
Starmerella bacillaris Strains Used in Sequential Alcoholic Fermentation with Saccharomyces cerevisiae Improves Protein Stability in White Wines

 ,
, 
 , ,
, ,  ,
,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Yeast Strains
2.2. Fermentation Trials
2.3. Chemical Analysis
2.4. Protein Stability of White Wines
2.5. Quantification of Grape Proteins
2.6. Quantification of Total Polysaccharides by HRSEC-RID
2.7. Mannoprotein Quantification
2.8. Statistical Analysis
3. Results and Discussion
3.1. Sequential Fermentation Performance in Three Grape Juices
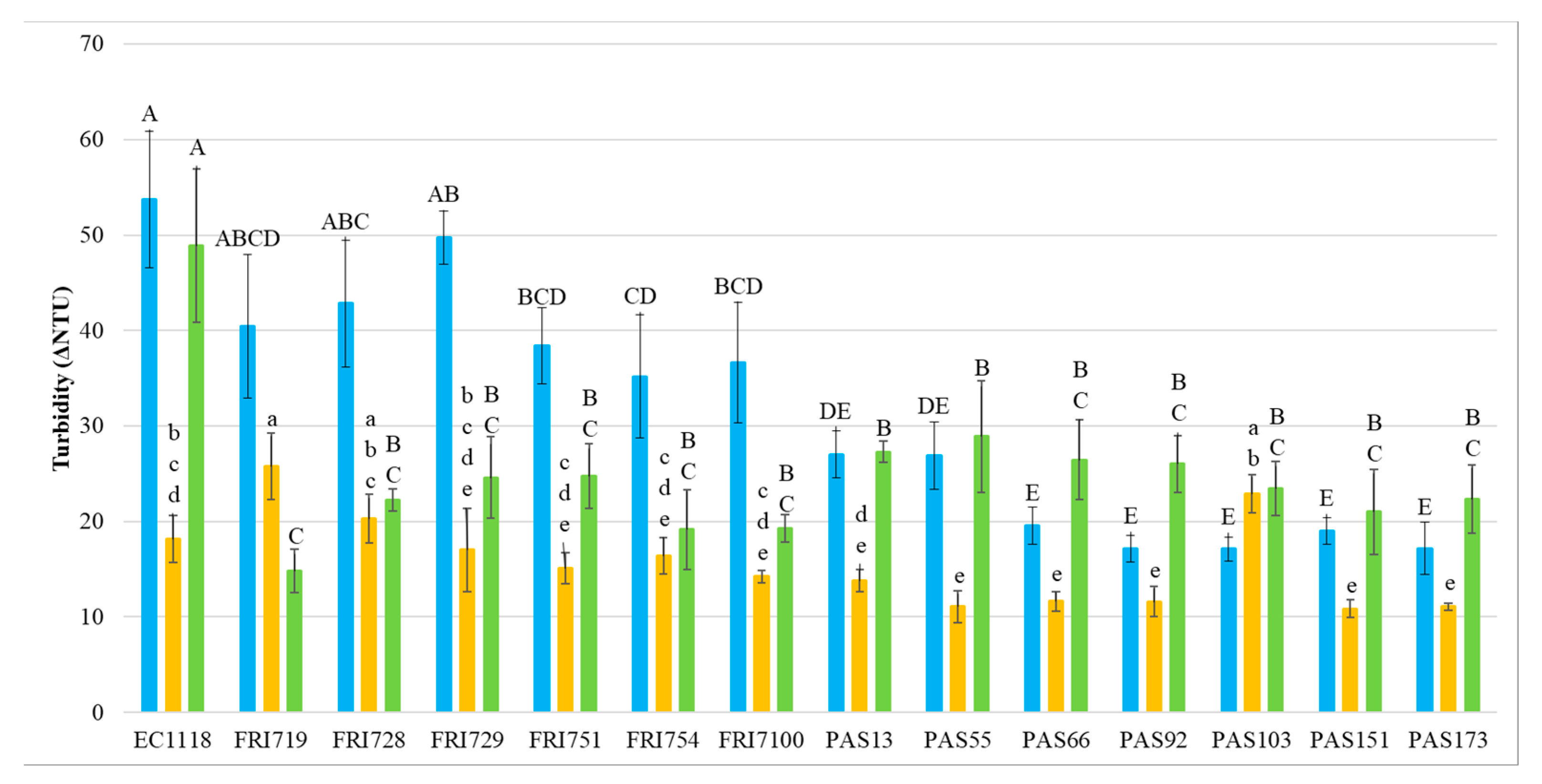
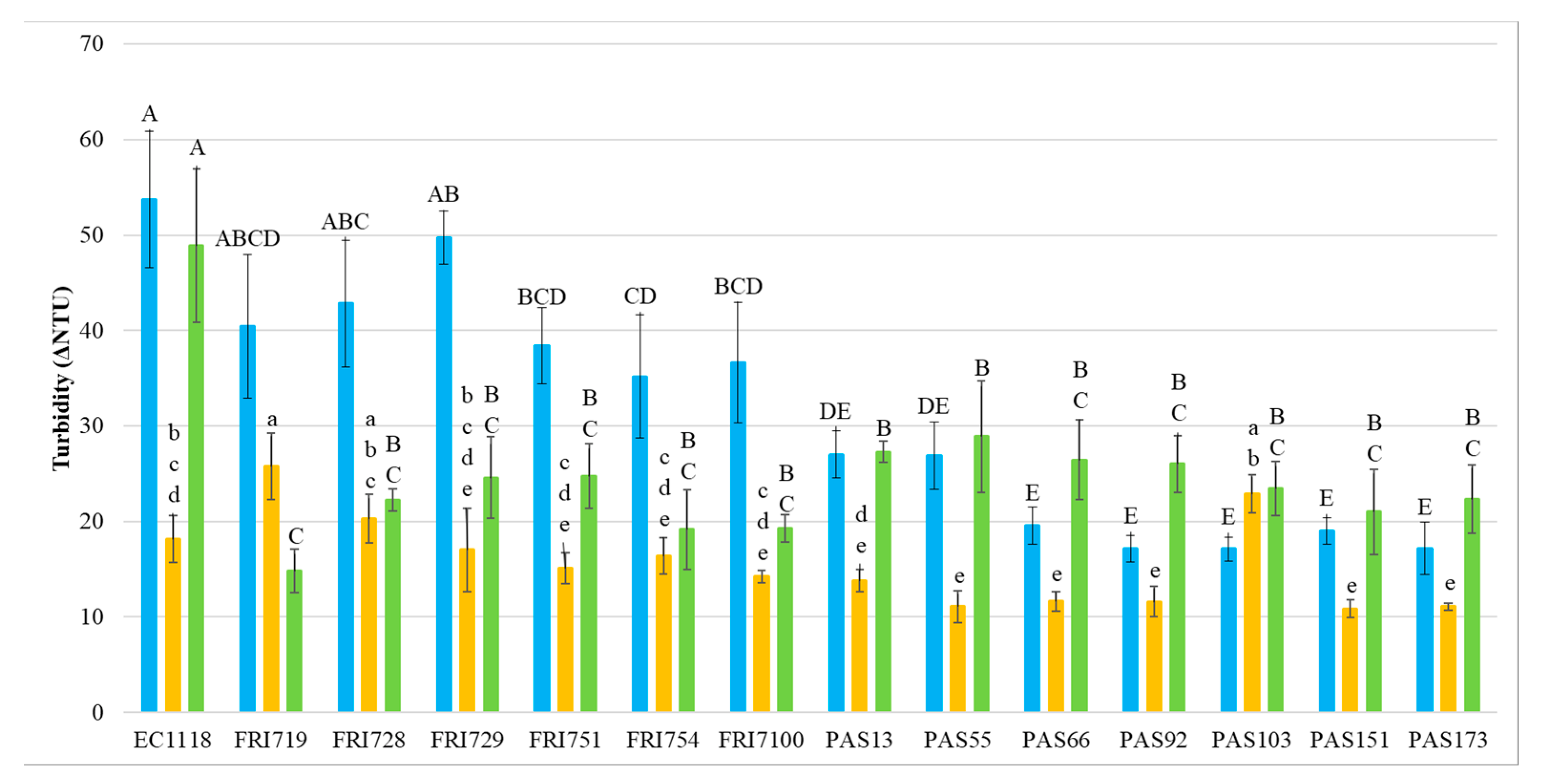
3.2. Wine Protein Stability
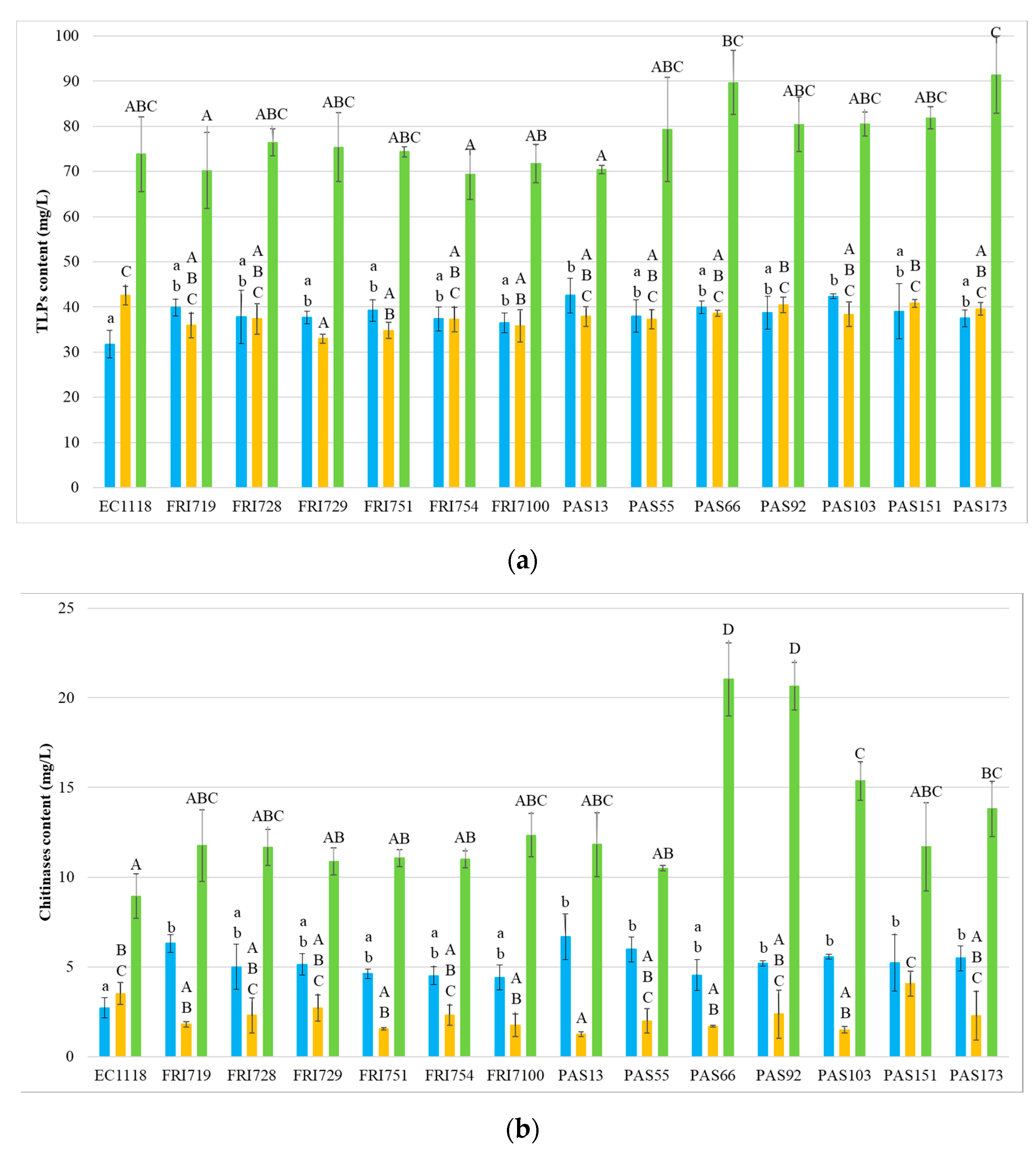
3.3. Wine Protein Profiles
3.4. Wine Polysaccharides
3.5. Mannoproteins Quantification
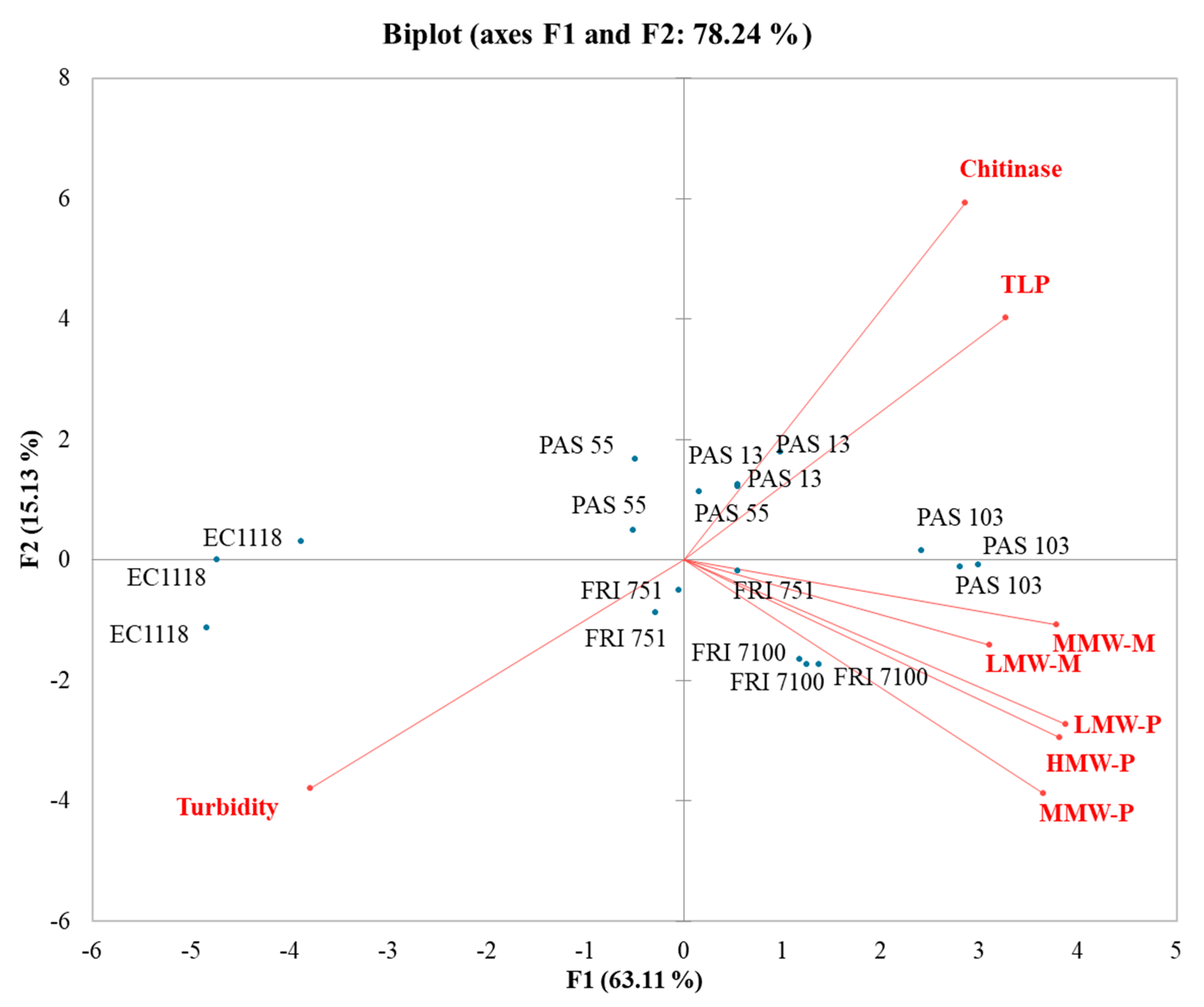
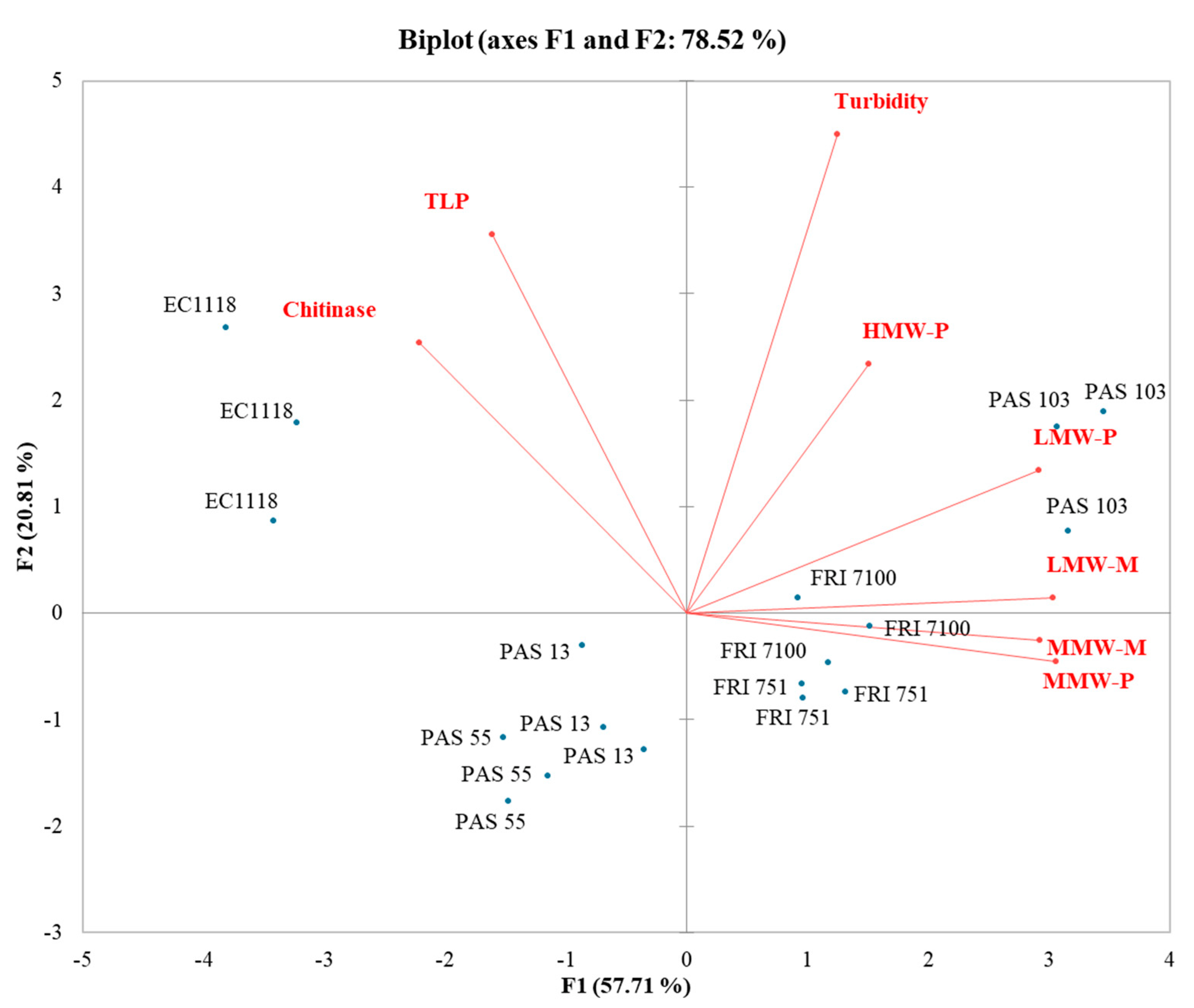
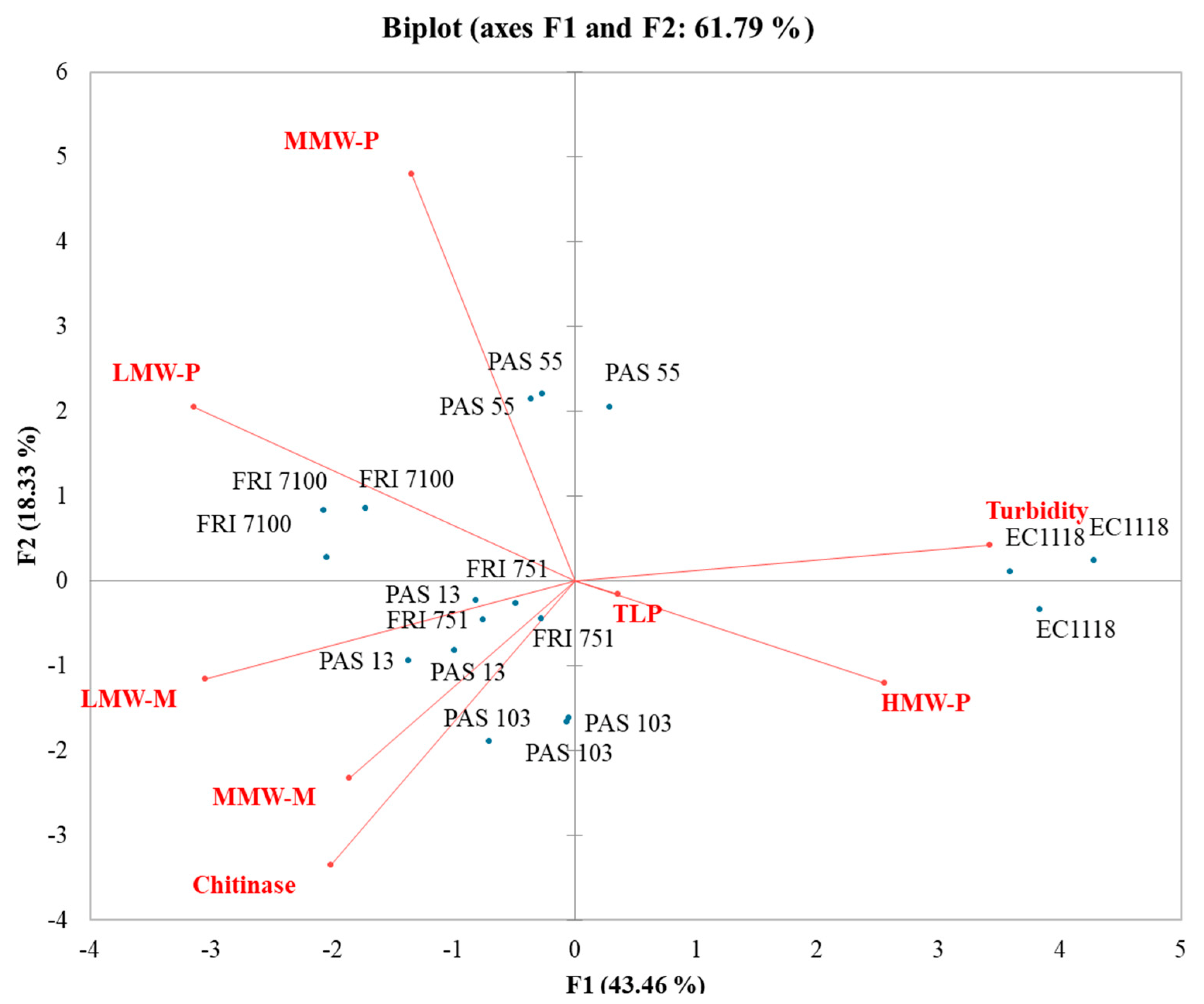
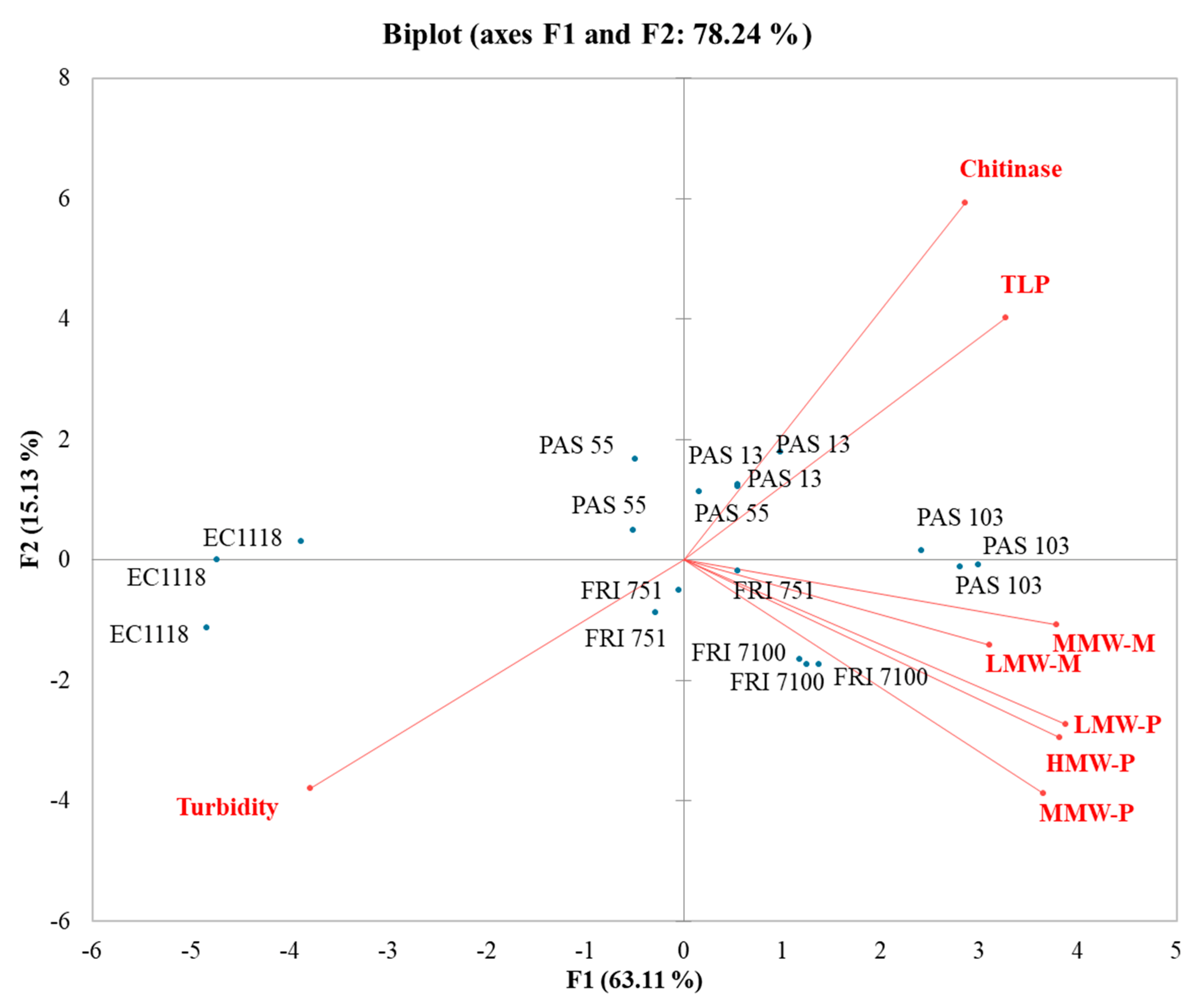
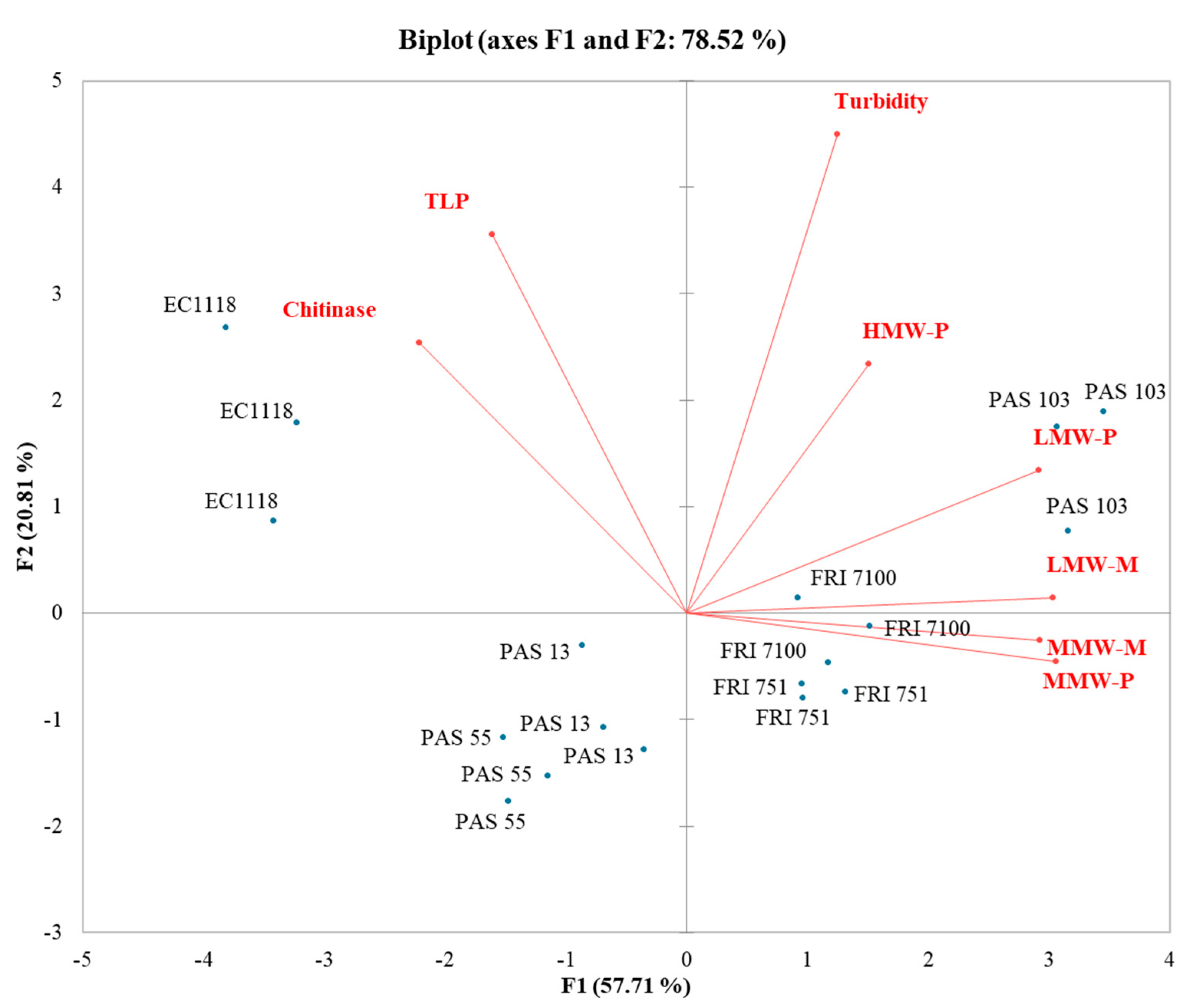
3.6. Influence of Polysaccharides and Mannoproteins on Wines Stability
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Acknowledgments
Conflicts of Interest
References
- Cosme, F.; Filipe-Ribeiro, L.; Nunes, F.M. Wine stabilisation: An overview of defects and treatments. In Chemistry and Biochemistry of Winemaking, Wine Stabilization and Aging; Cosme, F., Nunes, F.M., Filipe-Ribeiro, L., Eds.; IntechOpen: London, UK, 2021. [Google Scholar] [CrossRef]
- Van Sluyter, S.C.; McRae, J.M.; Falconer, R.J.; Smith, P.A.; Bacic, A.; Waters, E.J.; Marangon, M. Wine protein haze: Mechanisms of formation and advances in prevention. J. Agric. Food Chem. 2015, 63, 4020–4030. [Google Scholar] [CrossRef] [PubMed]
