
Production of Pigments by Filamentous Fungi Cultured on Agro-Industrial by-Products Using Submerged and Solid-State Fermentation Methods

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
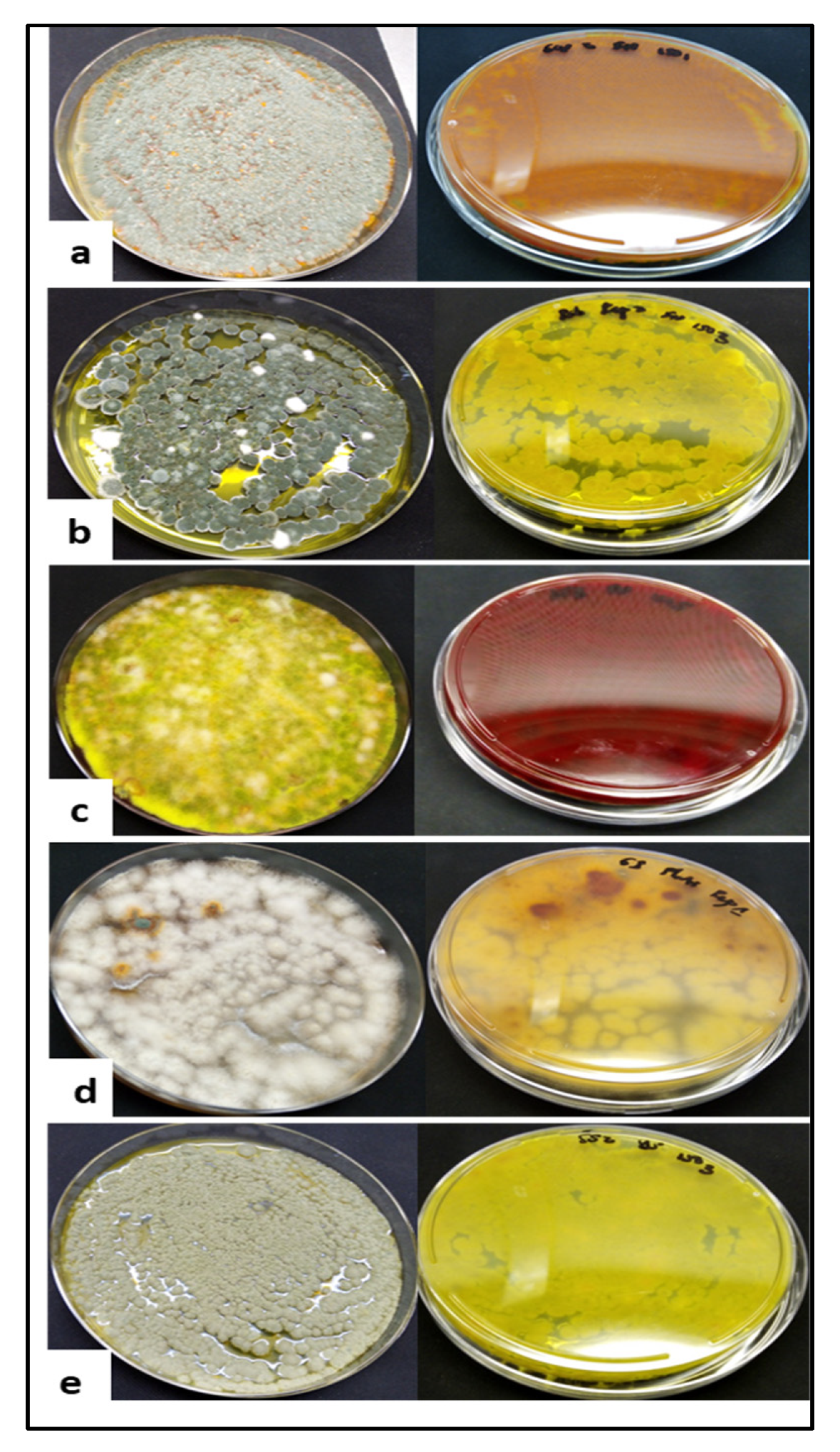
2.1. Fungal Isolates
2.1.1. Culture and Purification of the Fungi from Additional Soil Samples
2.1.2. Fungal DNA Extraction, Amplification and Sequencing
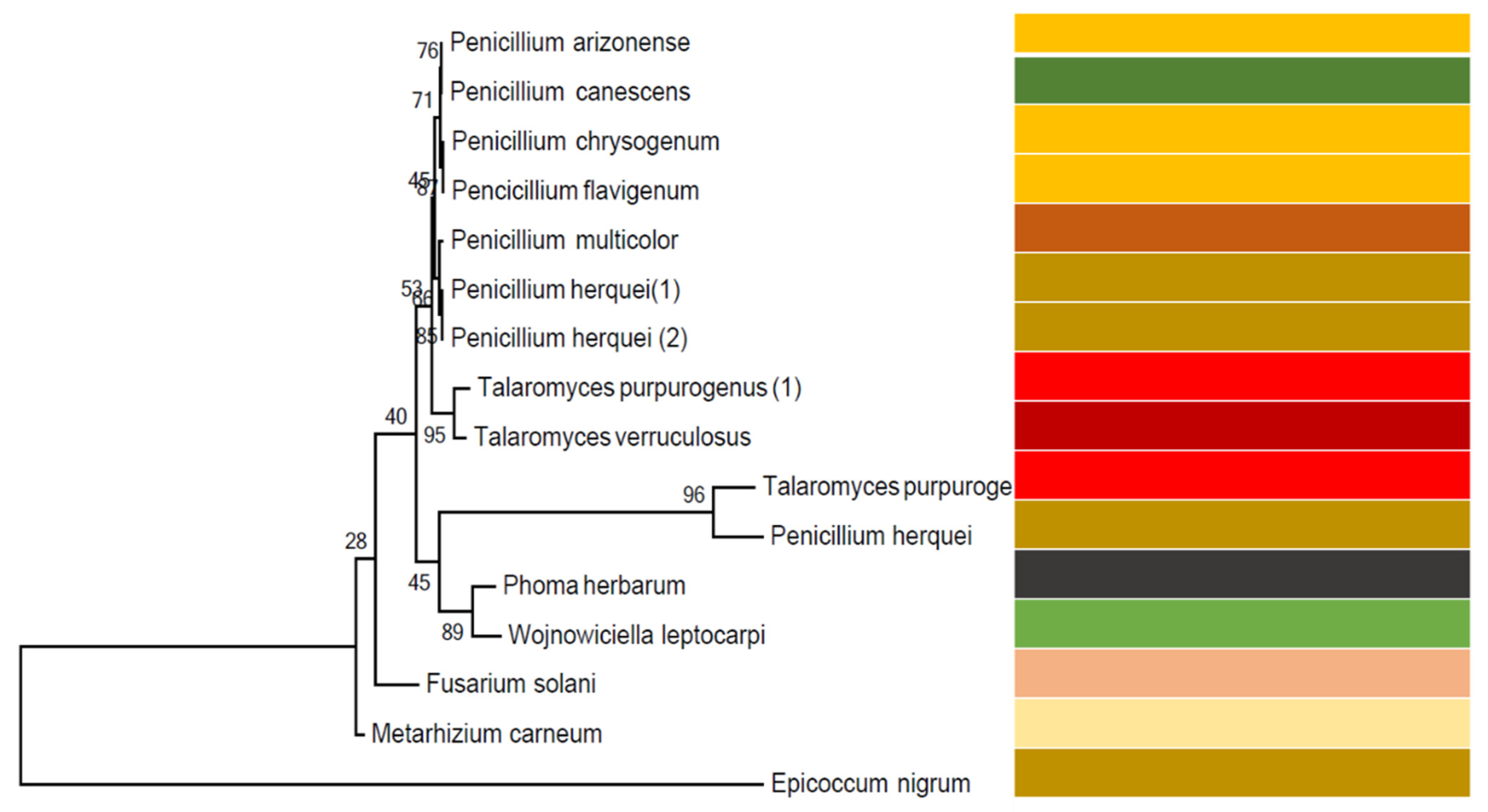
2.1.3. Fungal Identification and Construction of Phylogenetic Tree
2.2. Characterisation of Green Waste
2.3. Submerged Fermentation
