
Extractive Fermentation for Recovery of Bacteriocin-Like Inhibitory Substances Derived from Lactococcus lactis Gh1 Using PEG2000/Dextran T500 Aqueous Two-Phase System

 ,
,  ,
,  ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Media and Culture Conditions
2.2. Effect of Different Phase-Forming Reagents on BLIS Production and Bacterial Cell Growth
2.3. Partitioning Behavior of BLIS in ATPS
2.4. Effect of Orbital Agitation and pH on Partitioning Performance of BLIS
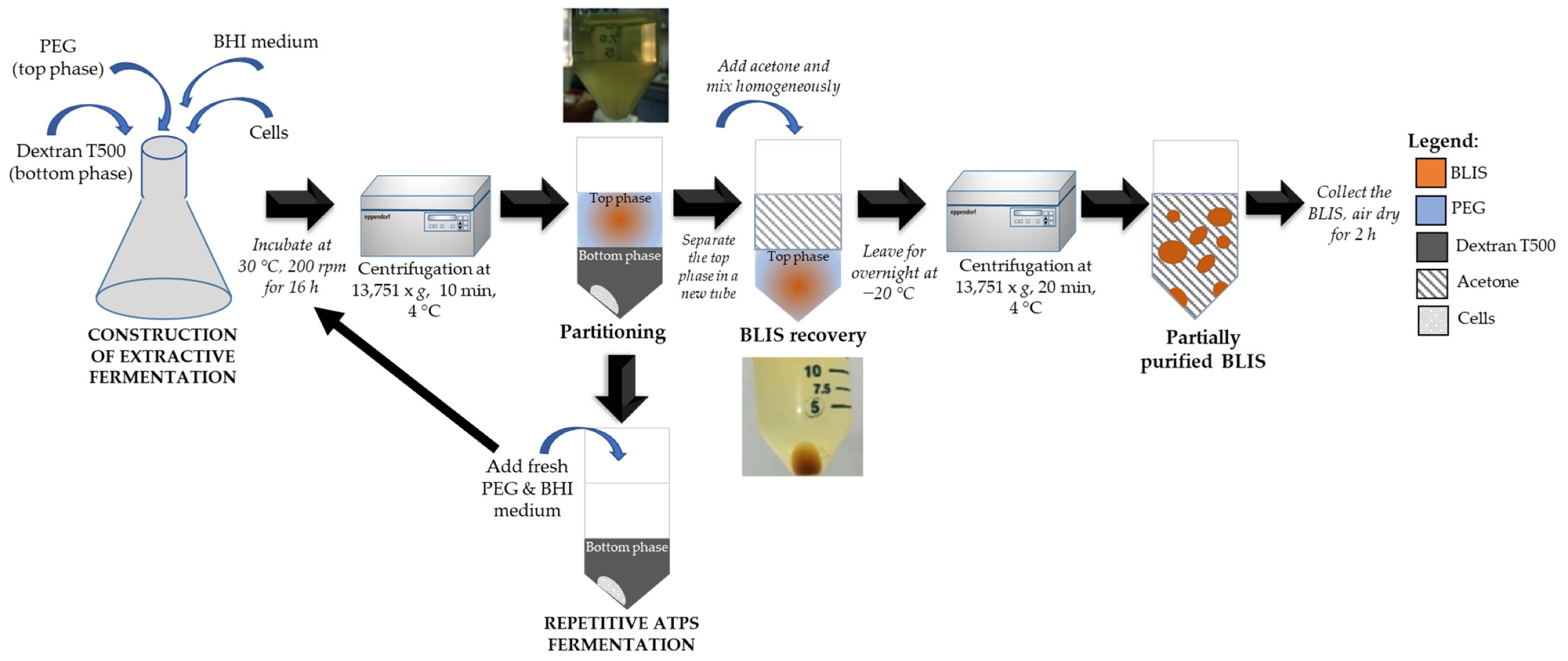
2.5. Scale-Up of ATPS Extractive Fermentation to 2 L Stirred Tank Bioreactor
2.6. Repetitive Batch of ATPS Extractive Fermentation
2.7. Analytical Methods
2.8. Determination of Relative Value, Volume Ratio, Partition Coefficient, Specific Activity, Selectivity, and Yield
3. Results
3.1. Selection of Polymer and Salt for Construction of ATPS and Their Influence on BLIS Production and Bacterial Cell Growth
3.2. Influencing Factors on Partitioning Behavior of BLIS in ATPS
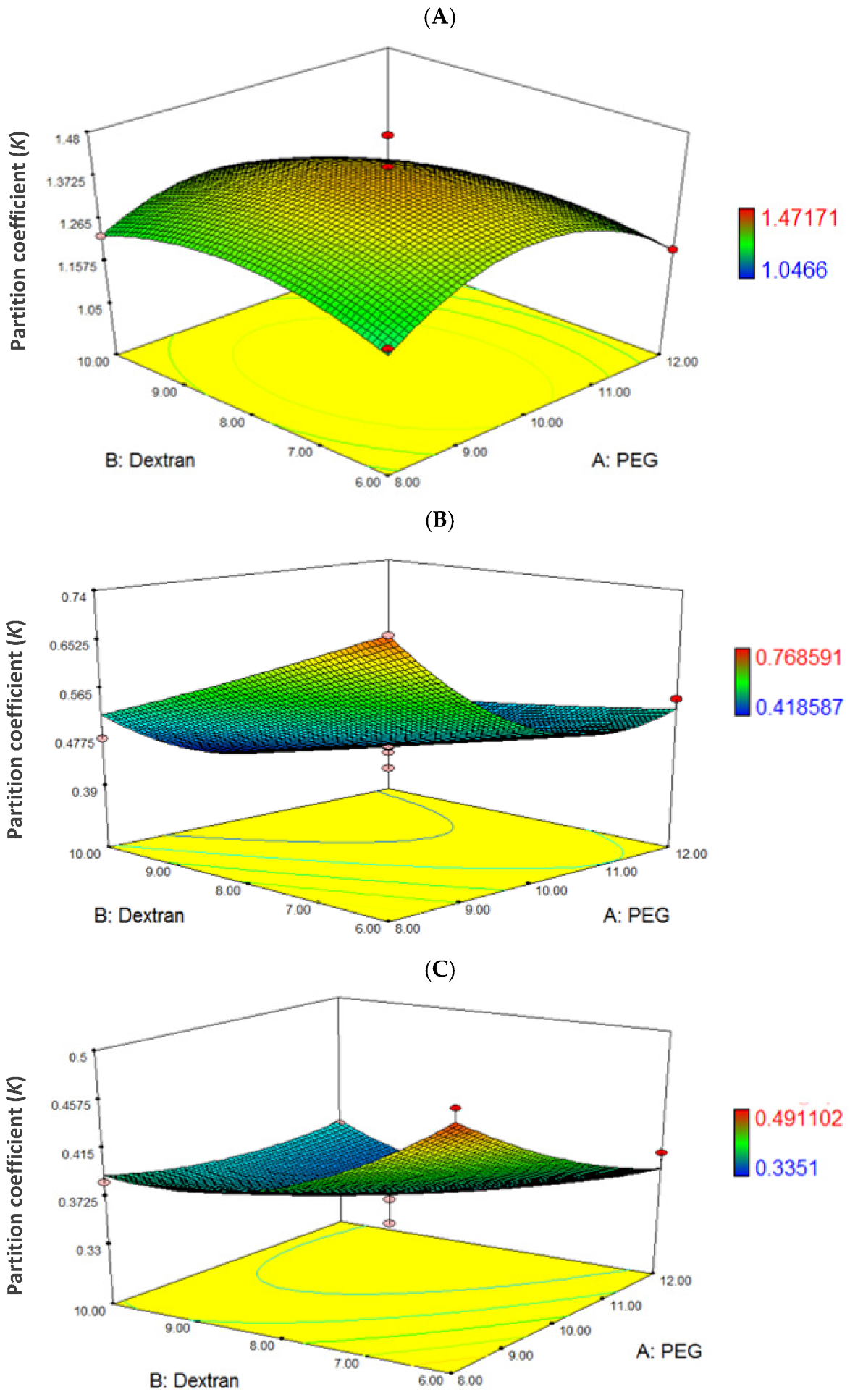
3.3. Effects of Model Parameters and Their Interactions
3.4. The Prediction of the Optimum Condition of Partition Coefficient (K)
3.5. Effect of Orbital Agitation and pH on Partitioning of BLIS
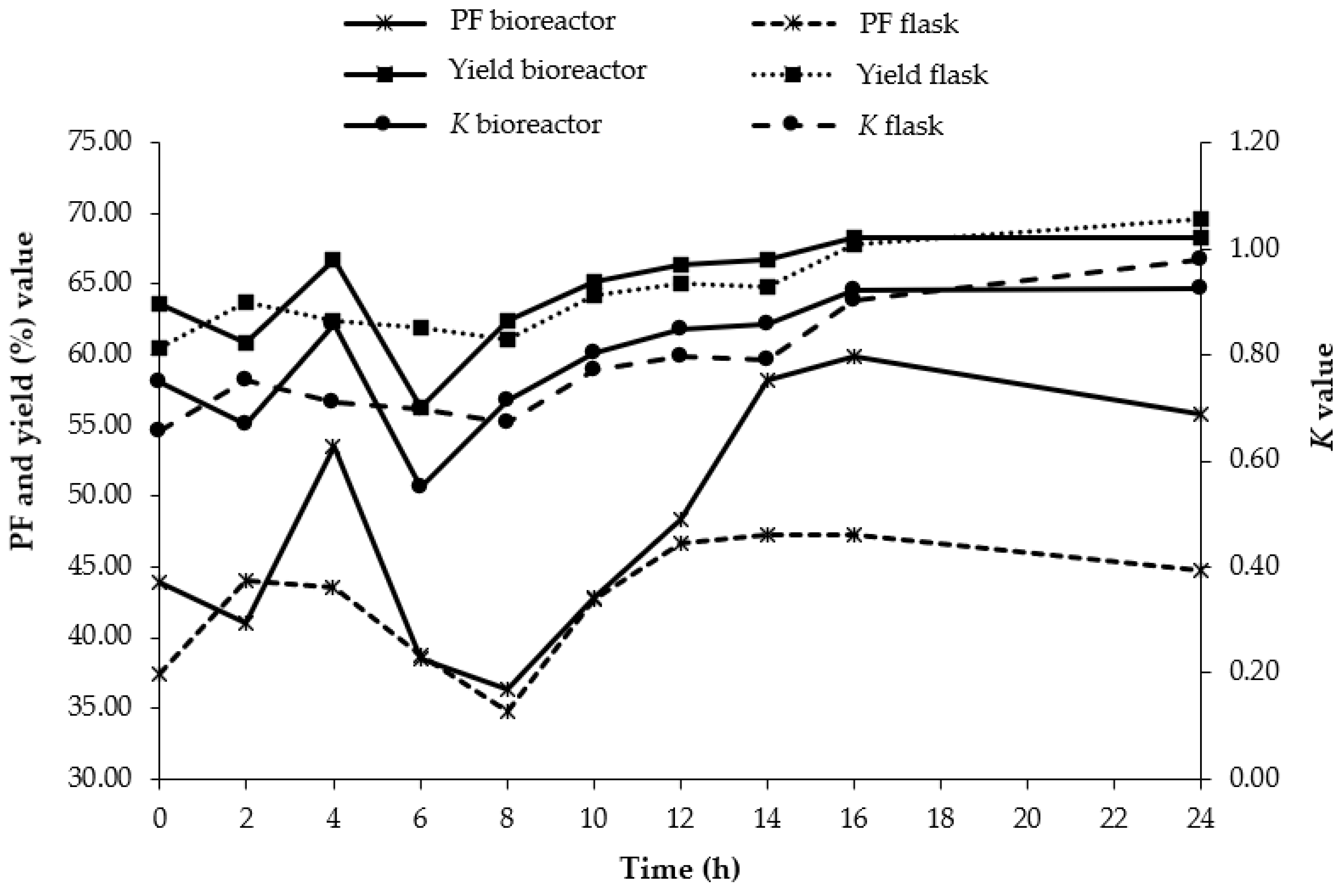
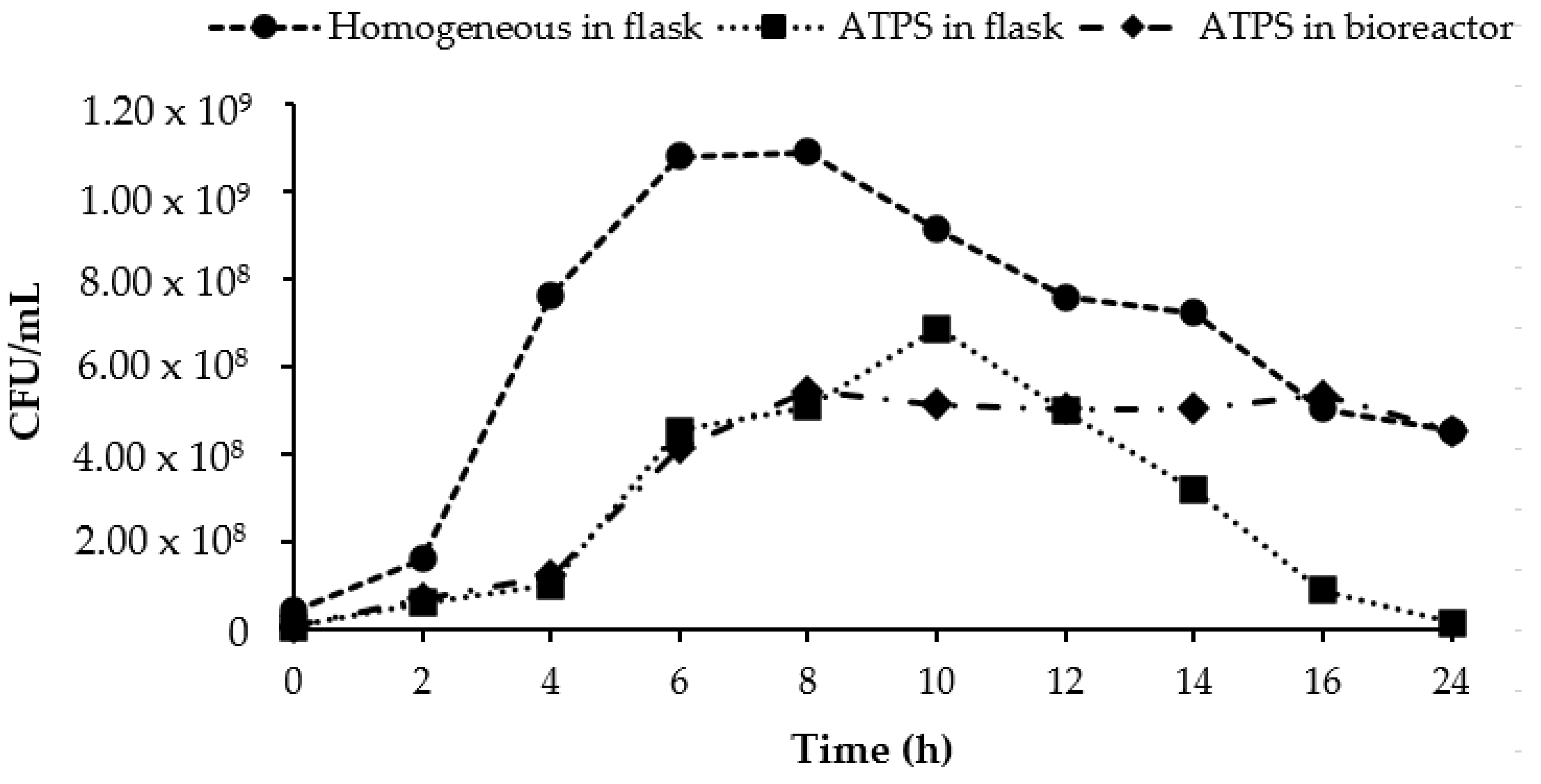
3.6. Scale-Up of ATPS in 2 L Stirred Tank Bioreactor
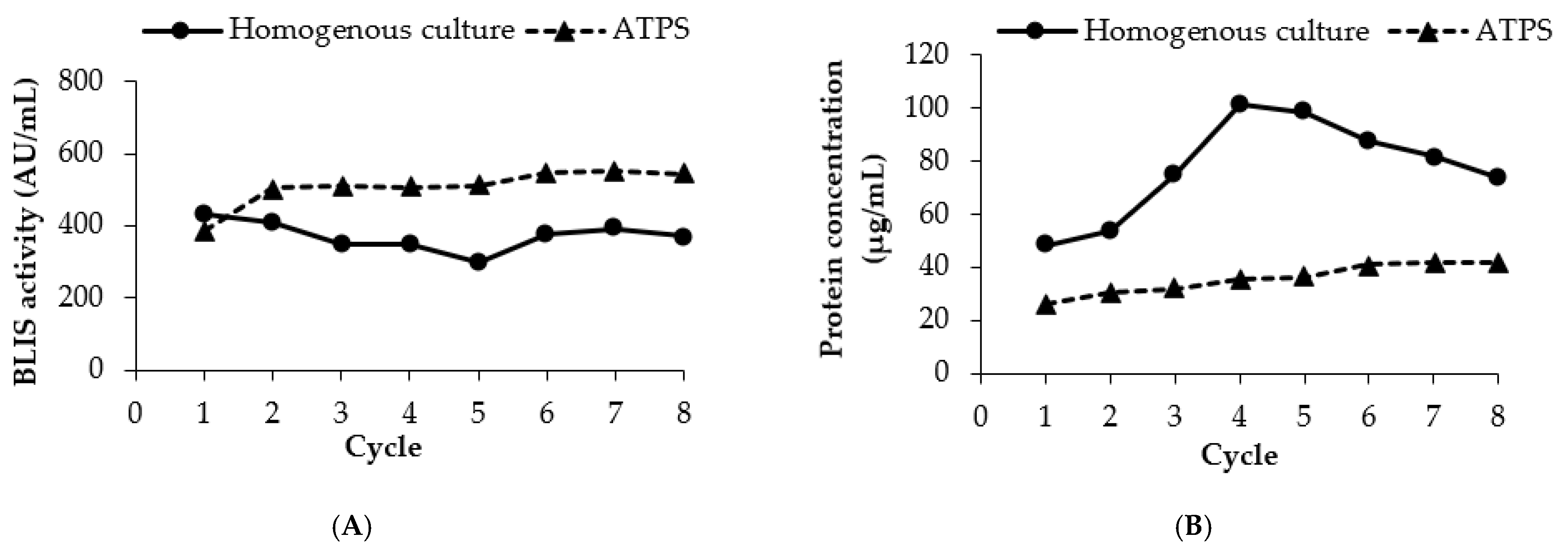
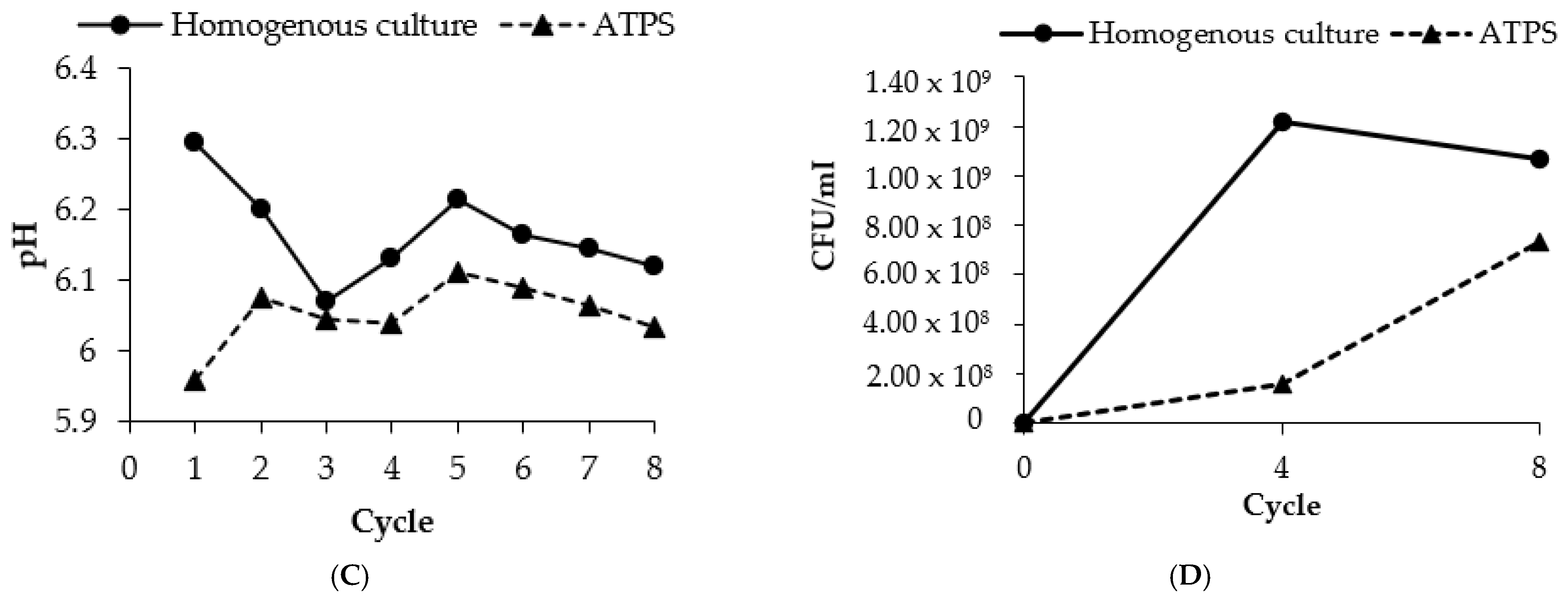
3.7. Repetitive ATPS Fermentation in an Erlenmeyer Flask
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Faour-Klingbeil, D.; Todd, E.C.D. Prevention and control of foodborne diseases in middle-east north African countries: Review of national control systems. Int. J. Environ. Res. Public Health 2020, 17, 70. [Google Scholar] [CrossRef] [PubMed]
- Reis, J.A.U.; Paula, A.T.U.; Casarotti, S.; Penna, A.L.B.U. Lactic Acid Bacteria Antimicrobial Compounds: Characteristics and Applications. Food Eng. Rev. 2012, 4, 124–140. [Google Scholar] [CrossRef]
- Sidek, N.L.M.; Halim, M.; Tan, J.S.; Abbasiliasi, S.; Mustafa, S.; Ariff, A.B. Stability of bacteriocin-like inhibitory substance (BLIS) produced by Pediococcus acidilactici Kp10 at different extreme conditions. Biomed. Res. Int. 2018, 2018, 5973484. [Google Scholar] [CrossRef]
- Alvarez-Sieiro, P.; Montalbán-López, M.; Mu, D.; Kuipers, O.P. Bacteriocins of lactic acid bacteria: Extending the family. Appl. Microbiol. Biotechnol. 2016, 100, 2939–2951. [Google Scholar] [CrossRef] [PubMed]
- Del Rocío López-Cuellar, M.; Rodríguez-Hernández, A.I.; Norberto, C.-H. LAB bacteriocin applications in the last decade. Biotechnol. Biotechnol. Equip. 2016, 30, 1039–1050. [Google Scholar] [CrossRef]
- Song, A.A.-L.; In, L.L.A.; Lim, E.; Rahim, R.A. A review on Lactococcus lactis: From food to factory. Microb. Cell Factories 2017, 16, 55. [Google Scholar] [CrossRef]