- Falconer, R.J.; Marangon, M.; Van Sluyter, S.C.; Neilson, K.A.; Chan, C.; Waters, E.J. Thermal stability of thaumatin-like protein, chitinase, and invertase isolated from Sauvignon blanc and Semillon juice and their role in haze formation in wine. J. Agric. Food Chem. 2010, 58, 975–980. [Google Scholar] [CrossRef] [PubMed]
- Marangon, M.; Sluyter, S.C.V.; Waters, E.J.; Menz, R.I. Structure of haze forming proteins in white wines: Vitis vinifera thaumatin-like proteins. PLoS ONE 2014, 9, e113757. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marangon, M.; Sauvage, F.-X.; Waters, E.J.; Vernhet, A. Effects of ionic strength and sulfate upon thermal aggregation of grape chitinases and thaumatin-like proteins in a model system. J. Agric. Food Chem. 2011, 59, 2652–2662. [Google Scholar] [CrossRef]
- Marangon, M.; Van Sluyter, S.C.; Neilson, K.A.; Chan, C.; Haynes, P.A.; Waters, E.J.; Falconer, R.J. Roles of grape thaumatin-like protein and chitinase in white wine haze formation. J. Agric. Food Chem. 2011, 59, 733–740. [Google Scholar] [CrossRef]
- Ruzza, P.; Honisch, C.; Marangon, M.; Curioni, A.; Bakalinsky, A.; Vincenzi, S. Influence of the reducing environment in the misfolding of wine proteins. Adv. Protein Chem. Struct. Biol. 2019, 118, 413–436. [Google Scholar] [CrossRef]
- Vernhet, A.; Meistermann, E.; Cottereau, P.; Charrier, F.; Chemardin, P.; Poncet-Legrand, C. Wine thermosensitive proteins adsorb first and better on bentonite during fining: Practical implications and proposition of alternative heat tests. J. Agric. Food Chem. 2020, 68, 13450–13458. [Google Scholar] [CrossRef]
- Di Gaspero, M.; Ruzza, P.; Hussain, R.; Honisch, C.; Biondi, B.; Siligardi, G.; Marangon, M.; Curioni, A.; Vincenzi, S. The secondary structure of a major wine protein is modified upon interaction with polyphenols. Molecules 2020, 25, 1646. [Google Scholar] [CrossRef] [Green Version]
