
Submerged Fermentation of Animal Fat By-Products by Oleaginous Filamentous Fungi for the Production of Unsaturated Single Cell Oil

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Microorganisms
2.2. Media Composition and Preparation of Inoculums
2.3. Shake Flask (SF) Cultivations
2.4. Bioreactor Cultivations
2.5. Sampling and Analytical Procedures
2.6. Determination of Lipid Content and Fatty Acid (FA) Composition of Biomass
2.7. Determination of Lipid Content in Emulsion
3. Results
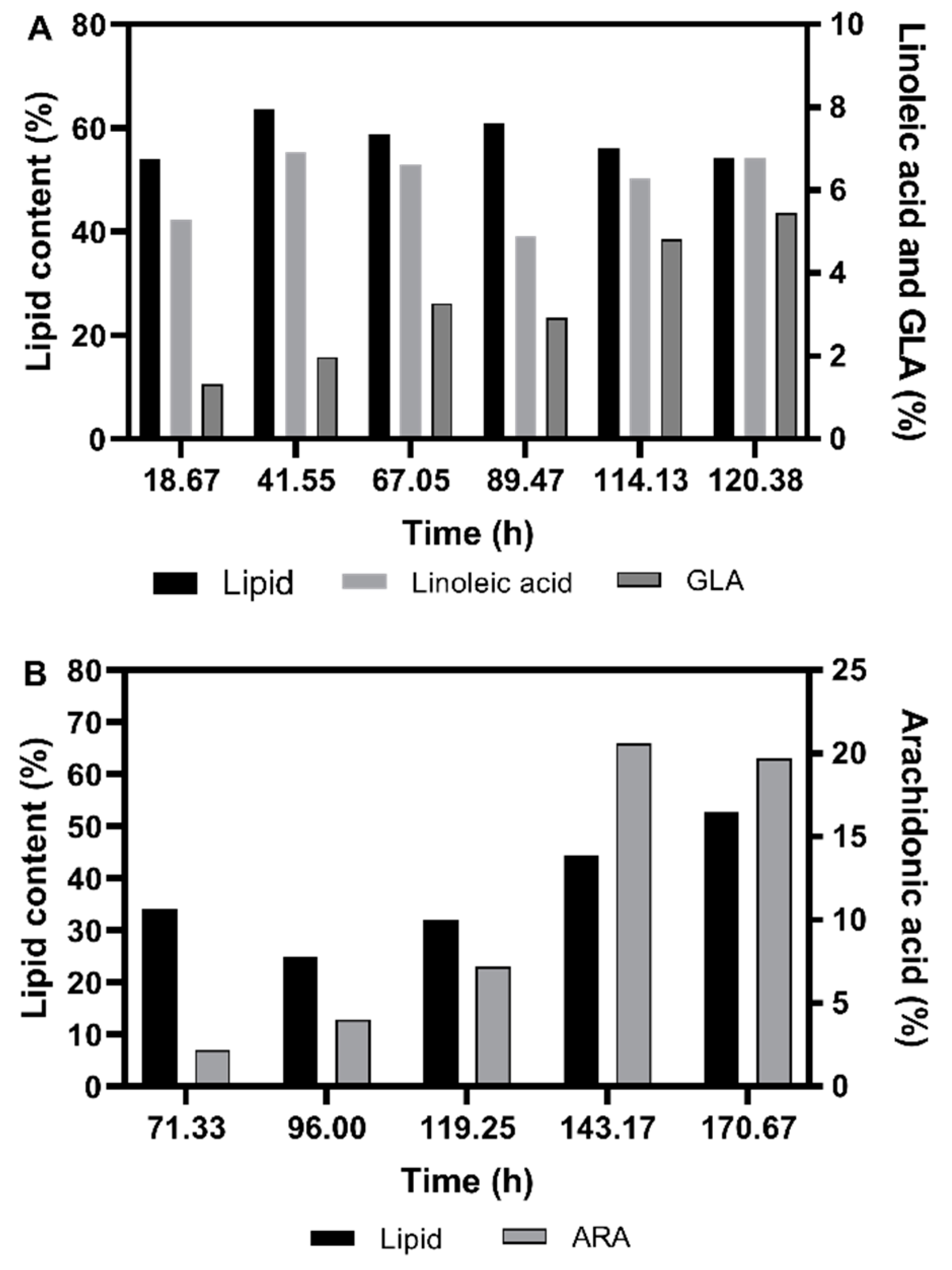
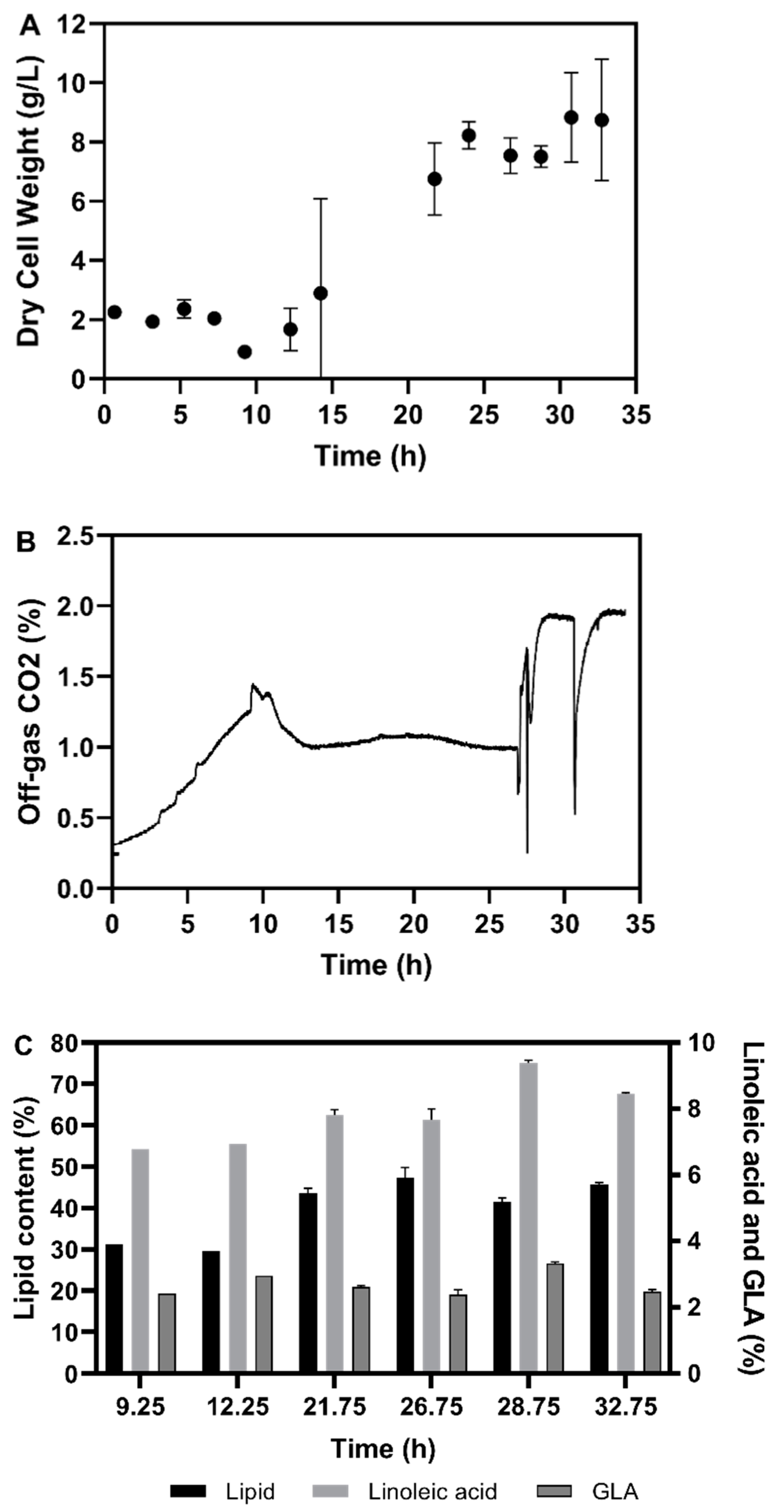
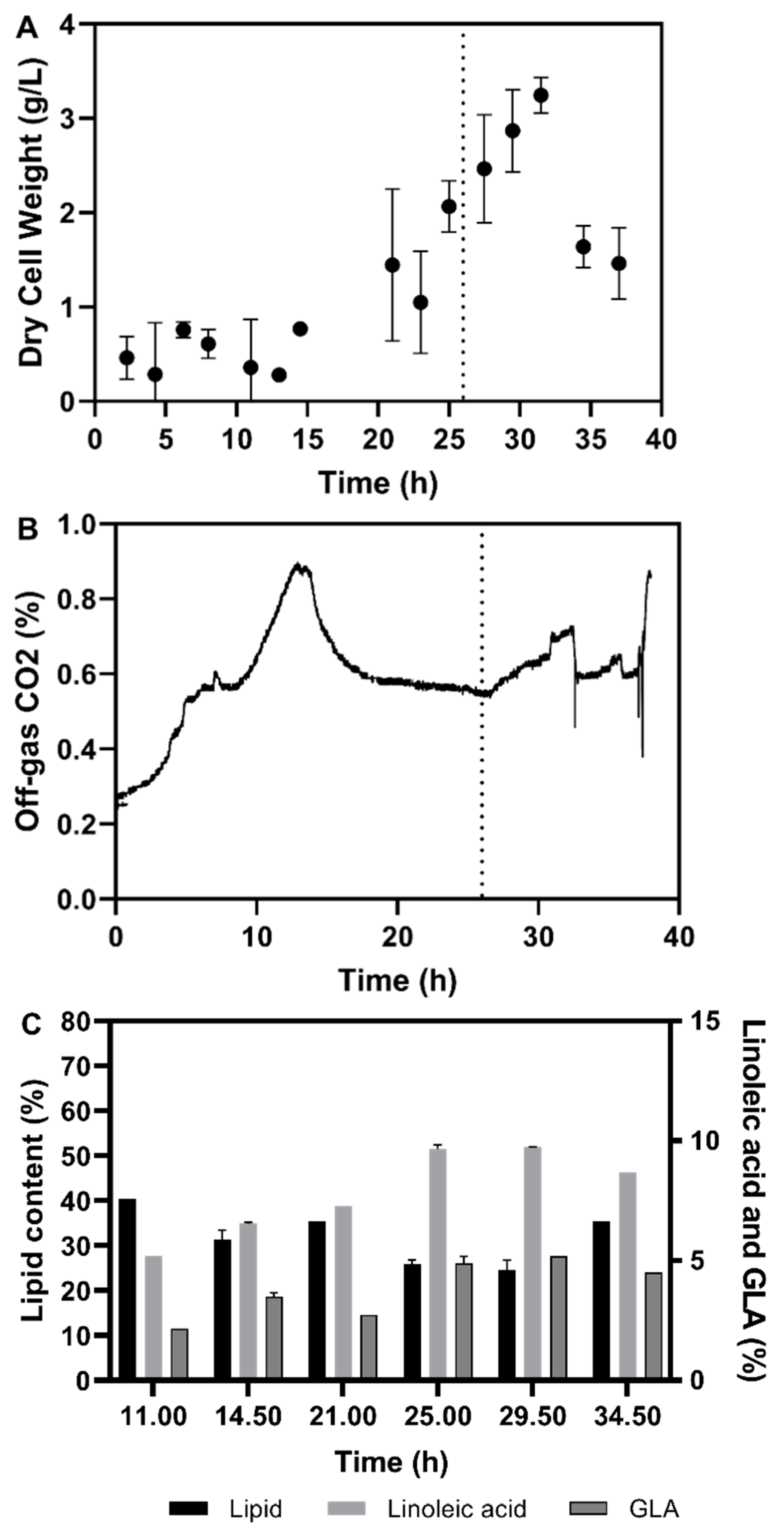
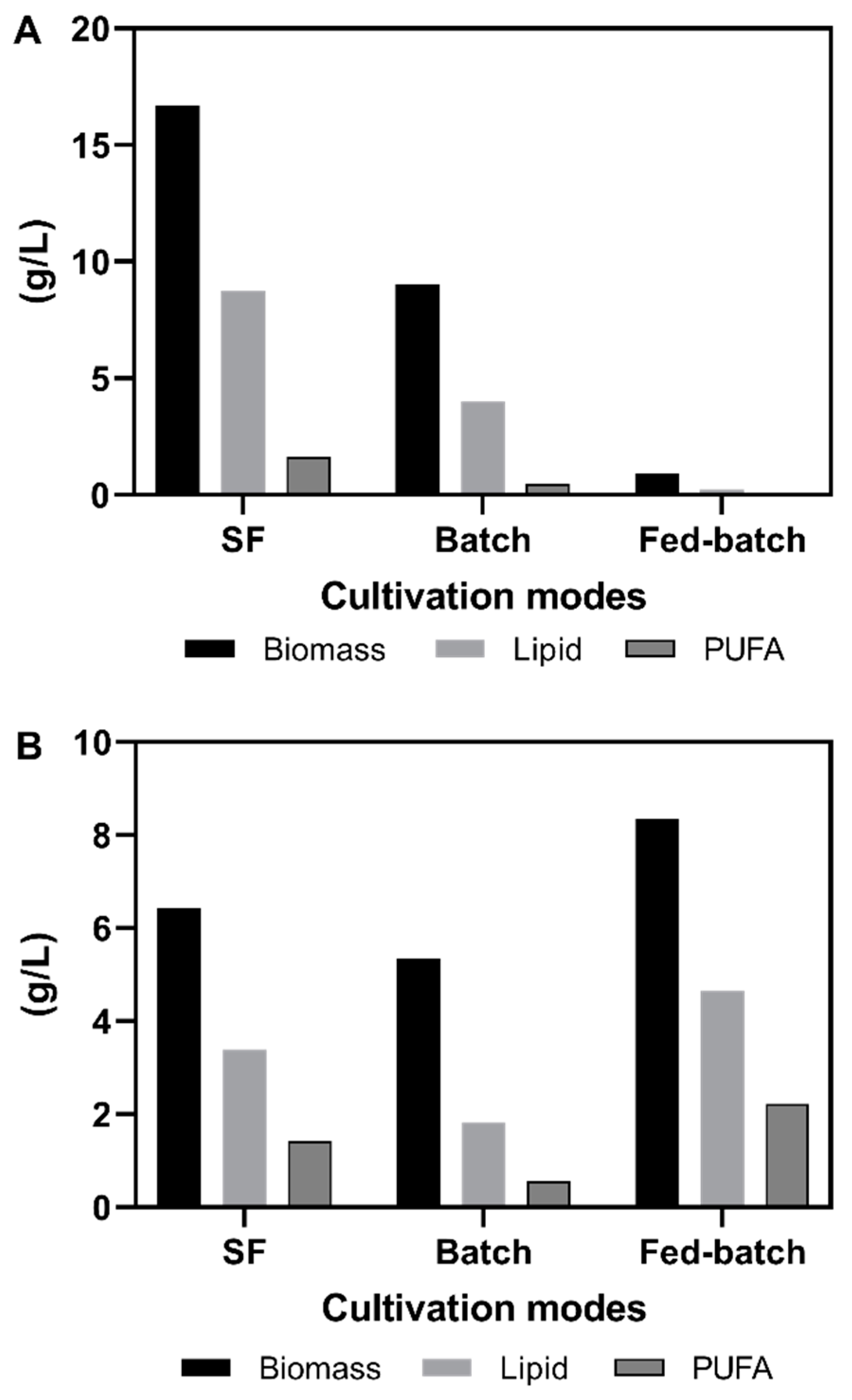
3.1. Shake Flask Cultivations
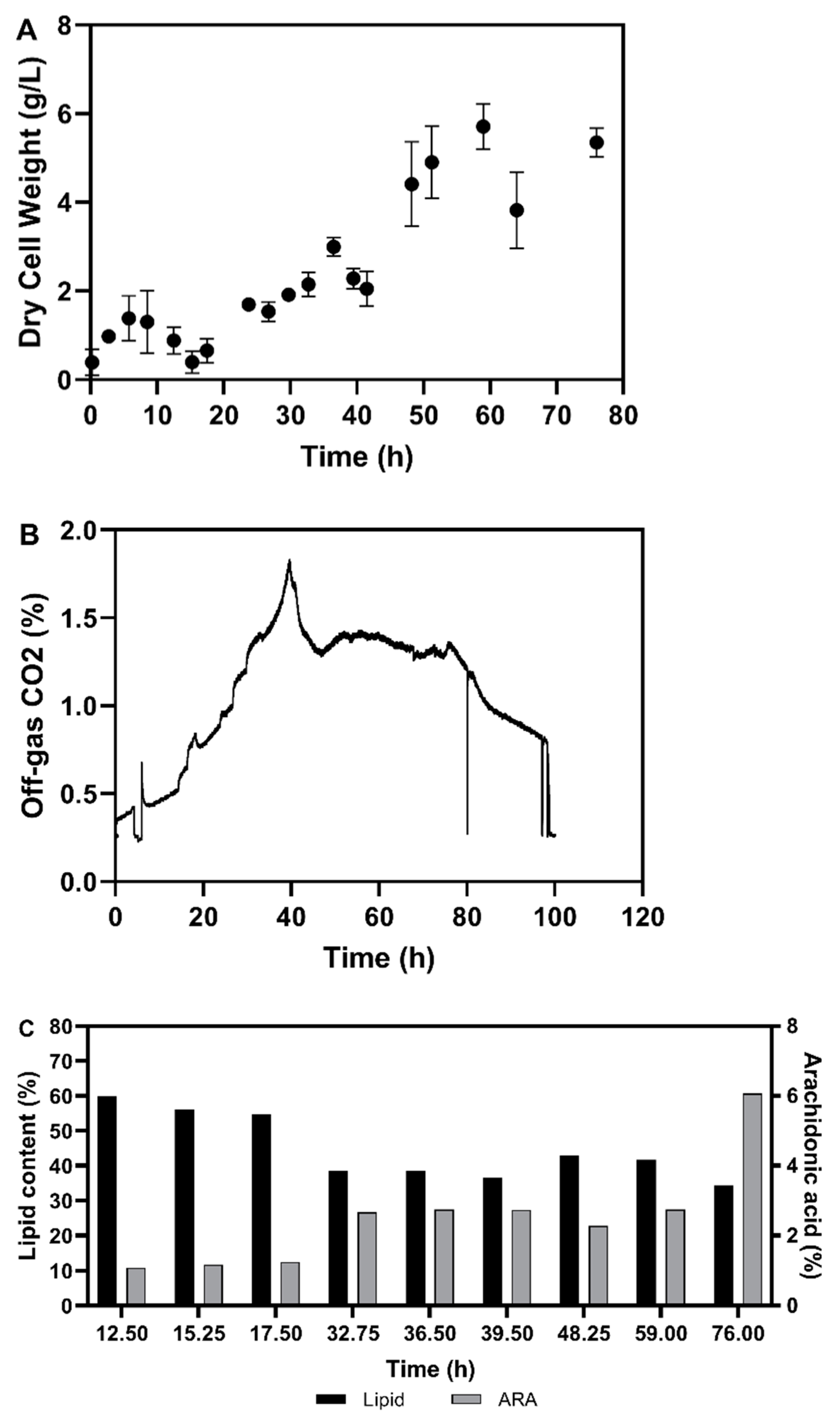
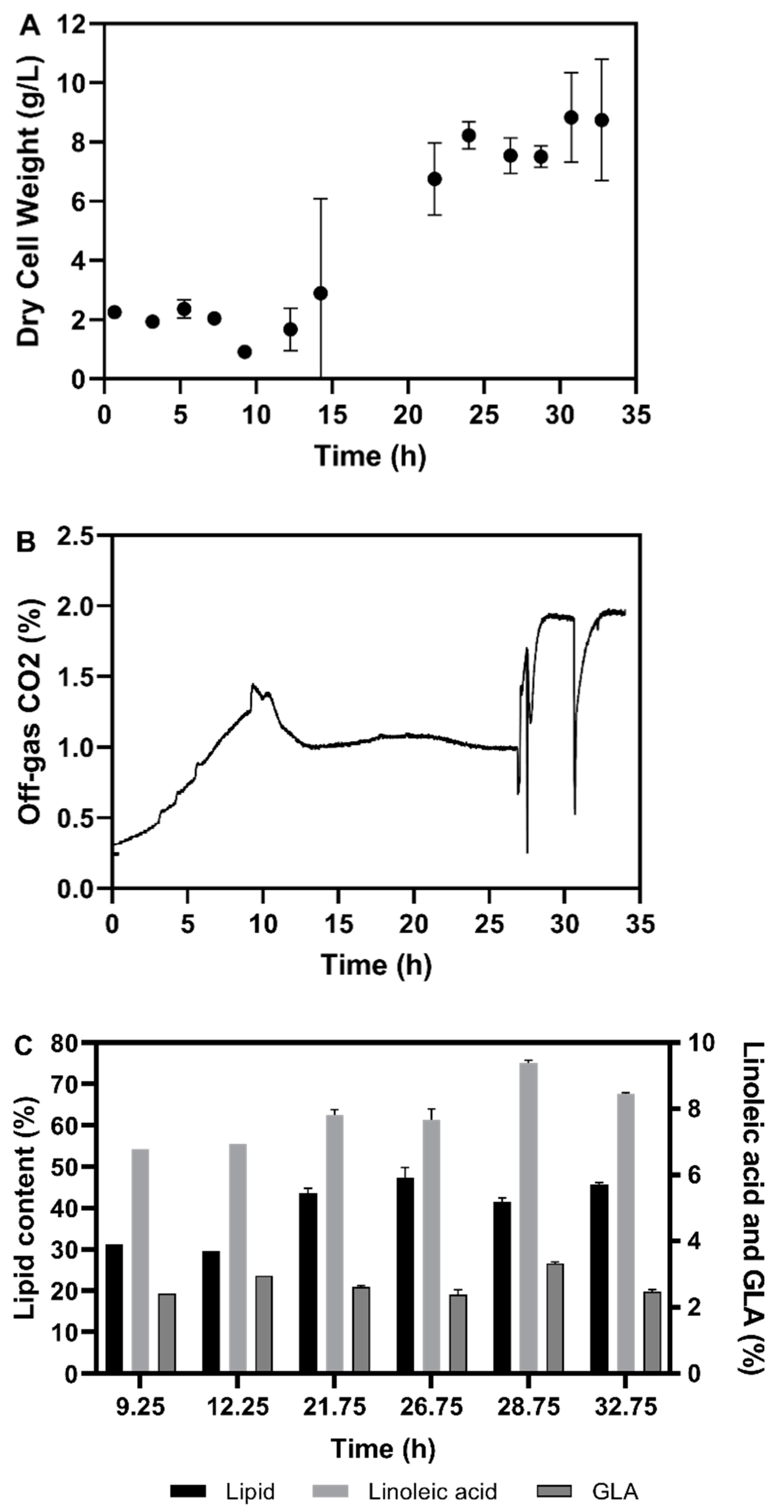
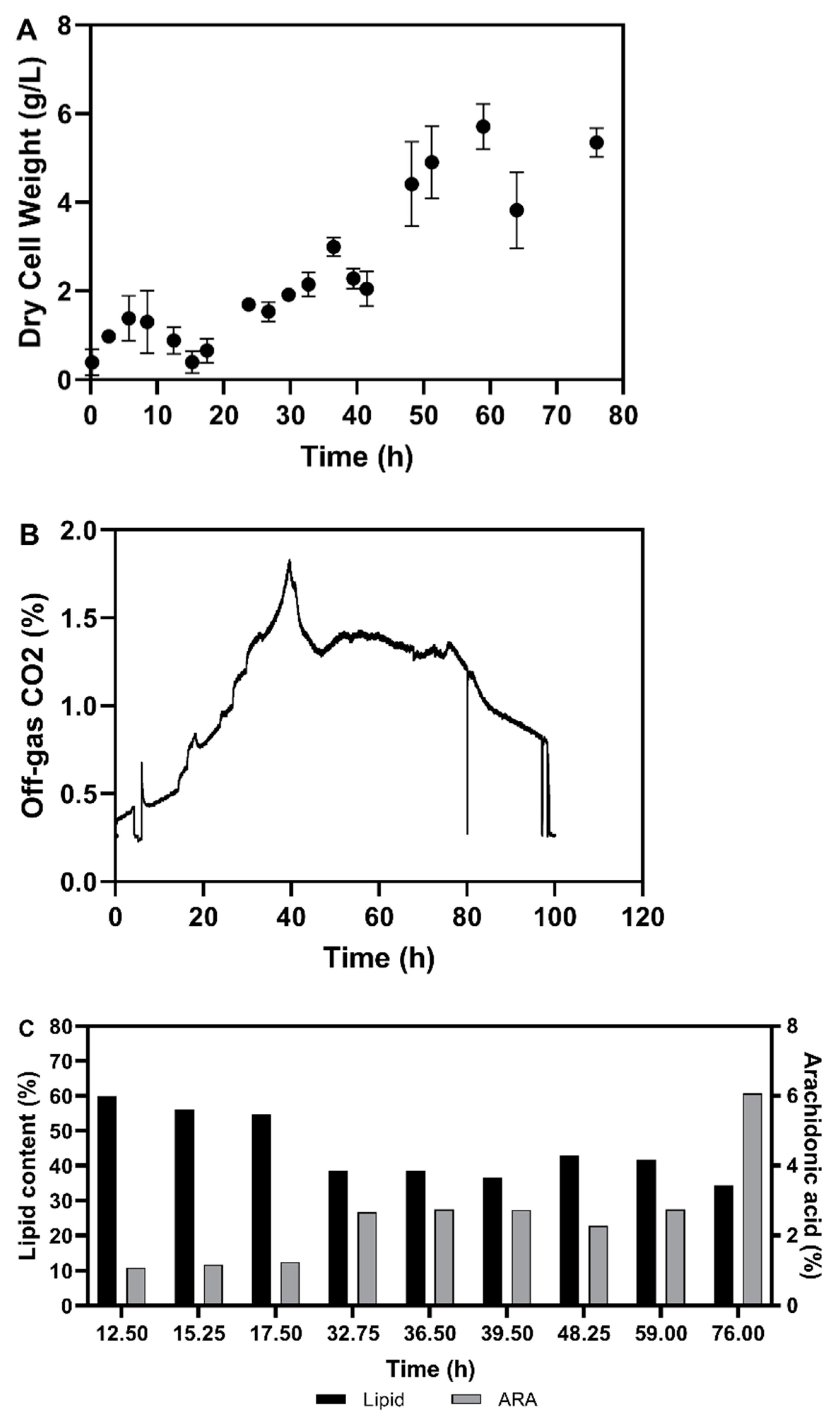
3.2. Fermentation Experiments Using Animal Fat By-Product Materials as C-Source
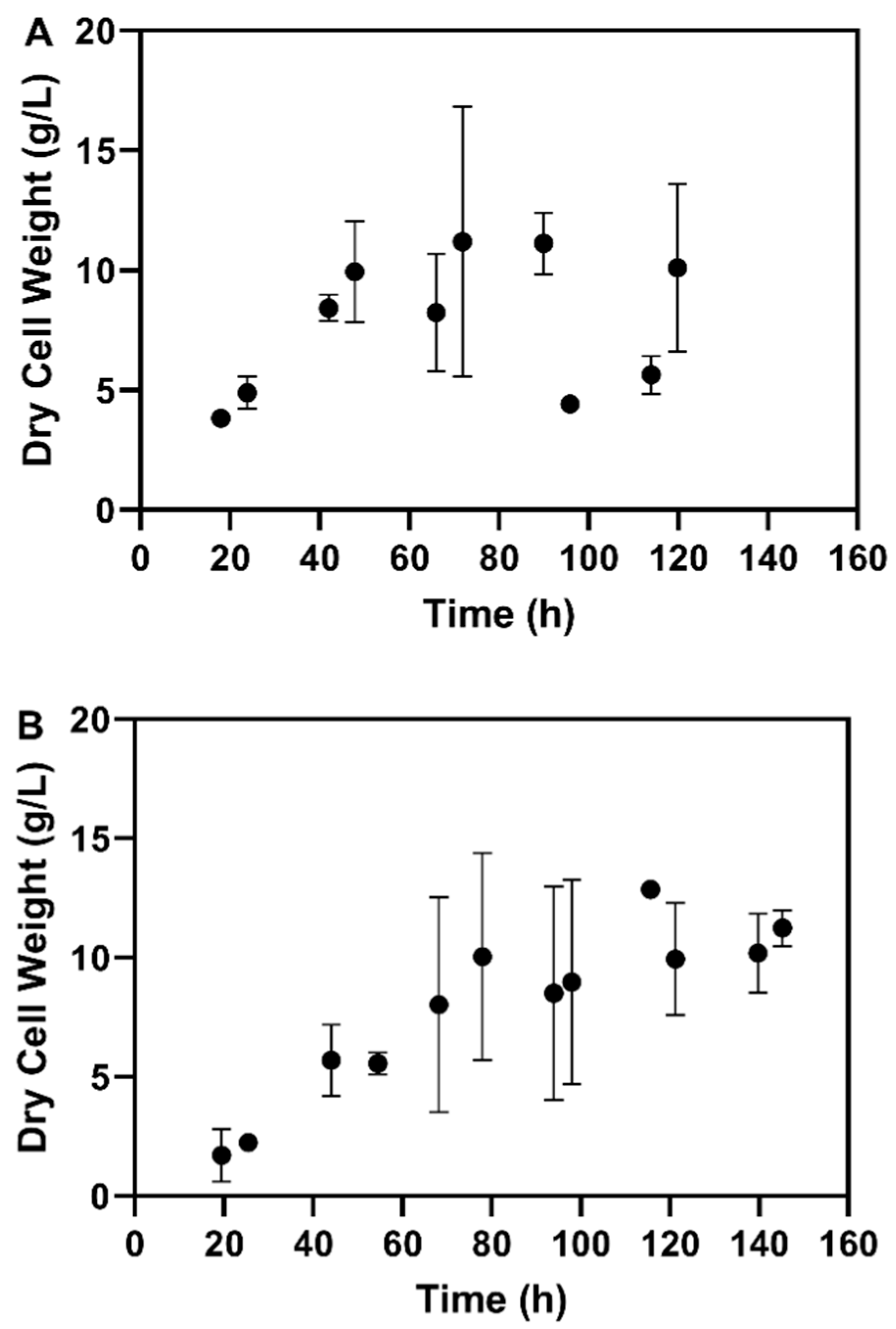
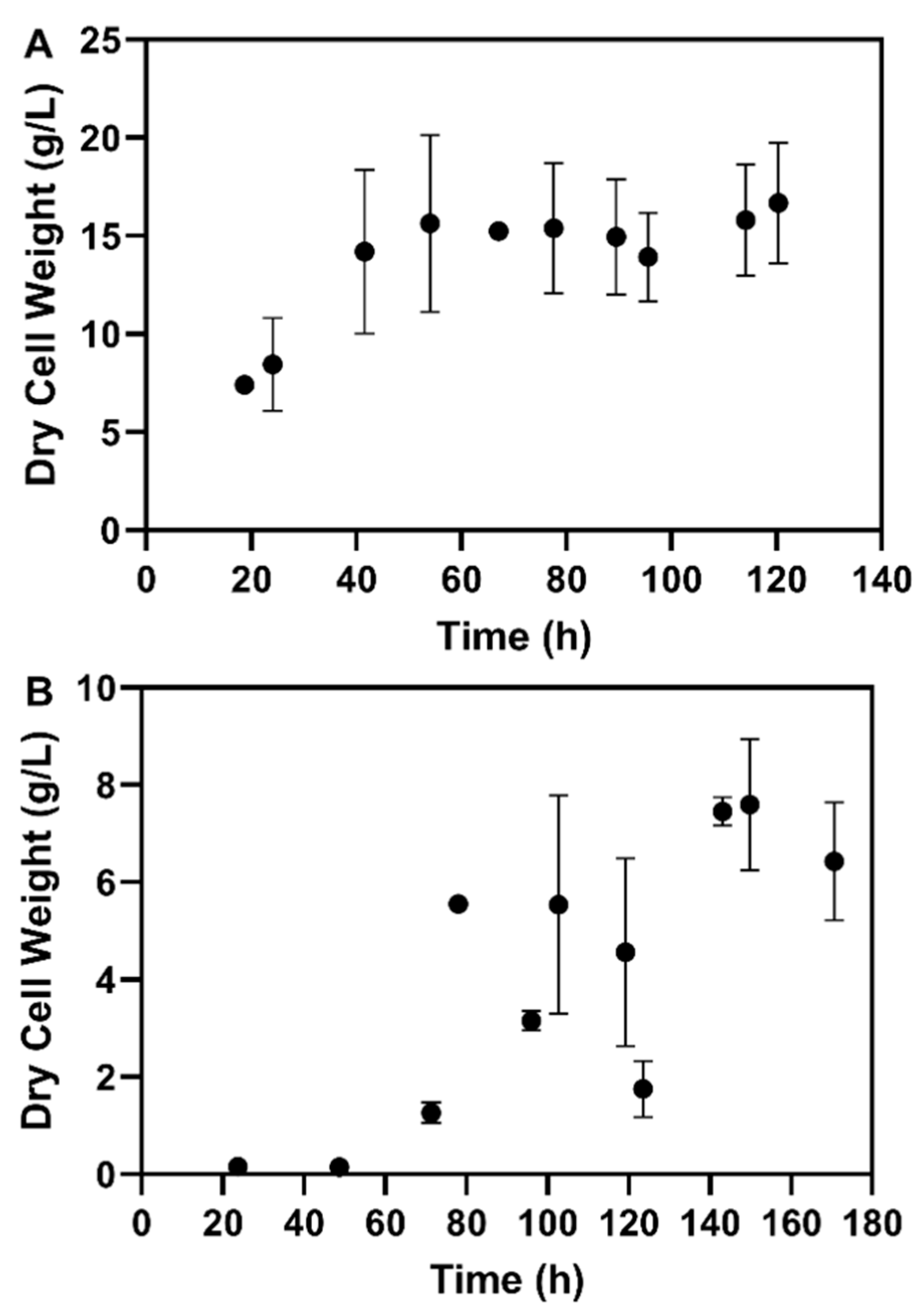
3.2.1. Fed-Batch Cultivation
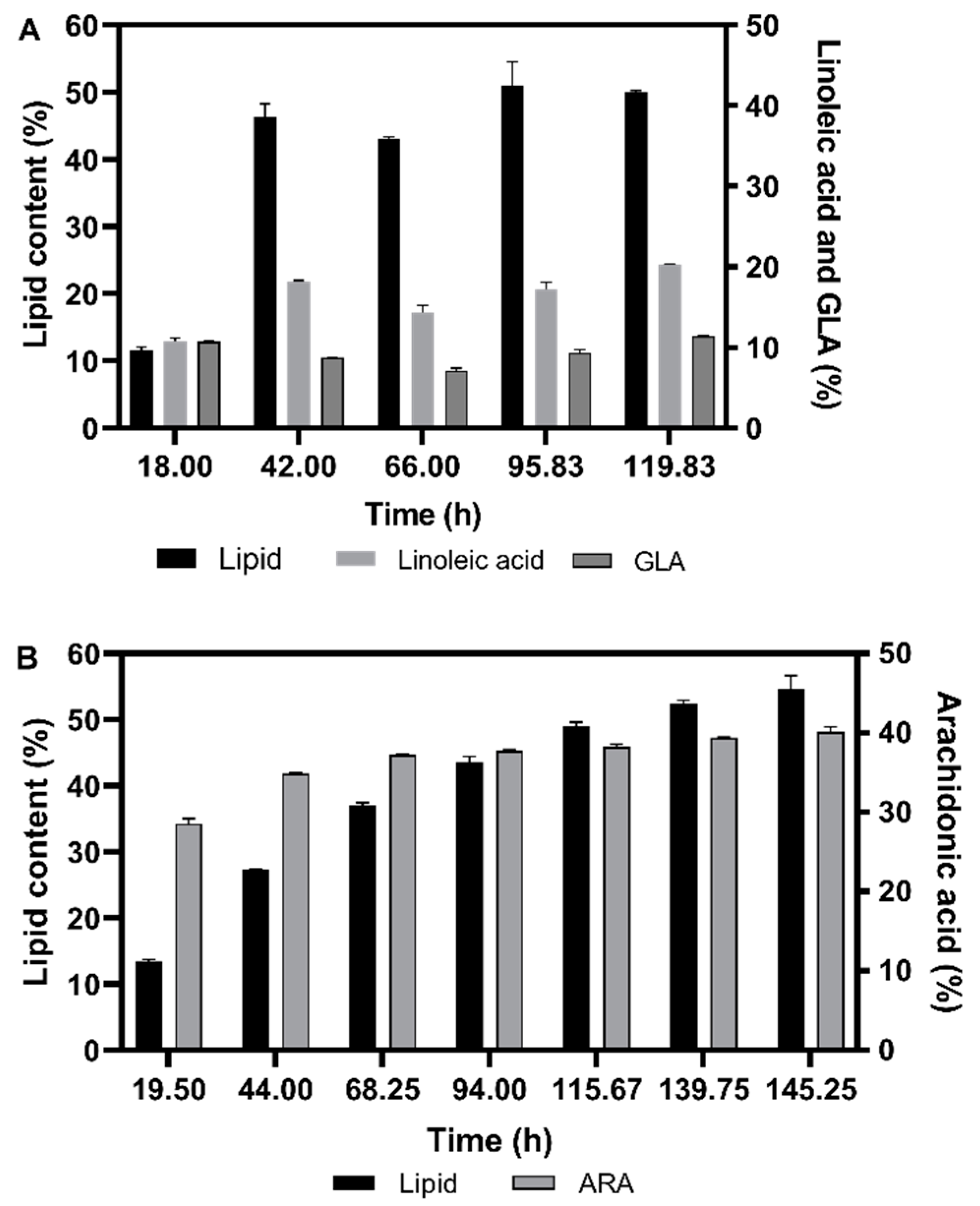
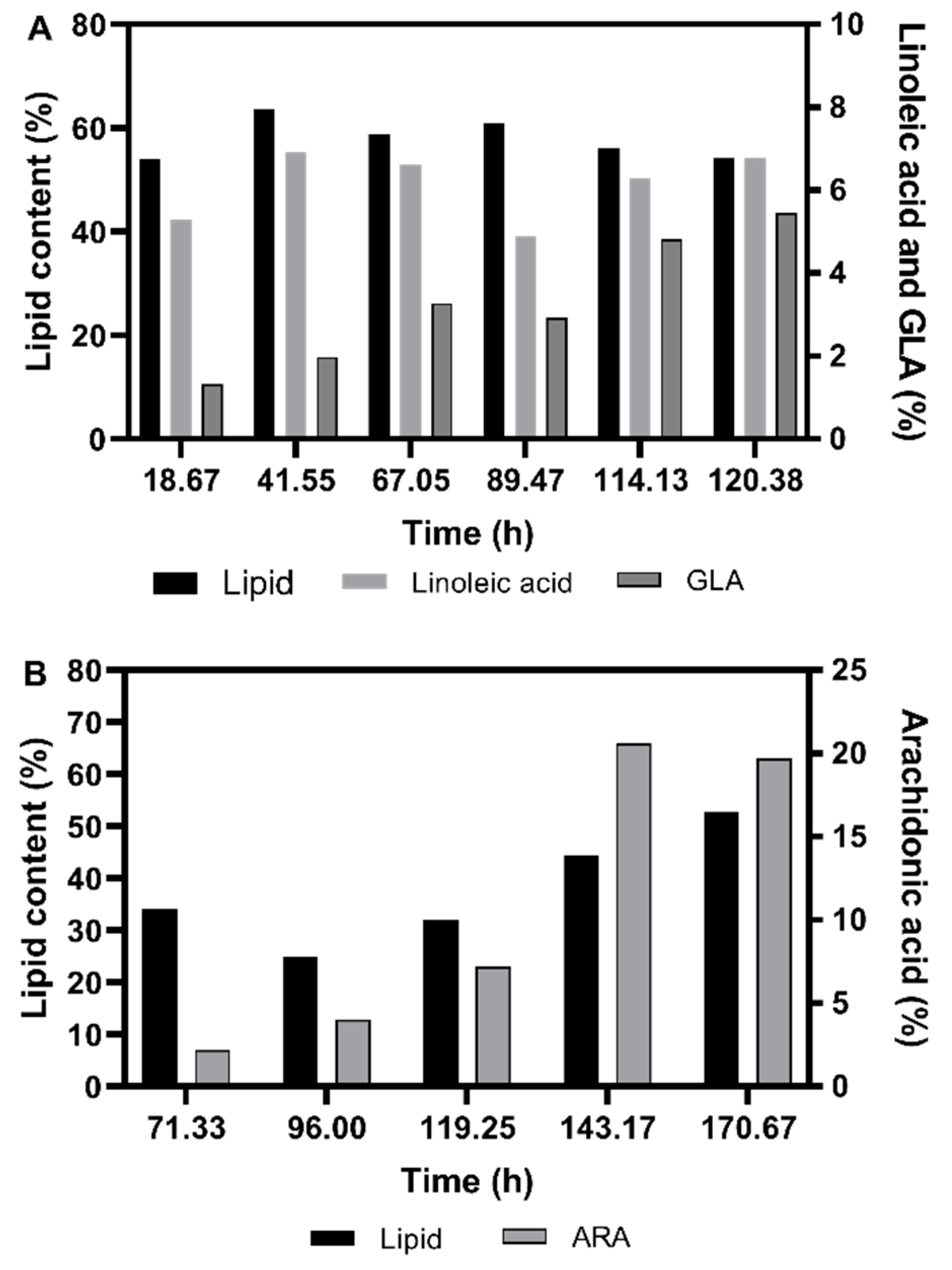
3.2.2. Fatty Acid Composition of the Single Cell Oil
3.3. Discussion
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- OECD/Food and Agriculture Organization of United Nations. Meat, OECD-FAO Agriculture Outlook 2021–2030; Food and Agriculture Organization of United Nations: Rome, Italy; OECD: Paris, France, 2021. [Google Scholar]
- Woodgate, S.L.; van der Veen, J.T. Fats and Oils—Animal Based. In Food Processing: Principles and Application, 2nd ed.; Clark, S., Jung, S., Lamsal, B., Eds.; John Wiley & Sons: Hoboken, NJ, USA, 2014. [Google Scholar]
- Marova, I.; Szotkowski, M.; Vanek, M.; Rapta, M.; Byrtusova, D.; Mikheichyk, N.; Haronikova, A.; Certik, M.; Shapaval, V. Utilization of animal fat waste as carbon source by carotenogenic yeasts—A screening study. EuroBiotech J. 2017, 1, 310–318. [Google Scholar] [CrossRef] [Green Version]