- Silva, C.; Silva, S.P.M.; Ribeiro, S.C. Application of Bacteriocins and Protective Cultures in Dairy Food Preservation. Front. Microbiol. 2018, 9, 594. [Google Scholar] [CrossRef]
- Market Survey Report. 2020. Available online: https://www.marketstudyreport.com/reports/global-food-additives-market-research-report-2020?gclid=Cj0KCQjw3f6HBhDHARIsAD_i3D8TQIqhmx-wMp8-htMLNdP8u8cUxqzqmZ2iGz5vLyCqWrycZx6hULQaAg9gEALw_wcB (accessed on 27 July 2021).
- Jawan, R.; Abbasiliasi, S.; Tan, J.S.; Mustafa, S.; Halim, M.; Ariff, A.B. Influence of Culture Conditions and Medium Compositions on the Production of Bacteriocin-Like Inhibitory Substances by Lactococcus lactis Gh1. Microorganisms 2020, 8, 1454. [Google Scholar] [CrossRef]
- Aguilar, O.; Rito-Palomares, M. Aqueous two-phase systems strategies for the recovery and characterization of biological products from plants. J. Sci. Food Agric. 2010, 90, 1385–1392. [Google Scholar] [CrossRef]
- Muhammad Khan, B.; Cheong, K.-L.; Liu, Y. ATPS: “Aqueous two-phase system” as the “answer to protein separation” for protein-processing food industry. Crit. Rev. Food Sci. Nutr. 2019, 59, 3165–3178. [Google Scholar] [CrossRef]
- Jamaluddin, N.; Stuckey, D.C.; Ariff, A.B.; Wong, F.W.F. Novel approaches to purifying bacteriocin: A review. Crit. Rev. Food Sci. Nutr. 2018, 58, 2453–2465. [Google Scholar] [CrossRef]
- Dutra-Molino, J.V.; Araujo-Feitosa, V.; de Lencastre-Novaes, L.C.; Santos-Ebinuma, V.C.; Lopes, A.M.; Jozala, A.F.; Marques, D.A.V.; Malpiedi, L.P.; Pessoa, A.J. Biomolecules extracted by ATPS: Practical examples. Rev. Mex. Ing. Química 2014, 13, 359–377. [Google Scholar]
- Zhao, L.; Peng, Y.-L.; Gao, J.-M.; Cai, W.-M. Bioprocess intensification: An aqueous two-phase process for the purification of C-phycocyanin from dry Spirulina platensis. Eur. Food Res. Technol. 2013, 238, 451–457. [Google Scholar] [CrossRef]
- Sabo, S.; Lopes, A.; Santos-Ebinuma, V.; Rangel-Yagui, C.; Oliveira, R. Bacteriocin partitioning from a clarified fermentation broth of Lactobacillus plantarum ST16Pa in aqueous two-phase systems with sodium sulfate and choline-based salts as additives. Process. Biochem. 2018, 66, 212–221. [Google Scholar] [CrossRef]
- Banik, R.M.; Santhiagu, A.; Kanari, B.; Sabarinath, C.; Upadhyay, S. Technological aspects of extractive fermentation using aqueous two-phase systems. World J. Microbiol. Biotechnol. 2003, 19, 337–348. [Google Scholar] [CrossRef]
- Li, C.; Ouyang, F.; Bai, J. Extractive cultivation of Lactococcus lactis using a polyethylene glycol/MgSO4 · 7H2O aqueous two-phase system to produce nisin. Biotechnol. Lett. 2000, 22, 843–847. [Google Scholar] [CrossRef]
- Ratanapongleka, K. Recovery of Biological Products in Aqueous Two Phase Systems. Int. J. Chem. Eng. Appl. 2010, 1, 191–198. [Google Scholar] [CrossRef]
