties in Sauvignon Blanc wines—Consumer preferences and contribution of key aroma compounds. Aust. J. Grape Wine Res. 2011, 17, 169–180. [Google Scholar] [CrossRef]
- Chambers, D.H.; Allison, A.-M.A.; Chambers, E. Training effects on performance of descriptive panelists. J. Sens. Stud. 2004, 19, 486–499. [Google Scholar] [CrossRef]
- Delwiche, J. Psychological Considerations in Sensory Analysis; Springer: New York, NY, USA, 2008; pp. 7–15. [Google Scholar]
- Chambers, E.; Koppel, K. Associations of Volatile Compounds with Sensory Aroma and Flavor: The Complex Nature of Flavor. Molecules 2013, 18, 4887–4905. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
| Strain | Background | Genotype |
|---|---|---|
| BY-London | - | wild-type |
| BY-989 | BY-London | FIG2::pPGK14-CSL11-tENO15 |
| BY-1200 | BY-London | FIG2::pPGK14-CSL11-tENO15, pHSP264-MpAAT12-tSSA15 |
| BY-1201 | BY-London | FIG2::pPGK14-CSL11-tENO15, pTDH34-MpAAT12-tSSA15 |
| BY-1203 | BY-London | FIG2::pPGK14-CSL11-tENO15, pHSP264-CATec33-tSSA15 |
| Analysis | Strain | |||||||
|---|---|---|---|---|---|---|---|---|
| 989 | 1200 | 1203 | 1201 | 1201 + WP | BYL | BYL + WP | BYL + DH | |
| Final Gravity (°P) | 4.4 | 4.4 | 4.2 | 4.3 | 4.3 | 4.2 | 4.1 | 4.4 |
| ABV (%v/v) | 4.7 | 4.7 | 4.9 | 4.9 | 4.8 | 4.9 | 5.0 | 5.1 |
| Real Extract (%w/w) | 6.3 | 6.4 | 5.9 | 6.3 | 5.9 | 6.2 | 6.2 | 6.3 |
| ADF (%w/w) | 66 | 65 | 69 | 67 | 68 | 67 | 68 | 68 |
| RDF (%) | 55 | 54 | 57 | 55 | 57 | 56 | 56 | 56 |
| BU | 20 | 21 | 18 | 27 | 31 | 17 | 32 | 44 |
| Final pH | 4.3 | 4.6 | 4.5 | 4.5 | 4.6 | 4.6 | 4.7 | 4.8 |
| Color (ASBC) | 5.6 | 6.4 | 7.5 | 7.7 | 5.7 | 5.6 | 6.1 | 7.1 |
| Time to attenuation (h) | 75 | 99 | 96 | 120 | 137 | 77 | 137 | 151 |
| Time to VDK pass (h) | 85 | 120 | 133 | 134 | 120 | 205 | 168 | 96 |
| Source | Degrees of Freedom | OHAI | Tropical Aroma Intensity | ||
|---|---|---|---|---|---|
| F | p-Value | F | p-Value | ||
| Panelist | 17 | 6.52 | <0.0001 * | 2.43 | 0.003 * |
| Sample | 7 | 8.56 | <0.0001 * | 6.23 | <0.0001 * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Molitor, R.W.; Roop, J.I.; Denby, C.M.; Depew, C.J.; Liu, D.S.; Stadulis, S.E.; Shellhammer, T.H. The Sensorial and Chemical Changes in Beer Brewed with Yeast Genetically Modified to Release Polyfunctional Thiols from Malt and Hops. Fermentation 2022, 8, 370. https://doi.org/10.3390/fermentation8080370
Molitor RW, Roop JI, Denby CM, Depew CJ, Liu DS, Stadulis SE, Shellhammer TH. The Sensorial and Chemical Changes in Beer Brewed with Yeast Genetically Modified to Release Polyfunctional Thiols from Malt and Hops. Fermentation. 2022; 8(8):370. https://doi.org/10.3390/fermentation8080370
Chicago/Turabian StyleMolitor, Richard W., Jeremy I. Roop, Charles M. Denby, Charles J. Depew, Daniel S. Liu, Sara E. Stadulis, and Thomas H. Shellhammer. 2022. "The Sensorial and Chemical Changes in Beer Brewed with Yeast Genetically Modified to Release Polyfunctional Thiols from Malt and Hops" Fermentation 8, no. 8: 370. https://doi.org/10.3390/fermentation8080370
APA StyleMolitor, R. W., Roop, J. I., Denby, C. M., Depew, C. J., Liu, D. S., Stadulis, S. E., & Shellhammer, T. H. (2022). The Sensorial and Chemical Changes in Beer Brewed with Yeast Genetically Modified to Release Polyfunctional Thiols from Malt and Hops. Fermentation, 8(8), 370. https://doi.org/10.3390/fermentation8080370