- Dordoni, R.; Colangelo, D.; Giribaldi, M.; Giuffrida, M.G.; De Faveri, D.M.; Lambri, M. Effect of bentonite characteristics on wine proteins, polyphenols, and metals under conditions of different pH. Am. J. Enol. Vitic. 2015, 66, 518–530. [Google Scholar] [CrossRef]
- Arenas, I.; Ribeiro, M.; Filipe-Ribeiro, L.; Vilamarim, R.; Costa, E.; Siopa, J.; Cosme, F.; Nunes, F.M. Effect of pre-fermentative maceration and fining agents on protein stability, macromolecular, and phenolic composition of Albariño white wines: Comparative efficiency of chitosan, k-carrageenan and bentonite as heat stabilisers. Foods 2021, 10, 608. [Google Scholar] [CrossRef]
- Marangon, M.; Lucchetta, M.; Duan, D.; Stockdale, V.J.; Hart, A.; Rogers, P.J.; Waters, E.J. Protein removal from a Chardonnay juice by addition of carrageenan and pectin: Adsorbents for protein stabilization of wines. Aust. J. Grape Wine Res. 2012, 18, 194–202. [Google Scholar] [CrossRef]
- Ratnayake, S.; Stockdale, V.; Grafton, S.; Munro, P.; Robinson, A.L.; Pearson, W.; McRae, J.M.; Bacic, A. Carrageenans as heat stabilisers of white wine. Aust. J. Grape Wine Res. 2019, 25, 439–450. [Google Scholar] [CrossRef]
- Romanini, E.; McRae, J.M.; Bilogrevic, E.; Colangelo, D.; Gabrielli, M.; Lambri, M. Use of grape seeds to reduce haze formation in white wines. Food Chem. 2021, 341, 128250. [Google Scholar] [CrossRef]
- Romanini, E.; McRae, J.M.; Colangelo, D.; Lambri, M. First trials to assess the feasibility of grape seed powder (GSP) as a novel and sustainable bentonite alternative. Food Chem. 2020, 305, 125484. [Google Scholar] [CrossRef]
