1. Introduction
Pigments have been applied as coloring agents since ancient times to improve the visual appearance of several products. Color is a key feature that stimulates consumers to buy certain products thus pigments have been widely explored by various industries, such as food, cosmetics, textile, pharmaceutical, and tanneries, for the development of different colorful products [
1].
Pigments can be classified as organic or inorganic, both classes being subdivided into natural or synthetic [
2,
3]. In the last decade, the replacement of synthetic pigments with natural ones has gained attention due to the increasing consciousness of consumers about ecological preservation and safety, sustainability, and health issues. Thus, the demand for new biological, non-toxic, and biocompatible pigments derived from natural, cleaner, and more ecological sources has been increasing [
4]. Furthermore, natural pigments may additionally provide valuable characteristics to the final products. They may present some useful biological properties for different industries, such as free radical protection, and antioxidant potential, among others [
5].
Natural pigments were primarily obtained from living organisms such as plants and insects [
3]. Nevertheless, to avoid the seasonal and ethical issues frequently associated wiith these organisms, microbial fermentation has been considered a very promising alternative to produce natural pigments, arousing industrial interest. Microorganisms are generally more accessible, can grow rapidly, and have the potential to be genetically modified, which consequently can provide better yields and productivity [
6,
7].
Among the different microorganisms, filamentous fungi have been reported as the main pigment producers due to their versatility in producing pigments of different colors [
7,
8,
9,
10]. Although there are several fungal pigments reported in the literature, their practical use is conditioned by regulatory approval criteria related to their toxicity and stability [
11].
Penicillium species have been identified as potential pigment producers. Their pigments are homologous to those produced by
Monascus (one of the most reported pigments producers), with the advantage of not producing the mycotoxin citrinin, known to be nephrotoxic, hepatotoxic, and possibly carcinogenic to humans and animals [
12,
13]. The potential of
Penicillium brevicompactum to produce pigments by submerged fermentation using a synthetic culture medium containing lactose was recently demonstrated [
14].
The use of agro-industrial byproducts, or residues, in fermentation processes has been widely studied as an alternative strategy to reduce their environmental impact and promote a circular economy. Additionally, the valorization of the residues/byproducts by fermentation generally lead to the production of value-added biocompounds with great commercial interest. Hence, the development of a fermentative process for pigment production using industrial byproducts, such as cheese whey and corn steep liquor, as substrates for microbial growth is of great relevance. Cheese whey (CW) is the main byproduct obtained from the dairy industry, with an estimated global production of approximately 200 million tons [
15]. Although being recognized as a suitable source of functional and bioactive compounds, around 50% of the CW produced worldwide is still not valorized, being generally discharged without appropriate treatment, thus causing severe environmental issues [
16,
17]. Nevertheless, it contains a high percentage of lactose which can be an important carbon source for microbial growth. Corn steep liquor (CSL) is a byproduct obtained from the corn-wet milling process to produce cornstarch. In the United States, one of the major producers, around 25 million tons of cornstarch are produced annually, generating about 16 million m
3 of CSL [
18]. CSL contains important amounts of free amino acids, sugars, and nitrogen, making it an interesting fermentation substrate for microbial growth [
19]. The successful use of CW and CSL in the preparation of alternative culture media for the production of different pigments was already reported for several
Monascus species [
20,
21,
22].
This work reports for the first time the development of a suitable bioprocess for pigment production by P. brevicompactum, using alternative low-cost media containing industrial byproducts, namely cheese whey, and corn steep liquor. Culture media composed of byproducts offer recognized economic advantages and, therefore, can result in a more attractive approach for pigment production at the industrial level. Additionally, different strategies for pigment production, such as solid-state fermentation (described as closer to the native growth conditions) and submerged fermentation with free and immobilized mycelium (to facilitate the biomass and product separation/recovery), were explored and compared in terms of production effectiveness.
2. Materials and Methods
2.1. Microorganism
P. brevicompactum (MUM 02.07) was obtained from the Mycology collection of the University of Minho (MUM). Stock cultures were maintained at room temperature (around 25 °C) by preparing spore suspensions in a semi-solid agar medium (2 g/L). For the inoculum preparation, stock cultures were sub-cultured on Petri dishes containing malt extract agar (MEA) medium (% w/v): malt extract (2), glucose (2), peptone (0.1), and agar (2); and grown for 7 days at 25 °C in the absence of light.
2.2. Industrial Byproducts
Cheese whey powder (CW), composed of (%
w/w): 58.5 lactose, 12.6 protein, less than 0.2 fat, and 1.2 moisture, was kindly provided by Lactogal Produtos Alimentares S.A., Modivas, Portugal. Corn steep liquor (CSL) with a composition of (%
w/v): 7.5 sugars and 0.5 proteins [
23] was supplied by COPAM (Companhia Portuguesa de Amidos, S.A.), São João da Talha, Portugal.
2.3. Inert Support
Cubes (0.125 cm
3) of commercial nylon sponge (Vileda Ultra Fresh
®, Freudenberg Home and Cleaning Solutions GmbH, Weinheim, Germany) were used as inert supports in both submerged and solid-state fermentations. The nylon sponge cubes were pre-treated by boiling for 15 min, washed with distilled water, and then dried at 60 °C. Prior to use, the cubes were autoclaved at 121 °C for 15 min [
24].
2.4. Inoculum Preparation
Inocula for fermentations were prepared by adding 1–2 mL of a sterile saline solution (0.85% (
w/v) NaCl containing 0.01% (
w/v) Tween 80) to fully sporulated agar plate cultures of
P. brevicompactum [
25]. The spores were then scraped from the agar plates under aseptic conditions, and the conidia suspension was used as the inoculum. Conidia density (10
6 conidia/mL) was determined using a Neubauer chamber.
2.5. Pigments Production
Four different culture media were prepared (g/L): medium A—peptone (8), yeast extract (8), KH
2PO
4 (2), Na
2HPO
4.12H
2O (8), MgSO
4.6H
2O (0.25), and lactose (20); medium B—CW (34.6); medium C—CSL (12.55); medium D—CSL (8), and CW (34.6). The optimal CSL concentration in medium C was previously defined as 12.55 g/L to reduce its color contribution to the culture medium. The different culture media (A, B, C, and D) were used for both submerged fermentation (free (SmF), immobilized mycelium (SmFi)), and solid-state fermentation (SSF). The fermentations were performed in an orbital shaker for 12 days at 23 °C and 150 rpm. These experimental conditions were determined in previous work and are optimized for pigment production by
P. brevicompactum cultivated in a synthetic fermentation medium [
14]. The experiments were performed in triplicate, using 250 mL cotton-plugged Erlenmeyer flasks containing an initial density of 10
6 conidia/mL. Samples were collected at 0, 3, 5, 7, 9, and 12 days, in aseptic conditions, for all the submerged fermentation processes. For SSF samples collected at the beginning and at the end of the fermentation time were evaluated.
