
Interaction between Chocolate Polyphenols and Encapsulated Probiotics during In Vitro Digestion and Colonic Fermentation



Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Reagents
2.2. Methods
2.2.1. Preparation of Encapsulated Probiotics
2.2.2. The In Vitro Gastrointestinal Digestion and Colonic Fermentation of Probiotic-Chocolate
2.2.3. Extraction of Polyphenols from the Digested/Fermented Chocolate Samples
Analysis of Total Polyphenols Content in the Extracted Supernatant
Analyses of Individual Flavonoids Using an HPLC Technique
2.3. Statistical Analysis
3. Results
3.1. Total Polyphenol Bioaccessibility in Chocolate Fortified with Encapsulated or Non-Encapsulated Probiotics during the In Vitro Gastrointestinal Digestion and Colonic Fermentation
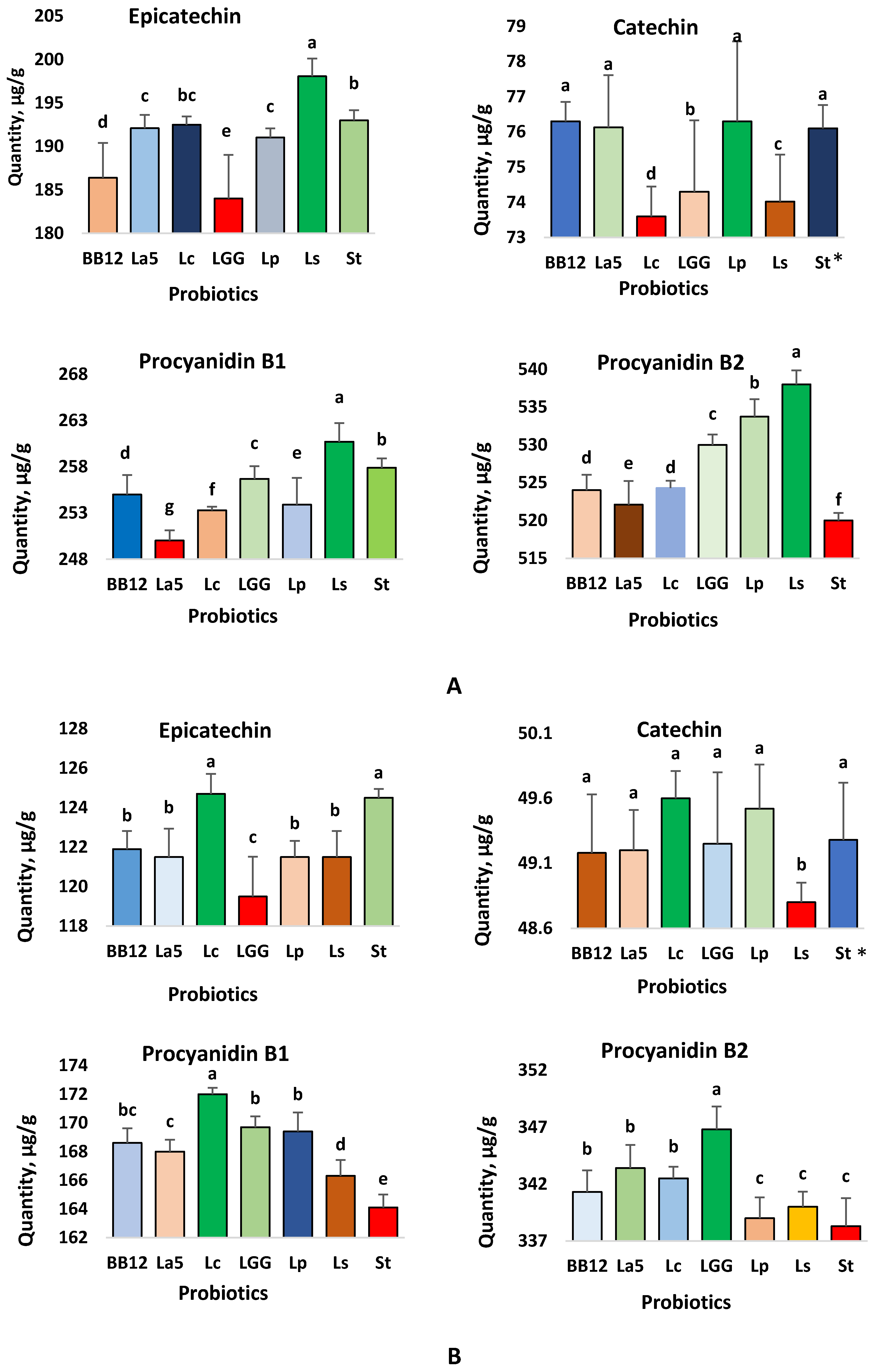
3.2. In Vitro Bioaccessibility of Individual Flavonoids from the Probiotic-Chocolates
3.2.1. Probiotic-Chocolate Containing 70% Cocoa Powder
3.2.2. Probiotic-Chocolate Containing 45% Cocoa Powder
3.3. Interaction between Flavonoids and Encapsulated Probiotics in the Chocolates
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Granato, D.; Barba, F.J.; Kovačević, D.B.; Lorenzo, J.M.; Cruz, A.G.; Putnik, P. Functional Foods: Product Development, Technological Trends, Efficacy Testing, and Safety. Annu. Rev. Food Sci. Technol. 2020, 11, 93–118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Min, M.; Bunt, C.R.; Mason, S.L.; Hussain, M.A. Non-dairy probiotic food products: An emerging group of functional foods. Crit. Rev. Food Sci. Nutr. 2019, 59, 2626–2641. [Google Scholar] [CrossRef] [PubMed]
- Socci, V.; Tempesta, D.; Desideri, G.; De Gennaro, L.; Ferrara, M. Enhancing human cognition with cocoa flavonoids. Front. Nutr. 2017, 4, 19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martín, M.A.; Ramos, S. Cocoa polyphenols in oxidative stress: Potential health implications. J. Funct. Foods 2016, 27, 570–588. [Google Scholar] [CrossRef] [Green Version]
- Davinelli, S.; Corbi, G.; Righetti, S.; Sears, B.; Olarte, H.H.; Grassi, D.; Scapagnini, G. Cardioprotection by cocoa polyphenols and ω-3 fatty acids: A disease-prevention perspective on aging-associated cardiovascular risk. J. Med. Food 2018, 21, 1060–1069. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Álvarez-Cilleros, D.; Ramos, S.; López-Oliva, M.E.; Escrivá, F.; Álvarez, C.; Fernández-Millán, E.; Martin, M.A. Cocoa diet modulates gut microbiota composition and improves intestinal health in Zucker diabetic rats. Food Res. Int. 2020, 132, 109058. [Google Scholar] [CrossRef] [PubMed]