- Marangon, M.; Van Sluyter, S.C.; Robinson, E.M.C.; Muhlack, R.A.; Holt, H.E.; Haynes, P.A.; Godden, P.W.; Smith, P.A.; Waters, E.J. Degradation of white wine haze proteins by Aspergillopepsin I and II during juice flash pasteurization. Food Chem. 2012, 135, 1157–1165. [Google Scholar] [CrossRef]
- Sui, Y.; McRae, J.M.; Wollan, D.; Muhlack, R.A.; Godden, P.; Wilkinson, K.L. Use of ultrafiltration and proteolytic enzymes as alternative approaches for protein stabilisation of white wine. Aust. J. Grape Wine Res. 2021, 27, 234–245. [Google Scholar] [CrossRef]
- Brown, S.L.; Stockdale, V.J.; Pettolino, F.; Pocock, K.F.; de Barros Lopes, M.; Williams, P.J.; Bacic, A.; Fincher, G.B.; Høj, P.B.; Waters, E.J. Reducing haziness in white wine by overexpression of Saccharomyces cerevisiae genes YOL155c and YDR055w. Appl. Microbiol. Biotechnol. 2007, 73, 1363–1376. [Google Scholar] [CrossRef]
- Dupin, I.V.S.; Stockdale, V.J.; Williams, P.J.; Jones, G.P.; Markides, A.J.; Waters, E.J. Saccharomycescerevisiae mannoproteins that protect wine from protein haze: Evaluation of extraction methods and immunolocalization. J. Agric. Food Chem. 2000, 48, 1086–1095. [Google Scholar] [CrossRef]
- Llaubères, R.M.; Dubourdieu, D.; Villettaz, J.C. Exocellular polysaccharides from Saccharomyces in Wine. J. Sci. Food Agric. 1987, 41, 277–286. [Google Scholar] [CrossRef]
- Charpentier, C.; Dos Santos, A.M.; Feuillat, M. Release of macromolecules by Saccharomyces cerevisiae during ageing of French flor sherry wine “Vin jaune”. Int. J. Food Microbiol. 2004, 96, 253–262. [Google Scholar] [CrossRef]
- FAO. Yeast Extracts Containing Mannoproteins. 2021. Available online: https://www.fao.org/3/cb3376en/cb3376en.pdf (accessed on 11 January 2022).
