
Inhibitory and Stimulatory Effects of Fruit Bioactive Compounds on Edible Filamentous Fungi: Potential for Innovative Food Applications

 ,
,  ,
,  ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Microorganisms
2.2. Cultivation Medium and Chemicals
2.3. Analytical Methods
2.4. Statistical Analysis
3. Results and Discussions
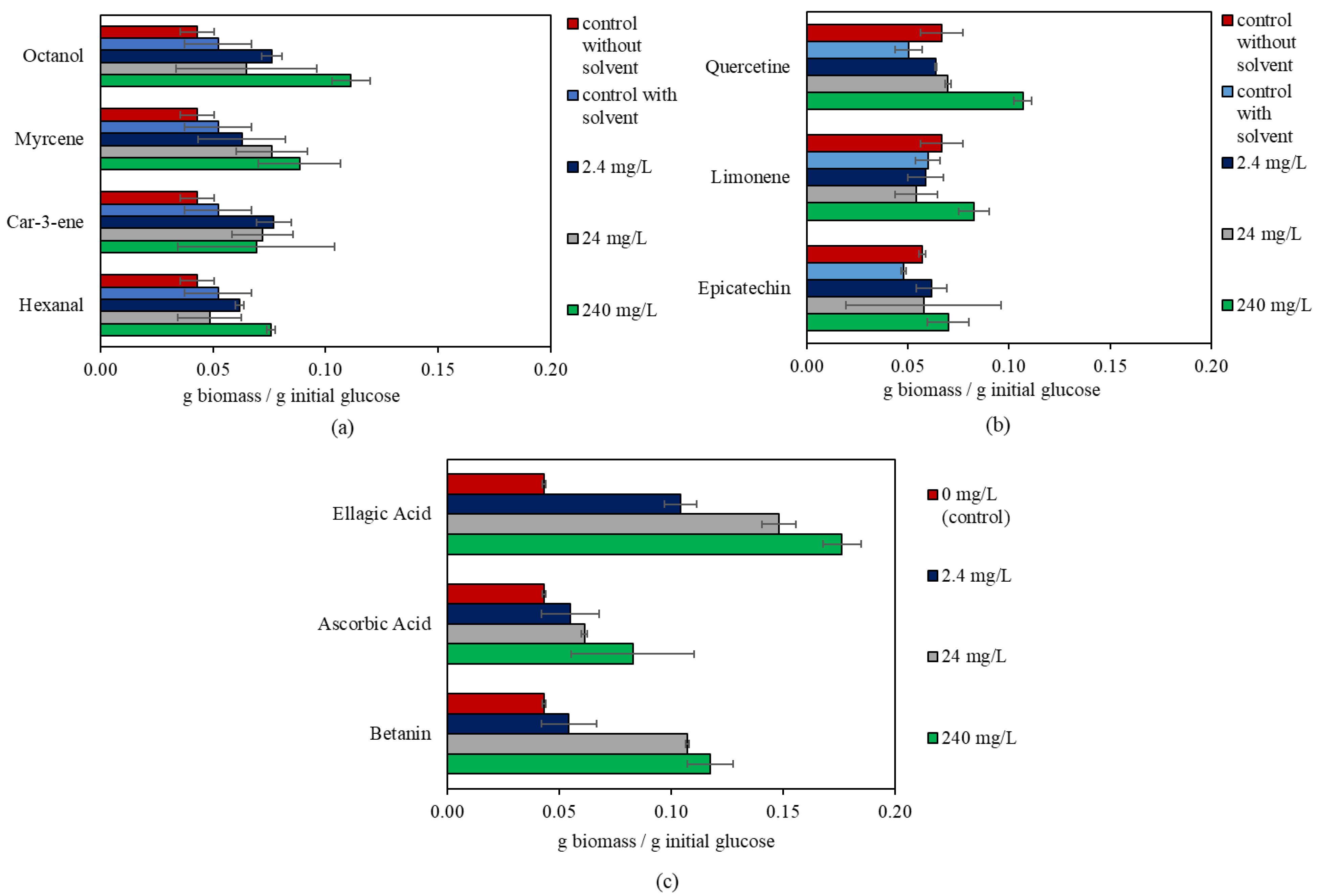
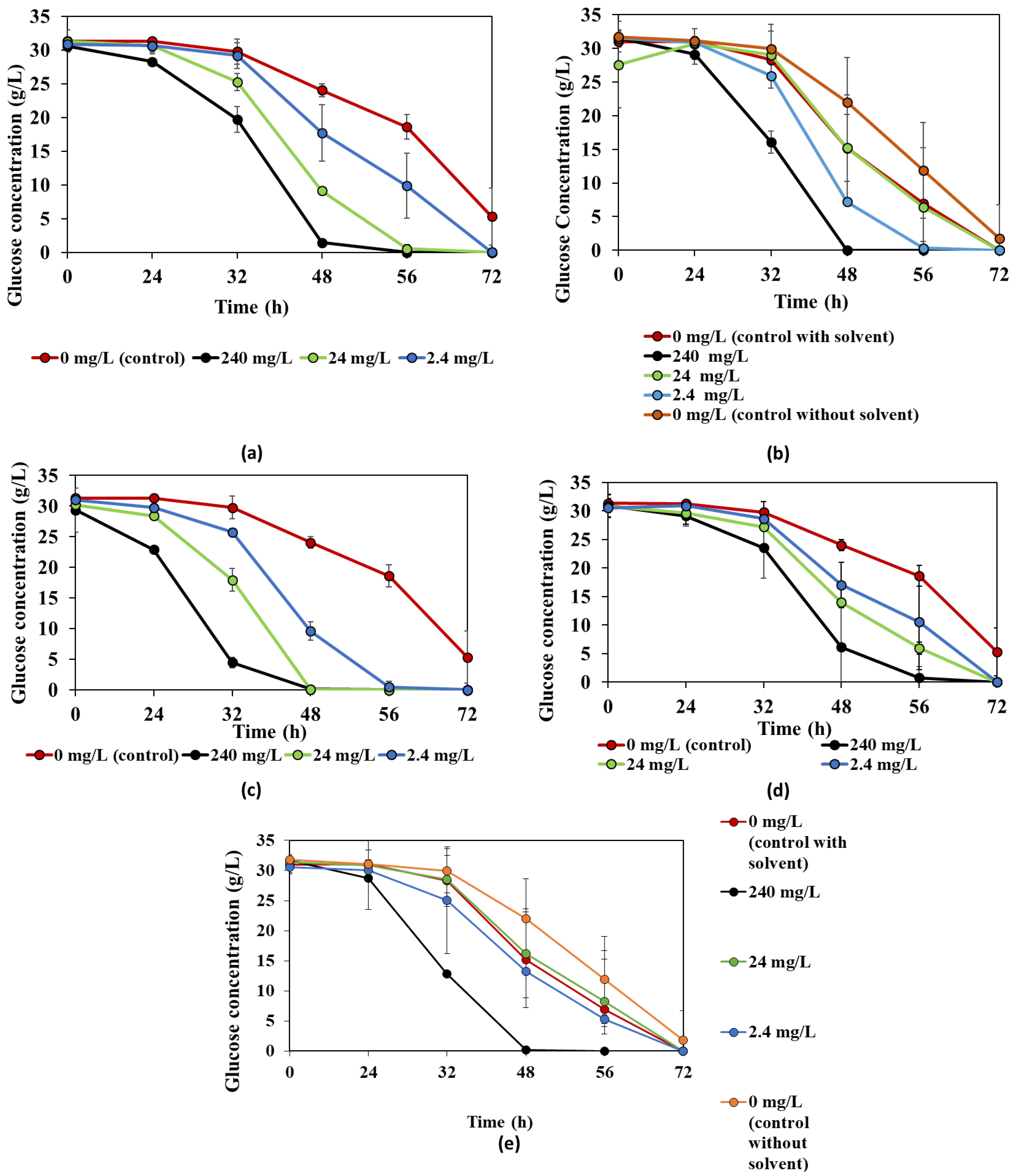
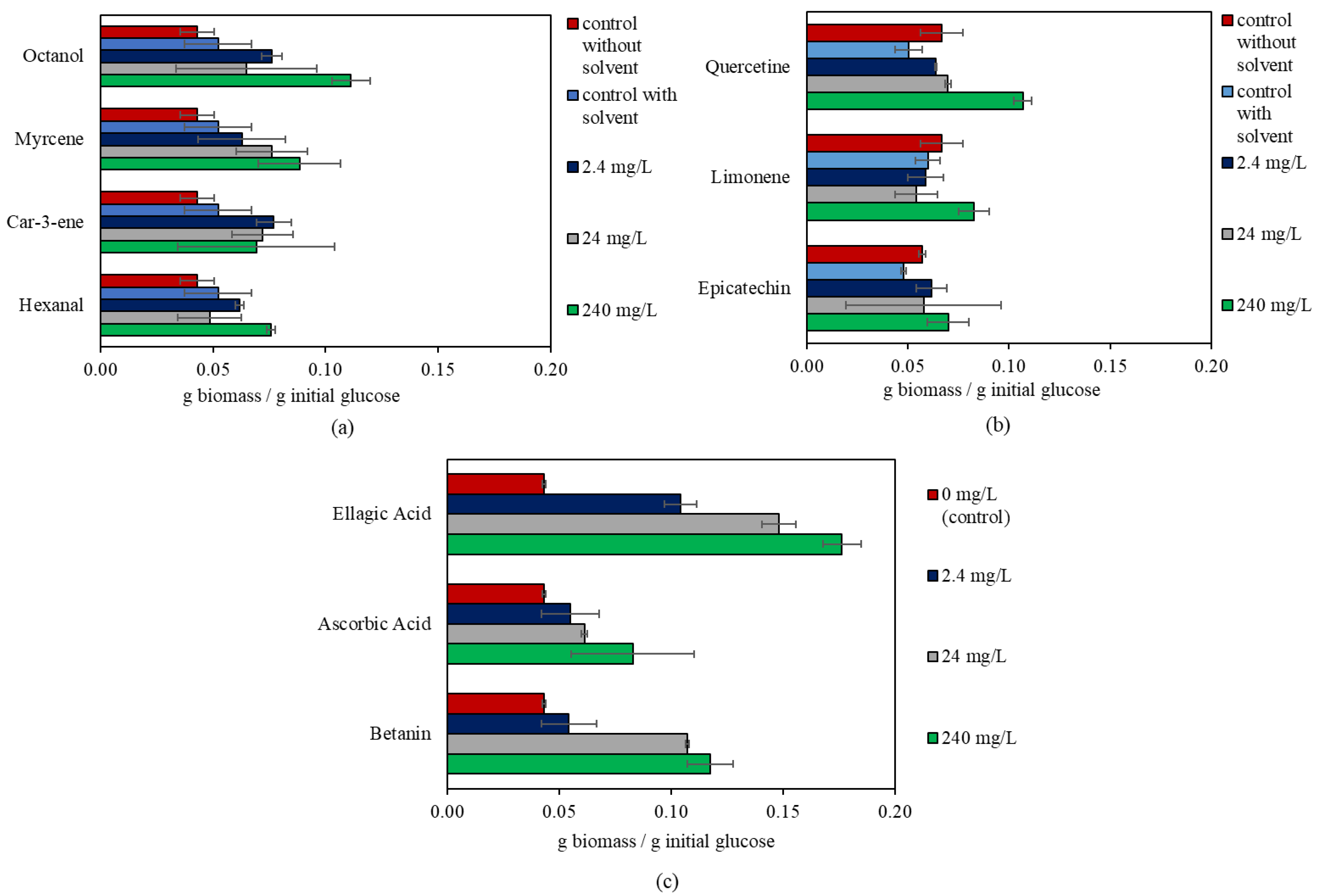
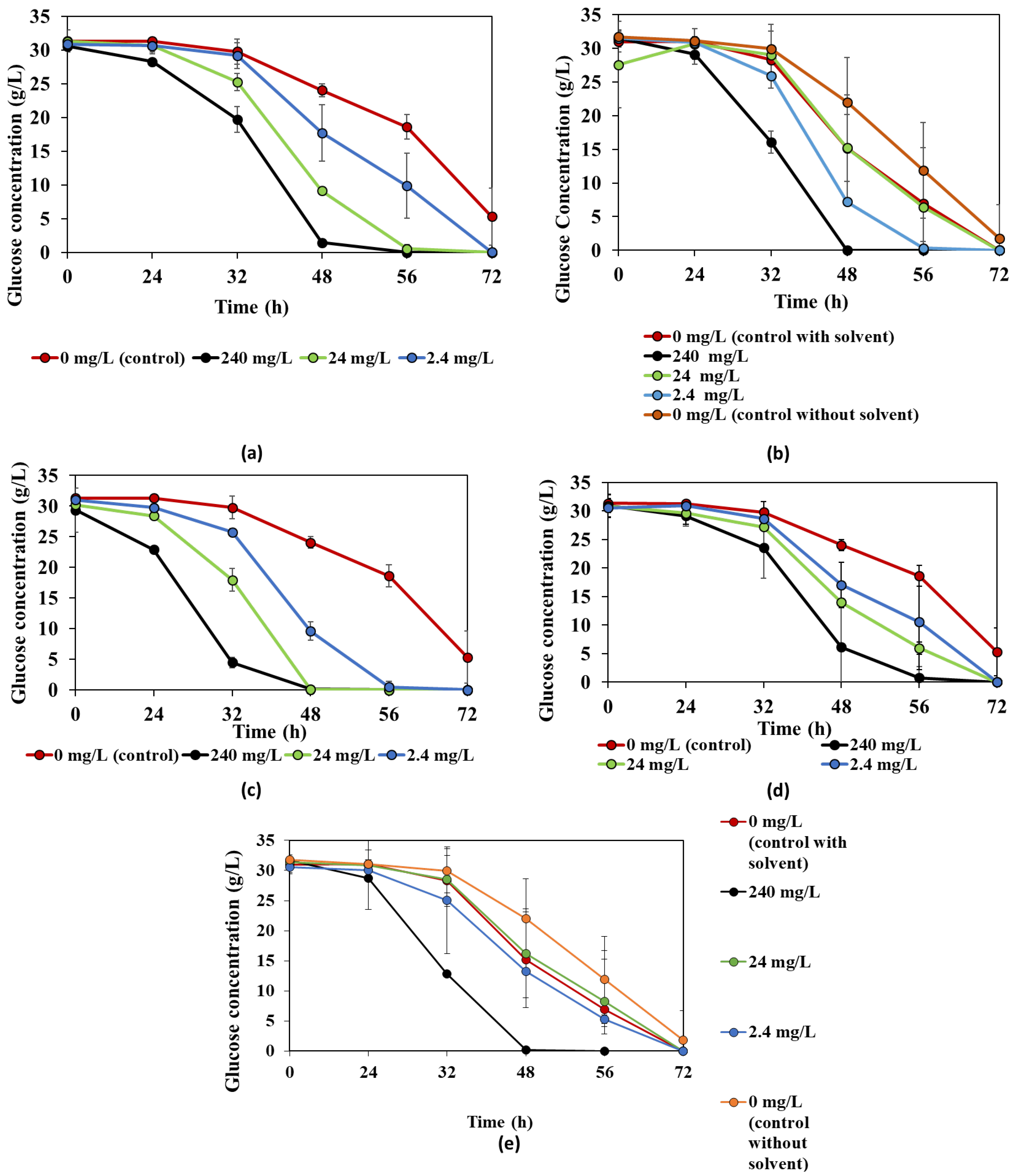
3.1. The Effect of Bioactive Compounds on R. oligosporus
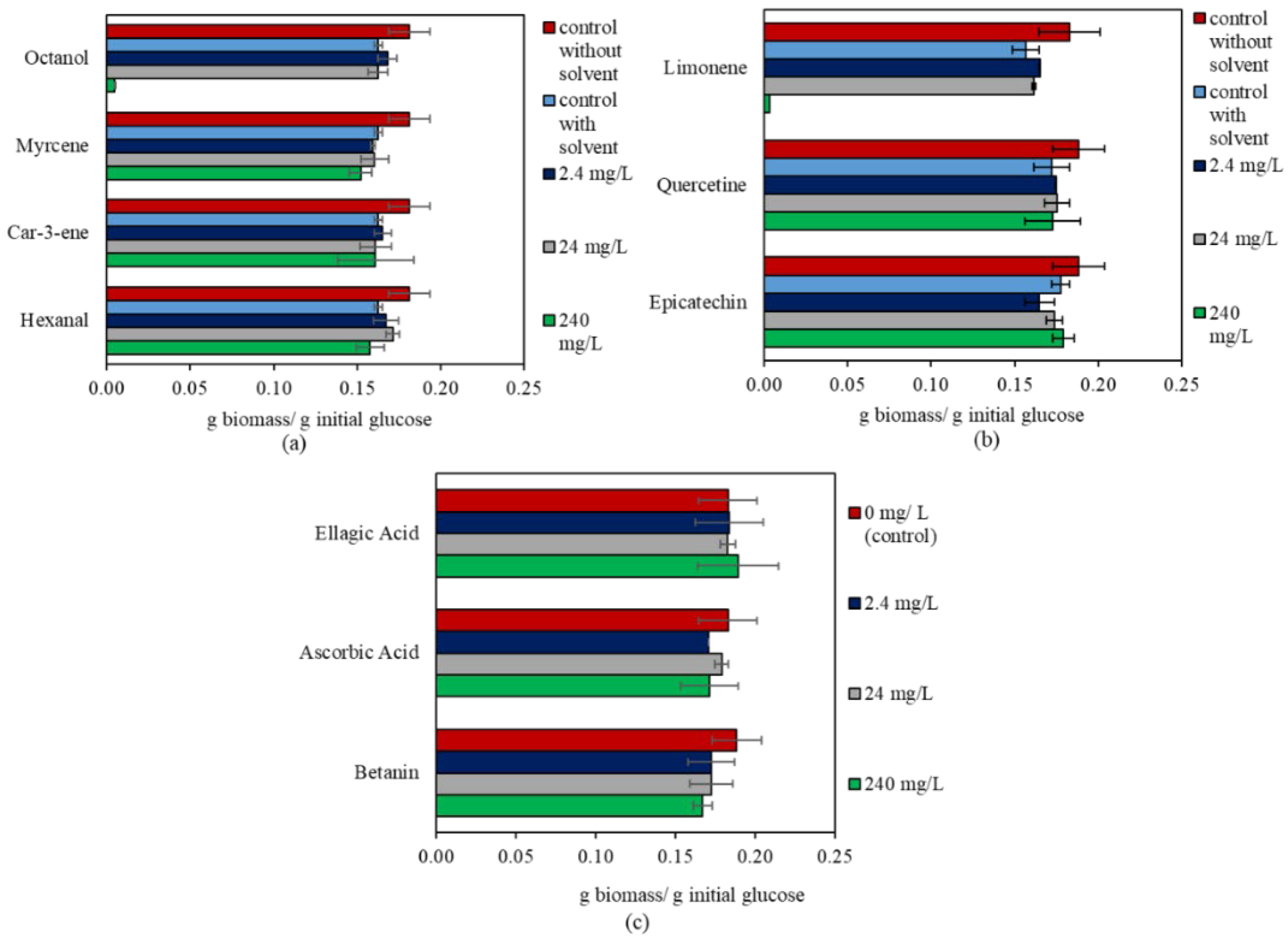
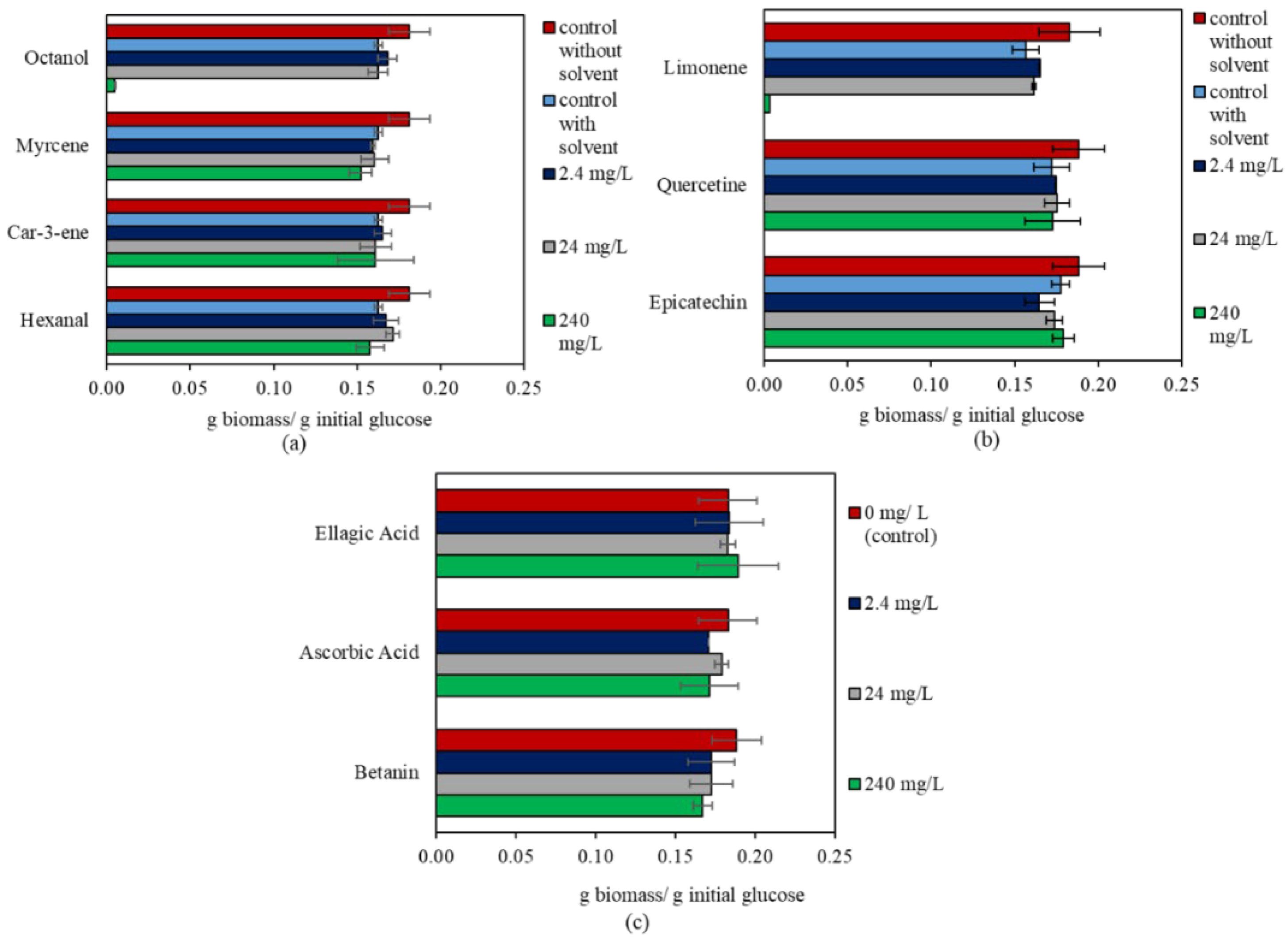
3.2. The Effect of Bioactive Compounds on N. intermedia
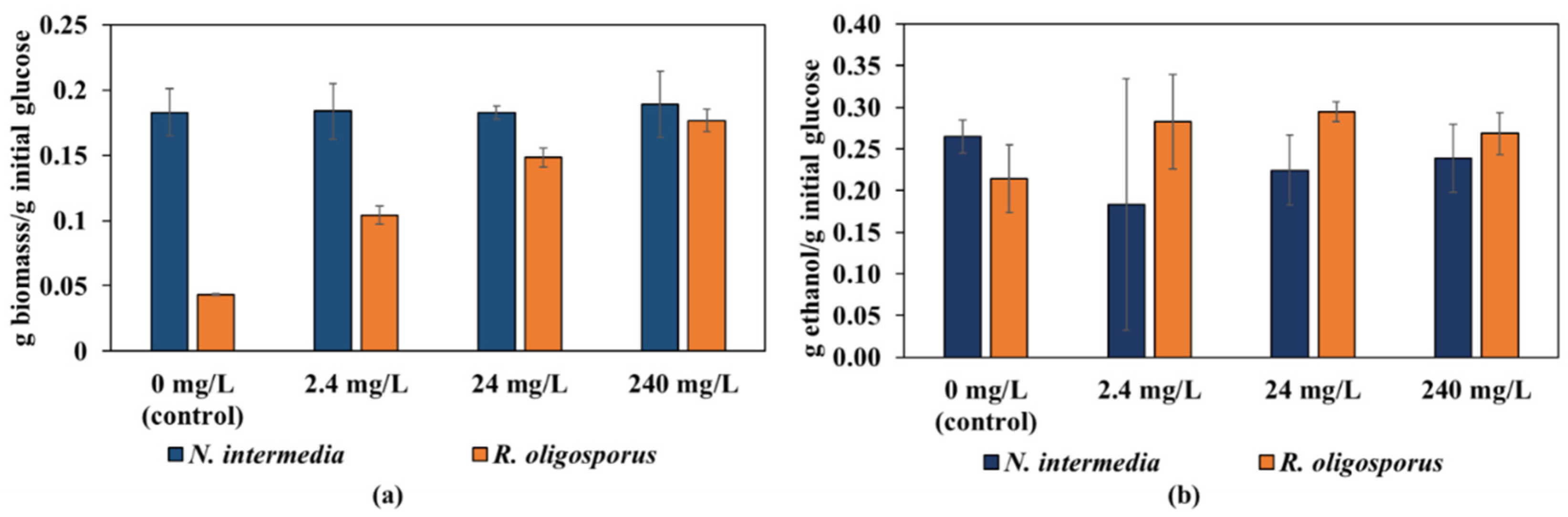
3.3. Comparison of R. oligosporus and N. intermedia
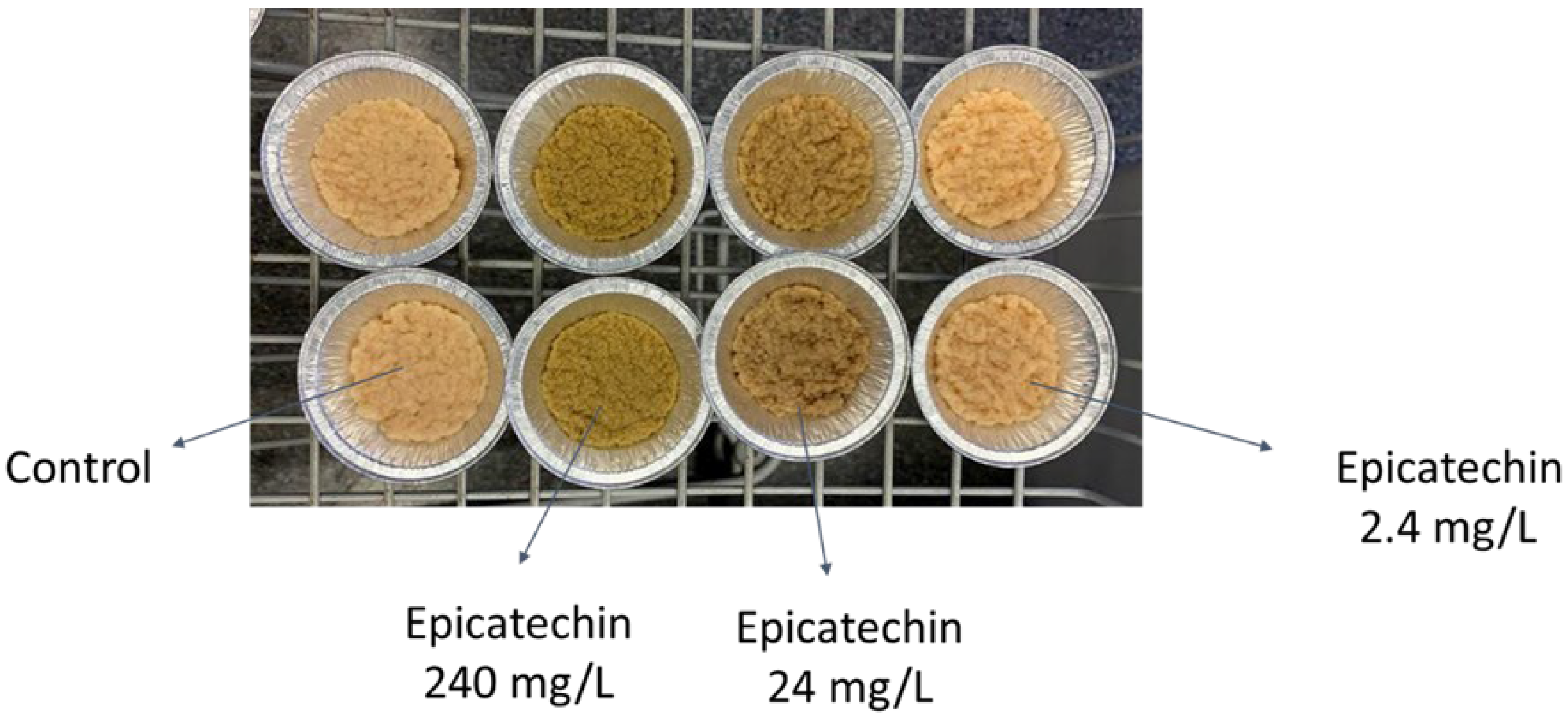
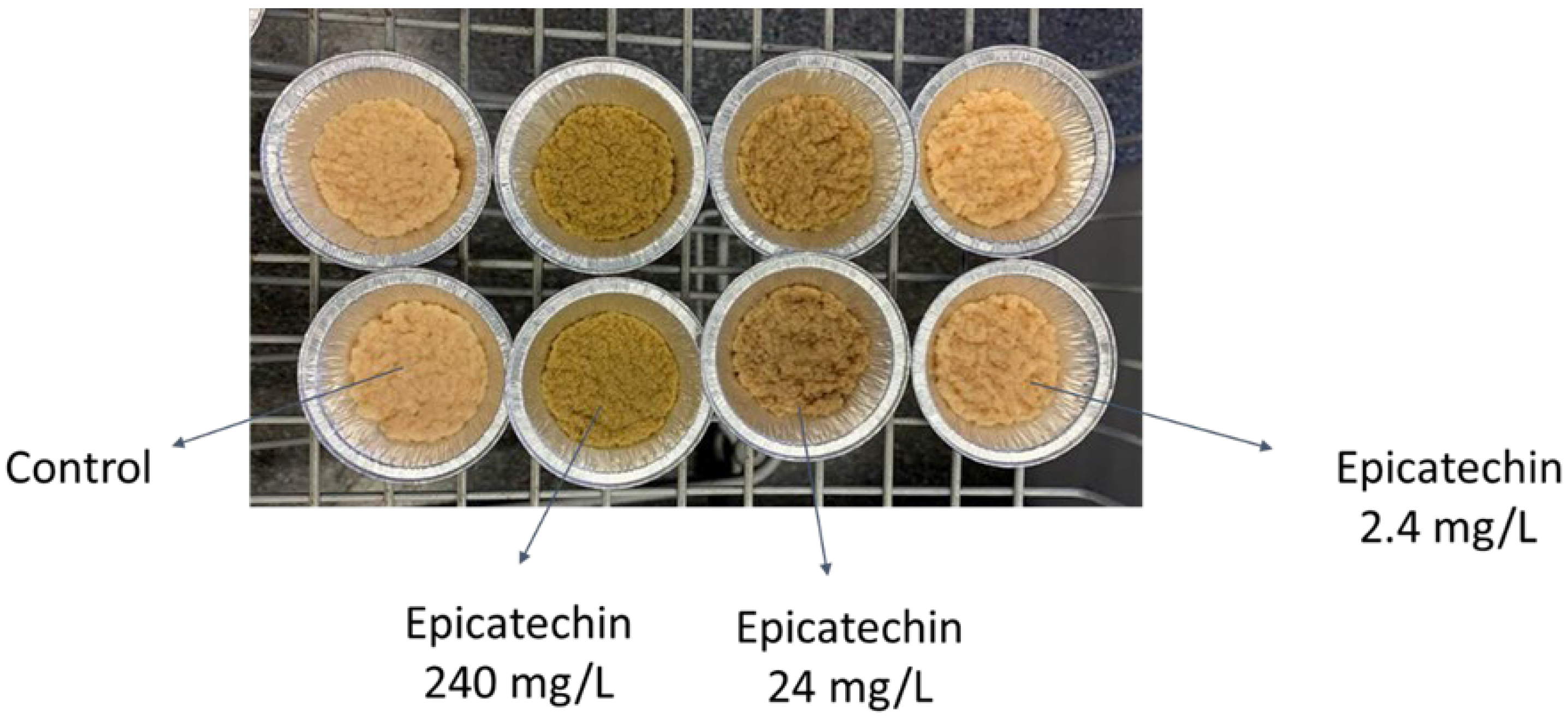
3.4. Morphology
3.5. Potential Applications of Fruit Bioactive Compounds in the Food Industry
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Javourez, U.; O’Donohue, M.; Hamelin, L. Waste-to-nutrition: A review of current and emerging conversion pathways. Biotechnol. Adv. 2021, 53, 107857. [Google Scholar] [CrossRef] [PubMed]
- Hashempour-Baltork, F.; Khosravi-Darani, K.; Hosseini, H.; Farshi, P.; Reihani, S.F.S. Mycoproteins as safe meat substitutes. J. Clean. Prod. 2020, 253, 119958. [Google Scholar] [CrossRef]
- Steinfeld, H.; Gerber, P.; Wassenaar, T.D.; Castel, V.; Rosales, M.; Rosales, M.; De Haan, C. Livestock’s Long Shadow: Environmental Issues and Options; Food & Agriculture Organization: Rome, Italy, 2006. [Google Scholar]
- Rousta, N.; Hellwig, C.; Wainaina, S.; Lukitawesa, L.; Agnihotri, S.; Rousta, K.; Taherzadeh, M.J. Filamentous fungus aspergillus oryzae for food: From submerged cultivation to fungal burgers and their sensory evaluation—A pilot study. Foods 2021, 10, 2774. [Google Scholar] [CrossRef] [PubMed]
- Gmoser, R.; Sintca, C.; Taherzadeh, M.J.; Lennartsson, P.R. Combining submerged and solid state fermentation to convert waste bread into protein and pigment using the edible filamentous fungus N. intermedia. Waste Manag. 2019, 97, 63–70. [Google Scholar] [CrossRef] [PubMed]
- Barzee, T.J.; Cao, L.; Pan, Z.; Zhang, R. Fungi for future foods. J. Future Foods 2021, 1, 25–37. [Google Scholar] [CrossRef]
- Ferreira, J. Integration of Filamentous Fungi in Ethanol Dry-Mill Biorefinery. Ph.D. Thesis, University of Borås, Borås, Sweden, 2015. [Google Scholar]
- Wikandari, R.; Hasniah, N.; Taherzadeh, M.J. The role of filamentous fungi in advancing the development of a sustainable circular bioeconomy. Bioresour. Technol. 2021, 345, 126531. [Google Scholar] [CrossRef]
- Wiebe, M. Myco-protein from Fusarium venenatum: A well-established product for human consumption. Appl. Microbiol. Biotechnol. 2002, 58, 421–427. [Google Scholar] [CrossRef]
