
Effect of Ellagic Acid on Fermentation Quality and Bacterial Community of Stylo Silage


Abstract
:1. Introduction
2. Materials and Methods
2.1. Raw Material and Silage Preparation
2.2. Microbial and Chemical Composition
2.3. Bacteria Community Analysis
2.4. Antioxidant Activity
2.5. Statistical Analysis
3. Results and Discussion
3.1. Characteristics of Fresh Stylo
3.2. Fermentation Quality of Stylo Silage
3.3. Protein Content of Stylo Silage
3.4. Antioxidant Activity
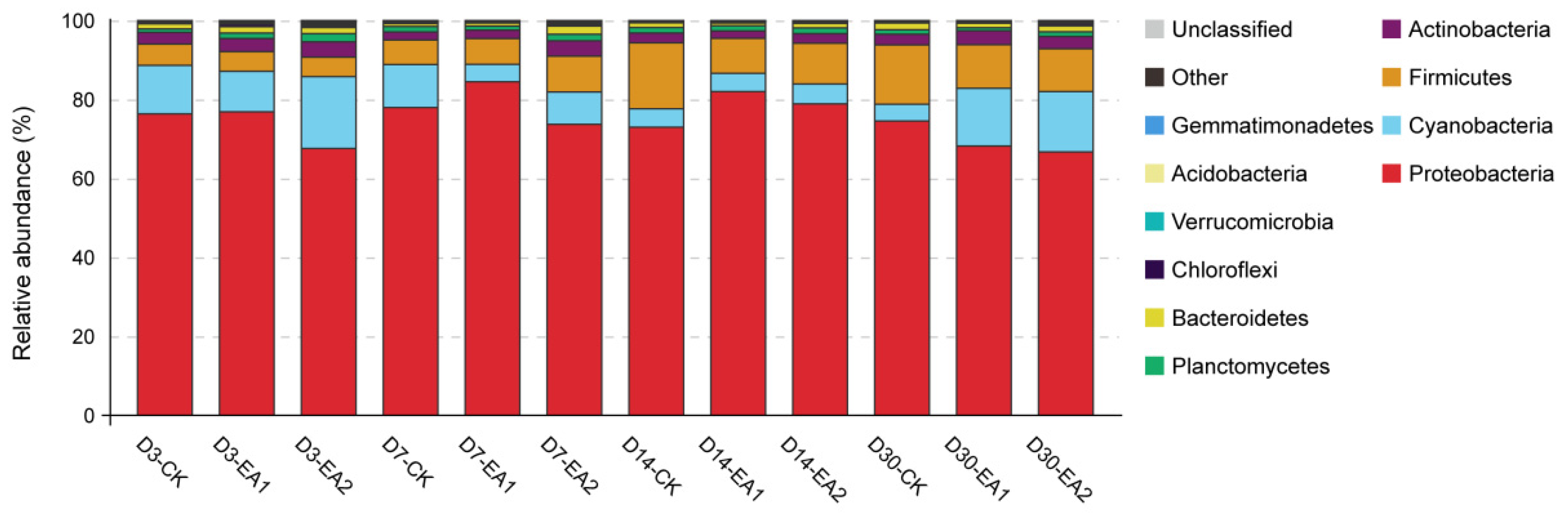
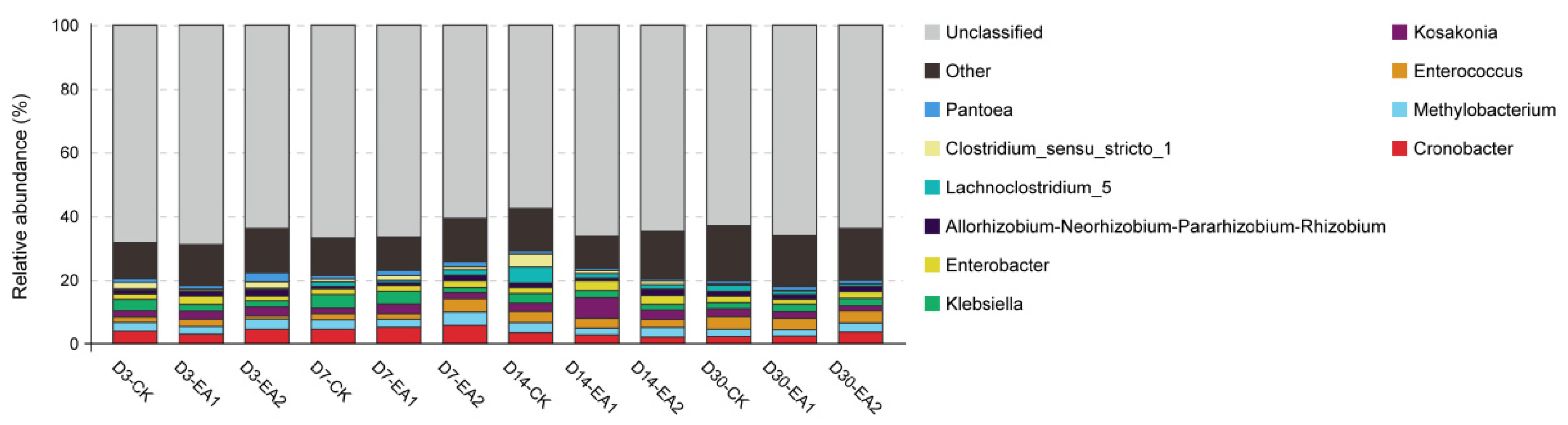
3.5. Bacteria Community
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- He, L.; Wang, C.; Xing, Y.; Zhou, W.; Pian, R.; Chen, X.; Zhang, Q. Ensiling characteristics, proteolysis and bacterial community of high-moisture corn stalk and stylo silage prepared with Bauhinia variegate flower. Bioresour. Technol. 2020, 296, 122336. [Google Scholar] [CrossRef] [PubMed]
- Jessie, G.; Vern, S.B.; Karen, A.B. Corn Forage Yield and Quality for Silage in Short Growing Season Areas of the Canadian Prairies. Agronomy 2018, 8, 164. [Google Scholar]
- Wang, Y.; Wang, C.; Zhou, W.; Yang, F.Y.; Chen, X.Y.; Zhang, Q. Effects of Wilting and Lactobacillus plantarum Addition on the Fermentation Quality and Microbial Community of Moringa oleifera Leaf Silage. Front. Microbiol. 2018, 9, 1817. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cai, Y.; Benno, Y.; Ogawa, M.; Ohmomo, S.; Kumai, S.; Nakase, T. Influence of lactobacillus spp. from An inoculant and of weissella and leuconostoc spp. from forage crops on silage fermentation. Appl. Environ. Microbiol. 1998, 64, 2982–2987. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, C.; Pian, R.; Chen, X.; Lv, H.; Zhou, W.; Zhang, Q. Beneficial Effects of Tannic Acid on the Quality of Bacterial Communities Present in High-Moisture Mulberry Leaf and Stylo Silage. Front. Microbiol. 2020, 11, 586412. [Google Scholar] [CrossRef] [PubMed]
- Barroso, A.L.P.; Pittarello, M.; Neto, A.C.; Busato, J.G.; Santos, J.L.A.; Dobbss, L.B. Humic Acids from Vermicompost and Eucalyptus urograndis Essential Oil: Biological Activity on Stylosanthes guianensis (Leguminosae) Seedlings. J. Essent. Oil-Bear. Plants (Dehra Dun) 2019, 22, 1322–1332. [Google Scholar] [CrossRef]
- Zhang, Q.; Zhao, M.; Wang, X.; Yu, Z.; Na, R. Ensiling alfalfa with whole crop corn improves the silage quality and in vitro digestibility of the silage mixtures. Grassl. Sci. 2017, 63, 211–217. [Google Scholar] [CrossRef]
- Denek, N.; Avci, C.A. The effect of molasses-based pre-fermented juice on the fermentation quality of first-cut lucerne silage. Grass Forage Sci. 2011, 66, 243–250. [Google Scholar] [CrossRef]