- Iqbal, M.; Tao, Y.; Xie, S.; Zhu, Y.; Chen, D.; Wang, X.; Huang, L.; Peng, D.; Sattar, A.; Shabbir, M.A.B.; et al. Aqueous two-phase system (ATPS): An overview and advances in its applications. Proced. Online 2016, 18, 18. [Google Scholar] [CrossRef]
- Raja, S.; Murty, V.R.; Thivaharan, V.; Rajasekar, V.; Ramesh, V. Aqueous Two Phase Systems for the Recovery of Biomolecules—A Review. Sci. Technol. 2011, 1, 7–16. [Google Scholar] [CrossRef]
- Raja, S.; Murty, V.R. Development and Evaluation of Environmentally Benign Aqueous Two Phase Systems for the Recovery of Proteins from Tannery Waste Water. ISRN Chem. Eng. 2012, 2012, 290471. [Google Scholar] [CrossRef]
- Abbasiliasi, S.; Tan, J.S.; Ibrahim, T.A.T.; Ramanan, R.N.; Vakhshiteh, F.; Mustafa, S.; Ling, T.C.; Rahim, R.A.; Ariff, A.B. Isolation of Pediococcus acidilactici Kp10 with ability to secrete bacteriocin-like inhibitory substance from milk products for applications in food industry. BMC Microbiol. 2012, 12, 260. [Google Scholar] [CrossRef]
- Jawan, R.; Abbasiliasi, S.; Mustafa, S.; Kapri, M.R.; Halim, M.; Ariff, A.B. In Vitro Evaluation of Potential Probiotic Strain Lactococcus lactis Gh1 and Its Bacteriocin-Like Inhibitory Substances for Potential Use in the Food Industry. Probiotics Antimicrob. Proteins 2021, 13, 422–440. [Google Scholar] [CrossRef] [PubMed]
- Abbasiliasi, S.; Ramanan, R.N.; Tengku Azmi, T.I.; Shuhaimi, M.; Mohammad, R.; Ariff, A.B. Partial characterization of antimicrobial compound produced by Lactobacillus paracasei LA 07, a strain isolated from Budu. Minerva Biotechnol. 2010, 22, 75–82. [Google Scholar]
- Jawan, R.; Abbasiliasi, S.; Tan, J.; Kapri, M.; Mustafa, S.; Halim, M.; Ariff, A. Evaluation of the Estimation Capability of Response Surface Methodology and Artificial Neural Network for the Optimization of Bacteriocin-Like Inhibitory Substances Production by Lactococcus lactis Gh1. Microorganisms 2021, 9, 579. [Google Scholar] [CrossRef]
- Abbasiliasi, S.; Tan, J.S.; Ibrahim, T.A.T.; Kadkhodaei, S.; Ng, H.S.; Vakhshiteh, F.; Ajdari, Z.; Mustafa, S.; Ling, T.C.; Rahim, R.A.; et al. Primary recovery of a bacteriocin-like inhibitory substance derived from Pediococcus acidilactici Kp10 by an aqueous two-phase system. Food Chem. 2014, 151, 93–100. [Google Scholar] [CrossRef]
- Bernbom, N.; Licht, T.R.; Brogren, C.-H.; Jelle, B.; Johansen, A.H.; Badiola, I.; Vogensen, F.K.; Nørrung, B. Effects of Lactococcus lactis on Composition of Intestinal Microbiota: Role of Nisin. Appl. Environ. Microbiol. 2006, 72, 54–58. [Google Scholar] [CrossRef] [PubMed]
- Aziz, N.F.H.A.; Abbasiliasi, S.; Ng, Z.J.; Abu Zarin, M.; Oslan, S.N.; Tan, J.S.; Bin Ariff, A. Recovery of a Bacteriocin-Like Inhibitory Substance from Lactobacillus bulgaricus FTDC 1211 Using Polyethylene-Glycol Impregnated Amberlite XAD-4 Resins System. Molecules 2020, 25, 5332. [Google Scholar] [CrossRef] [PubMed]