- Wiebe, M.G. Quorn TM myco-protein-overview of a successful fungal product. Mycologist 2004, 18, 17–20. [Google Scholar] [CrossRef]
- Gmoser, R.; Fristedt, R.; Larsson, K.; Undeland, I.; Taherzadeh, M.J.; Lennartsson, P.R. From stale bread and brewers spent grain to a new food source using edible filamentous fungi. Bioengineered 2020, 11, 582–598. [Google Scholar] [CrossRef]
- Parchami, M.; Ferreira, J.; Taherzadeh, M. Brewing process development by integration of edible filamentous fungi to upgrade the quality of Brewer’s spent grain (BSG). BioResources 2021, 16, 1686–1701. [Google Scholar] [CrossRef]
- Sar, T.; Ozturk, M.; Taherzadeh, M.J.; Ferreira, J.A. New insights on protein recovery from olive oil mill wastewater through bioconversion with edible filamentous fungi. Processes 2020, 8, 1210. [Google Scholar] [CrossRef]
- Borujeni, N.E.; Karimi, K.; Denayer, J.; Kumar, R. Apple pomace biorefinery for ethanol, mycoprotein, and value-added biochemicals production by Mucor indicus. Energy 2021, 240, 122469. [Google Scholar] [CrossRef]
- Lima, T.M.D.; Almeida, A.B.D.; Peres, D.S.; Oliveira, R.M.D.S.F.D.; Sousa, T.L.D.; Freitas, B.S.M.D.; Silva, F.G.; Egea, M.B. Rhizopus oligosporus as a biotransforming microorganism of Anacardium othonianum Rizz. byproduct for production of high -protein, -antioxidant, and -fiber ingredient. LWT 2021, 135, 110030. [Google Scholar] [CrossRef]
- Jiang, Y.; Song, J. Fruits and fruit flavor: Classification and biological characterization. In Handbook of Fruit and Vegetable Flavors; John Wiley and Sons: Hoboken, NJ, USA, 2010. [Google Scholar]
- Wikandari, R. Effect of Fruit Flavors on Anaerobic Digestion: Inhibitions and Solutions. Ph.D. Thesis, University of Borås, School of Engineering, Borås, Sweden, 2014. [Google Scholar]
- Song, J.; Forney, C.F. Flavour volatile production and regulation in fruit. Can. J. Plant Sci. 2008, 88, 537–550. [Google Scholar] [CrossRef]
- Balaban, M.; Koc, C.; Sar, T.; Yesilcimen Akbas, M. Screening for bioactive compound rich pomegranate peel extracts and their antimicrobial activities. Johns. Matthey Technol. Rev. 2021, 66, 81–89. [Google Scholar] [CrossRef]
- Banerjee, J.; Singh, R.; Vijayaraghavan, R.; MacFarlane, D.; Patti, A.F.; Arora, A. Bioactives from fruit processing wastes: Green approaches to valuable chemicals. Food Chem. 2017, 225, 10–22. [Google Scholar] [CrossRef]
- Panche, A.N.; Diwan, A.D.; Chandra, S.R. Flavonoids: An overview. J. Nutr. Sci. 2016, 5, e47. [Google Scholar] [CrossRef] [Green Version]
- Balaban, M.; Koc, C.; Sar, T.; Yesilcimen Akbas, M. Antibiofilm effects of pomegranate peel extracts against B. cereus, B. subtilis and E. faecalis. Int. J. Food Sci. Technol. 2021, 56, 4915–4924. [Google Scholar] [CrossRef]
- Sagar, N.A.; Pareek, S.; Sharma, S.; Yahia, E.M.; Lobo, M.G. Fruit and vegetable waste: Bioactive compounds, their extraction, and possible utilization. Compr. Rev. Food Sci. Food Saf. 2018, 17, 512–531. [Google Scholar] [CrossRef] [Green Version]