- Silva, V.P.; Pereira, O.G.; Leandro, E.S.; Da, S.T.; Ribeiro, K.G.; Mantovani, H.C.; Santos, S.A. Effects of lactic acid bacteria with bacteriocinogenic potential on the fermentation profile and chemical composition of alfalfa silage in tropical conditions. J. Dairy Sci. 2016, 99, 1895–1902. [Google Scholar] [CrossRef]
- Landete, J.M. Ellagitannins, ellagic acid and their derived metabolites: A review about source, metabolism, functions and health. Food Res. Int. 2011, 44, 1150–1160. [Google Scholar] [CrossRef]
- Umesalma, S.; Sudhandiran, G. Differential Inhibitory Effects of the Polyphenol Ellagic Acid on Inflammatory Mediators NF-κB, iNOS, COX-2, TNF-α, and IL-6 in 1,2-Dimethylhydrazine-Induced Rat Colon Carcinogenesis. Basic Clin. Pharm. 2010, 107, 650–655. [Google Scholar] [CrossRef] [PubMed]
- Howell, A.B.; D’Souza, D.H. The pomegranate: Effects on bacteria and viruses that influence human health. Evid.-Based Complement. Altern. Med. 2013, 2013, 606212. [Google Scholar] [CrossRef] [Green Version]
- Gommersall, L.M.; Albrecht, M.; Jiang, W.; Lansky, E.P.; Kumi-Diaka, J.; Campbell, M.J. Pomegranate extracts potently suppress proliferation, xenograft growth and invasion of human prostate cancer cells. Eur. Urol. Suppl. Off. J. Eur. Assoc. Urol. 2003, 2, 138. [Google Scholar] [CrossRef]
- Reddy, M.K.; Gupta, S.K.; Jacob, M.R.; Khan, S.I.; Ferreira, D. Antioxidant, antimalarial and antimicrobial activities of tannin-rich fractions, ellagitannins and phenolic acids from Punica granatum L. Planta Med. 2007, 73, 461–467. [Google Scholar] [CrossRef]
- Yamaguchi, M.U.; Garcia, F.P.; Cortez, D.A.; Ueda-Nakamura, T.; Filho, B.P.; Nakamura, C.V. Antifungal effects of Ellagitannin isolated from leaves of Ocotea odorifera (Lauraceae). Antonie Leeuwenhoek 2011, 99, 507–514. [Google Scholar] [CrossRef] [PubMed]
- Gil, M.I.; Tomas-Barberan, F.A.; Hess-Pierce, B.; Holcroft, D.M.; Kader, A.A. Antioxidant activity of pomegranate juice and its relationship with phenolic composition and processing. J. Agric. Food Chem. 2000, 48, 4581–4589. [Google Scholar] [CrossRef]
- Seeram, N.; Lee, R.; Hardy, M.; Heber, D. Rapid large scale purification of ellagitannins from pomegranate husk, a by-product of the commercial juice industry. Sep. Purif. Technol. 2004, 41, 49–55. [Google Scholar] [CrossRef] [Green Version]
- Kotsampasi, B.; Christodoulou, V.; Zotos, A.; Liakopoulou-Kyriakides, M.; Goulas, P.; Petrotos, K.; Natas, P.; Bampidis, V.A. Effects of dietary pomegranate byproduct silage supplementation on performance, carcass characteristics and meat quality of growing lambs. Anim. Feed. Sci. Technol. 2014, 197, 92–102. [Google Scholar] [CrossRef]