- Kerimi, A.; Williamson, G. The cardiovascular benefits of dark chocolate. Vascul. Pharmacol. 2015, 71, 11–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santos, D.I.; Saraiva, J.M.A.; Vicente, A.A.; Moldão-Martins, M. Methods for determining bioavailability and bioaccessibility of bioactive compounds and nutrients. In Innovative Thermal and Non-Thermal Processing, Bioaccessibility and Bioavailability of Nutrients and Bioactive Compounds; Elsevier: Amsterdam, The Netherlands, 2019; pp. 23–54. [Google Scholar]
- Toutain, P.-L.; Bousquet-Mélou, A. Bioavailability and its assessment. J. Vet. Pharmacol. Ther. 2004, 27, 455–466. [Google Scholar] [CrossRef] [PubMed]
- Lavefve, L.; Howard, L.R.; Carbonero, F. Berry polyphenols metabolism and impact on human gut microbiota and health. Food Funct. 2020, 11, 45–65. [Google Scholar] [CrossRef] [PubMed]
- Oracz, J.; Nebesny, E.; Zyzelewicz, D.; Budryn, G.; Luzak, B. Bioavailability and metabolism of selected cocoa bioactive compounds: A comprehensive review. Crit. Rev. Food Sci. Nutr. 2020, 60, 1947–1985. [Google Scholar] [CrossRef] [PubMed]
- Holst, B.; Williamson, G. Nutrients and phytochemicals: From bioavailability to bioefficacy beyond antioxidants. Curr. Opin. Biotechnol. 2008, 19, 73–82. [Google Scholar] [CrossRef] [PubMed]
- Wasilewski, A.; Zielińska, M.; Storr, M.; Fichna, J. Beneficial effects of probiotics, prebiotics, synbiotics, and psychobiotics in inflammatory bowel disease. Inflamm. Bowel Dis. 2015, 21, 1674–1682. [Google Scholar] [CrossRef] [PubMed]
- Hill, C.; Guarner, F.; Reid, G.; Gibson, G.R.; Merenstein, D.J.; Pot, B.; Morelli, L.; Canani, R.B.; Flint, H.J.; Salminen, S.; et al. Expert consensus document. The International Scientific Association for Probiotics and Prebiotics consensus statement on the scope and appropriate use of the term probiotic. Nat. Rev. Gastroenterol. 2014, 11, 506–514. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hossain, M.N.; Ranadheera, C.S.; Fang, Z.; Ajlouni, S. Healthy chocolate enriched with probiotics: A review. Food Sci. Technol. 2021, 41, 531–543. [Google Scholar] [CrossRef]
- Camps-Bossacoma, M.; Pérez-Cano, F.J.; Franch, À.; Castell, M. Gut microbiota in a rat oral sensitization model: Effect of a cocoa-enriched diet. Oxid. Med. Cell. Longev. 2017, 2017, 7417505. [Google Scholar] [CrossRef] [PubMed]