- Vattem, D.A.; Shetty, K. Solid-state production of phenolic antioxidants from cranberry pomace by Rhizopus oligosporus. Food Biotechnol. 2002, 16, 189–210. [Google Scholar] [CrossRef]
- Wikandari, R.; Nguyen, H.; Millati, R.; Niklasson, C.; Taherzadeh, M.J. Improvement of biogas production from orange peel waste by leaching of limonene. BioMed Res. Int. 2015, 2015, 494182. [Google Scholar] [CrossRef] [PubMed]
- Albuquerque, P.M.; Koch, F.; Trossini, T.G.; Esposito, E.; Ninow, J.L. Production of Rhizopus oligosporus protein by solid state fermentation of apple pomace. Braz. Arch. Biol. Technol. 2006, 49, 91–100. [Google Scholar]
- Jin, B.; Zepf, F.; Bai, Z.; Gao, B.; Zhu, N. A biotech-systematic approach to select fungi for bioconversion of winery biomass wastes to nutrient-rich feed. Process Saf. Environ. Prot. 2016, 103, 60–68. [Google Scholar] [CrossRef]
- Kayode, R.M.O.; Sani, A. Physicochemical and proximate composition of mango (Mangifera indica) kernel cake fermented with mono-culture of fungal isolates obtained from naturally decomposed mango kernel. Life Sci. J. 2008, 5, 55–63. [Google Scholar]
- Bulkan, G.; Sitaresmi, S.; Yudhanti, G.T.; Millati, R.; Wikandari, R.; Taherzadeh, M.J. Enhancing or inhibitory effect of fruit or vegetable bioactive compound on aspergillus niger and A. Oryzae. J. Fungi 2022, 8, 12. [Google Scholar] [CrossRef]
- Ferreira, J.A.; Lennartsson, P.R.; Taherzadeh, M.J. Production of ethanol and biomass from thin stillage using food-grade Zygomycetes and Ascomycetes filamentous fungi. Energies 2014, 7, 3872–3885. [Google Scholar] [CrossRef] [Green Version]
- Sues, A.; Millati, R.; Edebo, L.; Taherzadeh, M.J. Ethanol production from hexoses, pentoses, and dilute-acid hydrolyzate by Mucor indicus. FEMS Yeast Res. 2005, 5, 669–676. [Google Scholar] [CrossRef] [Green Version]
- Yudhanti, G.T. Effect of Terpenoids, Aldehyde, and Organic Acid on Filamentous Fungal Growth and Ethanol Production. Bachelor’s Thesis, Universitas Gadjah Mada, Sleman, Indonesia, 2021. [Google Scholar]
- Sitaresmi, S. Effect of Alcohol, Phenolic Compounds, and Glucoside on Filamentous Fungal Growth and Ethanol Production: Beneficial or Detrimental? Bachelor’s Thesis, Universitas Gadjah Mada, Sleman, Indonesia, 2021. [Google Scholar]
- Ünal, M.Ü.; Ucan, F.; Şener, A.; Dincer, S.J. Research on antifungal and inhibitory effects of DL-limonene on some yeasts. Turk. J. Agric. For. 2012, 36, 576–582. [Google Scholar]
- Ferreira, J.A.; Lennartsson, P.R.; Taherzadeh, M.J. Production of ethanol and biomass from thin stillage by Neurospora intermedia: A pilot study for process diversification. Eng. Life Sci. 2015, 15, 751–759. [Google Scholar] [CrossRef] [Green Version]