2.4. Solid State Fermentation
2.5. Enzyme Activity
2.5.1. Media Composition for α-Amylase Activity
2.5.2. Media Composition for Pectinase Activity
2.5.3. Media Composition for Cellulase Activity
2.6. Colour Measurements
2.7. Statistical Analysis
3. Results and Discussion
3.1. Fungal Identities and Phylogenetic Tree
3.2. Green Waste Characterisation
3.3. Enzyme Activity
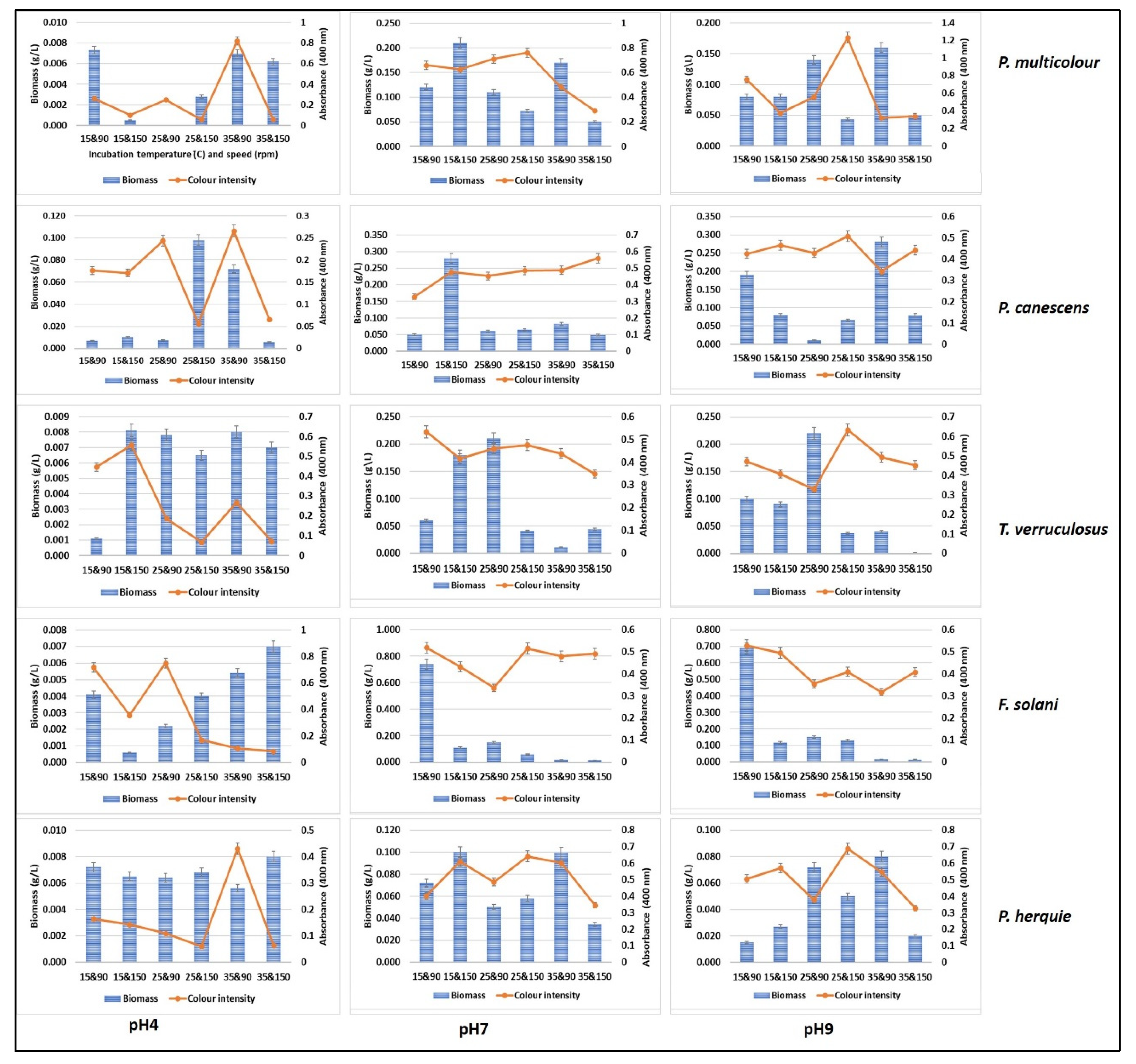
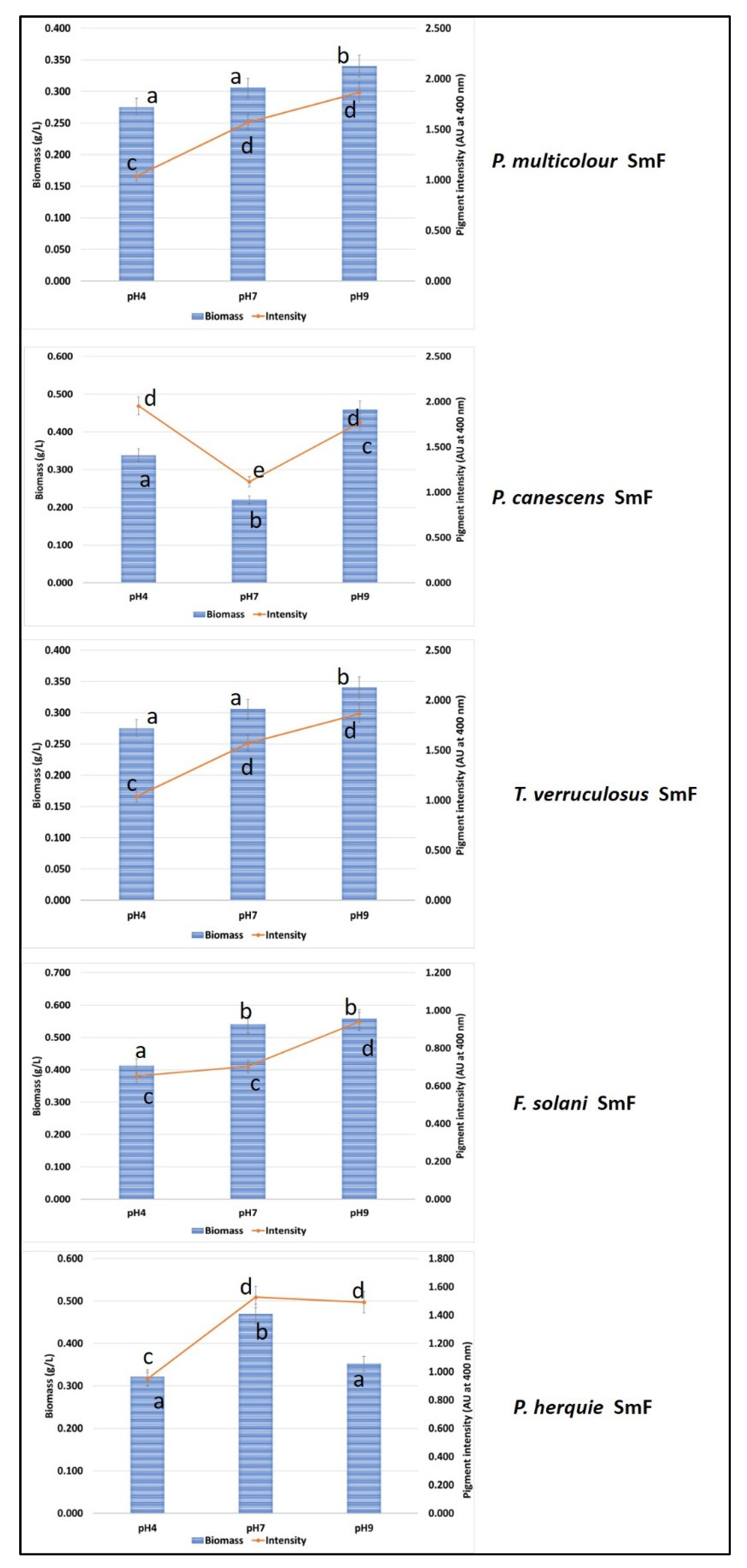
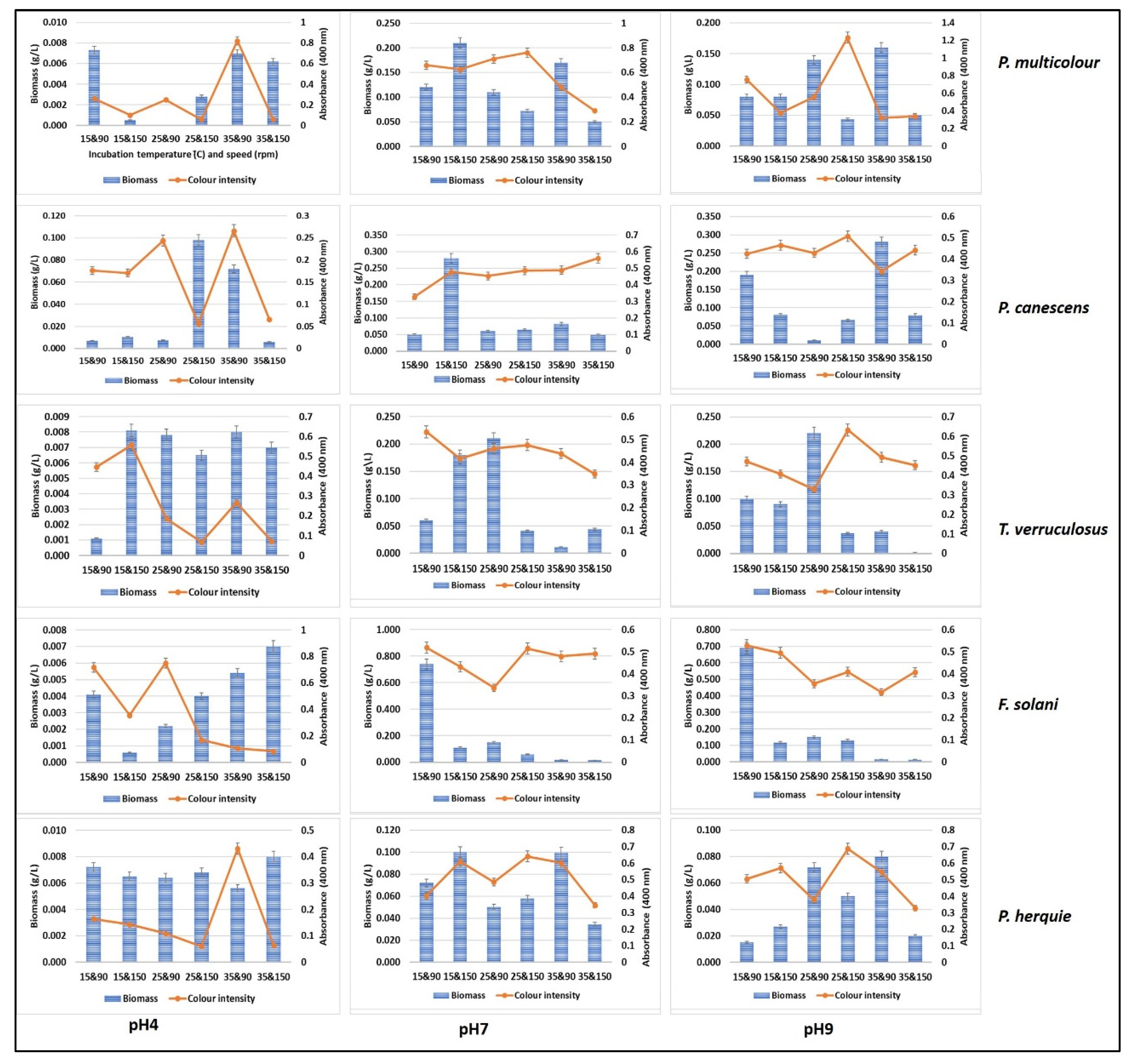
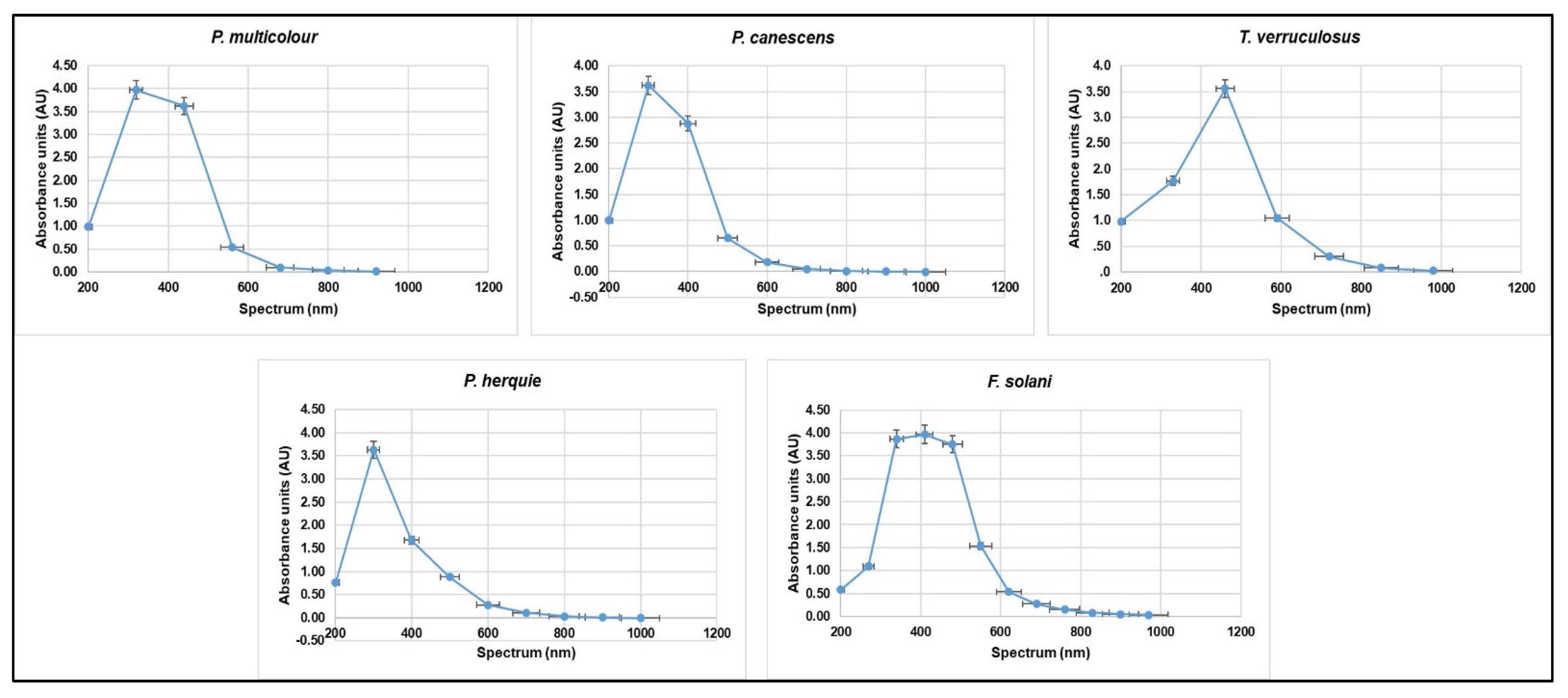