- Klis, F.M.; Boorsma, A.; De Groot, P.W.J. Cell wall construction in Saccharomyces cerevisiae. Yeast 2006, 23, 185–202. [Google Scholar] [CrossRef]
- Millarini, V.; Ignesti, S.; Cappelli, S.; Ferraro, G.; Adessi, A.; Zanoni, B.; Fratini, E.; Domizio, P. Protection of wine from protein haze using Schizosaccharomyces japonicus polysaccharides. Foods 2020, 9, 1407. [Google Scholar] [CrossRef]
- Vejarano, R. Non-Saccharomyces in winemaking: Source of mannoproteins, nitrogen, enzymes, and antimicrobial compounds. Fermentation 2020, 6, 76. [Google Scholar] [CrossRef]
- Giovani, G.; Rosi, I.; Bertuccioli, M. Quantification and characterization of cell wall polysaccharides released by non-Saccharomyces yeast strains during alcoholic fermentation. Int. J. Food Microbiol. 2012, 160, 113–118. [Google Scholar] [CrossRef]
- Romani, C.; Domizio, P.; Lencioni, L.; Gobbi, M.; Comitini, F.; Ciani, M.; Mannazzu, I. Polysaccharides and glycerol production by non-Saccharomyces wine yeasts in mixed fermentation. Quad. Vitic. Enol. Univ. Torino 2010, 31, 185–189. [Google Scholar]
- Goold, H.D.; Kroukamp, H.; Williams, T.C.; Paulsen, I.T.; Varela, C.; Pretorius, I.S. Yeast’s balancing act between ethanol and glycerol production in low-alcohol wines. Microb. Biotechnol. 2017, 10, 264–278. [Google Scholar] [CrossRef]
- Lemos Junior, W.J.F.; de Oliveira, V.S.; Guerra, A.F.; Giacomini, A.; Corich, V. From the vineyard to the cellar: New insights of Starmerella bacillaris (synonym Candida zemplinina) technological properties and genomic perspective. Appl. Microbiol. Biotechnol. 2021, 105, 493–501. [Google Scholar] [CrossRef]
- Lemos Junior, W.J.; Bovo, B.; Nadai, C.; Crosato, G.; Carlot, M.; Favaron, F.; Giacomini, A.; Corich, V. Biocontrol ability and action mechanism of Starmerella bacillaris (synonym Candida zemplinina) isolated from wine musts against gray mold disease agent Botrytis cinerea on grape and their effects on alcoholic fermentation. Front. Microbiol. 2016, 7, 1249. [Google Scholar] [CrossRef]
- Lemos Junior, W.J.F.; Treu, L.; Duarte, V.D.S.; Campanaro, S.; Nadai, C.; Giacomini, A.; Corich, V. Draft genome sequence of the yeast Starmerella bacillaris (syn. Candida zemplinina) FRI751 isolated from fermenting must of dried Raboso grapes. Genome Announc. 2017, 5, e00224–e00317. [Google Scholar] [CrossRef] [Green Version]
- Lemos Junior, W.J.F.; Treu, L.; da Silva Duarte, V.; Carlot, M.; Nadai, C.; Campanaro, S.; Giacomini, A.; Corich, V. Whole-genome sequence of Starmerella bacillaris PAS13, a non-conventional enological yeast with antifungal activity. Genome Announc. 2017, 5, e00788–e00817. [Google Scholar] [CrossRef] [Green Version]
- Bovo, B.; Nadai, C.; Vendramini, C.; Lemos Junior, W.J.; Carlot, M.; Skelin, A.; Giacomini, A.; Corich, V. Aptitude of Saccharomyces yeasts to ferment unripe grapes harvested during cluster thinning for reducing alcohol content of wine. Int. J. Food Microbiol. 2016, 236, 56–64. [Google Scholar] [CrossRef] [PubMed]
- Nadai, C.; Lemos Junior, W.J.; Favaron, F.; Giacomini, A.; Corich, V. Biocontrol activity of Starmerella bacillaris yeast against blue mold disease on apple fruit and its effect on cider fermentation. PLoS ONE 2018, 13, e0204350. [Google Scholar] [CrossRef] [PubMed]
- Lemos Junior, W.J.F.; Nadai, C.; Crepalde, L.T.; de Oliveira, V.S.; de Matos, A.D.; Giacomini, A.; Corich, V. Potential use of Starmerella bacillaris as fermentation starter for the production of low-alcohol beverages obtained from unripe grapes. Int. J. Food Microbiol. 2019, 303, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Pocock, K.F.; Rankine, B.C. Heat test for detecting protein instability in wine. Aust. Wine Brew. Spirit Rev. 1973, 91, 42–43. [Google Scholar]
- Van Sluyter, S.C.; Marangon, M.; Stranks, S.D.; Neilson, K.A.; Hayasaka, Y.; Haynes, P.A.; Ian Menz, R.; Waters, E.J. Two-Step purification of pathogenesis-related proteins from grape juice and crystallization of thaumatin-like proteins. J. Agric. Food Chem. 2009, 57, 11376–11382. [Google Scholar] [CrossRef]
- Ayestarán, B.; Guadalupe, Z.; León, D. Quantification of major grape polysaccharides (Tempranillo v.) released by maceration enzymes during the fermentation process. Anal. Chim. Acta 2004, 513, 29–39. [Google Scholar] [CrossRef]
- De Iseppi, A.; Marangon, M.; Vincenzi, S.; Lomolino, G.; Curioni, A.; Divol, B. A novel approach for the valorization of wine lees as a source of compounds able to modify wine properties. LWT 2021, 136, 110274. [Google Scholar] [CrossRef]
- Gobert, A.; Tourdot-Maréchal, R.; Morge, C.; Sparrow, C.; Liu, Y.; Quintanilla-Casas, B.; Vichi, S.; Alexandre, H. Non-Saccharomyces yeasts nitrogen source preferences: Impact on sequential fermentation and wine volatile compounds profile. Front. Microbiol. 2017, 8, 2175. [Google Scholar] [CrossRef] [Green Version]
- Nadai, C.; Giacomini, A.; Corich, V. The addition of wine yeast Starmerella bacillaris to grape skin surface influences must fermentation and glycerol production. OENO One 2021, 55, 47–55. [Google Scholar] [CrossRef]
- Horváth, B.O.; Sárdy, D.N.; Kellner, N.; Magyar, I. Effects of high sugar content on fermentation dynamics and some metabolites of wine-related yeast species Saccharomyces cerevisiae, S. uvarum and Starmerella bacillaris. Food Technol. Biotechnol. 2020, 58, 76–83. [Google Scholar] [CrossRef]
- Gawel, R.; Sluyter, S.V.; Waters, E.J. The effects of ethanol and glycerol on the body and other sensory characteristics of Riesling wines. Aust. J. Grape Wine Res. 2007, 13, 38–45. [Google Scholar] [CrossRef]
- Benito, S. The impacts of Schizosaccharomyces on winemaking. Appl. Microbiol. Biotechnol. 2019, 103, 4291–4312. [Google Scholar] [CrossRef]