- Kotsampasi, B.; Christodoulou, C.; Tsiplakou, E.; Mavrommatis, A.; Mitsiopoulou, C.; Karaiskou, C.; Dotas, V.; Robinson, P.H.; Bampidis, V.A.; Christodoulou, V.; et al. Effects of dietary pomegranate pulp silage supplementation on milk yield and composition, milk fatty acid profile and blood plasma antioxidant status of lactating dairy cows. Anim. Feed. Sci. Technol. 2017, 234, 228–236. [Google Scholar] [CrossRef]
- Pham-Huy, L.A.; He, H.; Pham-Huy, C. Free radicals, antioxidants in disease and health. Int. J. Biomed. Sci. IJBS 2008, 4, 89. [Google Scholar] [PubMed]
- Robinson, P.H.; Wiseman, J.; Udén, P.; Mateos, G. Some experimental design and statistical criteria for analysis of studies in manuscripts submitted for consideration for publication. Anim. Feed. Sci. Technol. 2006, 129, 1–11. [Google Scholar] [CrossRef]
- He, L.; Chen, N.; Lv, H.; Wang, C.; Zhou, W.; Chen, X.; Zhang, Q. Gallic acid influencing fermentation quality, nitrogen distribution and bacterial community of high-moisture mulberry leaves and stylo silage. Bioresour. Technol. 2020, 295, 122255. [Google Scholar] [CrossRef]
- Wang, C.; He, L.; Xing, Y.; Zhou, W.; Yang, F.; Chen, X.; Zhang, Q. Effects of mixing Neolamarckia cadamba leaves on fermentation quality, microbial community of high moisture alfalfa and stylo silage. Microb. Biotechnol. 2019, 12, 869–878. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; He, L.; Xing, Y.; Zhou, W.; Pian, R.; Yang, F.; Chen, X.; Zhang, Q. Bacterial diversity and fermentation quality of Moringa oleifera leaves silage prepared with lactic acid bacteria inoculants and stored at different temperatures. Bioresour. Technol. 2019, 284, 349–358. [Google Scholar] [CrossRef]
- Xie, X.; Yang, C.; Guan, L.L.; Wang, J.; Xue, M.; Liu, J.X. Persistence of Cellulolytic Bacteria Fibrobacter and Treponema After Short-Term Corn Stover-Based Dietary Intervention Reveals the Potential to Improve Rumen Fibrolytic Function. Front. Microbiol. 2018, 9, 1363. [Google Scholar] [CrossRef]
- Abdennacer, B.; Karim, M.; Yassine, M.; Nesrine, R.; Mouna, D.; Mohamed, B. Determination of phytochemicals and antioxidant activity of methanol extracts obtained from the fruit and leaves of Tunisian Lycium intricatum Boiss. Food Chem. 2015, 174, 577–584. [Google Scholar] [CrossRef]
- Li, X.; Jiang, Q.; Wang, T.; Liu, J.; Chen, D. Comparison of the Antioxidant Effects of Quercitrin and Isoquercitrin: Understanding the Role of the 6″-OH Group. Molecules 2016, 21, 1246. [Google Scholar] [CrossRef]
- Van Soest, J.P.; Mertens, R.D.; Deinum, B. Preharvest factors influencing quality of conserved forage. J. Anim. Sci. 1978, 47, 712–720. [Google Scholar] [CrossRef]
- McDonald, P. The Biochemistry of Silage; J. Wiley: Chichester, UK, 1981; p. 226. [Google Scholar]