- Cueva, C.; Silva, M.; Pinillos, I.; Bartolomé, B.; Moreno-Arribas, M. Interplay between Dietary Polyphenols and Oral and Gut Microbiota in the Development of Colorectal Cancer. Nutrients 2020, 12, 625. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sorrenti, V.; Ali, S.; Mancin, L.; Davinelli, S.; Paoli, A.; Scapagnini, G. Cocoa Polyphenols and Gut Microbiota Interplay: Bioavailability, Prebiotic Effect, and Impact on Human Health. Nutrients 2020, 12, 1908. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Aweya, J.J.; Huang, Z.X.; Kang, Z.Y.; Bai, Z.H.; Li, K.H.; He, X.T.; Liu, Y.; Chen, X.Q.; Cheong, K.L. In-Vitro fermentation of Gracilaria lemaneiformis sulfated polysaccharides and its agaro-oligosaccharides by human fecal inocula and its impact on microbiota. Carbohydr. Polym. 2020, 234, 115894. [Google Scholar] [CrossRef] [PubMed]
- Dala-Paula, B.M.; Deus, V.L.; Tavano, O.L.; Gloria, M.B.A. In-Vitro bioaccessibility of amino acids and bioactive amines in 70% cocoa dark chocolate: What you eat and what you get. Food Chem. 2021, 343, 128397. [Google Scholar] [CrossRef] [PubMed]
- Soto-Martin, E.C.; Warnke, I.; Farquharson, F.M.; Christodoulou, M.; Horgan, G.; Derrien, M.; Faurie, J.E.; Flint, H.J.; Duncan, S.H.; Louis, P. Vitamin Biosynthesis by Human Gut Butyrate-Producing Bacteria and Cross-Feeding in Synthetic Microbial Communities. Mbio 2020, 11, e00886-20. [Google Scholar] [CrossRef] [PubMed]
- Hossain, M.N.; Ranadheera, C.S.; Fang, Z.; Ajlouni, S. Impact of encapsulating probiotics with cocoa powder on the viability of probiotics during chocolate processing, storage, and in-vitro gastrointestinal digestion. J. Food Sci. 2021, 86, 1629–1641. [Google Scholar] [CrossRef] [PubMed]
- Minekus, M.; Alminger, M.; Alvito, P.; Ballance, S.; Bohn, T.; Bourlieu, C.; Carriere, F.; Boutrou, R.; Corredig, M.; Dupont, D.; et al. A standardised static In-Vitro digestion method suitable for food—An international consensus. Food Funct. 2014, 5, 1113–1124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tzounis, X.; Vulevic, J.; Kuhnle, G.G.; George, T.; Leonczak, J.; Gibson, G.R.; Kwik-Uribe, C.; Spencer, J.P. Flavanol monomer-induced changes to the human faecal microflora. Br. J. Nutr. 2008, 99, 782–792. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cooper, K.A.; Campos-Giménez, E.; Jiménez Alvarez, D.; Nagy, K.; Donovan, J.L.; Williamson, G. Rapid reversed phase ultra-performance liquid chromatography analysis of the major cocoa polyphenols and inter-relationships of their concentrations in chocolate. J. Agric. Food Chem. 2007, 55, 2841–2847. [Google Scholar] [CrossRef]