- Sinha, J.; Dey, P.K.; Panda, T. Aqueous two-phase: The system of choice for extractive fermentation. Appl. Microbiol. Biotechnol. 2000, 54, 476–486. [Google Scholar] [CrossRef]
- Chavez-Santoscoy, A.; Benavides, J.; Vermaas, W.; Rito-Palomares, M. Application of Aqueous Two-Phase Systems for the Potential Extractive Fermentation of Cyanobacterial Products. Chem. Eng. Technol. 2010, 33, 177–182. [Google Scholar] [CrossRef]
- Torres-Acosta, M.A.; Mayolo-Deloisa, K.; González-Valdez, J.; Rito-Palomares, M. Aqueous Two-Phase Systems at Large Scale: Challenges and Opportunities. Biotechnol. J. 2019, 14, e1800117. [Google Scholar] [CrossRef]
- Singh, V.; Haque, S.; Niwas, R.; Srivastava, A.; Pasupuleti, M.; Tripathi, C.K.M. Strategies for Fermentation Medium Optimization: An In-Depth Review. Front. Microbiol. 2017, 7, 2087. [Google Scholar] [CrossRef]
- Bayraktar, E. Response surface optimization of the separation of dl-tryptophan using an emulsion liquid membrane. Process. Biochem. 2001, 37, 169–175. [Google Scholar] [CrossRef]
- Gandhi, A.; Shah, N.P. Effect of salt on cell viability and membrane integrity of Lactobacillus acidophilus, Lactobacillus casei and Bifidobacterium longum as observed by flow cytometry. Food Microbiol. 2015, 49, 197–202. [Google Scholar] [CrossRef]
- Medveďová, A.; Šipošová, P.; Mančušková, T.; Valík, Ľ. The Effect of Salt and Temperature on the Growth of Fresco Culture. Fermentation 2018, 5, 2. [Google Scholar] [CrossRef]
- Ng, H.S.; Ooi, C.W.; Mokhtar, M.N.; Show, P.L.; Ariff, A.; Tan, J.S.; Ng, E.-P.; Ling, T.C. Extractive bioconversion of cyclodextrins by Bacillus cereus cyclodextrin glycosyltransferase in aqueous two-phase system. Bioresour. Technol. 2013, 142, 723–726. [Google Scholar] [CrossRef][Green Version]
- Ooi, C.W.; Hii, S.L.; Kamal, S.M.M.; Ariff, A.; Ling, T.C. Extractive fermentation using aqueous two-phase systems for integrated production and purification of extracellular lipase derived from Burkholderia pseudomallei. Process. Biochem. 2011, 46, 68–73. [Google Scholar] [CrossRef]
- Badhwar, P.; Kumar, P.; Dubey, K.K. Extractive Fermentation for Process integration and amplified pullulan production by A. pullulans in Aqueous Two Phase Systems. Sci. Rep. 2019, 9, 32. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Bai, J.; Li, W.; Cai, Z.; Ouyang, F. Optimization of Conditions for Bacteriocin Extraction in PEG/Salt Aqueous Two-Phase Systems Using Statistical Experimental Designs. Biotechnol. Prog. 2001, 17, 366–368. [Google Scholar] [CrossRef]
- Bhavani, A.L.; Nisha, J. Dextran—The polysaccharide with versatile uses. Int. J. Pharma. Biol. Sci. 2010, 1, 569–573. [Google Scholar]