2.5.1. Submerged Fermentation
Submerged fermentation was performed using free (SmF) or immobilized mycelium (SmFi) and 50 mL of each culture medium (A, B, C, and D). For (SmFi), approximately 20 cubes of inert support (corresponding to 0.8 g of nylon sponge) were used.
2.5.2. Solid-State Fermentation (SSF)
SSF was performed using nylon sponge cubes as an inert support. To achieve better results, a preliminary test was carried out to determine the most suitable moisture content % (
w/w). For this purpose, the production of pigments was evaluated when 3 g of nylon sponge cubes (75 cubes), and different volumes of culture medium A were used. The amount of support (3 g) was selected according to the volume occupied by the cubes in the flask. Moistures contents of 75, 80, 85, and 90% (
w/w) were assessed. The moisture content (wet basis) was calculated according to Ahmad and Munaim [
26] using Equation (1).
After selecting the most suitable moisture content, all the culture media (A, B, C, and D) were screened for pigment production under SSF conditions using 3 g of nylon sponge cubes.
2.6. Biomass Reuse under Submerged Fermentation Conditions
To evaluate if biomass reuse can be a suitable strategy to increase pigment production, two types of fermentation were carried out, namely SmF and SmFi.
For each type of fermentation, the experiments were performed in triplicate using 250 mL cotton-plugged Erlenmeyer flasks containing 50 mL of culture medium D and an initial density of 106 conidia/mL.
The study of biomass reuse was divided into 3 cycles of fermentation of 12 days. After 12 and 24 days of fermentation (first and second cycles, respectively), the culture medium was removed with the aid of a sterile net, and replaced by 50 mL of fresh medium D.
Simultaneously, for each condition (SmF and SmFi), control fermentations were performed in parallel for 36 days without replacing the fermentation medium. The fermentations were performed in an orbital shaker at 23 °C and 150 rpm. In the three cycles, samples were aseptically collected on days 0, 3, 5, 7, 9, and 12. In the control fermentation, samples were taken on days 0, 3, 5, 7, 10, 12, 14, 24, 26, and 36.
For each fermentation condition, pigment recovery was performed as described in the section below.
2.7. Pigments Recovery
For SmF and SmFi conditions, the culture media containing the produced pigments were recovered through vacuum filtration using a 0.45 μm Whatman filter and freeze-dried for further analysis. Additionally, for SmFi, the support containing the immobilized biomass was recovered for pigment extraction using a solution with 95:5% (
v/v) ethanol and water [
27]. The mixture was kept on a rotary shaker at 150 rpm for 24 h at room temperature. The support was then squeezed with the aid of a syringe, and the liquid fraction was filtered through a 0.45 μm Whatman filter. The ethanol was finally evaporated at 60 °C.
For SSF, pigments were also extracted from supports using a solution of 95:5% (
v/v) ethanol and water, as described for the SmFi. In this case, all the content of the culture flasks was soaked in ethanol. The volume of added ethanol corresponded to three times the volume of the culture medium initially used [
27].
2.8. Pigments Analysis
The samples collected during the fermentations were spectrophotometrically analyzed to monitor the production of pigments. The samples were centrifuged prior to measuring the absorbance at 400, 470, and 500 nm the corresponding wavelengths of the yellow, orange, and red regions of the visible spectrum, respectively [
28,
29].
To compare the different fermentation conditions for pigments production, freeze-dried media (collected at the end of SmF and SmFi) and dried ethanolic extracts (obtained from the SmFi and SSF immobilization supports) were dissolved in 2.5 mL of distilled water, and the absorbance was measured at the same wavelengths.
To determine the best conditions for pigment production, the sum of the three absorbances (at 400, 470, and 500 nm) was considered.
The fermentation media and the ethanolic extracts containing the pigments were also qualitatively analyzed by Thin Layer Chromatography (TLC). For each fermentation condition, 3 µL of a pigment solution with a concentration of 100 g/L was loaded on a silica plate (Macherey-Nagel, DC-Fertigfolien ALUGRAM SIL G/UV254, Macherey-Nagel GmbH & Co., Düren, Germany). Pigments were separated using a mixture of 50:50% (
v/v) water and ethanol as eluent [
14]. The plates were exposed to UV light (254 nm and 366 nm) for spot visualization.
2.9. Lactose Quantification
Lactose concentration in the fermentation media was determined by HPLC analysis using a Jasco chromatograph equipped with a refractive index detector (K-2300, Knauer) and a Prevail Carbohydrate ES column (5 μm particle size, 250 mm length × 4.6 mm internal diameter, Alltech, Grace Davison Discovery Sciences, Bannockburn, IL, USA) fitted with a pre-column using the same stationary phase.
A mixture of acetonitrile and water (70:30 %(
v/v)) pumped at 0.9 mL/min was used as the mobile phase. The injection volume was defined as 20 μL [
14]. Standards of lactose in the range of 1–15 g/L were used to previously prepare a calibration curve.
The samples collected during the submerged fermentations (SmF and SmFi) were centrifuged, filtered through a 0.45 μm sterile syringe filter, and analyzed to monitor the consumption of lactose during the fermentation time.
2.10. Statistical Analysis
All the experiments were performed in triplicate, and the values are presented as the means and respective standard deviations. One-way and two-way ANOVA tests were performed using GraphPad Prism 6.0 software (GraphPad Software, San Diego, CA, USA) to estimate significant differences among samples with a confidence interval of 95%.
3. Results and Discussion
During a screening study aiming to identify novel fungal producers of β-galactosidase, Silvério et al. [
30] observed for the first time the ability of
P. brevicompactum to synthesize soluble extracellular pigments. Later, Fonseca et al. [
14] proved the potential of
P. brevicompactum to successfully produce a mixture of yellow, orange, and red pigments, under SmF conditions using a synthetic culture medium (medium A) that combines lactose with peptone and yeast extract. It was also demonstrated that both the carbon and the nitrogen sources are essential for cellular metabolism, directly affecting the fungal growth and the type of synthesized pigments [
14].