- Ferreira, J.A.; Lennartsson, P.R.; Niklasson, C.; Lundin, M.; Edebo, L.; Taherzadeh, M.J. Spent sulphite liquor for cultivation of an edible Rhizopus sp. BioResources 2012, 7, 173–188. [Google Scholar]
- Rousta, N.; Ferreira, J.A.; Taherzadeh, M.J. Production of L-carnitine-enriched edible filamentous fungal biomass through submerged cultivation. Bioengineered 2021, 12, 358–368. [Google Scholar] [CrossRef] [PubMed]
- Derosa, G.; Maffioli, P.; Sahebkar, A. Ellagic Acid and Its Role in Chronic Diseases. In Anti-Inflammatory Nutraceuticals and Chronic Diseases; Gupta, S.C., Prasad, S., Aggarwal, B.B., Eds.; Springer International Publishing: Cham, Switzerland, 2016; pp. 473–479. [Google Scholar]
- Ramadan, D.T.; Ali, M.A.M.; Yahya, S.M.; El-Sayed, W.M. Correlation between Antioxidant/Antimutagenic and Antiproliferative Activity of Some Phytochemicals. Anti-Cancer Agents Med. Chem. 2019, 19, 1481–1490. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.-J.; Guo, X.; Dawuti, G.; Aibai, S. Antifungal activity of ellagic acid in vitro and in vivo. Phytother. Res. 2015, 29, 1019–1025. [Google Scholar] [CrossRef] [PubMed]
- Zamani, A. Superabsorbent polymers from the cell wall of zygomycetes fungi. Ph.D. Thesis, Chalmers University of Technology, Göteborg, Sweden, 2010. [Google Scholar]
- Garcia-Rubio, R.; de Oliveira, H.C.; Rivera, J.; Trevijano-Contador, N. The fungal cell wall: Candida, cryptococcus, and aspergillus species. Front. Microbiol. 2020, 10, 2993. [Google Scholar] [CrossRef]
- Patel, P.K.; Free, S.J. The Genetics and biochemistry of cell wall structure and synthesis in Neurospora crassa, a model filamentous fungus. Front. Microbiol. 2019, 10, 2294. [Google Scholar] [CrossRef]
- Madadi, E.; Mazloum-Ravasan, S.; Yu, J.S.; Ha, J.W.; Hamishehkar, H.; Kim, K.H. Therapeutic application of betalains: A review. Plants 2020, 9, 1219. [Google Scholar] [CrossRef]
- Hirai, I.; Okuno, M.; Katsuma, R.; Arita, N.; Tachibana, M.; Yamamoto, Y. Characterisation of anti-staphylococcus aureus activity of quercetin. Int. J. Food Sci. Technol. 2010, 45, 1250–1254. [Google Scholar] [CrossRef]
- Kapešová, J.; Petrásková, L.; Markošová, K.; Rebroš, M.; Kotik, M.; Bojarová, P.; Křen, V. Bioproduction of quercetin and rutinose catalyzed by rutinosidase: Novel concept of “solid state biocatalysis”. Int. J. Mol. Sci. 2019, 20, 1112. [Google Scholar] [CrossRef] [Green Version]
- Purewal, S.S.; Salar, R.K.; Bhatti, M.S.; Sandhu, K.S.; Singh, S.K.; Kaur, P. Solid-state fermentation of pearl millet with Aspergillus oryzae and Rhizopus azygosporus: Effects on bioactive profile and DNA damage protection activity. J. Food Meas. Charact. 2020, 14, 150–162. [Google Scholar] [CrossRef]
- Rocha, M.F.G.; Sales, J.A.; da Rocha, M.G.; Galdino, L.M.; de Aguiar, L.; Pereira-Neto, W.d.A.; de Aguiar Cordeiro, R.; Castelo-Branco, D.d.S.C.M.; Sidrim, J.J.C.; Brilhante, R.S.N. Antifungal effects of the flavonoids kaempferol and quercetin: A possible alternative for the control of fungal biofilms. Biofouling 2019, 35, 320–328. [Google Scholar] [CrossRef]