3.4. Submerged Fermentation for Pigment Production Using Green Waste and Whey Cocktail
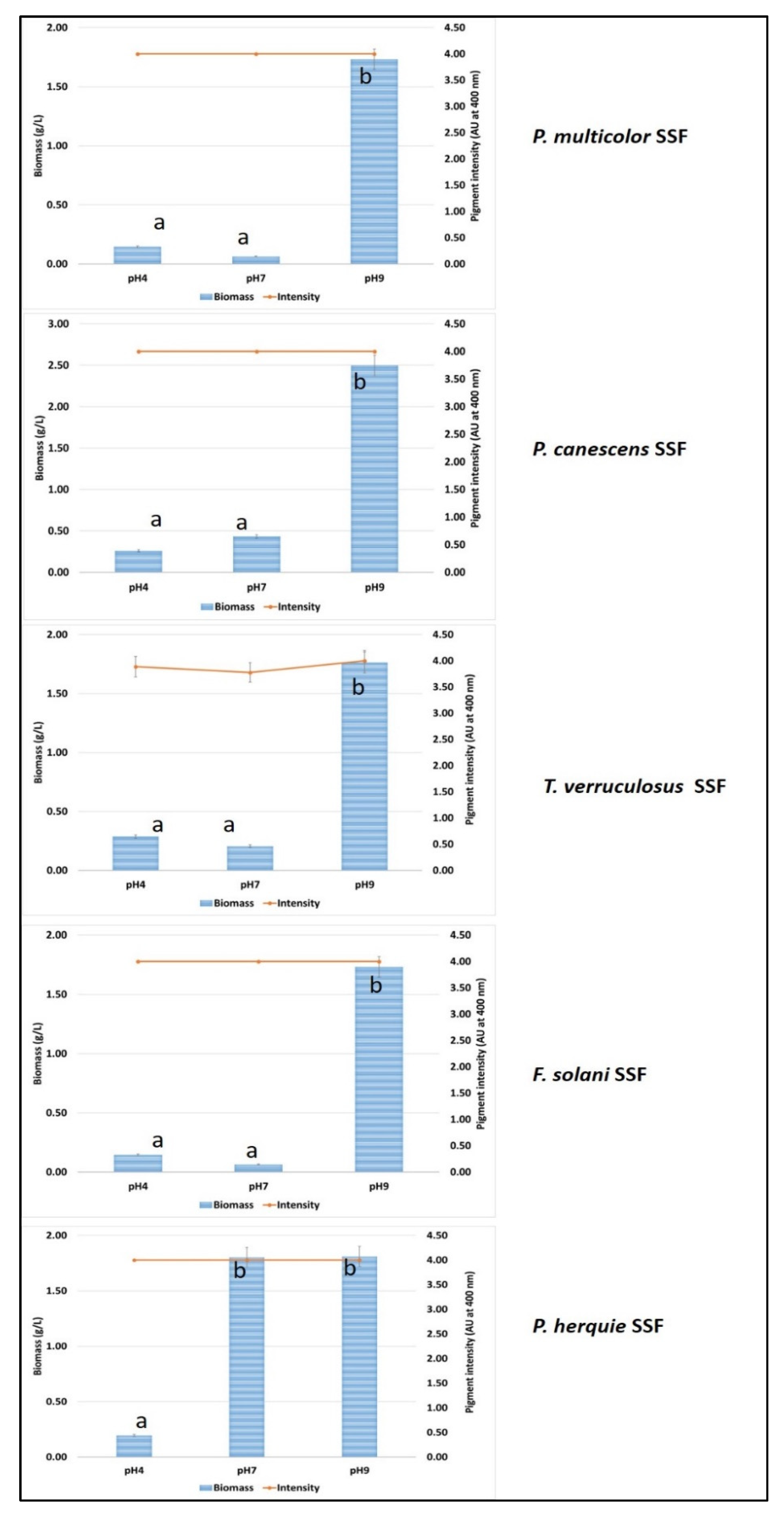
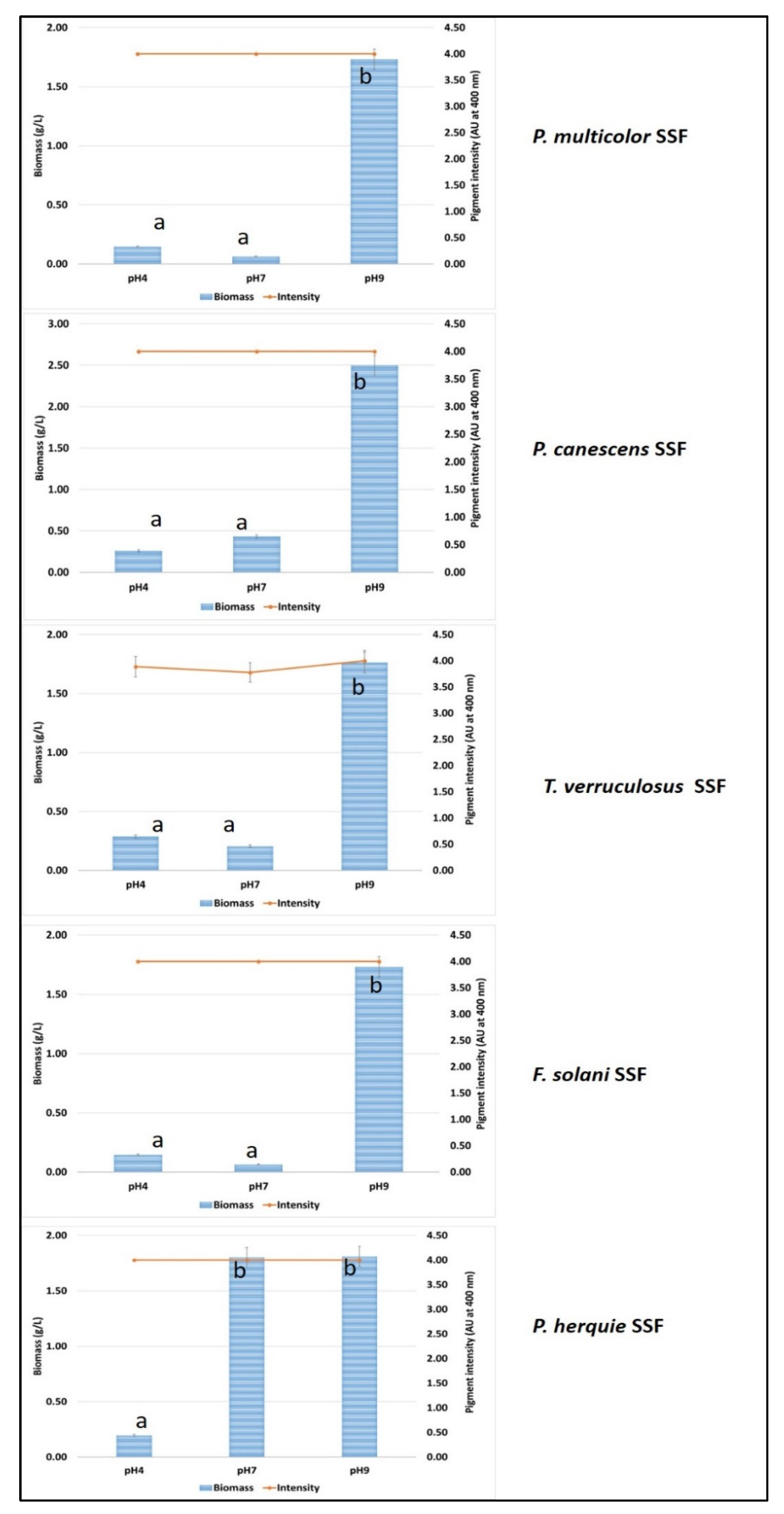
3.5. Solid State Fermentation Using Green Waste and Whey Cocktail
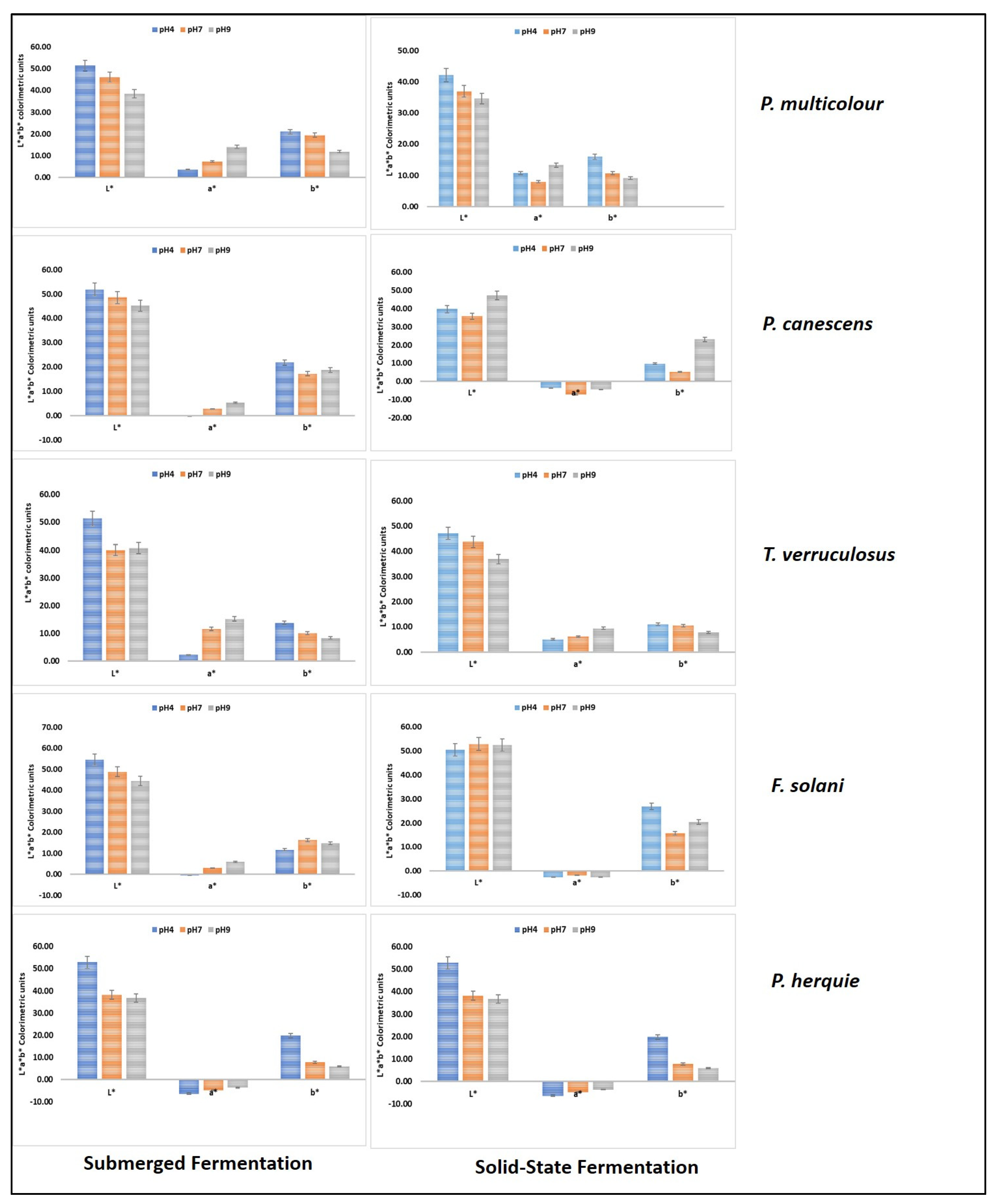
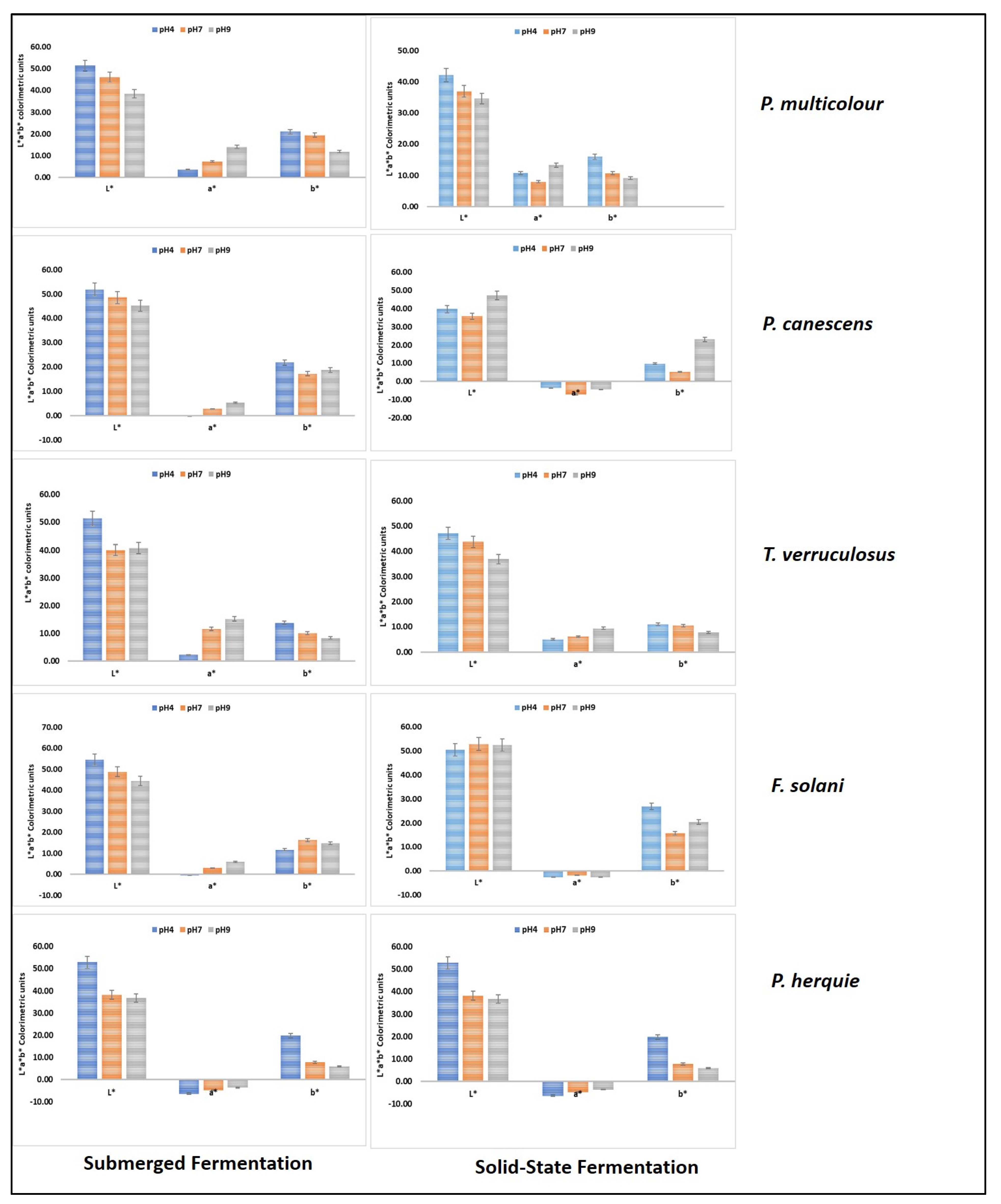
3.6. Colour Measurements
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bielčik, M.; Aguilar-Trigueros, C.A.; Lakovic, M.; Jeltsch, F.; Rillig, M.C. The role of active movement in fungal ecology and community assembly. Mov. Ecol. 2019, 7, 36. [Google Scholar] [CrossRef]