- Lv, H.; Pian, R.; Xing, Y.; Zhou, W.; Yang, F.; Chen, X.; Zhang, Q. Effects of citric acid on fermentation characteristics and bacterial diversity of Amomum villosum silage. Bioresour. Technol. 2020, 307, 123290. [Google Scholar] [CrossRef]
- Bialonska, D.; Ramnani, P.; Kasimsetty, S.G.; Muntha, K.R.; Gibson, G.R.; Ferreira, D. The influence of pomegranate by-product and punicalagins on selected groups of human intestinal microbiota. Int. J. Food Microbiol. 2010, 140, 175–182. [Google Scholar] [CrossRef]
- Hayrapetyan, H.; Hazeleger, W.C.; Beumer, R.R. Inhibition of Listeria monocytogenes by pomegranate (Punica granatum) peel extract in meat paté at different temperatures. Food Control. 2012, 23, 66–72. [Google Scholar] [CrossRef]
- Wang, C.; He, L.; Xing, Y.; Zhou, W.; Yang, F.; Chen, X.; Zhang, Q. Fermentation quality and microbial community of alfalfa and stylo silage mixed with Moringa oleifera leaves. Bioresour. Technol. 2019, 284, 240–247. [Google Scholar] [CrossRef] [PubMed]
- Peng, K.; Jin, L.; Niu, Y.D.; Huang, Q.; McAllister, T.A.; Yang, H.E.; Denise, H.; Xu, Z.; Acharya, S.; Wang, S.; et al. Condensed Tannins Affect Bacterial and Fungal Microbiomes and Mycotoxin Production during Ensiling and upon Aerobic Exposure. Appl. Environ. Microb. 2018, 84, e02274-17. [Google Scholar] [CrossRef] [Green Version]
- Grabber, J.H.; Coblentz, W.K. Polyphenol, Conditioning, and Conservation Effects on Protein Fractions and Degradability in Forage Legumes. Crop. Sci. 2009, 49, 1511–1522. [Google Scholar] [CrossRef]
- He, L.; Lv, H.; Wang, C.; Zhou, W.; Pian, R.; Zhang, Q.; Chen, X. Dynamics of fermentation quality, physiochemical property and enzymatic hydrolysis of high-moisture corn stover ensiled with sulfuric acid or sodium hydroxide. Bioresour. Technol. 2020, 298, 122510. [Google Scholar] [CrossRef]
- Li, X.; Tian, J.; Zhang, Q.; Jiang, Y.; Hou, J.; Wu, Z.; Yu, Z. Effects of applyingLactobacillus plantarum and Chinese gallnut tannin on the dynamics of protein degradation and proteases activity in alfalfa silage. Grass Forage Sci. 2018, 73, 648–659. [Google Scholar] [CrossRef]
- Cohen-Zinder, M.; Weinberg, Z.; Leibovich, H.; Chen, Y.; Rosen, M.; Sagi, G.; Orlov, A.; Agmon, R.; Yishay, M.; Miron, J.; et al. Ensiled Moringa oleifera: An antioxidant-rich feed that improves dairy cattle performance. J. Agric. Sci. 2017, 155, 1174–1186. [Google Scholar] [CrossRef]
- Papoutsi, Z.; Kassi, E.; Tsiapara, A.; Fokialakis, N.; Chrousos, G.P.; Moutsatsou, P. Evaluation of Estrogenic/Antiestrogenic Activity of Ellagic Acid via the Estrogen Receptor Subtypes ERα and ERβ. J. Agric. Food Chem. 2005, 53, 7715–7720. [Google Scholar] [CrossRef]