- Rosson, E.; Sgarbossa, P.; Pedrielli, F.; Mozzon, M.; Bertani, R. Bioliquids from raw waste animal fats: An alternative renewable energy source. Biomass Convers. Biorefinery 2020, 11, 1475–1490. [Google Scholar] [CrossRef]
- Toldrá-Reig, F.; Mora, L.; Toldrá, F. Trends in Biodiesel Production from Animal Fat Waste. Appl. Sci. 2020, 10, 3644. [Google Scholar] [CrossRef]
- Chakraborty, R.; Gupta, A.K.; Chowdhury, R. Conversion of slaughterhouse and poultry farm animal fats and wastes to biodiesel: Parametric sensitivity and fuel quality assessment. Renew. Sustain. Energy Rev. 2014, 29, 120–134. [Google Scholar] [CrossRef]
- Sai Akhil, U.; Alagumalai, A. A Short Review on Valorization of Slaughterhouse Wastes for Biodiesel Production. ChemistrySelect 2019, 4, 13356–13362. [Google Scholar] [CrossRef]
- Papanikolaou, S.; Aggelis, G. Lipids of oleaginous yeasts. Part I: Biochemistry of single cell oil production. Eur. J. Lipid Sci. Technol. 2011, 113, 1031–1051. [Google Scholar] [CrossRef]
- Adhikari, B.B.; Chae, M.; Bressler, D.C. Utilization of Slaughterhouse Waste in Value-Added Applications: Recent Advances in the Development of Wood Adhesives. Polymers 2018, 10, 176. [Google Scholar] [CrossRef] [Green Version]
- Papanikolaou, S.; Chevalot, I.; Komaitis, M.; Marc, I.; Aggelis, G. Single cell oil production by Yarrowia lipolytica growing on an industrial derivative of animal fat in batch cultures. Appl. Microbiol. Biotechnol. 2002, 58, 308–312. [Google Scholar] [CrossRef]
- El-Bialy, H.; Bullet, B.; Gomaa, O.; Khaled, B.; Azab, K. Conversion of oil waste to valuable fatty acids using Oleaginous yeast. J. Microbiol. Biotechnol. 2011, 27, 2791–2798. [Google Scholar] [CrossRef]
