Abstract
Background: Soy sauce is applied as a seasoning in daily life in East Asia. High amounts of salt in soy sauce can inhibit microbial growth in its production and preservation. However, the elevated salt content in food might increase the health risk. Low salt fermentation is rarely used in soil sauce production because of the potential harmful lactic acid bacteria growth. Therefore, dilution after high salt fermentation is commonly used to reduce the salt concentration. Methods: This study aims to treat the low salt fermented soy sauce with a high hydrostatic pressure process (HPP) to eliminate the harmful bacteria and to analyze the microbial community change compared with that in traditional high salt fermentation. Results: The results showed that the bacterial diversity was increased in low-salt and high-pressure (LS-HP)-treated soy sauce, though the bacterial abundance was decreased. Relative abundance in high-salt (HS), low-salt (LS), and LS-HP-treated soy sauce showed specific bacterial strains in the LS-HP group. Similarly, the fungal diversity was also increased in LS-HP-fermented soy sauce and the detected OTUs were increased. The fungi sensitive to salinity and pressure were indicated in our results. Conclusions: The present study suggests the enhanced bacterial and fungal diversity and different microbial community in HS, LS, and LS-HP-treated soy sauce, as well as the availability of LS and HPP treatment on soy sauce production.
1. Introduction
Soy sauce has a long history for its use as a traditional seasoning from the Orient. Due to its commercial importance and the interdependence of the microflora in production, the microbial community during soy sauce fermentation is generally researched with high interest. The molds can produce enzymes to saccharify the starch and decompose the protein in raw materials during preparation of koji. Moreover, ingredients from koji can be utilized by yeast and bacteria in the following brine fermentations [1]. Aspergillus oryzae is well known for its use in soy sauce fermentation. During the fermentation, when the temperature of mixture reaches 37 °C, the mold in the mixture can grow mycelium after stirring [2]. The single-strain fermentation of A. oryzae is widely used in industrial soy sauce production; therefore, soy sauce is commonly known as a fungal fermented seasoning. In addition, the microflora related to soy sauce also includes yeast and bacteria, and the microbial population in koji has been reported in some research [3].
During the fermentation of group 2A carcinogenic compounds, ethyl carbamate (EC) is formed as a byproduct [4,5]. EC can be found in a variety of foods, for example, soy sauce, and fermented food and beverages [6,7,8,9,10,11]. EC can be formed during some chemical reactions of ethanol and EC precursors such as urea, citrulline, hydrocyanic acid, and carbamyl phosphate [12]. Some reports have revealed that urea and citrulline can also be found in soy sauce, and citrulline is the major EC precursor among them [13,14]. Aspergilli and some film-forming yeasts can contribute to spoilage of non-pasteurized food or of food that contains low levels of salt [15]. Some strains of spoilage bacteria can grow in soy sauces with low levels of salt (<15%) and affect the food safety; accordingly, developing a healthier soy sauce product is always constricted by low-salt (LS) fermentation.
On the other side, Staphylococcus, Micrococcus, and Bacillus play major roles in alcoholic fermentation of soy sauce making, as Pediococcus is inhibited by salty brine [16,17]. The salty brine may partially affect arginine catabolism in Pediococcus acidilactici and cause formation of more citrulline instead of ornithine. This is also a significant pathway of citrulline formation that occurs in lactic acid fermentation in soy sauce [14]. Nevertheless, there is still no report found relevant to microbes, which can produce citrulline and citrulline accumulation during the alcoholic fermentation in soy sauce [18]. As a result, the potential methods of inhibiting formation and accumulation of citrulline may be essential for controlling EC level in LS fermented soy sauce.
The hydrostatic pressure process (HPP) has been used in food processing, such as soy sauce, sausages, and beverages, to inhibit the microbial activity without additional chemical reactions [19]. Therefore, soy sauce with no salt is possible to produce using HPP, which might be better for the patients who need to minimize the salt intake [19]. Other fermented foods including brine green asparagus, mango pulp, and pitted sweet cherries showed a change in microbial community and better preservation after HPP [20,21]. In addition, varied microbial community and metabolites in high-salt (HS) fermented soy sauce after HHP were also reported [22]. However, few studies are investigating the microbial change in LS-fermented soy sauce with HPP. Therefore, our study aims to analyze the microbial community change in LS fermentation with HPP using high-throughput sequencing. HS, LS, and LS with HHP (LS-HP)-treated soy sauce is assessed for bacterial and fungal community. In addition, microbial diversity and correlation between different treatments were studied. Our results provide insights into the microbial change in soy sauce under low levels of salt and HPP treatment, and the difference compared to traditional high levels of salt-fermented soy sauce.
2. Materials and Methods
2.1. Koji Fermentation
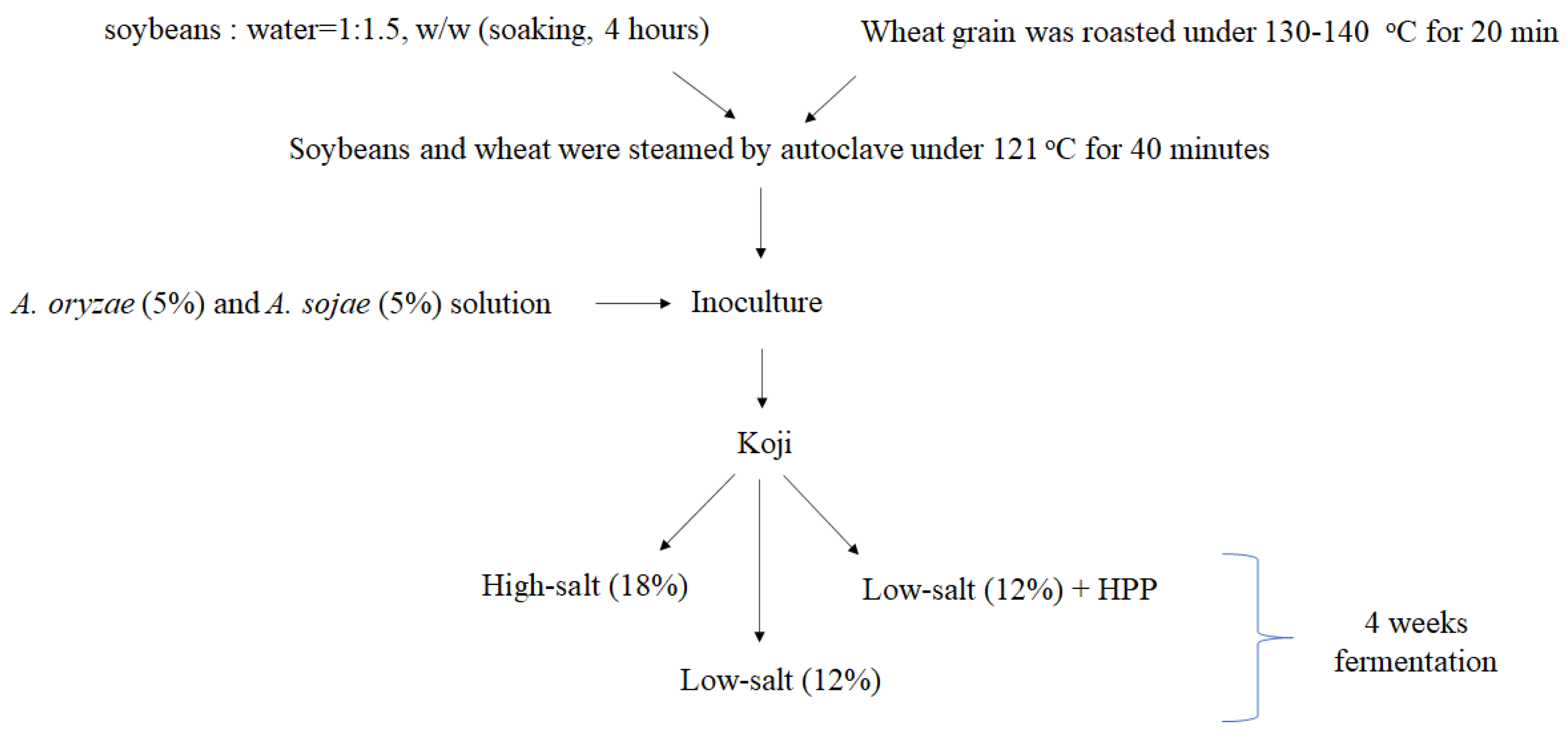
The soybeans with uniform shape and color were selected, washed, and then soaked in reverse osmosis water (soybeans: water = 1:1.5, w/w) for 4 h until being completely soften. Intact wheat grain with no mildew were selected and roasted under 130–140 °C for 20 min. During the roasting process, flavor was generated and color turned light brown. After roasting, the wheat was cooled down below 30 °C and then crushed for further use. The hydrated soybeans and wheat grain were mixed and steamed by means of autoclave under 121 °C for 40 min and then ground into paste. The soybean paste was cooled down below 37 °C. As starter mold (10%), the equal amount of Aspergillus oryzae and A. sojae were added into the soybean–wheat mixture for making the koji. The moisture content of mixture was adjusted to 39.5%. The koji mixture was loaded into trays and evened by 2 cm thickness. Each koji sample was then crushed with a 2.5 cm diameter hole at the center for mold to grow. In addition, the processes of soy sauce fermentation (soy koji:salt solution = 1:2) were divided into three groups including high-salt (HS; 18% salt saline), low-salt (HS; 12% salt saline), and low-salt with high hydrostatic pressure process (HPP) treatment (300 MPa) at 4 °C for 5 min. Moreover, the koji preparation and soy sauce fermentation were carried out in laminar flow to avoid contamination (Figure 1).