- Kula, M.R.; Korner, K.H.; Hustedt, H. Purification of Enzymes by Liquid-Liquid Extraction. In Reaction Engineering. Advances in Biochemical Engineering; Fechter, A., Ed.; Springer: Berlin/Heidelberg, Germany, 1982; pp. 73–118. [Google Scholar]
- Pandey, S.; Banik, R. Extractive fermentation for enhanced production of alkaline phosphatase from Bacillus licheniformis MTCC 1483 using aqueous two-phase systems. Bioresour. Technol. 2011, 102, 4226–4231. [Google Scholar] [CrossRef]
- Show, P.L.; Tan, C.P.; Anuar, M.S.; Ariff, A.; Yusof, Y.A.; Chen, S.K.; Ling, T.C. Extractive fermentation for improved production and recovery of lipase derived from Burkholderia cepacia using a thermoseparating polymer in aqueous two-phase systems. Bioresour. Technol. 2012, 116, 226–233. [Google Scholar] [CrossRef] [PubMed]
- Saravanan, S.; Rao, J.R.; Nair, B.U.; Ramasami, T. Aqueous two-phase poly(ethylene glycol)–poly(acrylic acid) system for protein partitioning: Influence of molecular weight, pH and temperature. Process. Biochem. 2008, 43, 905–911. [Google Scholar] [CrossRef]
- Wang, S.J.; Zhong, J.J. Bioreactor engineering. In Bioprocessing for Value-Added Products from Renewable Resources; Yang, S.T., Ed.; Elsevier: Amsterdam, The Netherlands, 2006; pp. 131–161. [Google Scholar]
- Marques, D.D.A.V.; Santos-Ebinuma, V.C.; Pessoa, A.; Porto, A.L.F.; Torres, B.R.; Converti, A. Effect of aeration and agitation on extractive fermentation of clavulanic acid by using aqueous two-phase system. Biotechnol. Prog. 2016, 32, 1444–1452. [Google Scholar] [CrossRef]
- Sankaran, R.; Manickam, S.; Yap, Y.J.; Ling, T.C.; Chang, J.-S.; Show, P.L. Extraction of proteins from microalgae using integrated method of sugaring-out assisted liquid biphasic flotation (LBF) and ultrasound. Ultrason. Sonochem. 2018, 48, 231–239. [Google Scholar] [CrossRef] [PubMed]
- Khoo, K.S.; Chew, K.W.; Ooi, C.W.; Ong, H.C.; Ling, T.C.; Show, P.L. Extraction of natural astaxanthin from Haematococcus pluvialis using liquid biphasic flotation system. Bioresour. Technol. 2019, 290, 121794. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable (%, w/v) | Symbol | Coded Values | ||||
|---|---|---|---|---|---|---|
| −α | −1 | 0 | +1 | +α | ||
| PEG | X1 | 7.0 | 8.0 | 10.0 | 12.0 | 13.0 |
| Dextran T500 | X2 | 5.0 | 6.0 | 8.0 | 10.0 | 11.0 |
| Type of PEG | Concentration (%, w/w) | BLIS Activity (AU/mL) | Protein Concentration (μg/mL) | Cell Concentration (g/L) |
|---|---|---|---|---|
| Control (BHI medium) | 552.00 ± 34.34 cd | 83.65 ± 0.122 a | 0.33 ± 0.002 a | |
| PEG2000 | 5 | 577.15 ± 17.43 abcd | 32.00 ± 0.032 cde | 0.26 ± 0.003 e |
| 10 | 580.40 ± 8.15 abcd | 10.59 ± 0.013 hi | 0.27 ± 0.003 d | |
| 15 | 580.39 ± 1.16 abcd | 6.77 ± 0.020 i | 0.22 ± 0.008 h | |
| 20 | 553.56 ± 27.50 cd | 6.91 ± 0.018 i | 0.19 ± 0.002 j | |
| PEG4000 | 5 | 556.73 ± 16.07 cd | 34.07 ± 0.016 cd | 0.30 ± 0.002 c |
| 10 | 538.76 ± 64.70 d | 23.58 ± 0.026 efg | 0.23 ± 0.002 g | |