- Huynh, N.T.; Smagghe, G.; Gonzales, G.B.; Van Camp, J.; Raes, K. Bioconversion of kaempferol and quercetin glucosides from plant sources using rhizopus spp. Fermentation 2018, 4, 102. [Google Scholar] [CrossRef] [Green Version]
- Fujita, K.I.; Fujita, T.; Kubo, I. Antifungal activity of alkanols against Zygosaccharomyces bailii and their effects on fungal plasma membrane. Phytother. Res. Int. J. Devoted Pharmacol. Toxicol. Eval. Nat. Prod. Deriv. 2008, 22, 1349–1355. [Google Scholar] [CrossRef] [PubMed]
- Miller, J.A.; Thompson, P.A.; Hakim, I.A.; Chow, H.H.S.; Thomson, C.A. d-Limonene: A bioactive food component from citrus and evidence for a potential role in breast cancer prevention and treatment. Oncol. Rev. 2011, 5, 31–42. [Google Scholar] [CrossRef]
- Lennartsson, P.R.; Ylitervo, P.; Larsson, C.; Edebo, L.; Taherzadeh, M.J. Growth tolerance of zygomycetes mucor indicus in orange peel hydrolysate without detoxification. Process Biochem. 2012, 47, 836–842. [Google Scholar] [CrossRef]
- Bier, M.C.J.; Medeiros, A.B.P.; Soccol, C.R. Biotransformation of limonene by an endophytic fungus using synthetic and orange residue-based media. Fungal Biol. 2017, 121, 137–144. [Google Scholar] [CrossRef]
- Setzer, W.N.; Vogler, B.; Schmidt, J.M.; Leahy, J.G.; Rives, R. Antimicrobial activity of Artemisia douglasiana leaf essential oil. Fitoterapia 2004, 75, 192–200. [Google Scholar] [CrossRef]
- Li, S.-F.; Zhang, S.-B.; Zhai, H.-C.; Lv, Y.-Y.; Hu, Y.-S.; Cai, J.-P. Hexanal induces early apoptosis of Aspergillus flavus conidia by disrupting mitochondrial function and expression of key genes. Appl. Microbiol. Biotechnol. 2021, 105, 6871–6886. [Google Scholar] [CrossRef]
- Millati, R.; Edebo, L.; Taherzadeh, M.J. Performance of rhizopus, rhizomucor, and mucor in ethanol production from glucose, xylose, and wood hydrolyzates. Enzym. Microb. Technol. 2005, 36, 294–300. [Google Scholar] [CrossRef]
- Pietrzak, W.; Kawa-Rygielska, J. Backset valorization in dry-grind ethanol process by co-culture of edible filamentous fungi and fodder yeast. J. Clean. Prod. 2019, 220, 376–385. [Google Scholar] [CrossRef]
- Taherzadeh, M.J.; Fox, M.; Hjorth, H.; Edebo, L. Production of mycelium biomass and ethanol from paper pulp sulfite liquor by Rhizopus oryzae. Bioresour. Technol. 2003, 88, 167–177. [Google Scholar] [CrossRef]
- Zhang, Z.Y.; Jin, B.; Kelly, J.M. Production of lactic acid from renewable materials by Rhizopus fungi. Biochem. Eng. J. 2007, 35, 251–263. [Google Scholar] [CrossRef]
- Marei, G.I.K.; Rasoul, M.A.A.; Abdelgaleil, S.A. Comparative antifungal activities and biochemical effects of monoterpenes on plant pathogenic fungi. Pestic. Biochem. Physiol. 2012, 103, 56–61. [Google Scholar] [CrossRef]