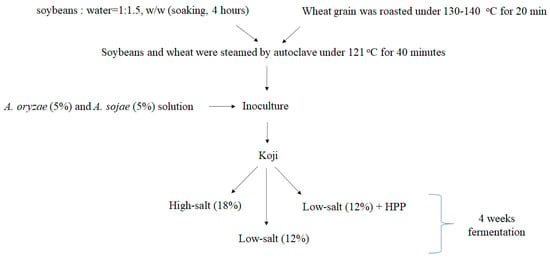
Figure 1.
The schematic representation of soy sauce.
2.2. Microbiota Assay
The DNA was extracted from HS, LS, and LS-HP-fermented soy sauce at week 0 and week 4 according to manufacturer’s instruction, and stored at −80 °C for the following analysis. The quality and quantity of the DNA samples were checked by NanoDrop spectrophotometer (Thermo Fisher Scientific, Waltham, MA, USA). The V3–V4 region of bacterial 16s rDNA was amplified by primers: 338F (5′-ACTCCTACGGGAGGCAGCA-3′) and 806R (5′-GGACTACHVGGGTWTCTAAT-3′). For fungi, primers ITS5 (5′-GGAAGTAAAAGTCGTAACAAGG-3′) and ITS1 (5′-GCTGCGTTCTTCATCGATGC-3′) were used to amplify internal transcribed spacer (ITS) regions. The PCR amplicons were incorporated with 5′-barcode and purified for following sequencing. The high-throughput sequencing was conducted by Illumina MiSeq platform and Miseq Reagent kit v3 (Illumina, Inc., Santiago, CA, USA). The sequences would be removed if they were <150 bp, with ambiguous base, and no barcodes. The DNA fragments were clustering into operational taxonomic units (OTUs) with threshold of 97% sequence identity. The OTUs were classified using BLAST searching and Greengenes Database.
2.3. Statistical Analysis
All data were recorded as mean ± SD. The statistical significance was determined by one-way analysis of variance (ANOVA) using the SAS general linear model procedure (SAS Inc., Cary, NC, USA), followed by ANOVA with Duncan’s test (p < 0.05). The microbial alpha and beta diversity including Shannon and Simpson diversity, and uniFrac principal coordinate analysis (PCoA), respectively. The difference between fungal OTUs was presented as Venn diagrams. Heat map and heat trees were used to illustrate the diversity change in bacteria and fungi.
3. Results and Discussion
3.1. Bacterial Diversity in Soy Sauce after High-Salt (HS), Low-Salt (LS), and Low-Salt High Pressure (LS-HP)
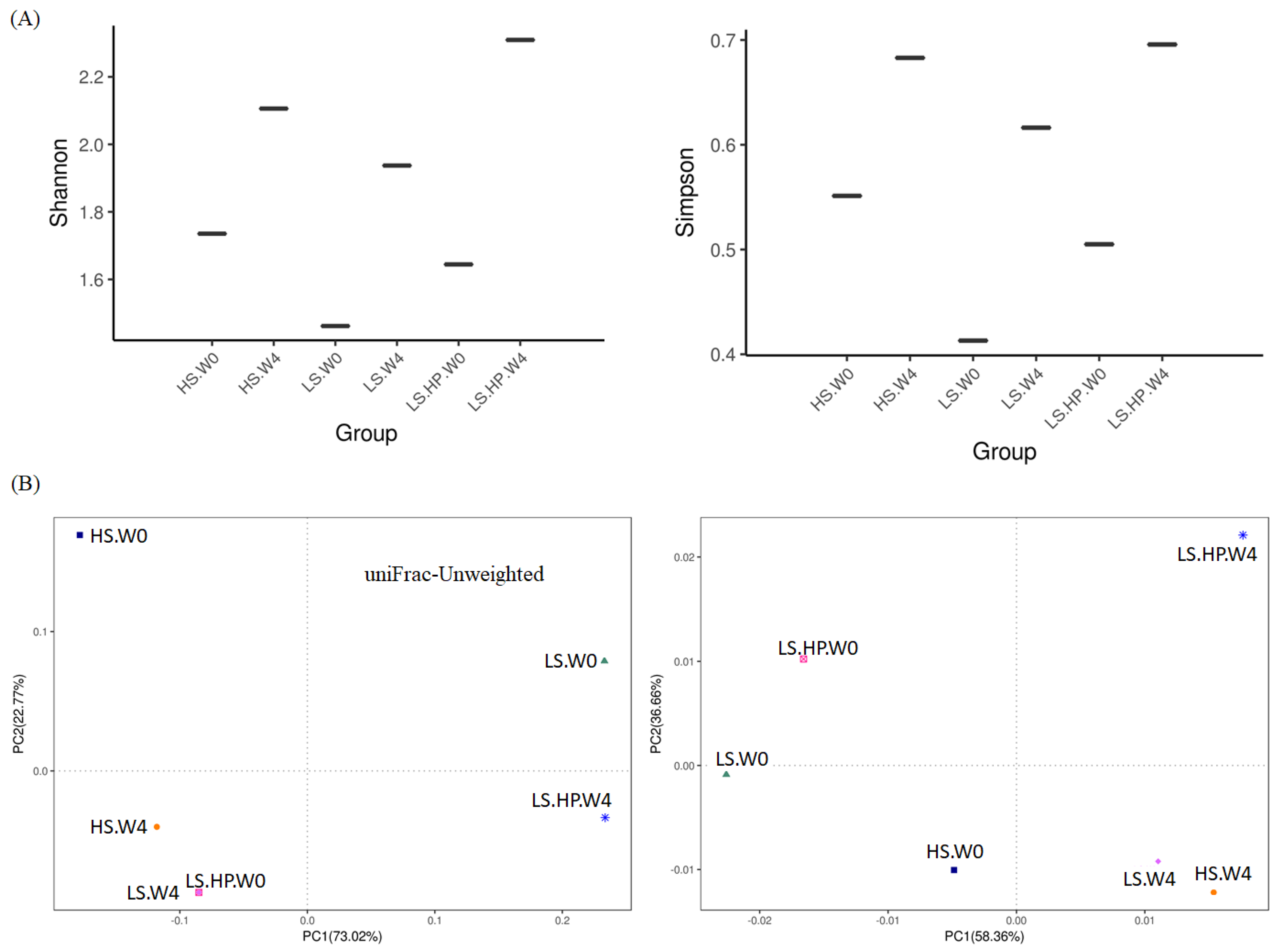
Figure 2A are Shannon–Wiener and Simpson’s indexes of alpha diversity, which are commonly used to evaluate diversity of organism community in certain sample, the higher value is regard as the greater diversity of the sample. From Figure 2A, both the Shannon–Weiner and Simpson’s indexes showed that the LS-HP-treated sample after 4-week fermentation exhibited the highest values among the samples tested, which were thought to have a high degree of community diversities. Figure 2B shows the uniFrac diagrams of PCoA, which can be used to measure the distance metric of biological relationships in microorganisms. The unweighted uniFrac is qualitative method for evaluating diversity of the community by measuring the relative distance between plots of samples, while the weighted uniFrac is a quantitative one [23]. Figure 2B shows that plots of HS and LS after 4-week fermentation were relatively close to each other compared to other groups, namely the bacteria communities from these two samples might be similar. Despite the difference in week 0, the related result was also found in the weighted-uniFrac diagram, which indicates the presence of similar bacterial microbiota in the HS and LS groups after 4-week fermentation. Thus, the predominant bacteria in HS and LS soy sauce might grow similarly during fermentation. In addition, the microbiota in the LS-HP group was found greatly different from other two groups after 4-week fermentation, which can be reasonably speculated that the HPP treatment changed the microbial community and influenced growth of primeval bacteria. Compared with our results, other soy sauce research using high-throughput sequencing on microbial diversity shared the similar range of Shannon and Simpson indexes, which were 1.57–1.97 and 0.29–0.34, respectively [24,25]. On the other hand, HS soy sauce mash inoculated with yeasts or yeasts + Tetragenococcus halophilus showed different diversity indexes (Shannon: 4.08–4.42; Simpson: 0.81–0.87) and only slight change on bacterial diversity after HPP [22]. The difference might be due to the additional yeasts and bacterium inoculation, HS fermentation, and the longer fermentation time (180 days) applied by Liang et al. [22]. Our results suggest that the HPP increased the bacterial diversity in LS-fermented soy sauce (Figure 2A), which is further confirmed by the PCoA of beta diversity results showing the high difference in the bacterial community of the LS-HP group compared with the HS and LS ones (Figure 2B).