| 15 | 568.93 ± 12.72 bcd | 17.15 ± 0.030 gh | 0.21 ± 0.001 h | |
| 20 | 580.42 ± 15.13 abcd | 19.44 ± 0.024 fgh | 0.18 ± 0.001 k | |
| PEG6000 | 5 | 590.60 ± 45.66 abcd | 39.86 ± 0.082 c | 0.25 ± 0.008 f |
| 10 | 585.50 ± 33.85 abcd | 36.21 ± 0.056 cd | 0.22 ± 0.001 h | |
| 15 | 611.31 ± 37.91 abcd | 35.89 ± 0.043 cd | 0.20 ± 0.001 i | |
| 20 | 622.92 ± 16.69 abc | 27.40 ± 0.083 def | 0.18 ± 0.001 k | |
| PEG8000 | 5 | 638.12 ± 7.21 ab | 55.96 ± 0.036 b | 0.31 ± 0.002 b |
| 10 | 645.97 ± 37.43 a | 49.36 ± 0.030 b | 0.27 ± 0.003 d | |
| 15 | 626.60 ± 50.17 abc | 35.93 ± 0.053 cd | 0.23 ± 0.004 g | |
| 20 | 622.04 ± 3.58 abc | 26.35 ± 0.031 defg | 0.20 ± 0.001 i | |
| Effect of type | F = 8.94 (S) | F = 90.14 (S) | F = 307.08 (S) | |
| Effect of concentration | F = 0.14 (NS) | F = 47.42 (S) | F = 1509.89 (S) | |
| Interaction | F = 0.61 (NS) | F = 4.46 (S) | F = 74.78 (S) | |
| Type of Bottom Phase Components | Concentration (%, w/w) | BLIS Activity (AU/mL) | Protein Concentration (μg/mL) | Cell Concentration (g/L) |
|---|---|---|---|---|
| Control (BHI medium) | 624.57 ± 4.77 d | 73.68± 0.010 acbd | 0.34 ± 0.005 b | |
| Ammonium sulphate | 5 | 618.67 ± 3.57 e | 73.93 ± 0.058 abcd | 0.37 ± 0.002 a |
| 10 | 602.84 ± 23.58 de | 55.58 ± 0.106 cdef | 0.17 ± 0.003 g | |
| 15 | 711.80 ± 6.25 bc | 50.42 ± 0.028 def | 0.17 ± 0.001 g | |
| 20 | 216.05 ± 7.08 g | 40.24 ± 0.029 f | 0.002 ± 0.001 k | |
| Sodium citrate | 5 | 558.31 ± 2.30 e | 73.86 ± 0.140 acbd | 0.22 ± 0.003 e |
| 10 | 342.05 ± 15.84 f | 54.66 ± 0.070 cdef | 0.01 ± 0.0002 i | |
| 15 | 385.50 ± 17.41 f | 46.98 ± 0.066 ef | 0.01 ± 0.0003 j | |
| 20 | 0.00 ± 0.00 h | 74.38 ± 0.161 abcd | 0.00 ± 0.00 k | |
| Sodium phosphate | 5 | 629.66 ± 14.36 d | 77.72 ± 0.136 abc | 0.27 ± 0.004 c |
| 10 | 729.56 ± 8.82 d | 90.70 ± 0.188 a | 0.19 ± 0.001 f | |
| 15 | 805.81 ± 6.54 a | 76.70 ± 0.165 abc | 0.17 ± 0.001 g | |
| 20 | 0.00 ± 0.00 h | 90.66 ± 0.244 a | 0.00 ± 0.0003 k | |
| Dextran T500 | 5 | 565.94 ± 45.01 e | 89.61 ± 0.135 ab | 0.22 ± 0.001 d |
| 10 | 694.40 ± 38.39 c | 69.75 ± 0.052 abcde | 0.22 ± 0.001 d | |
| 15 | 730.55 ± 30.27 bc | 66.14 ± 0.076 bcde | 0.17 ± 0.001 g | |
| 20 | 749.42 ± 36.91 b | 67.54 ± 0.015 abcde | 0.13 ± 0.0002 h | |
| Effect of type | F = 392.83 (S) | F = 11.58 (S) | F = 14868.54 (S) | |
| Effect of concentration | F = 202.19 (S) | F = 5.61 (S) | F = 36171.86 (S) | |
| Interaction effect | F = 289.63 (S) | F = 1.59 (NS) | F = 6241.57 (S) | |
| PEG (X1) (%, w/w) | Dextran (X2) (%, w/w) | Partition Coefficient (K) | ||
|---|---|---|---|---|
| Experimental | Predicted by RSM (% Difference) * | |||
| 10 | 8 | 1.37 | 1.39 (−1.46) | |
| 10 | 5 | 1.27 | 1.25 (1.57) | |
| 10 | 8 | 1.47 | 1.39 (5.44) | |
| 10 | 8 | 1.35 | 1.39 (−2.96) | |
| 10 | 11 | 1.22 | 1.22 (0.82) | |
| 2000 | 10 | 8 | 1.40 | 1.39 (0.71) |