- Weiss, R.; Eischer, A.; Tadic, T.; Gritsch, S.M.; Ortner, M.; Prall, K.; Neunteufel, E.; Putz, R.F.; Guebitz, G.M.; Nyanhongo, G.S. Valorisation of slaughter house and deinking paper waste streams for the production of enzyme by Trichoderma reesei. J. Clean. Prod. 2020, 275, 122882. [Google Scholar] [CrossRef]
- Szotkowski, M.; Byrtusova, D.; Haronikova, A.; Vysoka, M.; Rapta, M.; Shapaval, V.; Marova, I. Study of Metabolic Adaptation of Red Yeasts to Waste Animal Fat Substrate. Microorganisms 2019, 7, 578. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Slaný, O.; Klempová, T.; Shapaval, V.; Zimmermann, B.; Kohler, A.; Čertík, M. Biotransformation of Animal Fat-By Products into ARA-Enriched Fermented Bioproducts by Solid-State Fermentation of Mortierella alpina. J. Fungi 2020, 6, 236. [Google Scholar] [CrossRef]
- Byrtusová, D.; Szotkowski, M.; Kurowska, K.; Shapaval, V.; Márová, I. Rhodotorula kratochvilovae CCY 20-2-26—The Source of Multifunctional Metabolites. Microorganisms 2021, 9, 1280. [Google Scholar] [CrossRef]
- Slaný, O.; Klempová, T.; Shapaval, V.; Zimmermann, B.; Kohler, A.; Čertík, M. Animal Fat as a Substrate for Production of n-6 Fatty Acids by Fungal Solid-State Fermentation. Microorganisms 2021, 9, 170. [Google Scholar] [CrossRef]
- Marova, I.; Rapta, M.; Vanek, M.; Haronikova, A.; Szotkowski, M.; Shapaval, V. Use of high-throughput techniques to study simultaneous production of lipid metabolites in carotenogenic yeasts grown on waste animal fat. J. Biotechnol. 2017, 256, S42. [Google Scholar] [CrossRef]
- Tzimorotas, D.; Afseth, N.K.; Lindberg, D.; Kjørlaug, O.; Axelsson, L.; Shapaval, V. Pretreatment of different food rest materials for bioconversion into fungal lipid-rich biomass. Bioprocess Biosyst. Eng. 2018, 41, 1039–1049. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kosa, G.; Kohler, A.; Tafintseva, V.; Zimmermann, B.; Forfang, K.; Afseth, N.K.; Tzimorotas, D.; Vuoristo, K.S.; Horn, S.J.; Mounier, J.; et al. Microtiter plate cultivation of oleaginous fungi and monitoring of lipogenesis by high-throughput FTIR spectroscopy. Microb. Cell Factories 2017, 16, 101. [Google Scholar] [CrossRef] [Green Version]