- Olicón-Hernández, D.R.; González-López, J.; Aranda, E. Overview on the Biochemical Potential of Filamentous Fungi to Degrade Pharmaceutical Compounds. Front. Microbiol. 2017, 8, 1792. [Google Scholar] [CrossRef] [PubMed]
- Perlin, M.H.; Andrews, J.; Su San, T. Chapter Four: ABC and MFS Transporters and Their Roles in Survival and Pathogenicity. In Essential Letters in the Fungal Alphabet; Friedmann, T., Dunlap, J.C., Goodwin, S.F., Eds.; Academic Press: Cambridge, MA, USA, 2014; Volume 85, pp. 201–253. [Google Scholar]
- Werner, G.D.A.; Kiers, T. Friends in Fungi. 2012. Available online: https://science.sciencemag.org/content/sci/337/6101/1452.1.full.pdf?casa_token=GlztZ81gCGMAAAAA:o-Q3Wiz-WOXMJmiQtriQ9fcwMKH6WNJCImel6HixqdP67QsRru0O9WK2oEUKvWT_QWoasw2X496G (accessed on 25 August 2021).
- Venkatachalam, M.; Zelena, M.; Cacciola, F.; Ceslova, L.; Girard-Valenciennes, E.; Clerc, P.; Dugo, P.; Mondello, L.; Fouillaud, M.; Rotondo, A.; et al. Partial characterization of the pigments produced by the marine-derived fungus Talaromyces albobiverticillius 30548. Towards a new fungal red colorant for the food industry. J. Food Compos. Anal. 2018, 67, 38–47. [Google Scholar] [CrossRef]
- Lagashetti, A.C.; Dufossé, L.; Singh, S.K.; Singh, P.N. Fungal Pigments and Their Prospects in Different Industries. Microorganisms 2019, 7, 604. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spiteller, P. Chemical ecology of fungi. Nat. Prod. Rep. 2015, 32, 971–993. [Google Scholar] [CrossRef]
- Sezgin, A.; Sibel, A. Food Additives: Colorants. 2017. Available online: https://www.researchgate.net/publication/322404775_Food_Additives_Colorants (accessed on 29 July 2021).