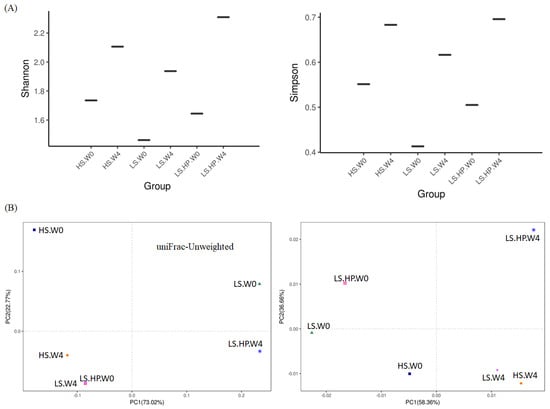
Figure 2.
(A) The Shannon–Weiner and Simpson’s indexes of alpha-diversity, and (B) The unweighted and weighted uniFrac PCoA plots of beta-diversity obtained from analysis of bacteria microbiota in high-salt (HS), low-salt (LS), and low-salt high-pressure (LS-HP) soy sauce during fermentation.
3.2. Bacterial Variation in HS, LS, and LS-HP-Treated Soy Sauce
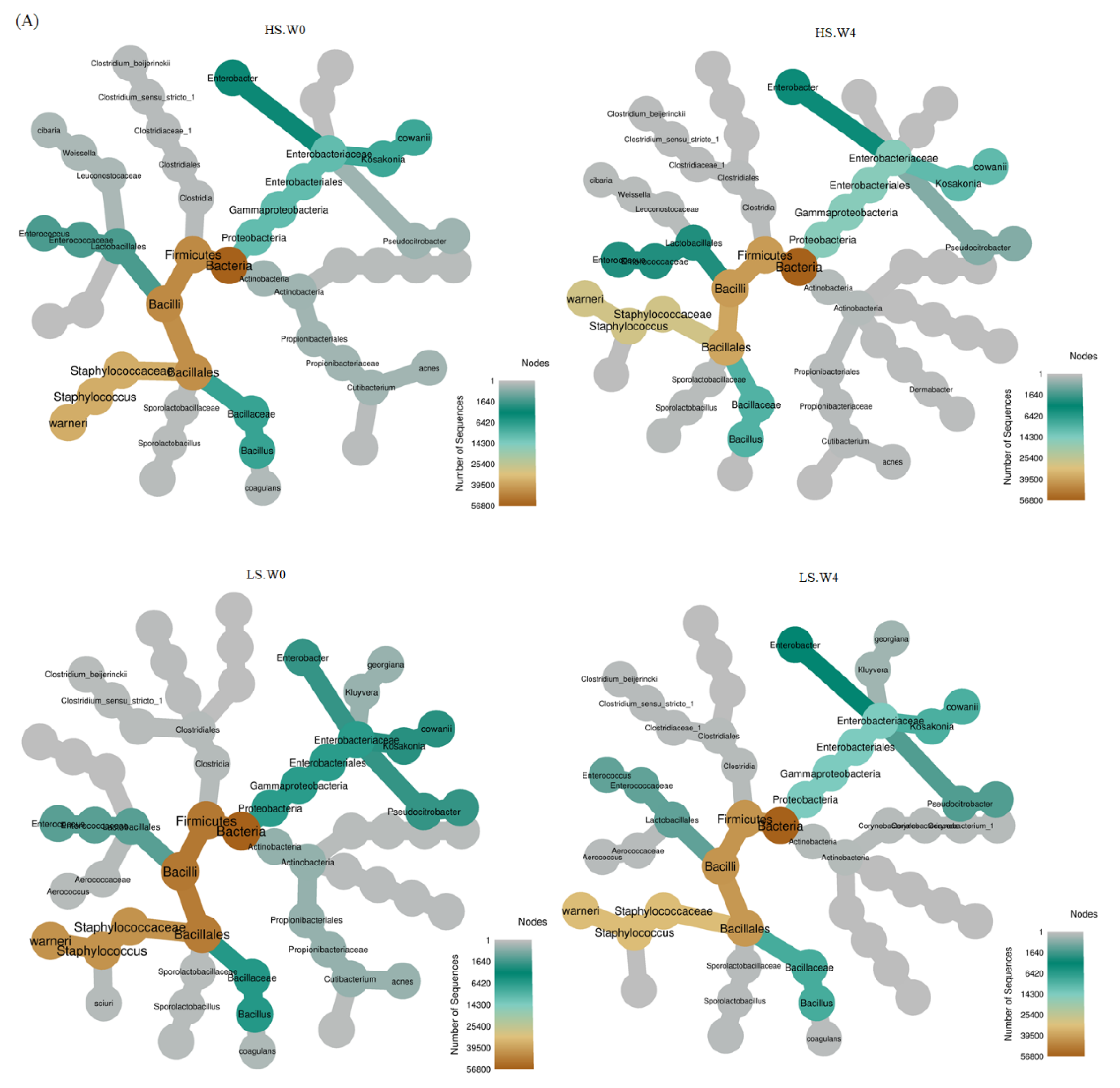
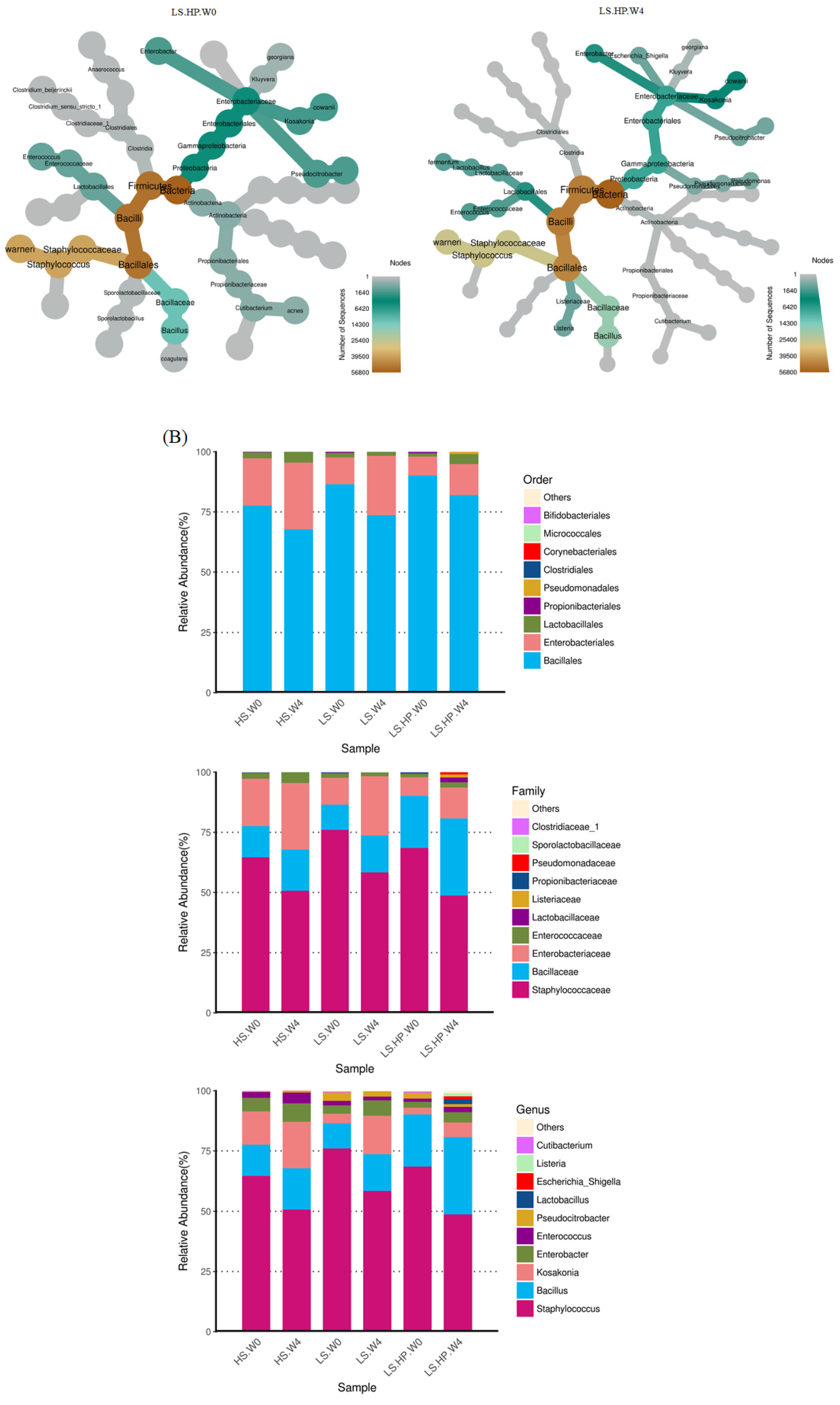
The decrease in Staphylococcus suggested in Figure 3B is similar with other bacterial community study using DGGE [16]. In addition, Staphylococcus warneri has been reported to be present in salted and smoke fish products [26], suggesting its relationship with salinity. On the other hand, the Pseudomonas of Gammaproteobacteria was not observed in HS-fermented soy sauce under HHP [24], indicating the different influence of HHP on LS- and HS-fermented soy sauce. On the other hand, the lactic acid bacteria, such as Enterococcus and Lactobacillus, were both presented in LS-HP-treated soy sauce after 4-week fermentation, suggesting the increased diversity of lactic acid bacteria compared with other groups (Figure 3). Enterococcus and Lactobacillus existed together in soy sauce koji [27], but only one kind was observed in soy sauce products [16,17]. Our results suggest the significant bacterial change in LS-HP soy sauce, which is different from LS and HS groups (Figure 3). Therefore, we further identified the bacterial taxonomy and relative abundance in HS, LS, and LS-HP-treated soy sauce.
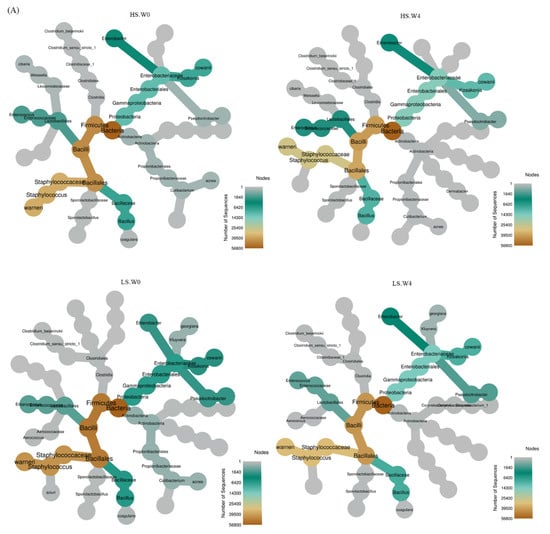
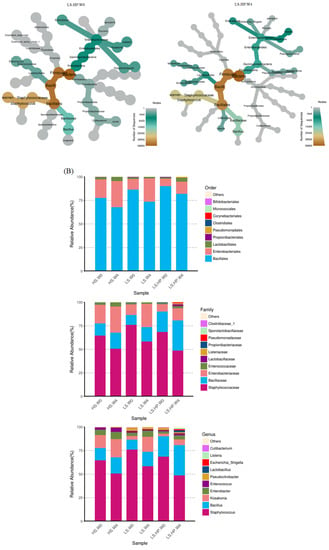
Figure 3.
(A) The heat trees obtained from analysis of bacteria microbiota in high-salt (HS), low-salt (LS), and low-salt high-pressure (LS-HP) soy sauce during fermentation. (B) The relative abundance (%) obtained from analysis of bacteria OTUs in high-saline (HS), low-saline (LS), and high-pressure-treated low-saline (LS-HP) soy sauce during fermentation.
3.3. Bacterial Abundance in HS, LS, and LS-HP-Treated Soy Sauce