- Sirisena, S.; Zabaras, D.; Ng, K.; Ajlouni, S. Characterization of Date (Deglet Nour) Seed Free and Bound Polyphenols by High-Performance Liquid Chromatography-Mass Spectrometry. J. Food Sci. 2017, 82, 333–340. [Google Scholar] [CrossRef]
- Sirisena, S.; Ajlouni, S.; Ng, K. Simulated gastrointestinal digestion and in vitro colonic fermentation of date (Phoenix dactylifera L.) seed polyphenols. Int. J. Food Sci. Technol. 2018, 53, 412–422. [Google Scholar] [CrossRef]
- Katz, D.L.; Doughty, K.; Ali, A. Cocoa and chocolate in human health and disease. Antioxid. Redox Signal. 2011, 15, 2779–2811. [Google Scholar] [CrossRef] [Green Version]
- Barisic, V.; Stokanovic, M.C.; Flanjak, I.; Doko, K.; Jozinovic, A.; Babic, J.; Subaric, D.; Milicevic, B.; Cindric, I.; Ackar, D. Cocoa Shell as a Step Forward to Functional Chocolates-Bioactive Components in Chocolates with Different Composition. Molecules 2020, 25, 5470. [Google Scholar] [CrossRef]
- Magrone, T.; Russo, M.A.; Jirillo, E. Cocoa and dark chocolate polyphenols: From biology to clinical applications. Front. Immunol. 2017, 8, 677. [Google Scholar] [CrossRef] [Green Version]
- Todorovic, V.; Redovnikovic, I.R.; Todorovic, Z.; Jankovic, G.; Dodevska, M.; Sobajic, S. Polyphenols, methylxanthines, and antioxidant capacity of chocolates produced in Serbia. J. Food Compos. Anal. 2015, 41, 137–143. [Google Scholar] [CrossRef]
- Urbańska, B.; Kowalska, J. Comparison of the Total Polyphenol Content and Antioxidant Activity of Chocolate Obtained from Roasted and Unroasted Cocoa Beans from Different Regions of the World. Antioxidants 2019, 8, 283. [Google Scholar] [CrossRef] [Green Version]
- Gomez-Juaristi, M.; Sarria, B.; Martinez-Lopez, S.; Bravo Clemente, L.; Mateos, R. Flavanol Bioavailability in Two Cocoa Products with Different Phenolic Content. A Comparative Study in Humans. Nutrients 2019, 11, 1441. [Google Scholar] [CrossRef] [Green Version]
- Montagnana, M.; Danese, E.; Angelino, D.; Mena, P.; Rosi, A.; Benati, M.; Gelati, M.; Salvagno, G.L.; Favaloro, E.J.; Del Rio, D.; et al. Dark chocolate modulates platelet function with a mechanism mediated by flavan-3-ol metabolites. Medicine 2018, 97, e13432. [Google Scholar] [CrossRef]
- Cantele, C.; Rojo-Poveda, O.; Bertolino, M.; Ghirardello, D.; Cardenia, V.; Barbosa-Pereira, L.; Zeppa, G. In Vitro Bioaccessibility and Functional Properties of Phenolic Compounds from Enriched Beverages Based on Cocoa Bean Shell. Foods 2020, 9, 715. [Google Scholar] [CrossRef]