- Kosa, G.; Zimmermann, B.; Kohler, A.; Ekeberg, D.; Afseth, N.K.; Mounier, J.; Shapaval, V. High-throughput screening of Mucoromycota fungi for production of low- and high-value lipids. Biotechnol. Biofuels 2018, 11, 66. [Google Scholar] [CrossRef]
- Dzurendova, S.; Zimmermann, B.; Tafintseva, V.; Kohler, A.; Ekeberg, D.; Shapaval, V. The influence of phosphorus source and the nature of nitrogen substrate on the biomass production and lipid accumulation in oleaginous Mucoromycota fungi. Appl. Microbiol. Biotechnol. 2020, 104, 8065–8076. [Google Scholar] [CrossRef]
- Dzurendova, S.; Zimmermann, B.; Kohler, A.; Tafintseva, V.; Slany, O.; Certik, M.; Shapaval, V. Microcultivation and FTIR spectroscopy-based screening revealed a nutrient-induced co-production of high-value metabolites in oleaginous Mucoromycota fungi. PLoS ONE 2020, 15, e0234870. [Google Scholar] [CrossRef] [PubMed]
- Dzurendova, S.; Zimmermann, B.; Tafintseva, V.; Kohler, A.; Horn, S.J.; Shapaval, V. Metal and Phosphate Ions Show Remarkable Influence on the Biomass Production and Lipid Accumulation in Oleaginous Mucor circinelloides. J. Fungi 2020, 6, 260. [Google Scholar] [CrossRef]
- Dzurendova, S.; Zimmermann, B.; Kohler, A.; Reitzel, K.; Nielsen, U.G.; Dupuy-Galet, B.X.; Leivers, S.; Horn, S.J.; Shapaval, V. Calcium Affects Polyphosphate and Lipid Accumulation in Mucoromycota Fungi. J. Fungi 2021, 7, 300. [Google Scholar] [CrossRef]