Figure 3B are the diagrams of the relative abundance from taxonomic analysis of bacteria in HS, LS, and LS-HP-treated soy sauce in accordance with “order”, “family”, and “genus” taxa. This figure shows the top 10 bacteria in soy sauce discussed in this study. As shown in Figure 3B, the majority of the bacteria in soy sauce was Bacillales, Staphylococcaceae, and Staphylococcus in terms of order, family, and genus, respectively. Rather than a high-salt environment, the bacteria of Bacillales survived well in a low-salt soy sauce. In spite of being treated with high-pressure processing, the number of bacteria species in the LS-HP group did not go down, and increased on the contrary. This might be related with the spore-formation properties of these bacteria. The 300 MPa high-pressure treatment might have unfavorable influences on some bacteria that are not able to tolerate high pressure, therefore facilitating the dominant condition of spore-forming bacteria of Bacillales. From family and genus taxonomic points of view, the LS environment is favorable for bacteria belonging to Staphylococcaceae and Staphylococcus. In addition, the amount of these bacteria was decreasing while the second majority in soy sauce, Bacillus (of Bacillaceae), grew well during fermentation in all groups. Whereby we can see the high-pressure treatment did not inhibit the growth of bacteria in Bacillaceae. At the genus level, the diversity of bacteria in the high-pressure-treated LS-HP group greatly increased after 4 weeks fermentation comparing to the beginning (0 week), which was the highest among the other groups. The reason for this status might be that high pressure changed the original growth environment for dominant bacteria, elevating the possibility for other bacteria to grow during fermentation.
Staphylococcus, Bacillus, and Lactobacillus are the most common bacteria during soy sauce fermentation [16,24,28], which is consistent with our results (Figure 3B). In addition, increased Bacillus abundance was also observed in high-pressure-fermented HS soy sauce, whereas the Staphylococcus abundance was only slightly increased [22]. Other research reported the Bacillus abundance remained stable in HS soy sauce during 4 weeks [24]. Our results show the high bacterial diversity after LS-HP treatment, and this is similar with the diversity indexes. Despite the decreased whole bacterial abundance in the LS-HP group after 4 weeks, the bacterial diversity was increased probably due to the newly observed bacteria, such as Lactobacillus (Figure 3B). Other studies have reported that some Lactobacillus strains could survive and acclimate in high-pressure environments [29,30], which might explain the occurrence of Lactobacillus in the LS-HP group.