- Gonzalez-Barrio, R.; Nunez-Gomez, V.; Cienfuegos-Jovellanos, E.; Garcia-Alonso, F.J.; Periago-Caston, M.J. Improvement of the Flavanol Profile and the Antioxidant Capacity of Chocolate Using a Phenolic Rich Cocoa Powder. Foods 2020, 9, 189. [Google Scholar] [CrossRef] [Green Version]
- Michael, K.; Papagiannopoulos, M.; Galensa, R. (-)-Catechin in Cocoa and Chocolate: Occurence and Analysis of an Atypical Flavan-3-ol Enantiomer. Molecules 2007, 12, 1274–1288. [Google Scholar]
- Gottumukkala, R.V.; Nadimpalli, N.; Sukala, K.; Subbaraju, G.V. Determination of Catechin and Epicatechin Content in Chocolates by High-Performance Liquid Chromatography. Int. Sch. Res. Not. 2014, 2014, 628196. [Google Scholar] [CrossRef] [Green Version]
- Ma, G.; Chen, Y. Polyphenol supplementation benefits human health via gut microbiota: A systematic review via meta-analysis. J. Funct. Foods 2020, 66, 103829. [Google Scholar] [CrossRef]
- Yang, Q.; Liang, Q.; Balakrishnan, B.; Belobrajdic, D.P.; Feng, Q.J.; Zhang, W. Role of dietary nutrients in the modulation of gut microbiota: A narrative review. Nutrients 2020, 12, 381. [Google Scholar] [CrossRef] [Green Version]
- Konar, N.; Palabiyik, I.; Toker, Ö.S.; Saputro, A.D.; Pirouzian, H.R. Improving Functionality of Chocolate. In Trends in Sustainable Chocolate Production; Springer Nature: Cham, Switzerland, 2022; pp. 75–112. [Google Scholar]
- Faccinetto-Beltran, P.; Gomez-Fernandez, A.R.; Santacruz, A.; Jacobo-Velazquez, D.A. Chocolate as Carrier to Deliver Bioactive Ingredients: Current Advances and Future Perspectives. Foods 2021, 10, 2065. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
| Flavonoids, µg/g | |||||||
|---|---|---|---|---|---|---|---|
| Probiotic-Chocolates | Digestion Stage | EC * | C | Pro B1 | Pro B2 | QC Gal | QC Glu |
| LGG *** | Initial | 298.66 ± 5.2 a **** | 115.66 ± 1.56 a | 387.86 ± 1.05 a | 793.0 ± 5.52 a | 644.33 ± 4.55 a | 587.0 ± 3.93 a |
| GD | 1830.0 ± 1.59 b | 82.0 ± 1.22 b | 313.5 ± 1.60 b | 636.33 ± 5.07 b | 631.67 ± 3.50 a | 582.0 ± 1.28 a | |
| CF | 87.33 ± 2.16 c | 26.0 ± 1.3 c | 68.7 ± 0.12 c | 146.33 ± 1.33 c | nd ** | nd | |
| Lc | Initial | 296.36 ± 4.12 a | 121.16 ± 2.16 a | 382.18 ± 2.23 a | 798.13 ± 3.22 a | 649.13 ± 2.51 a | 586.06 ± 4.06 a |
| GD | 194.66 ± 2.06 b | 73.33 ± 1.08 b | 321.33 ± 2.92 b | 704.24 ± 1.68 b | 623.37 ± 2.59 a | 581.33 ± 4.81 a | |
| CF | 96.33 ± 2.82 c | 32.0 ± 2.0 c | 58.3 ± 0.2 c | 68.0 ± 2.24 c | nd | nd | |
| La5 | Initial | 299.61 ± 1.27 a | 117.34 ± 1.06 a | 388.46 ± 1.15 a | 794.24 ± 2.52 a | 644.87 ± 3.76 a | 589.52 ± 6.09 a |