- Athenaki, M.; Gardeli, C.; Diamantopoulou, P.; Tchakouteu, S.S.; Sarris, D.; Philippoussis, A.; Papanikolaou, S. Lipids from yeasts and fungi: Physiology, production and analytical considerations. J. Appl. Microbiol. 2018, 124, 336–367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caporusso, A.; Capece, A.; De Bari, I. Oleaginous Yeasts as Cell Factories for the Sustainable Production of Microbial Lipids by the Valorization of Agri-Food Wastes. Fermentation 2021, 7, 50. [Google Scholar] [CrossRef]
- Ratledge, C. Fatty acid biosynthesis in microorganisms being used for Single Cell Oil production. Biochimie 2004, 86, 807–815. [Google Scholar] [CrossRef] [PubMed]
- Mhlongo, S.I.; Ezeokoli, O.T.; Roopnarain, A.; Ndaba, B.; Sekoai, P.T.; Habimana, O.; Pohl, C.H. The Potential of Single-Cell Oils Derived From Filamentous Fungi as Alternative Feedstock Sources for Biodiesel Production. Front. Microbiol. 2021, 12, 57. [Google Scholar] [CrossRef]
- Patel, A.; Karageorgou, D.; Rova, E.; Katapodis, P.; Rova, U.; Christakopoulos, P.; Matsakas, L. An Overview of Potential Oleaginous Microorganisms and Their Role in Biodiesel and Omega-3 Fatty Acid-Based Industries. Microorganisms 2020, 8, 434. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McIntyre, M.; Breum, J.; Arnau, J.; Nielsen, J. Growth physiology and dimorphism of Mucor circinelloides (syn. racemosus) during submerged batch cultivation. Appl. Microbiol. Biotechnol. 2002, 58, 495–502. [Google Scholar] [CrossRef] [PubMed]
- Mironov, A.A.; Nemashkalov, V.A.; Stepanova, N.N.; Kamzolova, S.V.; Rymowicz, W.; Morgunov, I.G. The Effect of pH and Temperature on Arachidonic Acid Production by Glycerol-Grown Mortierella alpina NRRL-A-10995. Fermentation 2018, 4, 17. [Google Scholar] [CrossRef] [Green Version]
- Kosa, G.; Vuoristo, K.S.; Horn, S.J.; Zimmermann, B.; Afseth, N.K.; Kohler, A.; Shapaval, V. Assessment of the scalability of a microtiter plate system for screening of oleaginous microorganisms. Appl. Microbiol. Biotechnol. 2018, 102, 4915–4925. [Google Scholar] [CrossRef] [Green Version]