3.4. Fungal Diversity and Variation in HS, LS, and LS-HP-Treated Soil Sauce
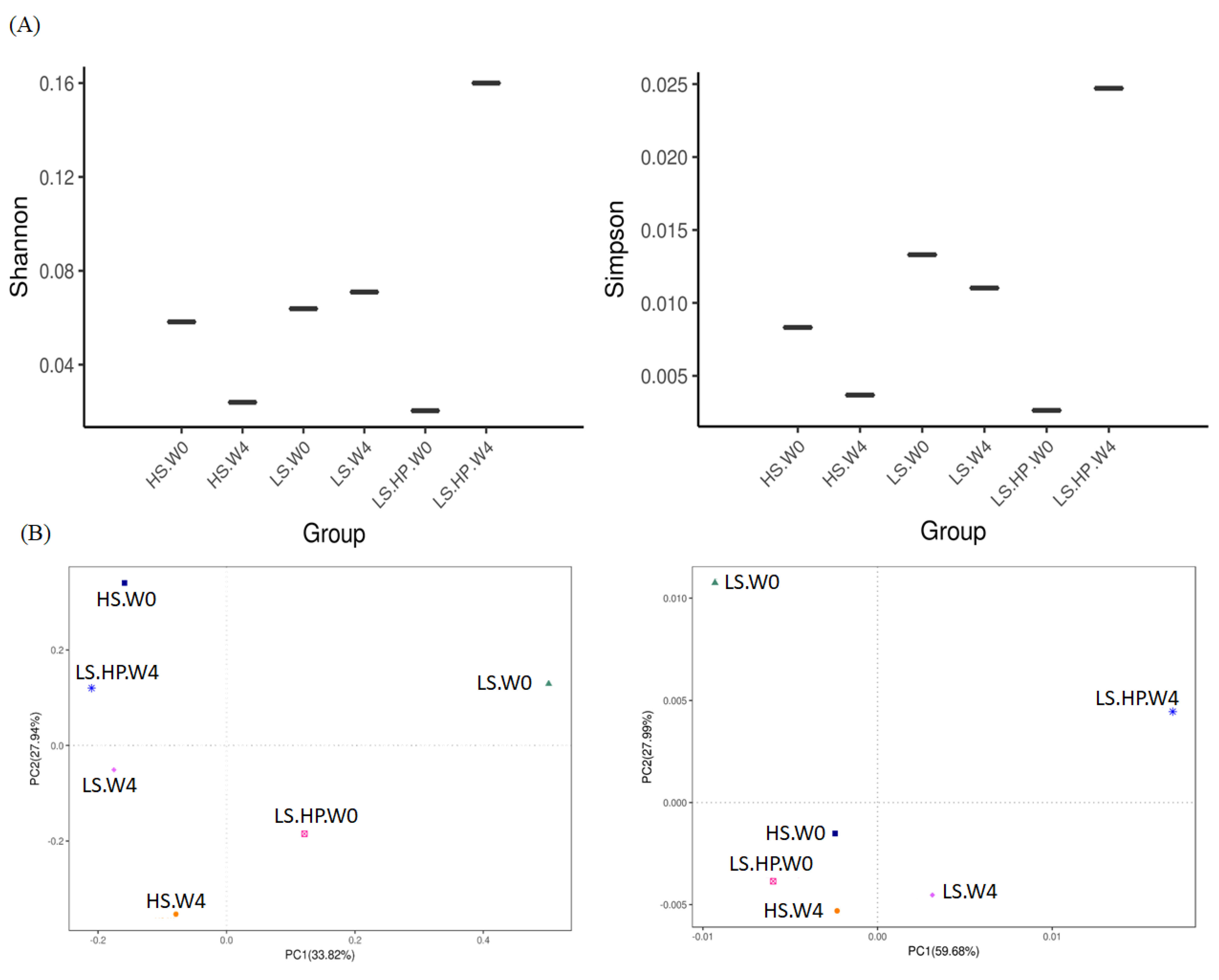
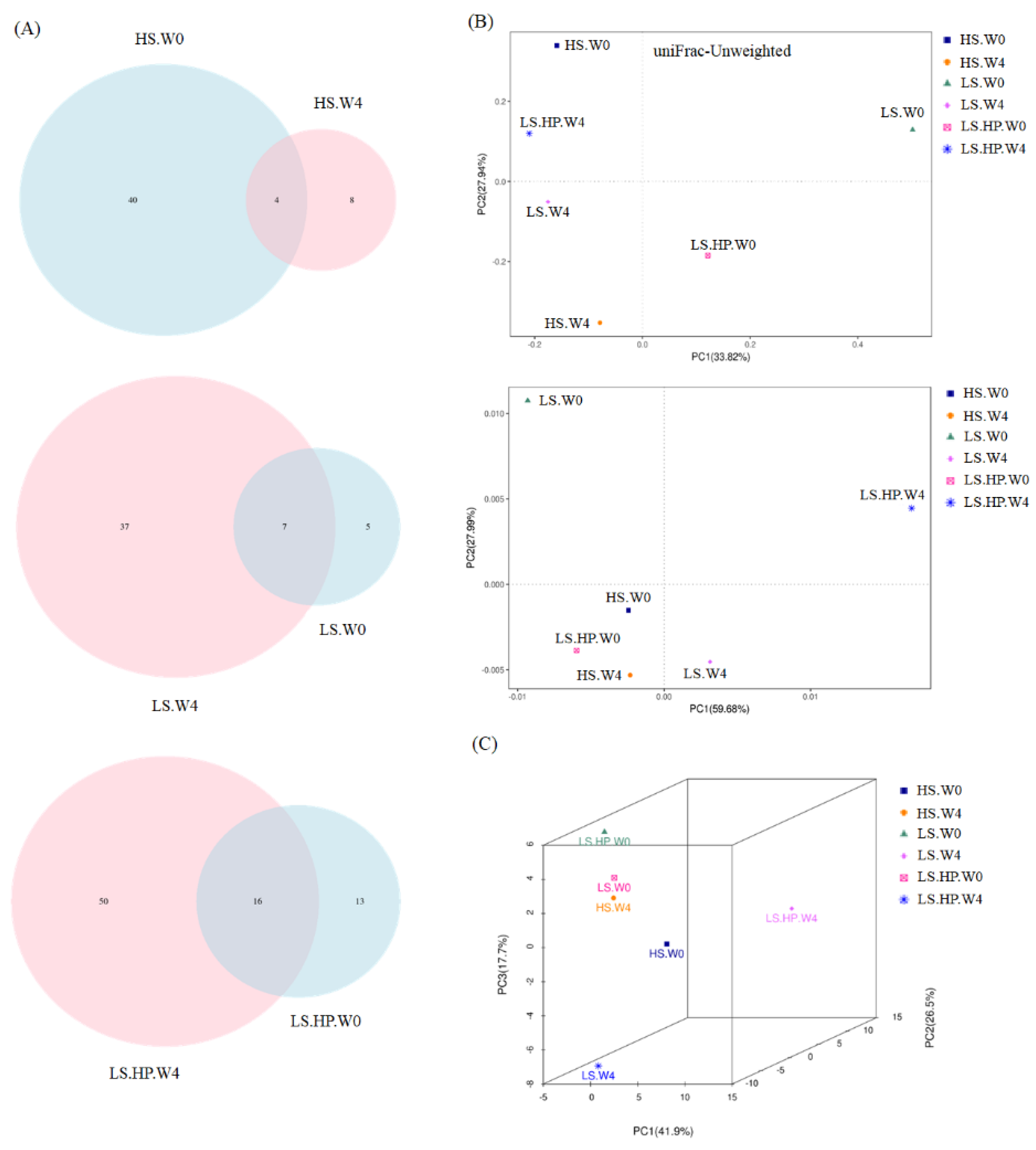
Figure 4 showed the Shannon and Simpson index of alpha-diversity, and the uniFrac-Unweighted and uniFrac-Weighted PCoA plots of beta-diversity obtained from analysis of fungal microbiota in high-salt (HS), low-salt (LS), and low-salt high-pressure (LS-HP) soy sauce during fermentation. To illustrate the similar and different OTU numbers in HS, LS, and LS-HPs groups, Figure 5A shows the Venn diagrams after cluster analysis. The fungal OTU number in HS-treated soy sauce was decreased from 44 to 12 (decreasing by 32) after 4-week fermentation, whereas that in the LS and LS-HP-treated one was increased by 32 and 37, respectively, after the same fermentation time. The results suggests that most fungal species could not survive in HS-treated soy sauce. The uniFrac PCA plots show that the difference between LS treatment in week 0 and LS-HP treatment in week 4 were much larger compared with other groups (Figure 5B).