| GD | 184.67 ± 1.58 b | 77.0 ± 0.66 b | 291.33 ± 3.56 b | 652.0 ± 4.29 b | 627.33 ± 6.53 a | 571.33 ± 3.24 a | |
| CF | 97.0 ± 2.13 c | 34.67 ± 0.05 c | 60.83 ± 2.19 c | 103.67 ± 2.03 c | nd | nd | |
| Lp | Initial | 298.49 ± 7.09 a | 115.27 ± 2.09 a | 390.66 ± 2.05 a | 793.56 ± 3.98 a | 643.45 ± 1.79 a | 582.74 ± 7.81 a |
| GD | 178.25 ± 2.15 b | 69.0 ± 1.0 b | 277.7 ± 3.79 b | 553.33 ± 2.24 b | 623.67 ± 4.86 a | 574.67 ± 7.0 a | |
| CF | 90.0 ± 0.71 c | 43.67 ± 1.5 c | 102.63 ± 2.58 c | 191.33 ± 4.7 c | nd | nd | |
| Ls | Initial | 301.24 ± 6.02 a | 118.57 ± 1.02 a | 387.23 ± 1.67 a | 797.21 ± 3.05 a | 647.87 ± 3.03 a | 589.26 ± 6.41 a |
| GD | 169.0 ± 2.80 b | 72.33 ± 1.3 b | 271.03 ± 2.64 b | 702 ± 3.77 b | 637.67 ± 2.5 a | 576.04 ± 3.66 a | |
| CF | 113.33 ± 1.83 c | 39.67 ± 0.93 c | 102.2 ± 2.54 c | 81.66 ± 1.13 c | nd | nd | |
| BB12 | Initial | 298.52 ± 5.02 a | 114.89 ± 3.16 a | 389.56 ± 3.25 a | 793.83 ± 2.35 a | 644.73 ± 5.72 a | 587.49 ± 2.93 a |
| GD | 192.33 ± 4.77 b | 75.33 ± 1.8 b | 304.7 ± 1.83 b | 719.33 ± 3.71 b | 638.36 ± 2.18 a | 581.33 ± 4.89 a | |
| CF | 96.67 ± 0.51 c | 30 ± 1.52 c | 74.97 ± 1.67 c | 69.23 ± 3.78 c | nd | nd | |
| St | Initial | 293.26 ± 3.76 a | 119.52 ± 1.35 a | 388.11 ± 2.59 a | 791.53 ± 2.46 a | 645.74 ± 1.04 a | 588.55 ± 7.01 a |
| GD | 164.12 ± 3.07 b | 88 ± 1.47 b | 266.36 ± 2.87 b | 632.0 ± 3.53 b | 623.67 ± 4.04 a | 564.67 ± 3.71 a | |
| CF | 123.33 ± 2.02 c | 25.67 ± 0.06 c | 112.03 ± 2.40 c | 134.37 ± 3.13 c | nd | nd | |
| Flavonoids, µg/g | |||||||
|---|---|---|---|---|---|---|---|
| Probiotic-Chocolates | Digestion Stage | EC * | C | Pro B1 | Pro B2 | QC Gal | QC Glu |
| LGG *** | Initial | 187.0 ± 1.2 a **** | 75.62 ± 1.42 a | 257.13 ± 2.05 a | 518.33 ± 1.52 a | 410.0 ± 3.55 a | 379.0 ± 5.93 a |
| GD | 110.0 ± 1.6 b | 60.6 ± 2.81 b | 198.67 ± 3.11 b | 458 ± 9.07 b | 397.63 ± 3.50 a | 364.0 ± 1.28 a | |
| CF | 70.33 ± 0.55 c | 13.93 ± 0.11 c | 57.7 ± 1.12 c | 61.13 ± 1.33 c | nd ** | nd | |
| Lc | Initial | 182.67 ± 3.09 a | 75.12 ± 1.02 a | 263.27 ± 2.85 a | 528.61 ± 7.52 a | 405.27 ± 7.09 a | 372.54 ± 5.23 a |
| GD | 122.66 ± 1.09 b | 63.67 ± 0.08 b | 208.0 ± 2.08 b | 464.66 ± 4.68 b | 373.37 ± 2.59 a | 357.33 ± 4.81 a | |
| CF | 62.0 ± 0.80 c | 8.67 ± 0.3 c | 53.3 ± 0.28 c | 48.0 ± 2.24 c | nd | nd | |
| La5 | Initial | 187.66 ± 3.98 a | 73.52 ± 2.26 a | 258.21 ± 3.05 a | 513.41 ± 1.92 a | 410.57 ± 3.05 a | 380.13 ± 8.07 a |
| GD | 114.33 ± 2.48 b | 62.16 ± 0.57 b | 200.66 ± 2.56 b | 409 ± 6.08 b | 401.24 ± 1.53 a | 370.27 ± 3.24 a | |