CW was selected due to its high lactose content (58.5%
w/w). The amount of CW was established to maintain a final lactose concentration of 20 g/L, reported by Fonseca et al. [
14] as the optimal lactose concentration for pigment production. In this work, CW was used as both a culture medium (medium B) and as a medium supplement (medium D).
To evaluate the effect of an alternative nitrogen source on pigment production, CSL was used (media C and D) since it has been widely reported as a relevant source of this element [
31]. As an alternative culture medium (medium C), CSL was used in a final concentration of 12.55 g/L. Since CSL is a dark brown viscous liquid, this concentration was previously defined as the most suitable for the culture medium without significantly altering its color, thus minimizing CSL interference in the absorbance measurements. As a supplement to the culture medium (medium D), CSL was used in the same concentration (8 g/L) reported by Fonseca et al. [
14] for the synthetic medium containing yeast extract and peptone.
3.1. Pigments Production under SmF
As reported before by Fonseca et al. [
14],
P. brevicompactum showed interesting productivity under SmF conditions. SmF is widely used both at laboratory and industrial-scale processes for the production of several valuable bioproducts. Thus, our first attempt was to investigate pigment production using the aforementioned alternative culture media under SmF conditions.
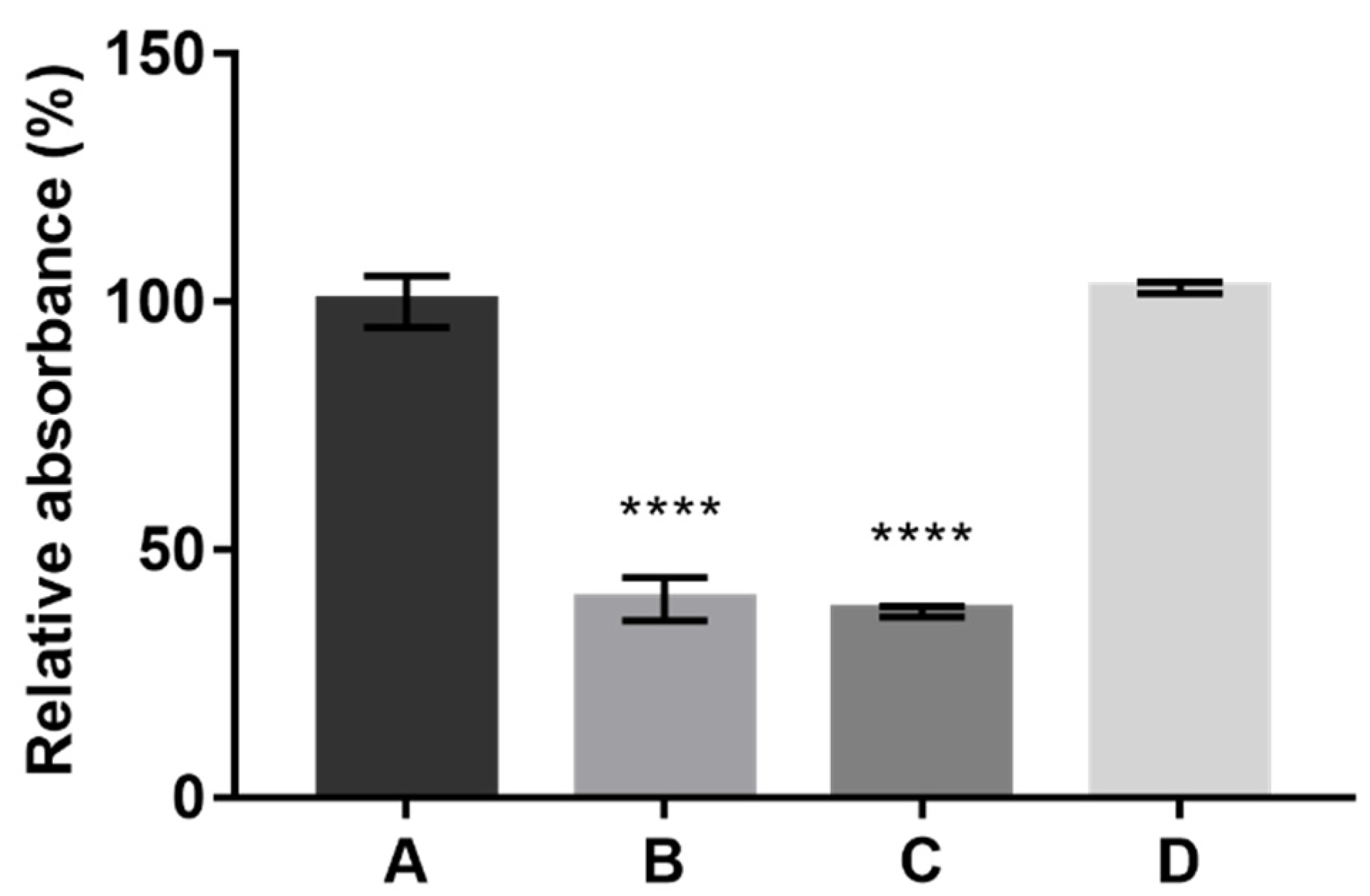
The results obtained for each culture media tested were expressed in relative absorbance (%) and statistically compared with the reference synthetic medium A (
Figure 1).
Figure 1 shows that, under SmF conditions, similar production of pigments was obtained using either the standard synthetic culture medium A or the alternative culture medium D composed of CW and CSL. Interestingly, both alternative culture media B and C, containing individually each one of the byproducts, led to the lowest pigments production. Previously, lactose was identified as the preferred sugar to produce pigments by
P. brevicompactum [
14]. However, according to the literature, the most common sugars in CSL are glucose and fructose [
32]. The concentration of these sugars in the alternative culture media C (12.55 g/L CSL) and D (8 g/L CSL) is very low since CSL generally contains only 2–3% (
w/v) of glucose and fructose [
32]. Furthermore, the HPLC analysis revealed a fast metabolization of CSL monosaccharides by
P. brevicompactum (the vestigial peaks corresponding to the monosaccharides from CSL were not present in the chromatograms after 3 days of fermentation). Thus, the very low concentration of monosaccharides and the absence of lactose in CSL composition probably led to lower production of pigments in culture medium C. Similarly, the lack of a nitrogen source in culture medium B seemed to reduce pigments production by the fungus.