- Lewis, T.; Nichols, P.D.; McMeekin, T.A. Evaluation of extraction methods for recovery of fatty acids from lipid-producing microheterotrophs. J. Microbiol. Methods 2000, 43, 107–116. [Google Scholar] [CrossRef]
- Langseter, A.M.; Dzurendova, S.; Shapaval, V.; Kohler, A.; Ekeberg, D.; Zimmermann, B. Evaluation and optimisation of direct transesterification methods for the assessment of lipid accumulation in oleaginous filamentous fungi. Microb. Cell Factories 2021, 20, 59. [Google Scholar] [CrossRef]
- Eroshin, V.K.; Satroutdinov, A.D.; Dedyukhina, E.G.; Chistyakova, T.I. Arachidonic acid production by Mortierella alpina with growth-coupled lipid synthesis. Process. Biochem. 2000, 35, 1171–1175. [Google Scholar] [CrossRef]
- Heeres, A.S.; Picone, C.S.F.; van der Wielen, L.A.M.; Cunha, R.L.; Cuellar, M.C. Microbial advanced biofuels production: Overcoming emulsification challenges for large-scale operation. Trends Biotechnol. 2014, 32, 221–229. [Google Scholar] [CrossRef]
- Wynn, J.P.; Hamid, A.A.; Li, Y.; Ratledge, C. Biochemical events leading to the diversion of carbon into storage lipids in the oleaginous fungi Mucor circinelloides and Mortierella alpina. Microbiology 2001, 147, 2857–2864. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Papanikolaou, S.; Galiotou-Panayotou, M.; Fakas, S.; Komaitis, M.; Aggelis, G. Lipid production by oleaginous Mucorales cultivated on renewable carbon sources. Eur. J. Lipid Sci. Technol. 2007, 109, 1060–1070. [Google Scholar] [CrossRef]
- Chen, H.; Hao, G.; Wang, L.; Wang, H.; Gu, Z.; Liu, L.; Zhang, H.; Chen, W.; Chen, Y.Q. Identification of a critical determinant that enables efficient fatty acid synthesis in oleaginous fungi. Sci. Rep. 2015, 5, 11247. [Google Scholar] [CrossRef]