- Yassin, S.; Wheals, A. Neurospora species in bakeries. J. Appl. Bacteriol. 1992, 72, 377–380. [Google Scholar] [CrossRef]
- Beltran-Garcia, M.J.; Estarron-Espinosa, M.; Ogura, T. Volatile compounds secreted by the oyster mushroom (Pleurotus ostreatus) and their antibacterial activities. J. Agric. Food Chem. 1997, 45, 4049–4052. [Google Scholar] [CrossRef]
- Nweke, C.O.; Okpokwasili, G.C. Removal of phenol from aqueous solution by adsorption onto activated carbon and fungal biomass. Int. J. Biosci. 2013, 3, 11–21. [Google Scholar]
- Aranda-Martinez, A.; Lopez-Moya, F.; Lopez-Llorca, L.V. Cell wall composition plays a key role on sensitivity of filamentous fungi to chitosan. J. Basic Microbiol. 2016, 56, 1059–1070. [Google Scholar] [CrossRef]
- Hasanin, M.S.; Hashem, A.H.; Abd El-Sayed, E.S.; El-Saied, H. Green ecofriendly bio-deinking of mixed office waste paper using various enzymes from Rhizopus microsporus AH3: Efficiency and characteristics. Cellulose 2020, 27, 4443–4453. [Google Scholar] [CrossRef]
- Jiménez, M.; García-Carmona, F. Oxidation of the flavonol quercetin by polyphenol oxidase. J. Agric. Food Chem. 1999, 47, 56–60. [Google Scholar] [CrossRef]
- Nedele, A.-K.; Schiebelbein, R.; Bär, A.; Kaup, A.; Zhang, Y. Reduction of aldehydes with green odor in soy products during fermentation with Lycoperdon pyriforme and analysis of their degradation products. Food Res. Int. 2022, 152, 110909. [Google Scholar] [CrossRef]
- Thitiprasert, S.; Sooksai, S.; Thongchul, N. In vivo regulation of alcohol dehydrogenase and lactate dehydrogenase in rhizopus oryzae to improve L-lactic acid fermentation. Appl. Biochem. Biotechnol. 2011, 164, 1305–1322. [Google Scholar] [CrossRef]
- Wainaina, S.; Kisworini, A.D.; Fanani, M.; Wikandari, R.; Millati, R.; Niklasson, C.; Taherzadeh, M.J. Utilization of food waste-derived volatile fatty acids for production of edible Rhizopus oligosporus fungal biomass. Bioresour. Technol. 2020, 310, 123444. [Google Scholar] [CrossRef]
- Xu, Q.; Li, S.; Huang, H.; Wen, J. Key technologies for the industrial production of fumaric acid by fermentation. Biotechnol. Adv. 2012, 30, 1685–1696. [Google Scholar] [CrossRef] [PubMed]
- Nair, R.B.; Lennartsson, P.R.; Taherzadeh, M.J. Mycelial pellet formation by edible ascomycete filamentous fungi, Neurospora intermedia. AMB Express 2016, 6, 31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cox, P.W.; Thomas, C.R. Classification and measurement of fungal pellets by automated image analysis. Biotechnol. Bioeng. 1992, 39, 945–952. [Google Scholar] [CrossRef] [PubMed]
- Souza Filho, P.F.; Zamani, A.; Taherzadeh, M.J. Production of edible fungi from potato protein liquor (PPL) in airlift bioreactor. Fermentation 2017, 3, 12. [Google Scholar] [CrossRef] [Green Version]
- Nair, R.B.; Osadolor, O.A.; Ravula, V.K.; Lennartsson, P.R.; Taherzadeh, M.J. Lignocellulose integration to 1G-ethanol process using filamentous fungi: Fermentation prospects of edible strain of Neurospora intermedia. BMC Biotechnol. 2018, 18, 49. [Google Scholar] [CrossRef] [Green Version]