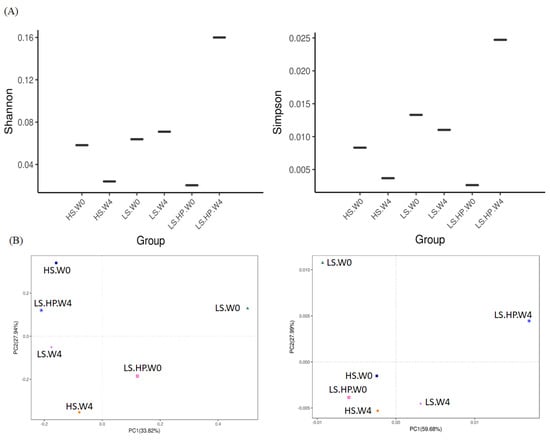
Figure 4.
(A) The Shannon and Simpson index of alpha-diversity, and (B) The uniFrac-Unweighted and uniFrac-Weighted PCoA plots of beta-diversity obtained from analysis of fungal microbiota in high-salt (HS), low-salt (LS), and low-salt high-pressure (LS-HP) soy sauce during fermentation.
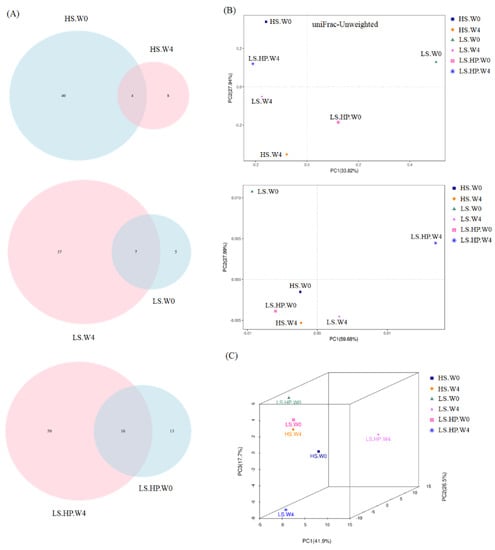
Figure 5.
(A) Venn diagrams of analysis for the fungal OTUs, (B) the unweighted and weighted uniFrac PCoA plots of beta-diversity obtained from analysis of fungal microbiota in high-salt (HS), low-salt (LS), and low-salt high-pressure (LS-HP) soy sauce during fermentation. (C) PCA assay for fungal microbiota in high-salt (HS), low-salt (LS), and low-salt high-pressure (LS-HP) soy sauce during fermentation.
The increase in fungal abundance in soy sauce after HS and HP fermentation has been suggested in recent study, whereas the fungal diversity was decreased [22]. However, other research showed an increase in fungal abundance and diversity in HS-fermented Cantonese soy sauce after 103 days [31], suggesting that different processing and fermentation times might affect the fungal community in HS-fermented soy sauce. Our results suggest LS and LS-HP groups increased the fungal OTU numbers compared with the HS one, indicating that the enhanced fungal diversity might be due to salinity change. In addition, the high fungal diversity after LS-HP treatment was also revealed in Figure 5C compared with the LS and HS groups, which is similar with our previous results about bacterial community.
3.5. Heat Map of Fungal Community in HS, LS, and LS-HP Soy Sauce
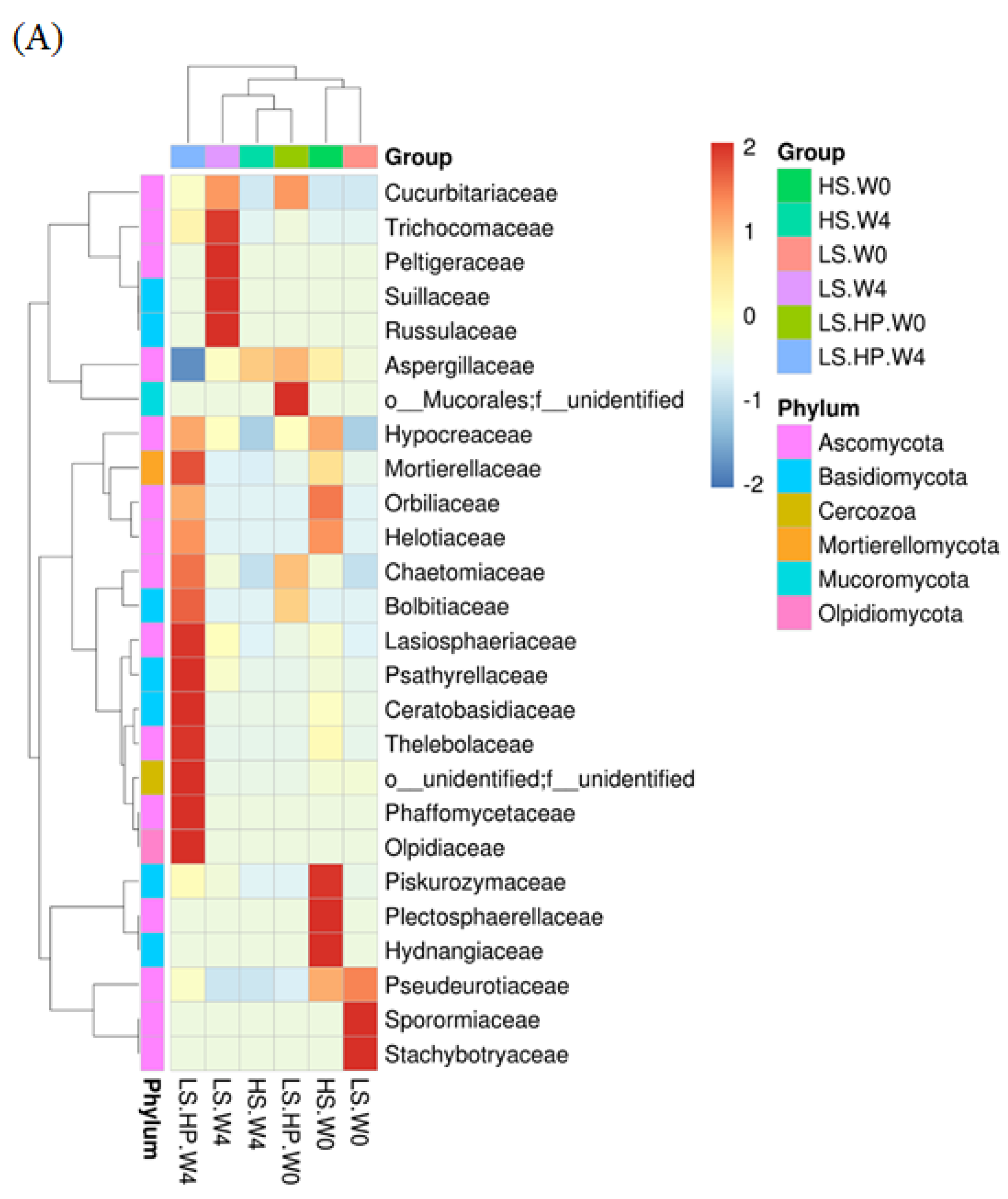
Figure 6A shows the heat map of fungal community in HS, LS, and LS-HP soy sauce. The x-axis indicates the different treatment groups, and the y-axis indicates the abundance of the top 26 fungi. The bar graph on the left panel illustrates the fungal classification based on the phylum taxa and is indicated by different colors. The tree diagrams show the similarity between different fungi and treatments. The abundance of fungi was illustrated by different colors, where red is the highest and blue is the lowest. The results show that HS and LS treatment at week 0 were similar and both have the high abundance of Pseudourotiaceae. After 4-week fermentation, a high abundance of Trichocomaceae, Peltigeraceae, Suillaceae, and Ressulaceae was observed in the LS group, suggesting that the growth of those fungi was not affected by salinity and grow normally in LS fermentation. However, the above fungi were not observed in the LS-HP group, which might have been due to the sensitivity to high pressure. In addition, LS-HP after 4-week fermentation was very different compared with the other groups, and showed the high abundance of Lasiosphaeriaceae, Psathyrellaceae, Ceratobasidiaceae, Thelebolaceae, Phaffomycetaceae, and Olpidiaceae.