The interesting results obtained for culture medium D indicate that CSL has a positive effect on pigment production when used in combination with CW, even at a lower concentration when compared to medium A (8 g/L of CSL in medium D versus 8 g/L of peptone + 8 g/L of yeast extract in medium A). The nitrogen from CSL seemed to work together with the lactose from the CW, enhancing pigments production, as culture medium D led to the same productivity as the reference synthetic medium A. Costa et al. [
22] also demonstrated that CW can be used as a carbon source for the growth, and consequent production of pigments by the filamentous fungi
Monascus ruber. Silbir and Goksungur [
33], reported the potential of CSL to be used by the filamentous fungus
Monascus purpureus as an alternative nitrogen source to monosodium glutamate, under SmF conditions.
After 12 days of fermentation, it was observed that in the culture media A and D lactose was completely or almost completely consumed (<1g/L), respectively, while for medium B, the concentration was higher than 8 g/L. In the work performed by Gmoser et al. [
34], aiming the production of carotenoids by
Neurospora sp., it was suggested that the use of a suitable carbon source could be a key factor to lead the metabolic pathway toward the acetyl-CoA pool, known to be the most important precursor for pigments production, rather than the cell growth pathways. Interestingly, we observed that the total, or almost total, consumption of lactose led to higher pigment production. Furthermore, an association between the timing determined as the starting point of lactose consumption and the appearance of color in the fermentation broth was observed (3 days of fermentation for culture medium A and 5 days of fermentation for the others). This fact led us to assume that the use of this carbon source by the fungus may be favoring or activating secondary metabolic pathways and consequently promoting pigments production.
3.2. Pigments Production by SmFi
The immobilization of mycelium is a common strategy used in the synthesis of secondary metabolites. Immobilization influences both diffusion properties of molecules through the support and the physiological behavior of immobilized cells, resulting in significant variations in the metabolism of nutrients and the production of metabolites [
35]. The immobilization of filamentous fungi is a relatively simple procedure facilitated by the ability of this kind of microorganisms to adhere to the surfaces of several materials. Fungal immobilization can bring some interesting advantages to the fermentation process. It eases the recovery and further reuse of the cells, which consequently may simplify the recovery of the bioproducts and increase the production and the purification yields [
36]. In a study conducted by Domínguez-Espinosa and Webb [
37], the immobilization of
Monascus on an inert support (foam) was tested under SmF conditions, resulting in enhanced production of red pigments, compared with SmF using the free mycelium.
To investigate whether biomass immobilization could influence pigment production, we repeated the previous conditions described for SmF but with the mycelium immobilized on a nylon sponge support (SmFi). The porosity of this material provides the fungus with an increased surface area (internal and superficial) to grow and have access to the culture medium nutrients [