- Liu, B.; Huan, H.; Gu, H.; Xu, N.; Shen, Q.; Ding, C. Dynamics of a microbial community during ensiling and upon aerobic exposure in lactic acid bacteria inoculation-treated and untreated barley silages. Bioresour. Technol. 2019, 273, 212–219. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Yuan, X.; Li, J.; Dong, Z.; Shao, T. Dynamics of microbial community and fermentation quality during ensiling of sterile and nonsterile alfalfa with or without Lactobacillus plantarum inoculant. Bioresour. Technol. 2019, 275, 280–287. [Google Scholar] [CrossRef] [PubMed]
- Romero, J.J.; Joo, Y.; Park, J.; Tiezzi, F.; Gutierrez-Rodriguez, E.; Castillo, M.S. Bacterial and fungal communities, fermentation, and aerobic stability of conventional hybrids and brown midrib hybrids ensiled at low moisture with or without a homo- and heterofermentative inoculant. J. Dairy Sci. 2018, 101, 3057–3076. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doronina, N.V.; Trotsenko, Y.A.; Kuznetsov, B.B.; Tourova, T.P.; Salkinoja-Salonen, M.S. Methylobacterium suomiense sp. nov. and Methylobacterium lusitanum sp. nov., aerobic, pink-pigmented, facultatively methylotrophic bacteria. Int. J. Syst. Evol. Microbiol. 2002, 52, 773–776. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ogunade, I.M.; Jiang, Y.; Pech, C.A.; Kim, D.H.; Oliveira, A.S.; Vyas, D.; Weinberg, Z.G.; Jeong, K.C.; Adesogan, A.T. Bacterial diversity and composition of alfalfa silage as analyzed by Illumina MiSeq sequencing: Effects of Escherichia coli O157:H7 and silage additives. J. Dairy Sci. 2018, 101, 2048–2059. [Google Scholar] [CrossRef] [PubMed]
- Ni, K.; Zhao, J.; Zhu, B.; Su, R.; Pan, Y.; Ma, J.; Zhou, G.; Tao, Y.; Liu, X.; Zhong, J. Assessing the fermentation quality and microbial community of the mixed silage of forage soybean with crop corn or sorghum. Bioresour. Technol. 2018, 265, 563–567. [Google Scholar] [CrossRef] [PubMed]
- Haixia, T.; Jiao, W.; Jiayu, L.; Yujie, W.; Megharaj, M.; Wenxiang, H. Six New Families of Aerobic Arsenate Reducing Bacteria: Leclercia, Raoultella, Kosakonia, Lelliottia, Yokenella, and Kluyvera. Geomicrobiol. J. 2019, 36, 339–347. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Items | Stylo |
|---|---|
| Dry matter (DM, %) | 34.3 ± 0.67 |
| Water-soluble carbohydrates (WSC, %DM) | 4.94 ± 0.67 |
| Crud protein (CP, %DM) | 13.3 ± 0.99 |
| True protein (% TN) | 58.0 ± 3.75 |
| Non-protein nitrogen (% TN) | 42.00 ± 3.75 |
| Neutral detergent fiber (NDF, %DM) | 60.2 ± 0.54 |
| Acid detergent fiber (ADF, %DM) | 47.0 ± 0.09 |
| Lactic acid bacteria (LAB, log10·cfu/g FM) | 4.93 ± 0.19 |
| Yeasts (log10·cfu/g FM) | 3.83 ± 0.35 |
| Coliform bacteria (log10·cfu/g FM) | 5.68 ± 0.18 |
| Molds (log10·cfu/g FM) | 3.71 ± 0.30 1 |