- Lemos Junior, W.J.F.; Nadai, C.; Rolle, L.; Da Silva Gulao, E.; Miguez da Rocha Leãoe, M.H.; Giacomini, A.; Corich, V.; Vincenzi, S. Influence of the mannoproteins of different strains of Starmerella bacillaris used in single and sequential fermentations on foamability, tartaric and protein stabilities of wines. OENO One 2020, 54, 231–243. [Google Scholar] [CrossRef] [Green Version]
- Englezos, V.; Rantsiou, K.; Giacosa, S.; Río Segade, S.; Rolle, L.; Cocolin, L. Cell-to-cell contact mechanism modulates Starmerella bacillaris death in mixed culture fermentations with Saccharomyces cerevisiae. Int. J. Food Microbiol. 2019, 289, 106–114. [Google Scholar] [CrossRef]
- Englezos, V.; Rantsiou, K.; Torchio, F.; Rolle, L.; Gerbi, V.; Cocolin, L. Exploitation of the non-Saccharomyces yeast Starmerella bacillaris (synonym Candida zemplinina) in wine fermentation: Physiological and molecular characterizations. Int. J. Food Microbiol. 2015, 199, 33–40. [Google Scholar] [CrossRef]
- Comitini, F.; Gobbi, M.; Domizio, P.; Romani, C.; Lencioni, L.; Mannazzu, I.; Ciani, M. Selected non-Saccharomyces wine yeasts in controlled multistarter fermentations with Saccharomyces cerevisiae. Food Microbiol. 2011, 28, 873–882. [Google Scholar] [CrossRef]
- Englezos, V.; Rantsiou, K.; Cravero, F.; Torchio, F.; Ortiz-Julien, A.; Gerbi, V.; Rolle, L.; Cocolin, L. Starmerella bacillaris and Saccharomyces cerevisiae mixed fermentations to reduce ethanol content in wine. Appl. Microbiol. Biotechnol. 2016, 100, 5515–5526. [Google Scholar] [CrossRef]
- Rantsiou, K.; Dolci, P.; Giacosa, S.; Torchio, F.; Tofalo, R.; Torriani, S.; Suzzi, G.; Rolle, L.; Cocolin, L. Candida zemplinina can reduce acetic acid produced by Saccharomyces cerevisiae in sweet wine fermentations. Appl. Environ. Microbiol. 2012, 78, 1987–1994. [Google Scholar] [CrossRef] [Green Version]
- Lambrechts, M.G.; Pretorius, I.S. Yeast and its importance to wine aroma-a review. S. Afr. J. Enol. Vitic. 2000, 21, 97–129. [Google Scholar] [CrossRef] [Green Version]
- Mendes Ferreira, A.; Mendes-Faia, A. The role of yeasts and lactic acid bacteria on the metabolism of organic acids during winemaking. Foods 2020, 9, 1231. [Google Scholar] [CrossRef]
- Esteruelas, M.; Poinsaut, P.; Sieczkowski, N.; Manteau, S.; Fort, M.F.; Canals, J.M.; Zamora, F. Characterization of natural haze protein in Sauvignon white wine. Food Chem. 2009, 113, 28–35. [Google Scholar] [CrossRef]
- Vincenzi, S.; Mosconi, S.; Zoccatelli, G.; Pellegrina, C.D.; Veneri, G.; Chignola, R.; Peruffo, A.; Curioni, A.; Rizzi, C. Development of a new procedure for protein recovery and quantification in wine. Am. J. Enol. Vitic. 2005, 56, 182–187. [Google Scholar]
- Waters, E.J.; Shirley, N.J.; Williams, P.J. Nuisance proteins of wine are grape pathogenesis related proteins. J. Agric. Food Chem. 1996, 44, 3–5. [Google Scholar] [CrossRef]
- Vincenzi, S.; Marangon, M.; Tolin, S.; Curioni, A. Protein evolution during the early stages of white winemaking and its relations with wine stability: Protein evolution in white wine during winemaking. Aust. J. Grape Wine Res. 2011, 17, 20–27. [Google Scholar] [CrossRef]
- Ndlovu, T.; Divol, B.; Bauer, F.F. Yeast cell wall chitin reduces wine haze formation. Appl. Environ. Microbiol. 2018, 84, e00668–e00718. [Google Scholar] [CrossRef] [Green Version]
- Vidal, S.; Doco, T.; Moutounet, M.; Pellerin, P. Soluble polysaccharide content at initial time of experimental must preparation. Am. J. Enol. Vitic. 2000, 51, 115–121. [Google Scholar]
- González-Royo, E.; Esteruelas, M.; Kontoudakis, N.; Fort, F.; Canals, J.M.; Zamora, F. The effect of supplementation with three commercial inactive dry yeasts on the colour, phenolic compounds, polysaccharides and astringency of a model wine solution and red wine. J. Sci. Food Agric. 2017, 97, 172–181. [Google Scholar] [CrossRef]
- Waters, E.J.; Pellerin, P.; Brillouet, J.M. A wine arabinogalactan-protein that reduces heat-induced wine protein haze. Biosci. Biotechnol. Biochem. 1994, 58, 43–48. [Google Scholar] [CrossRef] [Green Version]
- Moine-Ledoux, V.; Dubourdieu, D. An invertase fragment responsible for improving the protein stability of dry white wines. J. Sci. Food Agric. 1999, 79, 537–543. [Google Scholar] [CrossRef]
- Waters, E.J.; Alexander, G.; Muhlack, R.; Pocock, K.F.; Colby, C.; O’Neill, B.K.; Høj, P.B.; Jones, P. Preventing protein haze in bottled white wine. Aust. J. Grape Wine Res. 2005, 11, 215–225. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| 48 h | End of Fermentation | |||||
|---|---|---|---|---|---|---|
| Strains | Glucose (g/L) | Fructose (g/L) | Glycerol (g/L) | Ethanol (%v/v) | Succinic Acid (g/L) | Acetic Acid (g/L) |
| EC1118 | 90.14 ± 1.03 a | 91.13 ± 0.57 ab | 5.17 ± 0.07 a | 11.76 ± 0.01 b | 2.52 ± 0.05 ab | 0.43 ± 0.03 a |
| FRI719 | 120.12 ± 0.52 c | 91.28 ± 0.40 abc | 8.70 ± 0.02 fg | 11.47 ± 0.09 ab | 2.50 ± 0.35 ab | 0.72 ± 0.17 abc |
| FRI728 | 121.55 ± 1.08 c | 94.06 ± 0.29 bcde | 7.61 ± 0.14 bcd | 11.39 ± 0.25 ab | 2.67 ± 0.02 ab | 0.37 ± 0.03 a |
| FRI729 | 120.17 ± 2.95 c | 95.12 ± 0.36 e | 8.25 ± 0.10 ef | 11.50 ± 0.19 ab | 2.56 ± 0.01 ab | 0.40 ± 0.01 a |
| FRI751 | 122.34 ± 0.22 c | 94.92 ± 0.11 de | 7.92 ± 0.03 cde | 11.52 ± 0.23 ab | 2.72 ± 0.05 ab | 0.60 ± 0.27 abc |
| FRI754 | 121.97 ± 0.28 c | 94.2 ± 0.64 bcde | 8.53 ± 0.11 f | 11.43 ± 0.25 ab | 2.48 ± 0.27 ab | 0.67 ± 0.31 abc |
| FRI7100 | 122.73 ± 0.47 c | 94.43 ± 0.34 cde | 9.12 ± 0.15 g | 11.26 ± 0.09 ab | 3.16 ± 0.11 b | 1.10 ± 0.02 c |
| PAS13 | 121.84 ± 1.38 c | 95.49 ± 0.73 ef | 7.95 ± 0.17 cde | 11.48 ± 0.12 ab | 3.34 ± 0.12 b | 1.17 ± 0.09 c |
| PAS55 | 124.14 ± 5.04 c | 98.51 ± 2.05 fg | 7.39 ± 0.16 b | 11.34 ± 0.25 ab | 2.71 ± 0.05 ab | 0.56 ± 0.19 abc |