24]. The amount of support used (20 pieces) was previously chosen based on the amount of support that could be used and still be completely submerged in the total volume of the culture medium.
Remarkably, after 12 days of fermentation, the biomass grown attached to the nylon cubes was colorful (
Figure 2), indicating that some of the pigments produced were not released to the fermentation medium (note that the biomass grown under SmF conditions showed no indication of containing pigments, data not shown).
Consequently, the production of pigments under SmFi was not only evaluated in terms of the absorbance increase in the fermentation medium but also in terms of the absorbance obtained after the ethanolic extraction of pigments from the biomass grown after 12 days of fermentation. Ethanol was utilized as an extraction solvent due to its generally recognized as safe (GRAS) status [
23] and also because it can be easily recovered and recycled using distillation or rotary evaporation.
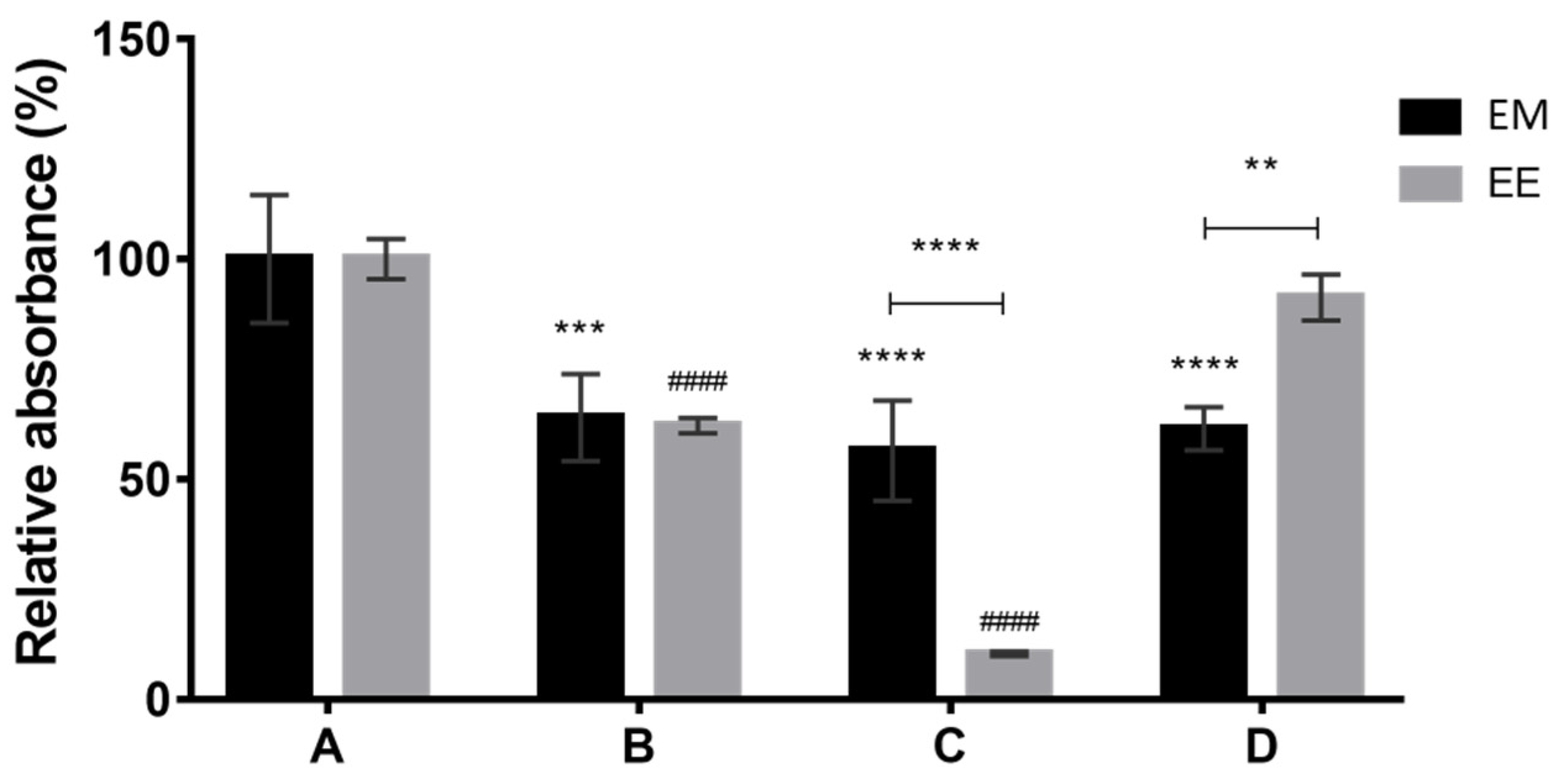
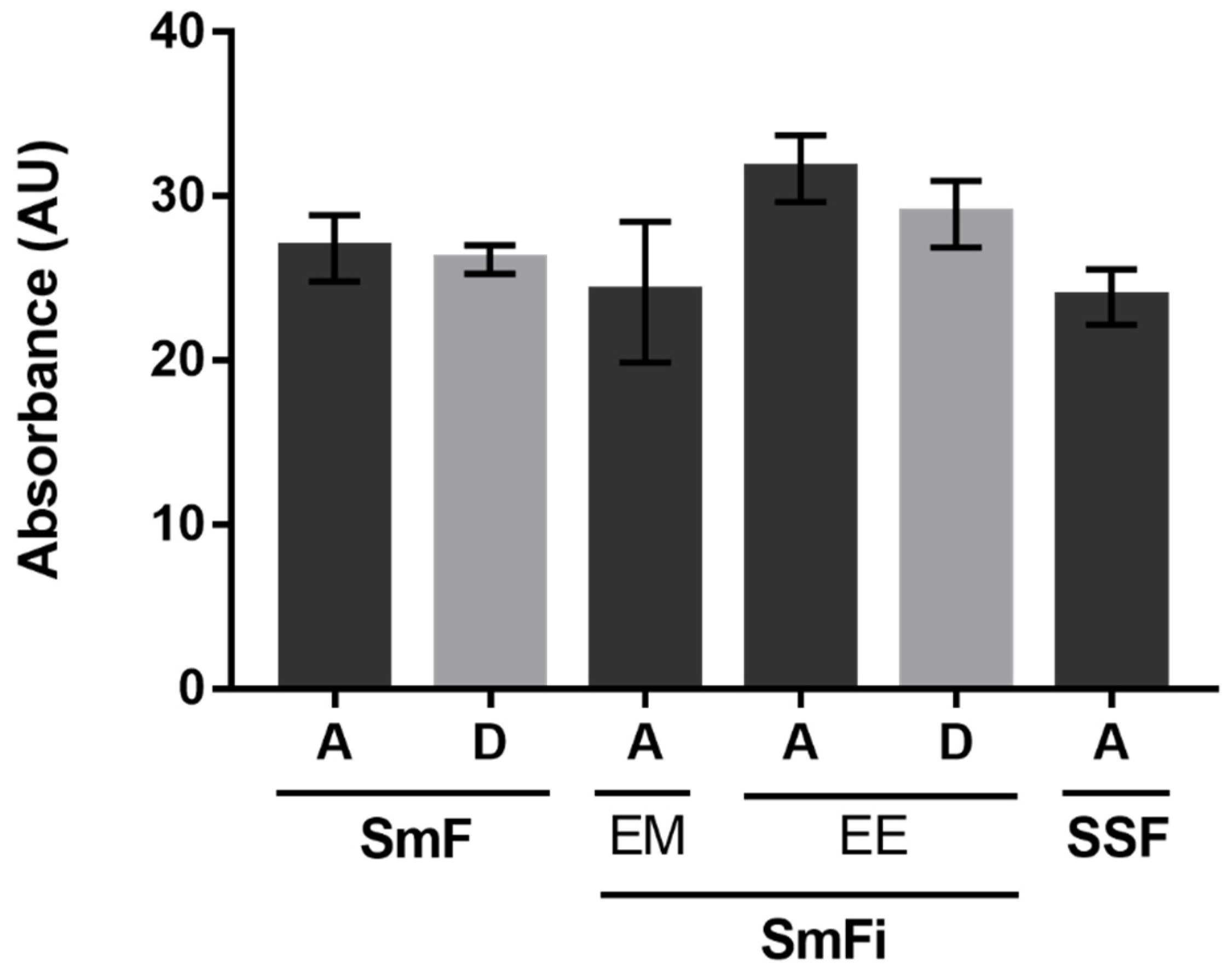
The results obtained under SmFi were expressed in relative absorbance (%) for the pigment mixtures from the extracellular medium (EM) and the ethanolic extraction (EE) of the biomass. All the alternative culture media were compared with the synthetic medium A, used as a reference, corresponding to 100% absorbance (
Figure 3).
Considering the absorbances of the EM, it is evident that the alternative culture media led to a poorer production of pigments compared to the reference medium A. Using the alternative culture media (B–D), the same production of pigments seems to be obtained in the EM. In this case, the medium composition seems to impact less the quantity of secreted pigments. On the other hand, for the EE, the production of pigments achieved with culture medium D is comparable to the one obtained with culture medium A, while the other alternative media presented a worse performance, especially medium C. For the medium composed only of CW (B), the same quantity of pigments was obtained from both the EM and the EE. For medium C, where only CSL was used, the quantity of pigments recovered from the biomass (EE) was extremely low. Taken together, these results suggest that, under SmFi conditions, the presence of the right carbon source is the primary factor affecting pigment synthesis, while the presence of a nitrogen source just helps to enhance it. Furthermore, comparing the obtained results using free (SmF) and immobilized mycelium (SmFi) for the same culture medium, it is suggested that pigment production (extra and intracellular) by P. brevicompactum is clearly affected by fungal immobilization.
The consumption of lactose over the fermentation time was also analyzed, and the results showed that culture medium A presented the lowest final lactose concentration (less than 5 g/L). For all the other media (B and D), more than 9 g/L of lactose was obtained after 12 days of fermentation. As the nylon sponge is a considerable porous material, the growth of biomass occurs both inside the support and on its surface, forming a dense coating. This fact may explain why lactose was not completely consumed. Probably, the biomass inside the support did not have the same access to the culture medium nutrients as the free biomass or the biomass on the support surface.