- Mansour, R.; Yusuf, M. Natural Dyes and Pigments: Extraction and Applications. In Handbook of Renewable Materials for Coloration and Finishing; Scrivener Publishing LLC: Beverly, MA, USA, 2018; pp. 75–102. [Google Scholar]
- Hoeksma, J.; Misset, T.; Wever, C.; Kemmink, J.; Kruijtzer, J.; Versluis, K.; Liskamp, R.M.J.; Boons, G.J.; Heck, A.; Boekhout, T.; et al. A new perspective on fungal metabolites: Identification of bioactive compounds from fungi using zebrafish embryogenesis as read-out. Sci. Rep. 2019, 9, 17546. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Panesar, R. Bio-utilization of Kinnow waste for the production of biopigments using submerged fermentation. Indian J. Nat. Prod. Resour. 2014, 3, 6. [Google Scholar]
- Finkelstein, M.; Huang, C.-C.; Byng, G.S.; Tsau, B.-R.; Leach, J. Blakeslea Trispora Mated Culture Capable of Increased be-ta-carotene Production. U.S. Patent 5,422,247, 6 June 1995. [Google Scholar]
- Xu, F.; Yuan, Q.-P.; Zhu, Y. Improved production of lycopene and β-carotene by Blakeslea trispora with oxygen-vectors. Process. Biochem. 2007, 42, 289–293. [Google Scholar] [CrossRef]
- Kurcz, A.; Błażejak, S.; Kot, A.M.; Bzducha-Wróbel, A.; Kieliszek, M. Application of Industrial Wastes for the Production of Microbial Single-Cell Protein by Fodder Yeast Candida utilis. Waste Biomass-Valorization 2018, 9, 57–64. [Google Scholar] [CrossRef] [Green Version]
- Kieliszek, M.; Piwowarek, K.; Kot, A.M.; Pobiega, K. The aspects of microbial biomass use in the utilization of selected waste from the agro-food industry. Open Life Sci. 2020, 15, 787–796. [Google Scholar] [CrossRef]
- O’Shea, N.; Ktenioudaki, A.; Smyth, T.; Mcloughlin, P.; Doran, L.; Auty, M.; Arendt, E.; Gallagher, E. Physicochemical assessment of two fruit by-products as functional ingredients: Apple and orange pomace. J. Food Eng. 2015, 153, 89–95. [Google Scholar] [CrossRef]
- Dufossé, L. Microbial Production of Food Grade Pigments. Food Technol Biotech. 2006, 44, 313–321. [Google Scholar]
- Benavente-Valdésa, J.R.; Aguilara, C.; Contreras-Esquivela, J.C.; Méndez-Zavalab, A.; Montañez, J. Strategies to enhance the production of photosynthetic pigments and lipids in Chlorophycae species. Biotechnol. Rep. 2016, 10, 117–125. [Google Scholar] [CrossRef] [Green Version]
- Department of Environmental Affairs. South Africa State of Waste. A Report on the State of the Environment. Final Draft Report; Department of Environmental Affairs: Pretoria, South Africa, 2018; 112p. [Google Scholar]
- Arikan, E.B.; Canli, O.; Caro, Y.; Dufossé, L.; Dizge, N. Production of Bio-Based Pigments from Food Processing Industry By-Products (Apple, Pomegranate, Black Carrot, Red Beet Pulps) Using Aspergillus carbonarius. J. Fungi 2020, 6, 240. [Google Scholar] [CrossRef]
- Babitha, S.; Soccol, C.R.; Pandey, A. Effect of stress on growth, pigment production and morphology of Monascus sp. in solid cultures. J. Basic Microbiol. 2007, 47, 118–126. [Google Scholar] [CrossRef] [PubMed]
- Hailu, S.L.; McCrindle, R.I.; Seopela, M.P.; Combrinck, S. Speciation of major and trace elements leached from coal fly ash and the kinetics involved. J. Environ. Sci. Heal. Part A 2019, 54, 1186–1196. [Google Scholar] [CrossRef] [PubMed]
- Rovera, M.R.; Johnston, P.A.; Lamsal, B.P.; Brown, R.C. Total water-soluble sugars quantification in bio-oil using the phenol–sulfuric acid assay. J. Anal. Appl. Pyrol. 2013, 104, 194–201. [Google Scholar] [CrossRef]
- Pratap, A.; Kumar, M.; Sibi, G. Fruit and Vegetable Waste Hydrolysates as Growth Medium for Higher Biomass and Lipid Production in Chlorella vulgaris. J. Environ. Manage. 2017, 4, 204–210. [Google Scholar]
- Gmoser, R.; Sintca, C.; Taherzadeh, M.; Lennartsson, P.R. Combining submerged and solid state fermentation to convert waste bread into protein and pigment using the edible filamentous fungus N. intermedia. Waste Manag. 2019, 97, 63–70. [Google Scholar] [CrossRef]
- Wang, W.; Li, Y.; Wang, H.; Zu, Y. Differences in the Activities of Eight Enzymes from Ten Soil Fungi and Their Possible In-fluences on the Surface Structure, Functional Groups, and Element Composition of Soil Colloids. PLoS ONE 2014, 9, e111740. [Google Scholar]
- Yakop, F.; Taha, H.; Shivanand, P. Isolation of Fungi from Various Habitats and their Possible Bioremediation. Curr. Sci. 2019, 116, 733. [Google Scholar] [CrossRef]
- Fouillaud, M.; Venkatachalam, M.; Llorente, M.; Magalon, H.; Cuet, P.; Dufossé, L. Biodiversity of Pigmented Fungi Isolated from Marine Environment in La Réunion Island, Indian Ocean: New Resources for Colored Metabolites. J. Fungi 2017, 3, 36. [Google Scholar] [CrossRef] [PubMed]
- FoodDive: Why Are Natural Colours Poised to Pop in 2021? Available online: http://www.fooddive.com/news/why-natural-colours-are-poised-to-pop-in-2021/591608 (accessed on 29 July 2021).
- Yanis, C.; Fouillaud, M.; Philippe, L.; Dufossé, L. Natural Hydroxyanthraquinoid Pigments: Current Situation and Future Op-Portunities in Food. Pigments in Food VII. 2013. Available online: Re-searchgate.net/publication/258553667_Natural_hydroxyanthraquinoid_pigments_current_situation_and_future_opportunities_in_food (accessed on 16 November 2021).
- Künzler, M. How fungi defend themselves against microbial competitors and animal predators. PLOS Pathog. 2018, 14, e1007184. [Google Scholar] [CrossRef] [PubMed]
- Gmoser, R.; Ferreira, J.A.; Lennartsson, P.R.; Taherzadeh, M.J. Filamentous ascomycetes fungi as a source of natural pigments. Fungal Biol. Biotechnol. 2017, 4, 1–25. [Google Scholar] [CrossRef] [Green Version]
- Lebeau, J.; Petit, T.; Clerc, P.; Dufossé, L.; Caro, Y. Isolation of two novel purple naphthoquinone pigments concomitant with the bioactive red bikaverin and derivates thereof produced by Fusarium oxysporum. Biotechnol. Prog. 2019, 35, e2738. [Google Scholar] [CrossRef] [Green Version]
- Joshi, V.K.; Rana, N.S. Optimization of apple pomace based medium and fermentation conditions for pigment production by Sarcina sp. Indian J. Nat. Prod. Resour. 2011, 2, 421–427. [Google Scholar]
- Tarangini, K.; Mishra, S. Production of melanin by soil microbial isolate on fruit waste extract: Two step optimization of key parameters. Biotechnol. Rep. 2014, 4, 139–146. [Google Scholar] [CrossRef] [Green Version]
- Usman, H.M.; Farouq, A.; Baki, A.; Abdulkadir, N.; Mustapha, G. Production and characterization of orange pigment produced by Halophilic bacterium Salinococcus roseus isolated from Abattoir soil. J. Microbiol. Exp. 2018, 6, 238–243. [Google Scholar] [CrossRef] [Green Version]
- Coultate, T.P. Food: The Chemistry of Its Components, 6th ed.; The Royal Society of Chemistry: Cambridge, UK, 2016. [Google Scholar]
- Nielsen, J.C. Development of fungal cell factories for the production of secondary metabolites: Linking genomics and metabolism. Synth. Syst. Biotechnol. 2017, 2, 5–12. [Google Scholar] [CrossRef]
- Wheltze, M. Measuring Color Using Hunter L, a, b versus CIE 1976 L*a*b*. 2016. Available online: https://www.hunterlab.com/media/documents/duplicate-of-an-1005-hunterlab-vs-cie-lab.pdf (accessed on 25 April 2020).