| 13 | 8 | 1.05 | 1.04 (0.95) | |
| 12 | 10 | 1.05 | 1.07 (−1.90) | |
| 10 | 8 | 1.36 | 1.39 (−2.21) | |
| 12 | 6 | 1.19 | 1.19 (0.000 | |
| 8 | 6 | 1.23 | 1.21 (1.63) | |
| 7 | 8 | 1.15 | 1.16 (−0.87) | |
| 8 | 10 | 1.22 | 1.23 (−0.82) | |
| 10 | 8 | 0.42 | 0.46 (−9.44) | |
| 10 | 5 | 0.55 | 0.57 (−2.95) | |
| 10 | 8 | 0.46 | 0.46 (0.12) | |
| 10 | 8 | 0.46 | 0.46 (0.44) | |
| 10 | 11 | 0.42 | 0.38 (9.22) | |
| 4000 | 10 | 8 | 0.52 | 0.46 (11.97) |
| 13 | 8 | 0.56 | 0.56 (0.55) | |
| 12 | 10 | 0.44 | 0.47 (−5.97) | |
| 10 | 8 | 0.45 | 0.46 (−2.24) | |
| 12 | 6 | 0.55 | 0.53 (3.13) | |
| 8 | 6 | 0.73 | 0.73 (0.36) | |
| 7 | 8 | 0.77 | 0.74 (3.72) | |
| 8 | 10 | 0.48 | 0.52 (−9.30) | |
| 10 | 8 | 0.39 | 0.36 (8.37) | |
| 10 | 5 | 0.44 | 0.46 (−4.11) | |
| 10 | 8 | 0.36 | 0.36 (−0.64) | |
| 10 | 8 | 0.34 | 0.36 (−7.43) | |
| 10 | 11 | 0.41 | 0.40 (1.38) | |
| 6000 | 10 | 8 | 0.37 | 0.36 (1.53) |
| 13 | 8 | 0.34 | 0.34 (−1.05) | |
| 12 | 10 | 0.37 | 0.38 (−1.36) | |
| 10 | 8 | 0.36 | 0.36 (1.22) | |
| 12 | 6 | 0.39 | 0.38 (2.47) | |
| 8 | 6 | 0.49 | 0.48 (2.26) | |
| 7 | 8 | 0.43 | 0.43 (−0.88) | |
| 8 | 10 | 0.39 | 0.39 (−1.23) | |
| Dependent Variables | Equation |
|---|---|
| PEG2000: dextran | +1.39 − 0.045X1 − 0.026X2 − 0.15X12 − 0.071X22 − 0.033X1X2 |
| PEG4000: dextran | +0.46 − 0.064X1 − 0.069X2 + 0.095X22 + 4.969 x 103X22 + 0.038X1X2 |
| PEG6000: dextran | +0.36 − 0.030X1 − 0.021X2 + 0.011X12 + 0.032X22 + 0.023X1X2 |
| Source | SS | DF | MS | F-Value | Prob (p) > F |
|---|---|---|---|---|---|
| PEG 2000 | |||||
| Model | 0.19 | 5 | 0.038 | 26.04 | 0.0002 ** |
| Residual | 0.010 | 7 | 1.474 × 103 | ||
| Lack of fit | 1.023 × 103 | 3 | 3.409 ×104 | 0.15 | 0.9266 * |
| Pure error | 9.294 × 103 | 4 | 2.323 × 103 | ||
| Total | 0.20 | 12 | |||
| R2 | 0.9490 | ||||
| Adj-R2 | 0.9126 | ||||
| Pred-R2 | 0.8922 | ||||
| CV | 3.05 | ||||
| PEG 4000 | |||||
| Model | 0.14 | 5 | 0.028 | 17.43 | 0.0008 ** |
| Residual | 0.011 | 7 | 1.596 × 103 | ||
| Lack of fit | 5.630 × 103 | 3 | 1.877 × 103 | 1.35 | 0.3762 * |
| Pure error | 5.544 × 103 | 4 | 1.386 × 103 | ||
| Total | 0.15 | 12 | |||
| R2 | 0.9257 | ||||
| Adj-R2 | 0.8726 | ||||
| Pred-R2 | 0.6760 | ||||
| CV | 7.62 | ||||
| PEG 6000 | |||||
| Model | 0.020 | 5 | 4.088 × 103 | 11.66 | 0.0027 ** |
| Residual | 2.454 × 103 | 7 | 3.506 × 104 | ||
| Lack of fit | 7.465 × 104 | 3 | 2.488 × 104 | 0.58 | 0.6572 * |
| Pure error | 1.708 × 103 | 4 | 4.270 × 104 | ||
| Total | 0.023 | 12 | |||
| R2 | 0.8928 | ||||
| Adj-R2 | 0.8162 | ||||
| Pred-R2 | 0.6516 | ||||
| CV | 4.80 |
| Parameters | Predicted by RSM | Experimental | % of Difference |
|---|---|---|---|
| Purification factor (PF) | 1.78 | 2.15 | −0.21 |
| Partition coefficient (K) | 1.39 | 1.07 | 0.23 |
| Recovery/yield (%) | 73.97 | 76.28 | −0.03 |
| Cell concentration (g/L) | 0.37 | 0.22 | 0.40 |