- Kavadia, A.; Komaitis, M.; Chevalot, I.; Blanchard, F.; Marc, I.; Aggelis, G. Lipid and γ-linolenic acid accumulation in strains of zygomycetes growing on glucose. J. Am. Oil Chem. Soc. 2001, 78, 341–346. [Google Scholar] [CrossRef]
- Jang, H.-D.; Lin, Y.-Y.; Yang, S.-S. Effect of culture media and conditions on polyunsaturated fatty acids production by Mortierella alpina. Bioresour. Technol. 2005, 96, 1633–1644. [Google Scholar] [CrossRef] [PubMed]
- Xia, C.; Zhang, J.; Zhang, W.; Hu, B. A new cultivation method for microbial oil production: Cell pelletization and lipid accumulation by Mucor circinelloides. Biotechnol. Biofuels 2011, 4, 15. [Google Scholar] [CrossRef] [Green Version]
- Tang, X.; Zan, X.; Zhao, L.; Chen, H.; Chen, Y.Q.; Chen, W.; Song, Y.; Ratledge, C. Proteomics analysis of high lipid-producing strain Mucor circinelloides WJ11: An explanation for the mechanism of lipid accumulation at the proteomic level. Microb. Cell Factories 2016, 15, 35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagao Totani, A.Y.; Yawata, M.; Ueda, T. The Role of Morphology during Growth of Mortierella alpina in Arachidonic Acid Production. J. Oleo Sci. 2002, 51, 531–538. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Time (h) | C14:0 | C16:0 | C16:1 | C17:0 | C18:0 | C18:1n9c | C18:2n6c | C18:3n6 | C18:3n3 | Other | Total Lipid Content (wt%) | ||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| SF | 50 mL | 120.38 | 0.50 | 19.87 | 0.95 | 1.18 | 29.28 | 32.04 | 6.77 | 5.45 | 0.00 | 3.96 | 54.14 |
| Batch | 8.5 L | 32.75 | 1.17 | 21.12 | 1.81 | 0.82 | 17.47 | 43.23 | 8.46 | 2.48 | 0.69 | 2.75 | 45.61 |
| Fed-batch | 8.0 L | 34.5 | 1.71 | 22.69 | 2.35 | 0.92 | 15.98 | 38.54 | 8.68 | 4.51 | 0.83 | 3.78 | 28.44 |