| CF | 71.0 ± 2.51 c | 11.0 ± 1.7 c | 50.83 ± 0.19 c | 101.1 ± 2.03 c | nd | nd | |
| Lp | Initial | 189.23 ± 2.03 a | 77.62 ± 1.15 a | 257.73 ± 5.82 a | 518.97 ± 6.02 a | 415.03 ± 5.87 a | 377.19 ± 3.78 a |
| GD | 111.67 ± 2.68 b | 60.33 ± 1.0 b | 179.40 ± 1.79 b | 386.33 ± 3.72 b | 398.67 ± 4.86 a | 374.67 ± 2.0 a | |
| CF | 72.33 ± 2.06 c | 12.56 ± 2.5 c | 72.43 ± 1.58 c | 121.33 ± 4.7 c | nd | nd | |
| Ls | Initial | 190.12 ± 4.08 a | 73.44 ± 3.42 a | 255.55 ± 4.09 a | 523.74 ± 6.97 a | 409.44 ± 2.07 a | 379.47 ± 6.09 a |
| GD | 117.0 ± 2.51 b | 64.33 ± 1.3 b | 201.03 ± 2.94 b | 447.67 ± 5.22 b | 387.67 ± 8.5 a | 366.04 ± 1.66 a | |
| CF | 59.33 ± 1.56 c | 8.23 ± 1.7 c | 43.2 ± 1.54 c | 61.66 ± 3.13 c | nd | nd | |
| BB12 | Initial | 186.87 ± 1.02 a | 78.23 ± 1.56 a | 258.35 ± 5.27 a | 522.36 ± 3.52 a | 412.67 ± 7.28 a | 378.93 ± 4.34 a |
| GD | 119.66 ± 1.6 b | 67.67 ± 1.6 b | 208.46 ± 4.83 b | 452.33 ± 5.05 b | 378.36 ± 2.18 a | 357.33 ± 4.89 a | |
| CF | 65.33 ± 3.8 c | 6.4 ± 1.73 c | 44.97 ± 2.67 c | 59.23 ± 3.78 c | nd | nd | |
| St | Initial | 187.74 ± 2.12 a | 76.56 ± 3.78 a | 264.70 ± 4.85 a | 518.43 ± 6.28 a | 410.71 ± 4.05 a | 381.33 ± 5.14 a |
| GD | 121.66 ± 2.21 b | 66 ± 0.06 b | 161.16 ± 1.28 b | 364.67 ± 1.53 b | 403.67 ± 6.04 a | 361.61 ± 2.83 a | |
| CF | 61.67 ± 2.02 c | 8.66 ± 0.09 c | 83.8 ± 1.40 c | 142.0 ± 3.13 c | nd | nd | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hossain, M.N.; Ranadheera, C.S.; Fang, Z.; Ajlouni, S. Interaction between Chocolate Polyphenols and Encapsulated Probiotics during In Vitro Digestion and Colonic Fermentation. Fermentation 2022, 8, 253. https://doi.org/10.3390/fermentation8060253
Hossain MN, Ranadheera CS, Fang Z, Ajlouni S. Interaction between Chocolate Polyphenols and Encapsulated Probiotics during In Vitro Digestion and Colonic Fermentation. Fermentation. 2022; 8(6):253. https://doi.org/10.3390/fermentation8060253
Chicago/Turabian StyleHossain, Md Nur, Chaminda Senaka Ranadheera, Zhongxiang Fang, and Said Ajlouni. 2022. "Interaction between Chocolate Polyphenols and Encapsulated Probiotics during In Vitro Digestion and Colonic Fermentation" Fermentation 8, no. 6: 253. https://doi.org/10.3390/fermentation8060253
APA StyleHossain, M. N., Ranadheera, C. S., Fang, Z., & Ajlouni, S. (2022). Interaction between Chocolate Polyphenols and Encapsulated Probiotics during In Vitro Digestion and Colonic Fermentation. Fermentation, 8(6), 253. https://doi.org/10.3390/fermentation8060253