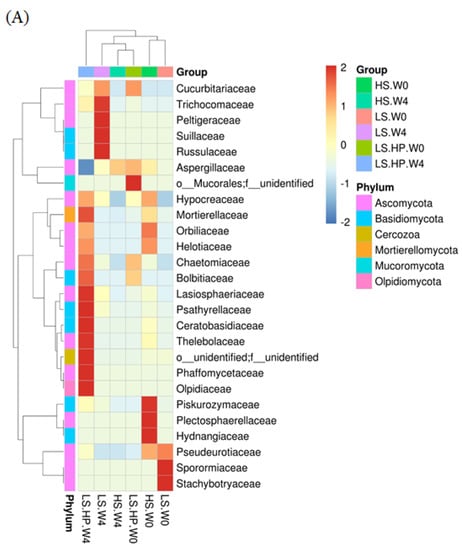
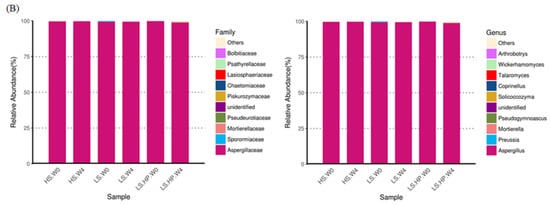
Figure 6.
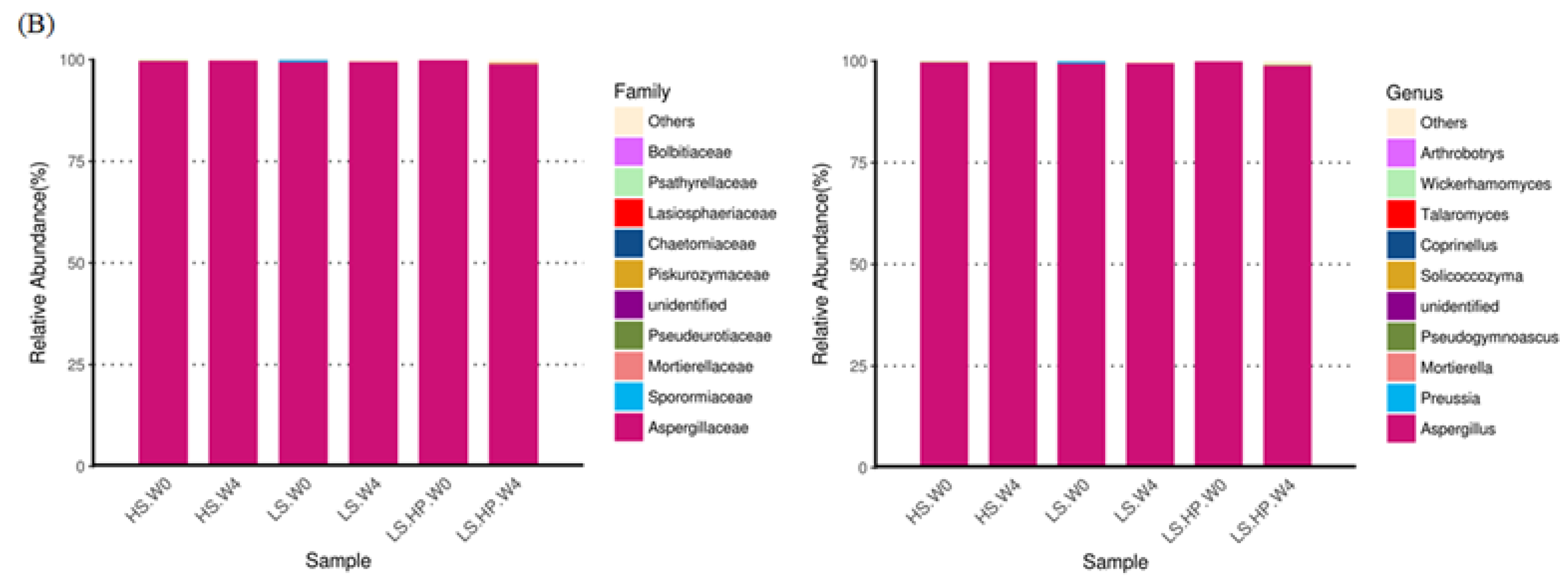
(A) The heat map obtained from analysis of fungal microbiota in high-salt (HS), low-salt (LS), and low-salt high-pressure (LS-HP) soy sauce during fermentation. (B) The relative abundance (%) obtained from analysis of fungal OTUs in high-salt (HS), low-salt (LS), and low-salt high-pressure (LS-HP) fermentation soy sauce.
In soy sauce koji, the dominant fungi were A. oryzae and Saccharomycopsis fibuligera, which might decrease following fermentation where other fungi would grow [17,32]. After 4-week fermentation, the major fungi in HS-fermented soy sauce was A. oryzae with a low abundance of other strains (Figure 6B), which is similar to other studies [22,27,28,30]. Our results further suggest that the significant difference in fungal diversity resulted from high-pressure treatment (Figure 6B). The growth of A. oryzae was significantly inhibited under high-pressure treatment [22,33], which is in consistent with our results. However, the influence of high pressure on fungi might be varied depending on their genera [22,33]. The results from our study suggest the different fungal genera occurred in HS, LS, and LS-HP-treated soy sauce, implying their sensitivity to salt and pressure.
4. Conclusions
Our study demonstrates that microbial community change in soy sauce under different fermentation conditions including HS, LS, and LS-HP after 4 weeks. The results showed different bacterial and fungal community especially in LS-HP-fermented soy sauce, suggesting that salinity and pressure would largely affect microbiota in soy sauce. Despite the decreased bacterial abundance, bacterial diversity was increased in LS-HP-treated soy sauce. Different bacteria, such as Lactobacillus, was especially observed in the LS-HP group compared with the HS and LS ones. In addition, fungal diversity was also increased in LS-HP-fermented soy sauce and different fungi were specifically observed accompanied by a lower abundance of Aspergillus. The present study suggests the availability of low levels of salt and high pressure on soy sauce production provides information about the bacterial and fungal community change under different methods.
Author Contributions
Conceptualization, Y.-C.S. and B.-H.L.; methodology, C.-Y.L.; software, S.-C.W.; formal analysis, S.-C.W.; investigation, S.-C.W. and B.-H.L.; resources, S.-C.W.; data curation, Y.-C.S.; writing—original draft preparation, Y.-C.S.; writing—review and editing, S.-C.W.; project administration, B.-H.L. All authors have read and agreed to the published version of the manuscript.