3.3. Pigments Production by SSF
The SmFi conditions showed that high amounts of pigments can be extracted from the immobilized biomass (
Figure 3), suggesting that SSF could be a promising fermentation process for pigment production by
P. brevicompactum. In fact, the use of SSF to produce pigments has been generally reported as the most suitable approach to obtain high yields with
Monascus strains. However, this type of fermentation was lesser explored for pigment production by
Penicillium strains, which mostly report SmF as an adequate condition to improve the yields [
38]. Nevertheless, SSF can present some advantages over SmF. In SSF, only the basic nutrients for fungal growth are provided, and the natural growth environment is mimicked, which can generally increase production yields with relatively low process costs [
39]. The moisture content is one of the key variables influencing the efficiency of the SSF. Using the most adequate moisture content for each support and microorganism, is crucial to promote favorable growth conditions and maximizingmetabolites production [
40,
41]. If the moisture content used is too high, gas diffusion will be restricted due to the occupancy of the cavities of the supports with water. On the other hand, if the moisture content is too low, fungal growth will be severely affected [
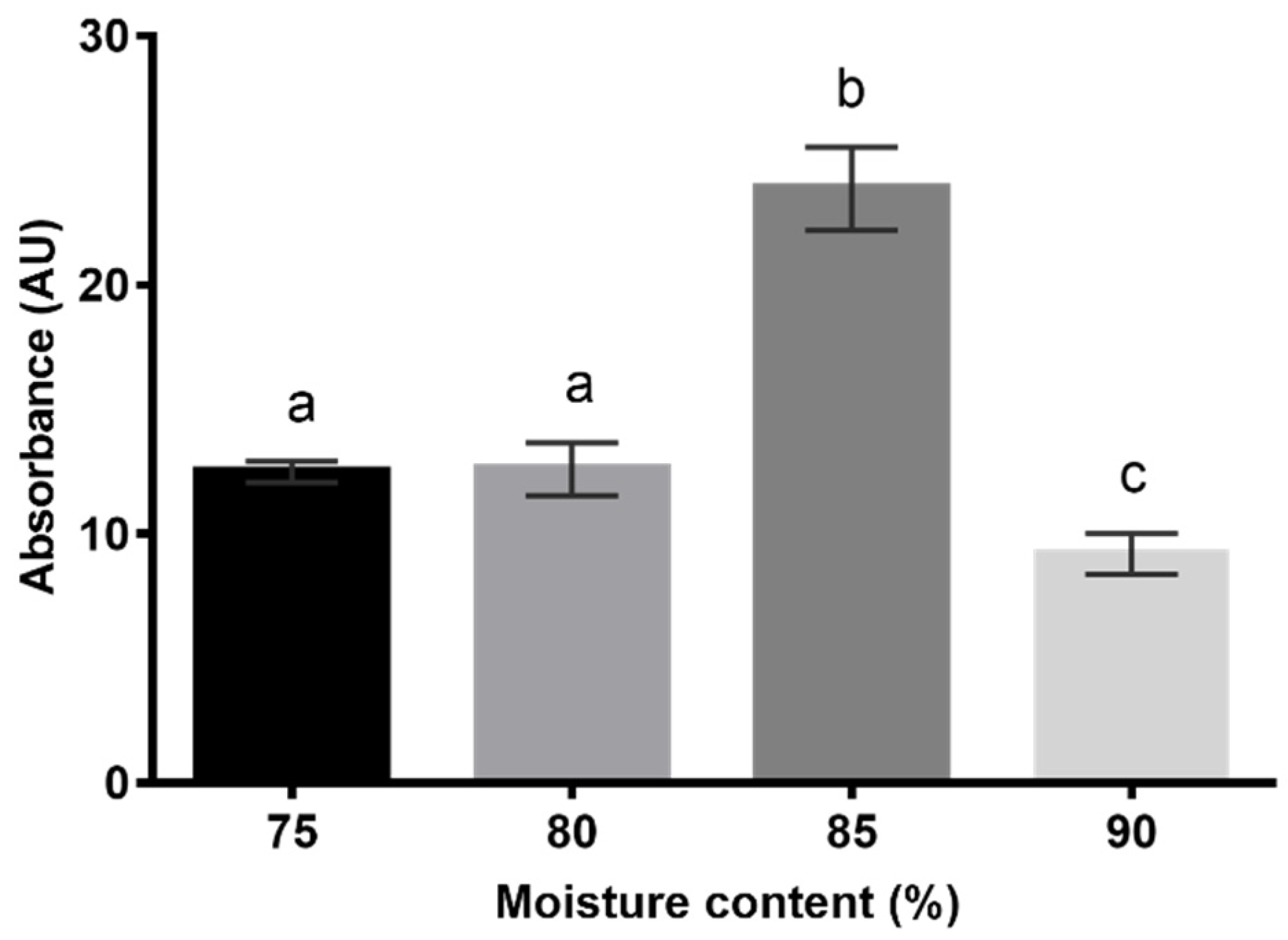
42]. Culture medium A was previously used to optimize the moisture content for pigment production under SSF. The amount of immobilization support (75 pieces corresponding to 3 g of nylon sponge) was established according to the volume occupied by it in the fermentation flask. The absorbances determined for the EE, obtained using the different moisture contents evaluated as a preliminary study, are represented in
Figure 4.
A moisture content of 85% (
w/w) resulted in 2 to 2.5 times more pigments than the other conditions, so it was selected for the studies of pigment production under SSF conditions.
Figure 5 summarizes the results obtained for the several culture media.
Despite the well-known SSF advantages, and the promising productivities previously obtained from SmFi, it was observed that none of the alternative culture media achieved pigment production similar to the one obtained with the synthetic medium A. Contrarily to the SmF conditions (free and immobilized), the supplementation of CW with CSL have no positive effect on pigments production under SSF conditions.
Nevertheless, our results illustrate the potential of SSF to be an economic strategy to produce pigments using lower volumes of culture medium. Furthermore, the three alternative culture media screened (B–D), composed of agro-industrial byproducts, were allowed to reach >50% of relative absorbance. It is important to highlight that, for comparison purposes, the concentration of the byproducts (carbon and nitrogen sources) was kept the same between experiments (SmF and SSF conditions). However, these values were previously optimized for the production of pigments under SmF conditions. Thus, our data suggest that under SSF conditions, perhaps higher pigment productivity might be achieved by optimizing the concentration of the byproducts (and consequently, the concentration of lactose and nitrogen) or by using different immobilization supports. The porosity of the nylon foam is suitable for fungal growth and nutrient access under SmF conditions. However, under SSF, this inert support showed some limitations, namely the poor capacity to retain the culture medium and preserve the moisture content in all the small cubes used. Based on these results and observations, it will be interesting to test other immobilization supports, including natural/organic supports composed of agro-industrial residues.