| PAS66 | 119.33 ± 0.74 c | 97.16 ± 1.94 ef | 7.55 ± 0.07 bc | 11.12 ± 0.32 ab | 3.05 ± 0.21 ab | 0.93 ± 0.23 abc |
| PAS92 | 109.63 ± 3.28 b | 101.22 ± 2.34 g | 8.05 ± 0.18 de | 11.48 ± 0.18 ab | 2.26 ± 0.79 ab | 0.61 ± 0.28 abc |
| PAS103 | 120.09 ± 0.48 c | 91.92 ± 0.21 abcd | 7.87 ± 0.35 cde | 11.40 ± 0.16 ab | 1.98 ± 1.02 a | 0.54 ± 0.45 abc |
| PAS151 | 118.89 ± 0.71 c | 91.39 ± 0.30 abc | 7.97 ± 0.18 cde | 11.19 ± 0.25 ab | 2.93 ± 0.29 ab | 0.44 ± 0.18 ab |
| PAS173 | 119.12 ± 0.16 c | 90.76 ± 0.68 a | 7.94 ± 0.10 cde | 10.91 ± 0.49 a | 3.09 ± 0.08 ab | 1.07 ± 0.05 bc |
| 48 h | End of Fermentation | |||||
|---|---|---|---|---|---|---|
| Strains | Glucose (g/L) | Fructose (g/L) | Glycerol (g/L) | Ethanol (%v/v) | Succinic Acid (g/L) | Acetic Acid (g/L) |
| EC1118 | 75.80 ± 2.13 a | 85.78 ± 1.33 a | 5.36 ± 0.21 a | 11.11 ± 0.58 c | 1.45 ± 0.05 a | 0.68 ± 0.01 abcde |
| FRI719 | 106.83 ± 1.77 d | 98.46 ±0.38 c | 6.79 ± 0.15 cd | 11.41 ± 0.07 c | 1.46 ± 0.08 a | 0.49 ± 0.06 a |
| FRI728 | 105.99 ± 1.70 cd | 94.17 ± 1.75 b | 7.23 ± 0.11 de | 11.30 ± 0.01 c | 1.62 ± 0.09 ab | 0.64 ± 0.07 abcd |
| FRI729 | 102.50 ± 1.89 bc | 92.71 ± 1.21 b | 7.89 ± 0.22 fg | 11.26 ± 0.02 c | 1.67 ± 0.17 abc | 0.81 ± 0.16 cde |
| FRI751 | 105.48 ± 0.51 cd | 98.67 ± 0.70 c | 7.38 ± 0.24 ef | 11.31 ± 0.10 c | 1.69 ± 0.06 abc | 0.66 ± 0.05 abcde |
| FRI754 | 98.75 ± 0.53 b | 93.40 ± 0.90 b | 7.77 ± 0.13 fg | 11.13 ± 0.03 c | 1.61 ± 0.09 ab | 0.84 ± 0.06 de |
| FRI7100 | 100.04 ± 0.62 b | 93.82 ± 0.34 b | 8.00 ± 0.05 g | 11.15 ± 0.10 c | 1.83 ± 0.15 bc | 0.86 ± 0.09 e |
| PAS13 | 102.88 ± 0.65 bcd | 95.73 ± 0.72 bc | 7.10 ± 0.05 cde | 11.39 ± 0.02 c | 1.69 ± 0.11 abc | 0.65 ± 0.03 abcde |
| PAS55 | 100.55 ± 1.39 b | 93.61 ± 0.45 b | 6.70 ± 0.29 c | 11.38 ± 0.06 c | 1.79 ± 0.07 bc | 0.69 ± 0.07 abcde |
| PAS66 | 100.77 ± 0.58 b | 93.17 ± 0.96 b | 7.07 ± 0.06 cde | 11.21 ± 0.06 c | 1.91 ± 0.10 bc | 0.64 ± 0.01 abcd |
| PAS92 | 102.76 ± 0.97 bcd | 94.12 ± 2.12 b | 5.94 ± 0.27 b | 9.58 ± 0.17 a | 1.64 ± 0.11 abc | 0.59 ± 0.11 ab |
| PAS103 | 102.13 ±2.74 bc | 92.25 ± 2.78 b | 5.83 ± 0.10 ab | 9.18 ± 0.38 a | 1.70 ± 0.18 abc | 0.63 ± 0.06 abc |
| PAS151 | 102.32 ± 1.25 bc | 95.96 ± 0.99 bc | 6.15 ± 0.01 b | 10.36 ± 0.13 b | 1.88 ± 0.04 bc | 0.57 ± 0.01 ab |
| PAS173 | 100.89 ± 0.61 b | 95.68 ± 0.70 bc | 7.04 ± 0.21 cde | 11.00 ± 0.34 bc | 1.95 ± 0.08 c | 0.71 ± 0.03 bcde |
| 48 h | End of Fermentation | |||||
|---|---|---|---|---|---|---|
| Strains | Glucose(g/L) | Fructose(g/L) | Glycerol(g/L) | Ethanol(%v/v) | Succinic Acid (g/L) | Acetic Acid (g/L) |
| EC1118 | 59.49 ± 2.93 a | 75.70 ± 1.65 a | 5.01 ± 0.17 a | 12.40 ± 0.11 c | 2.42 ± 0.21 a | 0.65 ± 0.01 abc |
| FRI719 | 103.18 ± 1.05 b | 86.19 ± 1.18 abcd | 8.21 ± 0.71 cd | 11.71 ± 0.96 bc | 2.30 ± 0.05 a | 0.70 ± 0.09 abc |
| FRI728 | 103.54 ± 2.59 b | 91.76 ± 6.83 cd | 7.05 ± 0.49 bcd | 11.17 ± 0.26 abc | 2.33 ± 0.09 a | 0.64 ± 0.09 abc |
| FRI729 | 97.21 ± 5.59 b | 87.04 ± 5.52 abcd | 6.86 ± 0.54 bcd | 10.97 ± 0.19 abc | 2.15 ± 0.09 a | 0.53 ± 0.03 a |
| FRI751 | 99.55 ± 5.05 b | 88.71 ± 6.60 bcd | 7.23 ± 0.57 bcd | 10.92 ± 0.25 abc | 2.31 ± 0.02 a | 0.62 ± 0.08 abc |
| FRI754 | 93.04 ± 4.53 b | 78.44 ± 4.16 ab | 7.70 ± 0.36 bcd | 10.89 ± 0.27 ab | 2.19 ± 0.07 a | 0.64 ± 0.09 abc |
| FRI7100 | 93.33 ± 5.95 b | 78.91 ± 5.59 abc | 8.31 ± 0.18 d | 10.71 ± 0.10 ab | 2.43 ± 0.16 a | 0.74 ± 0.12 abc |
| PAS13 | 91.63 ± 1.37 b | 76.42 ± 0.85 ab | 8.17 ± 0.64 cd | 10.70 ± 0.45 ab | 2.42 ± 0.13 a | 0.83 ± 0.10 c |
| PAS55 | 98.91 ± 3.70 b | 85.77 ± 3.48 abcd | 7.36 ± 0.58 bcd | 10.97 ± 0.28 abc | 2.26 ± 0.09 a | 0.60 ± 0.04 abc |
| PAS66 | 101.57 ± 3.36 b | 94.82 ± 1.75 d | 6.67 ± 0.52 bc | 10.90 ± 0.34 ab | 2.21 ± 0.09 a | 0.54 ± 0.03 ab |
| PAS92 | 100.87 ± 3.57 b | 94.11 ± 3.22 d | 6.42 ± 0.28 b | 10.23 ± 0.41 ab | 2.16 ± 0.06 a | 0.52 ± 0.02 a |
| PAS103 | 99.40 ± 4.04 b | 93.45 ± 5.22 d | 7.31 ± 0.68 bcd | 10.93 ± 0.30 abc | 2.34 ± 0.17 a | 0.65 ± 0.12 abc |
| PAS151 | 98.44 ± 5.91 b | 86.73 ± 5.43 abcd | 7.76 ± 0.44 bcd | 11.13 ± 0.21 abc | 2.09 ± 0.12 a | 0.60 ± 0.04 abc |
| PAS173 | 99.64 ± 1.89 b | 91.98 ± 1.69 d | 6.85 ± 0.77 bcd | 10.20 ± 1.26 a | 2.41 ± 0.30 a | 0.76 ± 0.09 bc |
| Polysaccharides * (mg/L) | ||||
|---|---|---|---|---|
| Strains | Total | High MW | Medium MW | Low MW |
| EC1118 | 135.54 ± 2.11 a | 14.53 ± 0.93 ab | 56.96 ± 1.54 a | 64.05 ± 0.78 a |
| FRI719 | 219.86 ± 0.44 f | 13.74 ± 0.28 a | 88.05 ± 0.96 d | 118.06 ± 1.59 e |
| FRI728 | 227.43 ± 2.20 g | 16.57 ± 0.74 abcde | 102.15 ± 1.30 fg | 108.71 ± 1.46 d |
| FRI729 | 187.33 ± 3.59 bc | 17.53 ± 1.35 cdef | 81.33 ± 1.13 b | 88.47 ± 1.62 b |
| FRI751 | 237.82 ± 1.18 h | 18.64 ± 0.66 defg | 103.30 ± 0.88 g | 115.89 ± 0.46 e |
| FRI754 | 216.52 ± 2.64 f | 16.50 ± 1.41 abcde | 92.56 ± 1.58 e | 107.46 ± 1.38 d |
| FRI7100 | 281.07 ± 0.11 i | 20.31 ± 0.30 fg | 111.84 ± 0.76 h | 148.91 ± 1.12 f |
| PAS13 | 199.52 ± 2.91 d | 19.16 ± 0.09 efg | 82.33 ± 1.68 b | 98.02 ± 1.36 c |
| PAS55 | 208.59 ± 2.17 e | 15.58 ± 1.23 abc | 86.69 ± 0.95 cd | 106.32 ± 1.20 d |
| PAS66 | 183.76 ± 1.40 b | 16.18 ± 0.47 abcd | 81.17 ± 0.95 b | 86.41 ± 1.75 b |
| PAS92 | 223.37 ± 0.68 fg | 17.08 ± 0.58 bcde | 92.04 ± 0.87 e | 114.25 ± 1.06 e |
| PAS103 | 308.64 ± 1.90 j | 20.54 ± 0.97 g | 98.74 ± 1.00 f | 189.36 ± 1.85 g |
| PAS151 | 193.23 ± 3.06 cd | 21.44 ± 1.23 g | 82.12 ± 0.96 b | 89.67 ± 1.89 b |
| PAS173 | 192.08 ± 4.15 c | 19.06 ± 1.73 defg | 83.34 ± 1.29 bc | 89.68 ± 1.88 b |
| Polysaccharides * (mg/L) | ||||
|---|---|---|---|---|
| Strains | Total | High MW | Medium MW | Low MW |
| EC1118 | 230.96 ± 3.90 b | 25.00 ± 1.72 de | 82.32 ± 1.22 a | 123.64 ± 2.94 b |