- Bushara, K.O.; Goldstein, S.R.; Grimes, G.J.; Burstein, A.H.; Hallett, M. Pilot trial of 1-octanol in essential tremor. Neurology 2004, 62, 122–124. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Bioactive Compound | Concentration (mg/L) | p-Value | g Biomass/g Initial Glucose |
|---|---|---|---|
| Betanin | 0 | 0.000 | 0.04 ± 0.00 b |
| 2.4 | 0.05 ± 0.01 b | ||
| 24 | 0.11 ± 0.00 a | ||
| 240 | 0.12 ± 0.01 a | ||
| Ascorbic Acid | 0 | 0.027 | 0.04 ± 0.00 b |
| 2.4 | 0.06 ± 0.01 ab | ||
| 24 | 0.06 ± 0.00 ab | ||
| 240 | 0.08 ± 0.03 a | ||
| Ellagic Acid | 0 | 0.000 | 0.04 ± 0.00 a |
| 2.4 | 0.10 ± 0.01 b | ||
| 24 | 0.15 ± 0.01 c | ||
| 240 | 0.18 ± 0.01 d | ||
| Limonene | 0 | 0.008 | 0.06 ± 0.01 b |
| 2.4 | 0.06 ± 0.01 b | ||
| 24 | 0.05 ± 0.01 b | ||
| 240 | 0.08 ± 0.01 a | ||
| Quercetin | 0 | 0.000 | 0.05 ± 0.01 c |
| 2.4 | 0.06 ± 0.00 b | ||
| 24 | 0.07 ± 0.00 b | ||
| 240 | 0.11 ± 0.00 a | ||
| Hexanal | 0 | 0.021 | 0.05 ± 0.01 b |
| 2.4 | 0.06 ± 0.00 ab | ||
| 24 | 0.05 ± 0.01 b | ||
| 240 | 0.08 ± 0.00 a | ||
| Myrcene | 0 | 0.048 | 0.05 ± 0.01 b |
| 2.4 | 0.06 ± 0.02 ab | ||
| 24 | 0.08 ± 0.02 ab | ||
| 240 | 0.09 ± 0.02 a | ||
| Octanol | 0 | 0.000 | 0.05 ± 0.01 b |
| 2.4 | 0.08 ± 0.00 ab | ||
| 24 | 0.06 ± 0.03 b | ||
| 240 | 0.11 ± 0.01 a |
| Metabolite | Bioactive Compound | Concentration (mg/L) | p-Value | g Metabolite/g Initial Glucose |
|---|---|---|---|---|
| Ethanol | Betanin | 0 | 0.012 | 0.21 ± 0.04 b |
| 2.4 | 0.23 ± 0.03 ab | |||
| 24 | 0.28 ± 0.01 a | |||
| 240 | 0.28 ± 0.00 a | |||
| Ellagic Acid | 0 | 0.044 | 0.21 ± 0.04 b | |
| 2.4 | 0.28 ± 0.06 ab | |||
| 24 | 0.29 ± 0.01 a | |||
| 240 | 0.27 ± 0.03 ab | |||
| Lactic Acid | Betanin | 0 | 0.036 | 0.18 ± 0.03 ab |
| 2.4 | 0.18 ± 0.01 b | |||
| 24 | 0.22 ± 0.00 ab | |||
| 240 | 0.23 ± 0.04 a | |||
| Ellagic Acid | 0 | 0.002 | 0.18 ± 0.03 a | |
| 2.4 | 0.14 ± 0.00 ab | |||
| 24 | 0.12 ± 0.00 b | |||
| 240 | 0.07 ± 0.03 c | |||
| Epicatechin | 0 | 0.034 | 0.15± 0.03 ab | |
| 2.4 | 0.17± 0.00 a | |||
| 24 | 0.17± 0.01 a | |||
| 240 | 0.13± 0.01 b | |||
| Quercetin | 0 | 0.000 | 0.22 ± 0.04 a | |
| 2.4 | 0.18 ± 0.00 a | |||
| 24 | 0.09± 0.00 b | |||
| 240 | 0.04± 0.03 c | |||
| Hexanal | 0 | 0.002 | 0.19 ± 0.01 b | |
| 2.4 | 0.18± 0.01 bc | |||
| 24 | 0.18± 0.00 c | |||
| 240 | 0.21± 0.00 a | |||
| Myrcene | 0 | 0.021 | 0.19± 0.01 a | |
| 2.4 | 0.15± 0.01 b | |||
| 24 | 0.17± 0.02 ab | |||
| 240 | 0.17± 0.01 ab | |||
| Octanol | 0 | 0.044 | 0.19± 0.01 a | |
| 2.4 | 0.17± 0.00 ab | |||
| 24 | 0.14± 0.03 b | |||
| 240 | 0.15± 0.03 ab |
| Bioactive Compound | Concentration (mg/L) | p-Value | g Biomass/g Initial Glucose |
|---|---|---|---|
| Epicatechin | 0 | 0.043 | 0.18 ± 0.01 ab |
| 2.4 | 0.16 ± 0.01 b | ||