Funding
Acknowledgement for this research work and subsidiary spending were supported by the Ministry of Science and Technology (MOST109-2314-B-415-001-) (Taiwan, R.O.C.).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Wicklow, D.T.; McAlpin, C.E.; Yeoh, Q.L. Diversity of Aspergillus oryzae genotypes (RFLP) isolated from traditional soy sauce production within Malaysia and Southeast Asia. Mycoscience 2007, 48, 373–380. [Google Scholar] [CrossRef]
- Luh, B.S. Industrial production of Soy sauce. J. Ind. Microbiol. 1995, 14, 467–471. [Google Scholar] [CrossRef]
- Yang, L.; Yang, H.L.; Tu, Z.C.; Wang, X.L. High-throughput sequencing of microbial community diversity and dynamics during douchi fermentation. PLoS ONE 2016, 11, e0168166. [Google Scholar] [CrossRef] [Green Version]
- Beland, F.A.; Benson, R.W.; Mellick, P.W.; Kovatch, R.M.; Roberts, D.W.; Fang, J.L.; Doerge, D.R. Effect of ethanol on the tumorigenicity of urethane (ethyl carbamate) in B6C3F1 mice. Food Chem. Toxicol. 2005, 43, 1–19. [Google Scholar] [CrossRef]
- Riachi, L.G.; Santos, A.; Moreira, R.F.; De Maria, C.A. A review of ethyl carbamate and polycyclic aromatic hydrocarbon contamination risk in cachaça and other Brazilian sugarcane spirits. Food Chem. 2014, 149, 159–169. [Google Scholar] [CrossRef]
- Chen, D.W.; Ren, Y.P.; Zhong, Q.D.; Shao, Y.; Zhao, Y.F.; Wu, Y.N. Ethyl carbamate in alcoholic beverages from China: Levels, dietary intake, and risk assessment. Food Control 2017, 72, 283–288. [Google Scholar] [CrossRef]
- Hasnip, S.; Crews, C.; Potter, N.; Christy, J.; Chan, D.; Bondu, T.; Matthews, W.; Walters, B.; Patel, K. Survey of ethyl carbamate in fermented foods sold in the United Kingdom in 2004. J. Agric. Food Chem. 2007, 55, 2755–2759. [Google Scholar] [CrossRef]
- Jiao, Z.; Dong, Y.; Chen, Q. Ethyl carbamate in fermented beverages: Presence, analytical chemistry, formation mechanism, and mitigation proposals. Compr. Rev. Food Sci. Food Saf. 2014, 13, 611–626. [Google Scholar] [CrossRef]
- Ryu, D.; Choi, B.; Kim, E.; Park, S.; Paeng, H.; Kim, C.I.; Lee, J.Y.; Yoon, H.J.; Koh, E. Determination of ethyl carbamate in alcoholic beverages and fermented foods sold in Korea. Toxicol. Res. 2015, 31, 289–297. [Google Scholar] [CrossRef]
- Weber, J.V.; Sharypov, V.I. Ethyl carbamate in foods and beverages—A review. In Climate Change, Intercropping, Pest Control and Beneficial Microorganisms; Lichtfouse, E., Ed.; Springer: Dordrecht, The Netherlands, 2009; pp. 429–452. [Google Scholar]
- Wu, P.G.; Pan, X.D.; Wang, L.Y.; Shen, X.H.; Yang, D.J. A survey of ethyl carbamate in fermented foods and beverages from Zhejiang, China. Food Control 2012, 23, 286–288. [Google Scholar] [CrossRef]
- Zhao, X.R.; Du, G.C.; Zou, H.J.; Fu, J.W.; Zhou, J.W.; Chen, J. Progress in preventing the accumulation of ethyl carbamate in alcoholic beverages. Trends Food Sci. Technol. 2013, 32, 97–107. [Google Scholar] [CrossRef]
- Matsudo, T.; Aoki, T.; Abe, K.; Fukuta, N.; Higuchi, T.; Sasaki, M.; Uchida, K. Determination of ethyl carbamate in soy-sauce and its possible precursor. J. Agric. Food Chem. 1993, 41, 352–356. [Google Scholar] [CrossRef]
- Zhang, J.R.; Fang, F.; Chen, J.; Du, G.C. The arginine deiminase pathway of koji bacteria is involved in ethyl carbamate precursor production in soy sauce. FEMS Microbiol. Lett. 2014, 358, 91–97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Betts, G.D.; Linton, P.; Betteridge, R.J. Food spoilage yeasts: Effects of pH, NaCl and temperature on growth. Food Control 1999, 10, 27–33. [Google Scholar] [CrossRef]
- Tanaka, Y.; Watanabe, J.; Mogi, Y. Monitoring of the microbial communities involved in the soy sauce manufacturing process by PCR-denaturing gradient gel electrophoresis. Food Microbiol. 2012, 31, 100–106. [Google Scholar] [CrossRef]
- Wei, Q.; Wang, H.; Chen, Z.; Lv, Z.; Xie, Y.; Lu, F. Profiling of dynamic changes in the microbial community during the soy sauce fermentation process. Appl. Microbiol. Biotechnol. 2013, 97, 9111–9119. [Google Scholar] [CrossRef]
- Fang, F.; Zhang, J.; Zhou, J.; Zhou, Z.; Li, T.; Lu, L.; Zeng, W.; Du, G.; Chen, J. Accumulation of citrulline by microbial arginine metabolism during alcoholic fermentation of soy sauce. J. Agric. Food Chem. 2018, 66, 2108–2113. [Google Scholar] [CrossRef]
- Yamamoto, K. Food processing by high hydrostatic pressure. Biosci. Biotechnol. Biochem. 2017, 81, 672–679. [Google Scholar] [CrossRef] [Green Version]
- Del Arbol, J.T.; Pulido, R.P.; La Storia, A.; Burgos, M.J.G.; Lucas, R.; Ercolini, D.; Galvez, A. Changes in microbial diversity of brined green asparagus upon treatment with high hydrostatic pressure. Int. J. Food Microbiol. 2016, 216, 1–8. [Google Scholar] [CrossRef]
- Pulido, R.P.; Burgos, M.J.G.; Galvez, A.; Lucas, R. Changes in bacterial diversity of refrigerated mango pulp before and after treatment by high hydrostatic pressure. LWT Food Sci. Technol. 2017, 78, 289–295. [Google Scholar] [CrossRef]
- Liang, R.; Huang, J.; Wu, X.M.; Xu, Y.; Fan, J.; Wu, C.D.; Jin, Y.; Zhou, R. Characterizing the metabolites and the microbial communities of the soy sauce mash affected by temperature and hydrostatic pressure. Food Res. Int. 2019, 123, 801–808. [Google Scholar] [CrossRef] [PubMed]
- Lozupone, C.A.; Hamady, M.; Kelley, S.T.; Knight, R. Quantitative and qualitative β diversity measures lead to different insights into factors that structure microbial communities. Appl. Environ. Microbiol. 2007, 73, 1576–1585. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Qian, M.; Shen, Y.; Qin, X.; Huang, H.; Yang, H.; He, Y.; Bai, W. A high-throughput sequencing approach to the preliminary analysis of bacterial communities associated with changes in amino acid nitrogen, organic acid and reducing sugar contents during soy sauce fermentation. Food Chem. 2021, 349, 129131. [Google Scholar] [CrossRef] [PubMed]
- Foster, Z.S.; Sharpton, T.J.; Grunwald, N.J. Metacoder: An R package for visualization and manipulation of community taxonomic diversity data. PLoS Comput. Biol. 2017, 13, e1005404. [Google Scholar] [CrossRef] [Green Version]
- Sergelidis, D.; Abrahim, A.; Papadopoulos, T.; Soultos, N.; Martziou, E.; Koulourida, V.; Govaris, A.; Pexara, A.; Zdragas, A.; Papa, A. Isolation of methicillin-resistant Staphylococcus spp. from ready-to-eat fish products. Lett. Appl. Microbiol. 2014, 59, 500–506. [Google Scholar] [CrossRef]
- Yan, Y.Z.; Qian, Y.L.; Ji, F.D.; Chen, J.Y.; Han, B.Z. Microbial composition during Chinese soy sauce koji-making based on culture dependent and independent methods. Food Microbiol. 2013, 34, 189–195. [Google Scholar] [CrossRef]
- Han, D.M.; Chun, B.H.; Feng, T.; Kim, H.M.; Jeon, C.O. Dynamics of microbial communities and metabolites in ganjang, a traditional Korean fermented soy sauce, during fermentation. Food Microbiol. 2020, 92, 103591. [Google Scholar] [CrossRef]
- Mota, M.J.; Lopes, R.P.; Sousa, S.; Gomes, A.M.; Delgadillo, I.; Saraiva, J.A. Lactobacillus reuteri growth and fermentation under high pressure towards the production of 1,3-propanediol. Food Res. Int. 2018, 113, 424–432. [Google Scholar] [CrossRef]
- Yang, D.; Zhang, Y.; Zhao, L.; Wang, Y.T.; Rao, L.; Liao, X.J. Pressure-resistant acclimation of lactic acid bacteria from a natural fermentation product using high pressure. Innov. Food Sci. Emerg. Technol. 2021, 69, 102660. [Google Scholar] [CrossRef]
- Qi, Q.; Huang, J.; Zhou, R.Q.; Yang, M.L.; Zhang, L.; Peng, C.; Jin, Y.; Wu, C.D. Characterizing microbial community and metabolites of Cantonese soy sauce. Food Biosci. 2021, 40, 100872. [Google Scholar] [CrossRef]
- Wei, C.L.; Chao, S.H.; Tsai, W.B.; Lee, P.S.; Tsau, N.H.; Chen, J.S.; Lai, W.L.; Tu, J.C.Y.; Tsai, Y.C. Analysis of bacterial diversity during the fermentation of inyu, a high-temperature fermented soy sauce, using nested PCR-denaturing gradient gel electrophoresis and the plate count method. Food Microbiol. 2013, 33, 252–261. [Google Scholar] [CrossRef] [PubMed]
- Del Arbol, J.T.; Pulido, R.P.; La Storia, A.; Burgos, M.J.G.; Lucas, R.; Ercolini, D.; Galvez, A. Microbial diversity in pitted sweet cherries (Prunus avium L.) as affected by high-hydrostatic pressure treatment. Food Res. Int. 2016, 89, 790–796. [Google Scholar] [CrossRef] [PubMed]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).