3.4. Direct Comparison of the Best Fermentation Conditions
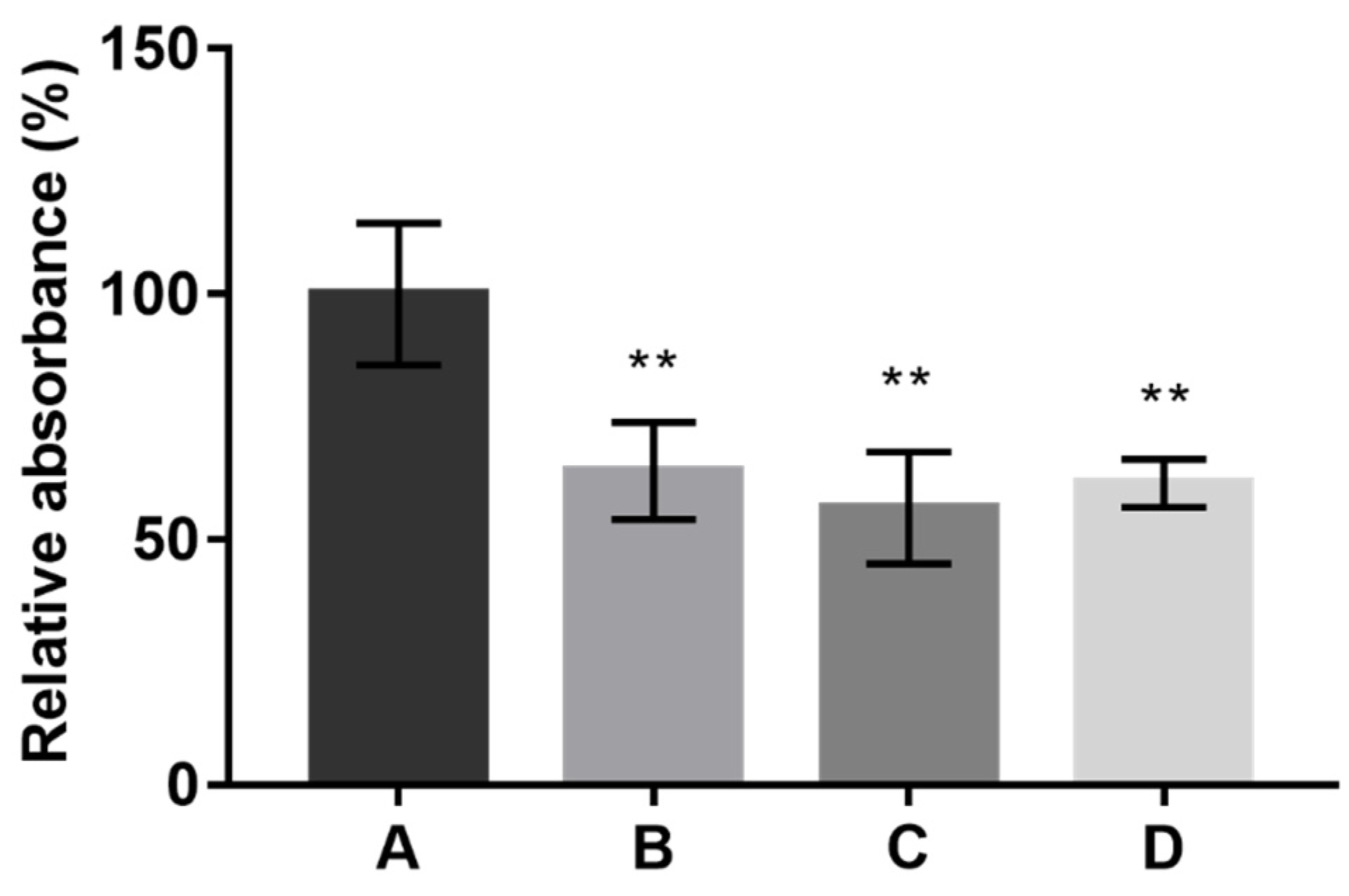
Attempting to identify which studied condition was the most suitable for pigment production, a direct comparison was established between the best results obtained in each type of fermentation. The best conditions selected are summarized in
Figure 6. For comparison purposes, the results are presented as absolute absorbance. The absolute absorbance corresponds to the sum of absorbances measured at 400, 470, and 500 nm for each specific solution (freeze-dried media or dried ethanolic extracts dissolved in 2.5 mL of distilled water) prepared with the mixture of pigments obtained from the different fermentation conditions.
As
Figure 6 illustrates, culture media A (synthetic, used as reference) and D (composed of CW and CSL) allowed us to obtain a higher production of pigments for all types of fermentation except for SSF. Interestingly, there are no statistical differences between the selected conditions, meaning that similar amounts of pigments were produced.
This analysis highlights the suitability of medium D for pigment production by
P. brevicompactum under SmF and SmFi (EE). Besides the comparable productivities with the synthetic medium A, its composition based only on agro-industrial byproducts also makes it more attractive from both the economic and environmental points of view. In fact, the use of byproducts can offer a significant economic advantage in the bioprocess since CSL represents one-fifth of the costs of yeast extract [
43], and CW is generally described as a byproduct of negligible cost [
44]. Using medium D, it is possible to reduce the accumulation of byproducts from the food industry and simultaneously generate new natural products with potential applications in several sectors.
Furthermore, in the case of SmFi, old sponge wastes or industrial leftovers can be reused as immobilization supports, thus contributing to lowering the costs of the entire process and still making it attractive from the circular economy perspective.
Finally, in order to determine if the same mixture of pigments was produced in all the selected conditions, the pigment profiles were qualitatively analyzed by TLC (
Figure 7). The mixtures of pigments were separated using water:ethanol (50:50) as eluent, which was shown in previous studies to be the best eluent for their separation [
45].
From the TLC analysis, it is evident that the medium composition affects the mixture of pigments being produced (e.g., media A and D under SmF, corresponding to 1 and 2, respectively, in
Figure 7). In addition, the fermentation type plays an important role too on pigment production (e.g., medium A under SmF and SmFi, corresponding to 1 and 3, respectively, in
Figure 7).