| FRI719 | 265.94 ± 3.67 de | 23.07 ± 0.92 cd | 96.26 ± 1.11 bcd | 146.61 ± 3.35 ef |
| FRI728 | 263.97 ± 0.83 de | 16.49 ± 1.02 a | 97.16 ± 0.87 bcd | 150.32 ± 0.96 f |
| FRI729 | 271.07 ± 1.30 ef | 24.50 ± 1.20 de | 105.30 ± 1.09 ef | 141.27 ± 1.06 de |
| FRI751 | 290.29 ± 5.32 h | 26.23 ± 1.12 de | 106.28 ± 3.73 fg | 157.78 ± 1.69 g |
| FRI754 | 215.38 ± 5.08 a | 15.41 ± 1.21 a | 79.96 ± 2.76 a | 120.01 ± 2.14 b |
| FRI7100 | 300.42 ± 2.52 i | 32.78 ± 1.61 f | 107.46 ± 1.12 fgh | 160.18 ± 1.83 g |
| PAS13 | 279.29 ± 1.71 fg | 20.60 ± 1.24 bc | 100.64 ± 0.70 de | 158.05 ± 1.18 g |
| PAS55 | 241.87 ± 6.10 c | 17.00 ± 1.56 a | 92.22 ± 2.63 b | 132.65 ± 2.47 c |
| PAS66 | 255.86 ± 2.14 d | 17.82 ± 0.60 ab | 97.99 ± 1.02 cd | 140.05 ± 1.57 d |
| PAS92 | 327.54 ± 0.64 j | 17.47 ± 1.00 ab | 111.49 ± 0.87 gh | 198.58 ± 1.17 h |
| PAS103 | 351.21 ± 3.86 k | 27.02 ± 0.50 e | 112.72 ± 1.14 h | 211.47 ± 2.78 i |
| PAS151 | 286.56 ± 2.01 gh | 18.41 ± 0.62 ab | 108.82 ± 1.39 fgh | 159.33 ± 1.05 g |
| PAS173 | 224.52 ± 2.10 ab | 18.56 ± 0.83 ab | 92.96 ± 2.35 bc | 113.00 ± 1.31 a |
| Polysaccharides * (mg/L) | ||||
|---|---|---|---|---|
| Strains | Total | High MW | Medium MW | Low MW |
| EC1118 | 276.10 ± 2.63 bc | 59.50 ± 1.02 h | 124.52 ± 0.97 c | 92.08 ± 1.60 a |
| FRI719 | 253.08 ± 4.24 a | 32.76 ± 2.62 ab | 97.45 ± 1.44 a | 122.87 ± 2.06 ef |
| FRI728 | 324.71 ± 1.05 g | 29.12 ± 1.19 a | 176.32 ± 1.96 i | 119.27 ± 1.10 de |
| FRI729 | 254.33 ± 0.21 a | 35.75 ± 1.80 bc | 117.18 ± 1.71 b | 101.39 ± 1.06 b |
| FRI751 | 317.70 ± 1.85 f | 49.44 ± 1.39 g | 148.98 ± 1.39 f | 119.29 ± 0.80 de |
| FRI754 | 322.49 ± 0.55 fg | 46.07 ± 1.79 efg | 157.20 ± 1.24 g | 119.22 ± 1.06 de |
| FRI7100 | 350.04 ± 1.32 i | 41.70 ± 1.37 def | 150.38 ± 1.90 f | 157.95 ± 1.25 h |
| PAS13 | 282.23 ± 2.98 d | 29.70 ± 2.30 a | 128.65 ± 1.03 cd | 123.88 ±1.40 f |
| PAS55 | 333.45 ± 1.56 h | 31.1 ± 1.61 ab | 165.66 ± 1.23 h | 136.69 ± 0.47 g |
| PAS66 | 298.90 ± 2.08 e | 40.12 ± 1.84 cd | 141.68 ± 1.32 e | 117.10 ± 1.72 d |
| PAS92 | 302.00 ± 0.71 e | 39.29 ± 1.07 cd | 143.62 ± 0.90 e | 119.09 ± 1.46 de |
| PAS103 | 280.14 ± 1.66 cd | 40.52 ± 1.51 cde | 116.57 ± 1.79 b | 123.04 ± 1.39 ef |
| PAS151 | 274.11 ± 1.37 b | 46.82 ± 2.94 fg | 118.73 ± 1.04 b | 108.56 ± 1.92 c |
| PAS173 | 303.00 ± 2.09 e | 47.21 ± 2.30 fg | 130.62 ± 0.91 d | 125.17 ± 1.52 f |
| Sauvignon Blanc | |||
|---|---|---|---|
| Yeast Strains | Total Mannoproteins (mg/L) | Medium MW Mannoproteins (mg/L) | Low MW Mannoproteins (mg/L) |
| EC1118 | 39.77 ± 1.87 a | 0.68 ± 0.33 a | 39.09 ± 1.69 a |
| FRI751 | 61.09 ± 0.55 b | 8.63 ± 0.56 c | 52.46 ± 0.89 b |
| FRI7100 | 85.83 ± 1.94 d | 9.04 ± 0.16 c | 76.79 ± 1.78 e |
| PAS13 | 76.39 ± 1.17 c | 4.60 ± 1.28 b | 71.79 ± 1.47 d |
| PAS55 | 60.00 ± 2.18 b | 9.55 ± 1.04 c | 50.46 ± 1.78 b |
| PAS103 | 72.70 ± 2.21 c | 14.34 ± 1.32 d | 58.37 ± 1.25 c |
| Pinot Grigio | |||
| Yeast Strains | Total Mannoproteins (mg/L) | Medium MW Mannoproteins (mg/L) | Low MW Mannoproteins (mg/L) |
| EC1118 | 42.29 ± 1.71 a | 1.57 ± 0.55 a | 40.72 ± 1.41 a |
| FRI751 | 75.69 ± 2.43 d | 20.05 ± 1.40 d | 55.64 ± 1.20 c |
| FRI7100 | 74.33 ± 2.37 d | 16.35 ± 1.06 c | 57.98 ± 1.51 c |
| PAS13 | 55.97 ± 2.12 b | 9.88 ± 0.65 b | 46.09 ± 1.76 b |
| PAS55 | 66.07 ± 2.59 c | 16.58 ± 1.17 c | 49.48 ± 1.45 b |
| PAS103 | 93.24 ± 2.14 e | 30.26 ± 0.48 e | 62.98 ± 1.94 d |
| Manzoni Bianco | |||
| Yeast Strains | Total Mannoproteins (mg/L) | Medium MW Mannoproteins (mg/L) | Low MW Mannoproteins (mg/L) |
| EC1118 | 31.57 ± 0.04 a | 0.16 ± 0.05 a | 31.44 ± 0.07 a |
| FRI751 | 96.46 ± 1.04 d | 25.03 ± 1.04 e | 71.42 ± 1.95 d |
| FRI7100 | 94.21 ± 1.35 d | 7.84 ± 0.55 c | 86.37 ± 1.08 e |
| PAS13 | 98.51 ± 2.97 d | 15.52 ± 1.37 d | 82.90 ± 1.61 e |
| PAS55 | 48.11 ± 2.05 b | 3.08 ± 0.70 b | 45.02 ± 1.89 b |
| PAS103 | 65.43 ± 2.57 c | 9.33 ± 1.01 c | 56.09 ± 1.56 c |
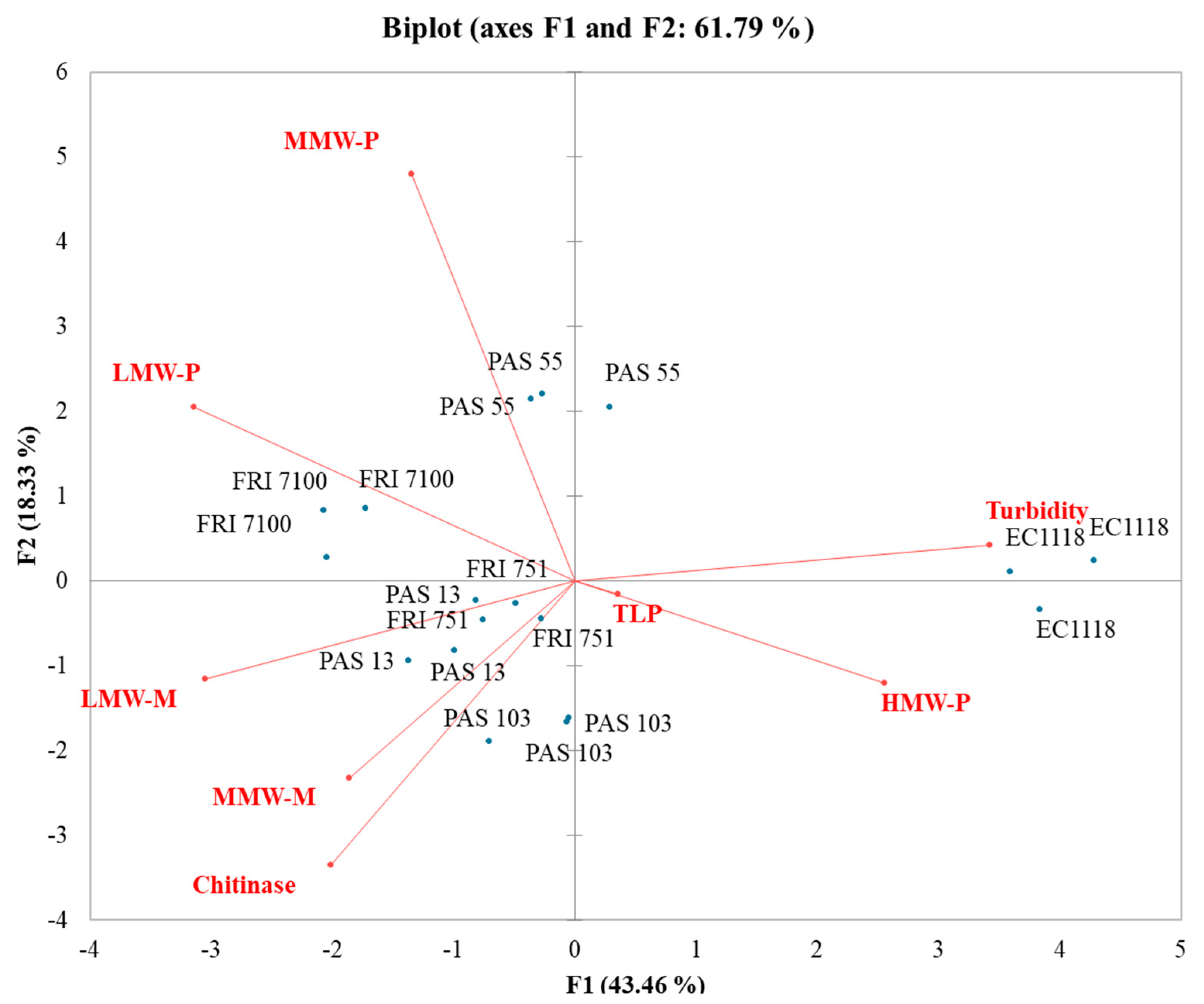
| Variables | SB | PG | MB |
|---|---|---|---|
| Turbidity | 1 | 1 | 1 |
| TLP | −0.751 | 0.331 | 0.097 |
| Chitinase | −0.765 | 0.096 | −0.586 |
| HMW-P | −0.560 | 0.384 | 0.568 |
| MMW-P | −0.474 | 0.262 | −0.292 |
| LMW-P | −0.681 | 0.618 | −0.770 |
| MMW-M | −0.761 | 0.351 | −0.477 |
| LMW-M | −0.422 | 0.350 | −0.692 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moreira, L.d.P.D.; Nadai, C.; da Silva Duarte, V.; Brearley-Smith, E.J.; Marangon, M.; Vincenzi, S.; Giacomini, A.; Corich, V. Starmerella bacillaris Strains Used in Sequential Alcoholic Fermentation with Saccharomyces cerevisiae Improves Protein Stability in White Wines. Fermentation 2022, 8, 252. https://doi.org/10.3390/fermentation8060252
Moreira LdPD, Nadai C, da Silva Duarte V, Brearley-Smith EJ, Marangon M, Vincenzi S, Giacomini A, Corich V. Starmerella bacillaris Strains Used in Sequential Alcoholic Fermentation with Saccharomyces cerevisiae Improves Protein Stability in White Wines. Fermentation. 2022; 8(6):252. https://doi.org/10.3390/fermentation8060252
Chicago/Turabian StyleMoreira, Luiza de Paula Dias, Chiara Nadai, Vinícius da Silva Duarte, Edward John Brearley-Smith, Matteo Marangon, Simone Vincenzi, Alessio Giacomini, and Viviana Corich. 2022. "Starmerella bacillaris Strains Used in Sequential Alcoholic Fermentation with Saccharomyces cerevisiae Improves Protein Stability in White Wines" Fermentation 8, no. 6: 252. https://doi.org/10.3390/fermentation8060252
APA StyleMoreira, L. d. P. D., Nadai, C., da Silva Duarte, V., Brearley-Smith, E. J., Marangon, M., Vincenzi, S., Giacomini, A., & Corich, V. (2022). Starmerella bacillaris Strains Used in Sequential Alcoholic Fermentation with Saccharomyces cerevisiae Improves Protein Stability in White Wines. Fermentation, 8(6), 252. https://doi.org/10.3390/fermentation8060252