| 24 | 0.17 ± 0.01 ab | ||
| 240 | 0.18 ± 0.01 a | ||
| Limonene | 0 | 0.000 | 0.16 ± 0.01 b |
| 2.4 | 0.16 ± 0.00 a | ||
| 24 | 0.16 ± 0.00 ab | ||
| 240 | 0.00 ± 0.00 c | ||
| Hexanal | 0 | 0.041 | 0.16 ± 0.00 ab |
| 2.4 | 0.17 ± 0.01 ab | ||
| 24 | 0.17 ± 0.00 a | ||
| 240 | 0.16 ± 0.01 b | ||
| Octanol | 0 | 0.000 | 0.16 ± 0.00 a |
| 2.4 | 0.17 ± 0.01 a | ||
| 24 | 0.16 ± 0.01 a | ||
| 240 | 0.00 ± 0.00 b |
| Bioactive Compound | Concentration (mg/L) | g Biomass/g Initial Glucose | Morphology | Final pH |
|---|---|---|---|---|
| Ellagic acid 1 | 0 | 0.04 ± 0.00 | Compact, round pellets with a smooth outer surface. | 3.5 |
| 2.4 | 0.10 ± 0.01 | Mixture of round and tubular-shaped pellets with a slightly fluffy (hairy) outer surface. | 5.2–5.5 | |
| 24 | 0.15 ± 0.01 | Mixture of round and tubular-shaped pellets with fluffy (hairy) structure. | 5 | |
| 240 | 0.18 ± 0.01 | Mycelial clumps. | 5 | |
| Betanin 1 | 0 | 0.04 ± 0.00 | Compact, round pellets with a smooth outer surface. | 3.5 |
| 2.4 | 0.05 ± 0.01 | Mixture of big and small pellets with a round shape and slightly hairy surface. | 4 | |
| 24 | 0.11 ± 0.00 | Mixture of round and tubular-shaped pellets with fluffy (hairy) structure. | 5 | |
| 240 | 0.12 ± 0.01 | Round pellets with fluffy (hairy) structure. | 5 | |
| Octanol 1 | 0–24 | 0.05–0.08 | Round pellets. | 4.5–5 |
| 240 | 0.11 ± 0.01 | Smaller and spherical pellets. | 5.3 | |
| Quercetin 2 | 0–2.4 | No significant change in biomass yield | Mixture of pellets and mycelium. | 3.8–4.7 |
| 24 | Pellets. | 4–4.5 | ||
| 240 | Smaller pellets. | 4.5 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bulkan, G.; Yudhanti, G.T.; Sitaresmi, S.; Millati, R.; Wikandari, R.; Taherzadeh, M.J. Inhibitory and Stimulatory Effects of Fruit Bioactive Compounds on Edible Filamentous Fungi: Potential for Innovative Food Applications. Fermentation 2022, 8, 270. https://doi.org/10.3390/fermentation8060270
Bulkan G, Yudhanti GT, Sitaresmi S, Millati R, Wikandari R, Taherzadeh MJ. Inhibitory and Stimulatory Effects of Fruit Bioactive Compounds on Edible Filamentous Fungi: Potential for Innovative Food Applications. Fermentation. 2022; 8(6):270. https://doi.org/10.3390/fermentation8060270
Chicago/Turabian StyleBulkan, Gülru, Gerarda Tania Yudhanti, Sitaresmi Sitaresmi, Ria Millati, Rachma Wikandari, and Mohammad J. Taherzadeh. 2022. "Inhibitory and Stimulatory Effects of Fruit Bioactive Compounds on Edible Filamentous Fungi: Potential for Innovative Food Applications" Fermentation 8, no. 6: 270. https://doi.org/10.3390/fermentation8060270
APA StyleBulkan, G., Yudhanti, G. T., Sitaresmi, S., Millati, R., Wikandari, R., & Taherzadeh, M. J. (2022). Inhibitory and Stimulatory Effects of Fruit Bioactive Compounds on Edible Filamentous Fungi: Potential for Innovative Food Applications. Fermentation, 8(6), 270. https://doi.org/10.3390/fermentation8060270