| Orbital Speed | Specific Activity (SA) | Partition Coefficient (K) | Purification Factor(PF) | Selectivity (S) | Yield (%) | Cell Concentration (g/L) |
|---|---|---|---|---|---|---|
| 150 | 16,741.63 ± 1079.75 a | 0.87 ± 0.017 ab | 1.76 ± 0.11 a | 2.55 ± 0.26 a | 75.03 ± 0.37 ab | 0.15 ± 0.02 b |
| 200 | 16,899.88 ± 951.29 a | 0.94 ± 0.040 a | 2.02 ± 0.11 a | 1.76 ± 0.29 ab | 76.46 ± 0.77 a | 0.16 ± 0.01 b |
| 250 | 19,530.09 ± 2173.03 a | 0.80 ± 0.032 b | 1.72 ± 0.19 a | 1.38 ± 0.33 b | 73.34 ± 0.78 b | 0.22 ± 0.01 a |
| pH | Specific Activity (SA) | Partition Coefficient (K) | Purification Factor (PF) | Selectivity (S) | Yield (%) | Cell Concentration (g/L) |
|---|---|---|---|---|---|---|
| * Control | 16707.71 ± 1352.99 bc | 0.83 ± 0.10 a | 2.38 ± 0.19b c | 1.08 ± 0.45 ab | 74.05 ± 2.24 a | 0.13 ± 0.02 c |
| 5 | 21820.84 ± 1900.45 a | 0.79 ± 0.07 a | 3.11 ± 0.27 a | 0.38 ± 0.26 b | 72.97 ± 1.72 a | 0.11 ± 0.01 c |
| 6 | 19775.51 ± 1306.68 ab | 0.81 ± 0.02 a | 2.82 ± 0.19 ab | 0.96 ± 0.16 ab | 73.61 ± 0.56 a | 0.13 ± 0.00 c |
| 7 | 20442.98 ± 2606.24 ab | 1.00 ± 0.16 a | 2.92 ± 0.37 ab | 2.06 ± 1.34 a | 77.24 ± 2.81 a | 0.12 ± 0.02 c |
| 8 | 15535.43 ± 1087.32 c | 0.91 ± 0.15 a | 2.22 ± 0.16 c | 0.93 ± 0.28 ab | 75.55 ± 3.05 a | 0.24 ± 0.00 b |
| 9 | 12697.92 ± 1215.92 c | 0.85 ± 0.01 a | 1.81 ± 0.17 c | 1.20 ± 0.38 ab | 74.62 ± 0.13 a | 0.29 ± 0.03 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jawan, R.; Abbasiliasi, S.; Tan, J.S.; Halim, M.; Mustafa, S.; Lee, B.H.; Kwa, J.S.; Ariff, A.B. Extractive Fermentation for Recovery of Bacteriocin-Like Inhibitory Substances Derived from Lactococcus lactis Gh1 Using PEG2000/Dextran T500 Aqueous Two-Phase System. Fermentation 2021, 7, 257. https://doi.org/10.3390/fermentation7040257
Jawan R, Abbasiliasi S, Tan JS, Halim M, Mustafa S, Lee BH, Kwa JS, Ariff AB. Extractive Fermentation for Recovery of Bacteriocin-Like Inhibitory Substances Derived from Lactococcus lactis Gh1 Using PEG2000/Dextran T500 Aqueous Two-Phase System. Fermentation. 2021; 7(4):257. https://doi.org/10.3390/fermentation7040257
Chicago/Turabian StyleJawan, Roslina, Sahar Abbasiliasi, Joo Shun Tan, Murni Halim, Shuhaimi Mustafa, Bin Hao Lee, Jia Sim Kwa, and Arbakariya B. Ariff. 2021. "Extractive Fermentation for Recovery of Bacteriocin-Like Inhibitory Substances Derived from Lactococcus lactis Gh1 Using PEG2000/Dextran T500 Aqueous Two-Phase System" Fermentation 7, no. 4: 257. https://doi.org/10.3390/fermentation7040257
APA StyleJawan, R., Abbasiliasi, S., Tan, J. S., Halim, M., Mustafa, S., Lee, B. H., Kwa, J. S., & Ariff, A. B. (2021). Extractive Fermentation for Recovery of Bacteriocin-Like Inhibitory Substances Derived from Lactococcus lactis Gh1 Using PEG2000/Dextran T500 Aqueous Two-Phase System. Fermentation, 7(4), 257. https://doi.org/10.3390/fermentation7040257