- Pombeiro-Sponchiado, S.R.; Sousa, G.S.; Andrade, J.C.R.; Lisboa, H.F.; Gonçalves, R.C.R. Production of Melanin Pigment by Fungi and Its Biotechnological Applications. In Melanin; Blumenber, M., Ed.; IntechOpen: London, UK, 2017; p. 3760. [Google Scholar]
- Parmar, R.S.; Singh, C. A comprehensive study of eco-friendly natural pigment and its applications. Biochem. Biophys. Rep. 2018, 13, 22–26. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Soil Sample | ITS Fugal Identity | Colour |
|---|---|---|
| Bio and Food Tech | Wojnowiciella leptocarpi | green |
| Bio and Food Tech | Epicoccum nigrum | Olive |
| Somerset West | Penicillium herquei | Olive |
| Somerset West | Fusarium solani | Olive |
| Somerset West | P. herquei | Olive-Yellow |
| Somerset West | P. herquei | Olive-Yellow |
| Somerset West | Tolaromyces verruculosus | Red |
| Guano Islands | P. flavigenum | Yellow |
| Guano Islands | Fusarium solani | Orange |
| Guano Islands | P. chrysogenum | Yellow |
| Pilanesberg | P. arizonense | Yellow |
| Moreleta Park nature reserve | P. mollochii | Dark red |
| Moreleta Park nature reserve | Phoma herbarum | Black |
| Moreleta Park nature reserve | P. multicolor | Dark Orange |
| Somerset West | Tolaromyces purpurogenus | Red |
| Pilanesberg | P. canescens | Green |
| Pilanesberg | Metarhizium carneum | Yellow |
| Fungal Isolate | α-Amylase | Cellulase | Pectinase |
|---|---|---|---|
| P. multicolour | ++ | ++ | - |
| P. canescens | + | ++ | +++ |
| T. verruculosus | + | +++ | - |
| F. solani | +++ | +++ | + |
| P. herquie | +++ | ++ | - |
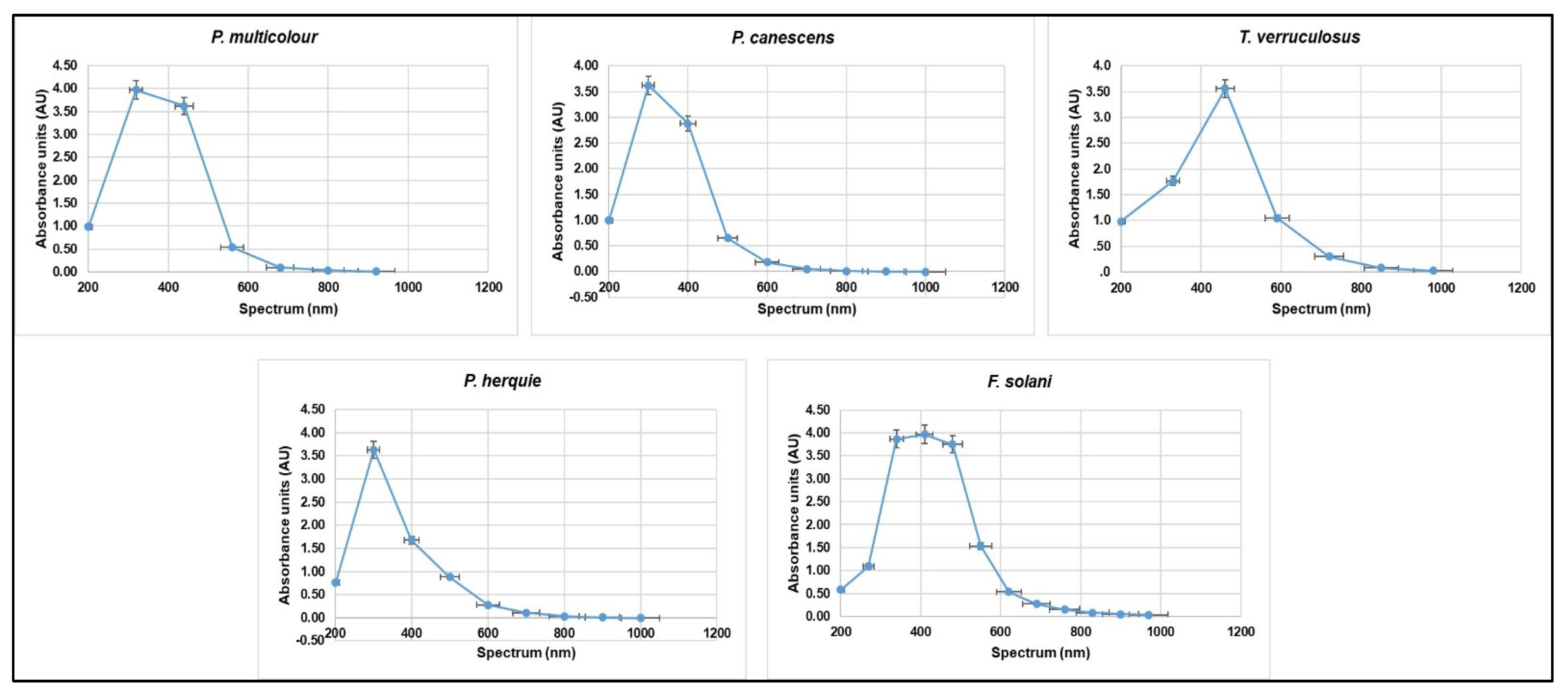
| Crude Pigment Sample | Spectrum (nm) | |
|---|---|---|
| λmax 1 | λmax 2 | |
| Peaks in UV Region 200–400 nm | Peaks Outside the UV Region >400 nm | |
| P. multicolor | 320 | 440–560 |
| P. canescens | 300–400 | 500 |
| T. verruculosus | 300 | 460–590 |
| F. solani | 270–340 | 410–550 |
| P. herquie | 330–400 | 500 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Molelekoa, T.B.J.; Regnier, T.; da Silva, L.S.; Augustyn, W. Production of Pigments by Filamentous Fungi Cultured on Agro-Industrial by-Products Using Submerged and Solid-State Fermentation Methods. Fermentation 2021, 7, 295. https://doi.org/10.3390/fermentation7040295
Molelekoa TBJ, Regnier T, da Silva LS, Augustyn W. Production of Pigments by Filamentous Fungi Cultured on Agro-Industrial by-Products Using Submerged and Solid-State Fermentation Methods. Fermentation. 2021; 7(4):295. https://doi.org/10.3390/fermentation7040295
Chicago/Turabian StyleMolelekoa, Tumisi Beiri Jeremiah, Thierry Regnier, Laura Suzanne da Silva, and Wilma Augustyn. 2021. "Production of Pigments by Filamentous Fungi Cultured on Agro-Industrial by-Products Using Submerged and Solid-State Fermentation Methods" Fermentation 7, no. 4: 295. https://doi.org/10.3390/fermentation7040295
APA StyleMolelekoa, T. B. J., Regnier, T., da Silva, L. S., & Augustyn, W. (2021). Production of Pigments by Filamentous Fungi Cultured on Agro-Industrial by-Products Using Submerged and Solid-State Fermentation Methods. Fermentation, 7(4), 295. https://doi.org/10.3390/fermentation7040295