| Time (h) | C14:0 | C16:0 | C16:1 | C18:0 | C18:1n9c | C18:2n6c | C18:3n6 | C18:3n3 | C20:3n6 | C20:4n6 | C24:0 | Other | Total Lipid Content (wt%) | ||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| SF | 50 mL | 170.67 | 1.58 | 18.91 | 1.73 | 16.46 | 25.80 | 5.24 | 2.48 | 0.55 | 0.00 | 19.72 | 1.42 | 6.11 | 52.67 |
| Batch | 8.5 L | 76 | 2.02 | 21.01 | 2.53 | 14.31 | 36.95 | 6.21 | 4.49 | 0.85 | 0.54 | 6.07 | 0.58 | 4.43 | 34.27 |
| Fed-batch | 8.0 L | 130.33 | 1.41 | 16.54 | 1.64 | 12.15 | 27.02 | 5.32 | 2.77 | 0.59 | 1.50 | 23.80 | 0.75 | 4.68 | 55.77 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gaykawad, S.S.; Ramanand, S.S.; Blomqvist, J.; Zimmermann, B.; Shapaval, V.; Kohler, A.; Oostindjer, M.; Boccadoro, C. Submerged Fermentation of Animal Fat By-Products by Oleaginous Filamentous Fungi for the Production of Unsaturated Single Cell Oil. Fermentation 2021, 7, 300. https://doi.org/10.3390/fermentation7040300
Gaykawad SS, Ramanand SS, Blomqvist J, Zimmermann B, Shapaval V, Kohler A, Oostindjer M, Boccadoro C. Submerged Fermentation of Animal Fat By-Products by Oleaginous Filamentous Fungi for the Production of Unsaturated Single Cell Oil. Fermentation. 2021; 7(4):300. https://doi.org/10.3390/fermentation7040300
Chicago/Turabian StyleGaykawad, Sushil S., Sreerekha S. Ramanand, Johanna Blomqvist, Boris Zimmermann, Volha Shapaval, Achim Kohler, Marije Oostindjer, and Catherine Boccadoro. 2021. "Submerged Fermentation of Animal Fat By-Products by Oleaginous Filamentous Fungi for the Production of Unsaturated Single Cell Oil" Fermentation 7, no. 4: 300. https://doi.org/10.3390/fermentation7040300
APA StyleGaykawad, S. S., Ramanand, S. S., Blomqvist, J., Zimmermann, B., Shapaval, V., Kohler, A., Oostindjer, M., & Boccadoro, C. (2021). Submerged Fermentation of Animal Fat By-Products by Oleaginous Filamentous Fungi for the Production of Unsaturated Single Cell Oil. Fermentation, 7(4), 300. https://doi.org/10.3390/fermentation7040300