Overall, these results demonstrate that pigment production by
P. brevicompactum can be fine-tuned by adjusting the medium composition and choosing a specific fermentation type. In fact, the composition of the culture medium has always been suggested as a key factor for biomass growth and pigment metabolism [
46]. Moussa and Azeiz (2017) [
47] reported the important effect of the medium composition on the type of pigments synthesized by
M. ruber, leading to improve yield of specific desired pigments. Similarly, the effect of the fermentation type (SmF and SSF) on the produced pigments was also described for several fungi, including
Penicillium [
48]. Additionally, to understand the impact of the fermentation conditions on the type of synthesized pigments by
P. brevicompactum, a detailed study of separation and characterization of the several obtained mixtures should be performed in the future.
3.5. Biomass Reuse under Submerged Fermentation Conditions
In previous studies, it was determined that 12 days of submerged fermentation was a suitable time for pigment production using the culture medium A [
14]. In this work, the same fermentation time was used for the studies carried out with the alternative culture media, which also led to good performances. Nevertheless, aiming to have a process with the highest productivity possible, we investigated the production of pigments over 36 days of fermentation. In this study, we allowed the biomass to grow in 3 cycles of 12 days, and at the end of each cycle, the fermentation medium was collected and replaced by a fresh culture medium with the same initial composition. As a control, an extended batch fermentation (36 days) was performed. Medium D was selected for these experiments due to its positive results as an alternative to synthetic medium A, and it was used under SmF and SmFi conditions, since in such conditions, we have previously observed the highest production of pigments.
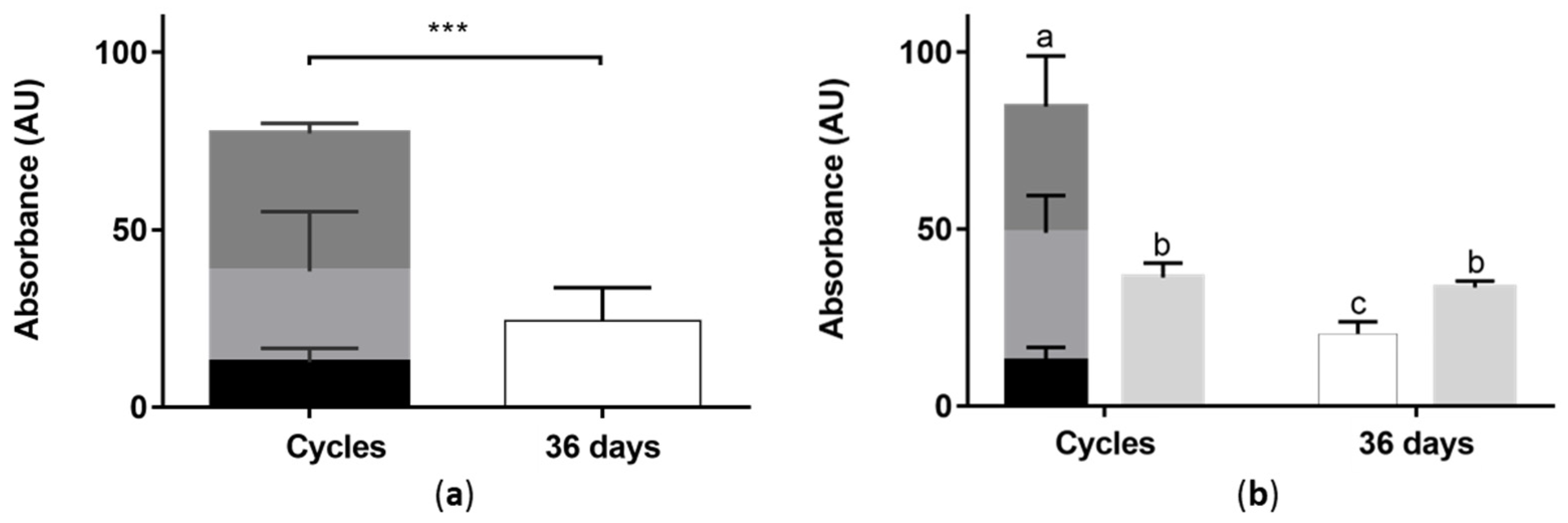
The sum of the absorbances determined for the different fermentation conditions (SmF, and SmFi with or without culture medium change) is presented in
Figure 8. As explained before, for SmFi, the extracellular medium (EM) and the ethanolic extract of the biomass (EE) were analyzed due to the presence of color on both.
Data show that for both SmF and SmFi, changing the culture medium every 12 days results in almost three times more pigments produced than in the control fermentations (36 days extended batch). Additionally, there are no statistical differences between the amounts of pigments produced in each cycle. These results demonstrate that a strategy based on the sequential removal of fermented broth and its replacement by fresh culture medium can indeed be an interesting strategy to promote higher productivity in pigment production.
Secondary metabolites are known to be produced under stress conditions [
49,
50]. However, some authors have stated that secondary metabolite production in fungi is more dependent on specific factors, including the type, concentration, source of nutrients, and adequate ratio in the culture medium [
51,
52]. Our data corroborate that fact since adequate carbon and nitrogen sources in the culture medium resulted in higher pigment production than when they are missing, or replaced by unsuitable ones. In the cycle operational mode, biomass always has higher amounts of available nutrients, producing pigments whenever the fermentation medium is renewed. In contrast, in the extended batch (36 days), the biomass probably consumes all the nutrients in the first 12 days, having no supplies to help or induce pigment synthesis later on. These results support the assumption that the concentration of carbon and nitrogen sources in the culture medium is essential to stimulate and activate the metabolic pathways involved in pigments production.
Interestingly, the absorbances of the ethanolic extracts from SmFi were similar for both extended batch and medium change conditions. In the extended batch, they were also higher than the absorbances of the extracellular medium, as seen before during the SmFi screening experiments. Contrarily, in the cycle operation mode, the absorbance values were higher in the extracellular medium, suggesting that more pigments were released from the biomass grown for 36 days with a fresh culture medium.
Overall, higher production of pigments was observed under SmFi conditions and cycle operation mode, considering both the pigments obtained from the extracellular medium and the ethanolic extraction of the biomass. Moreover, this type of fermentation with mycelium immobilization proved to be simpler and more suitable for the successive cycles of culture medium exchange. As far as we know, this strategy of biomass immobilization and reuse was never studied for pigment production. It represents an interesting approach for the development of low-cost and sustainable bioprocesses operating at an industrial scale and can be optimized in future studies of pigment production using agro-industrial byproducts/wastes.




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}