| Item | Treatments | Ensiling Days | Means | SEM | p-Value | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| 3 | 7 | 14 | 30 | D | T | D*T | ||||
| Dry matter (DM, %) | CK | 34.0 B | 34.9 C | 34.5 B | 35.1 B | 34.6 C | 0.09 | * | ** | NS |
| EA1 | 35.7 A | 35.9 B | 35.5 AB | 35.7 AB | 35.7 B | |||||
| EA2 | 35.5 A | 36.7 A | 36.4 A | 36.7 A | 36.7 A | |||||
| Dry matter recovery (%) | CK | 98.7 B | 101.1 C | 99.8 C | 100.3 B | 100.0 C | 0.22 | NS | ** | NS |
| EA1 | 103.8 Ba | 103.8 Ba | 102.4 Bb | 101.8 ABb | 103.0 B | |||||
| EA2 | 104.3 A | 106.1 A | 106.4 A | 105.1 A | 105.5 A | |||||
| pH | CK | 5.65 b | 5.94 Aa | 5.48 Ac | 5.29 Ad | 5.62 A | 0.01 | ** | ** | NS |
| EA1 | 5.53 b | 5.89 Ba | 5.44 Ab | 5.18 Bc | 5.47 B | |||||
| EA2 | 5.53 b | 5.80 Ca | 5.29 Bc | 5.07 Cd | 5.41 C | |||||
| Lactic acid (LA, g/kg) | CK | 1.62 | 1.65 | 1.67 | 1.70 | 1.66 | 0.000 | ** | NS | NS |
| EA1 | 1.62 | 1.63 | 1.70 | 1.72 | 1.67 | |||||
| EA2 | 1.62 | 1.65 | 1.68 | 1.74 | 1.67 | |||||
| Acetic acid (AA, g/kg) | CK | 0.27 | 0.62 A | 0.96 | 1.77 | 0.97 A | 0.002 | ** | ** | NS |
| EA1 | 0.28 | 0.43 B | 0.90 | 1.68 | 0.82 B | |||||
| EA2 | 0.23 | 0.37 B | 0.71 | 1.57 | 0.72C | |||||
| Propionic acid (PA, g/kg) | CK | ND | ND | ND | ND | - | - | - | - | - |
| EA1 | ND | ND | ND | ND | - | - | - | - | - | |
| EA2 | ND | ND | ND | ND | - | - | - | - | - | |
| Butyric acid (BA, g/kg) | CK | ND | ND | ND | ND | - | - | - | - | - |
| EA1 | ND | ND | ND | ND | - | - | - | - | - | |
| EA2 | ND | ND | ND | ND | - | - | - | - | - | |
| Lactic acid bacteria (LAB, log10·cfu/g FM) | CK | 7.93 a | 7.71 b | 7.27 c | 7.20 c | 7.52 | 0.03 | ** | NS | * |
| EA1 | 7.59 ab | 7.94 b | 7.35 b | 7.30 b | 7.55 | |||||
| EA2 | 7.86 a | 7.64 ab | 7.49 b | 7.12 c | 7.53 | |||||
| Coliform bacteria (log10·cfu/g FM) | CK | 7.10 Ab | 7.48 a | 6.56 c | 4.72 d | 6.62 A | 0.05 | ** | ** | ** |
| EA1 | 6.59 Bb | 7.73 a | 6.65 b | 3.74 c | 6.18 B | |||||
| EA2 | 6.29 Ca | 7.38 a | 6.63 b | 3.15 d | 6.11 B | |||||
| Yeasts (log10·cfu/g FM) | CK | 2.62 | <2.00 | <2.00 | 2.95 | 2.80 | 0.21 | NS | NS | NS 2 |
| EA1 | <2.00 | 3.01 | <2.00 | 2.45 | 2.73 | |||||
| EA2 | 3.52 | <2.00 | 2.54 | 2.00 | 2.69 | |||||
| Item | Treatment | Ensiling Days | Means | SEM | p-Value | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| 3 | 7 | 14 | 30 | D | T | D*T | ||||
| Crude protein (% DM) | CK | 11.1 Ba | 9.56 Bb | 10.0 b | 8.94 c | 9.91 B | 0.08 | ** | * | * |
| EA1 | 12.1 Aa | 9.82 Bb | 10.8 b | 9.34 b | 10.4 A | |||||
| EA2 | 10.8 Ba | 10.8 Aa | 10.7 a | 9.08 b | 10.3 A | |||||
| True protein (% TN) | CK | 51.9 A | 52.4 | 50.9 | 52.0 | 51.8 | 0.58 | NS | NS | NS |
| EA1 | 45.2 B | 56.3 | 49.4 | 51.3 | 51.05 | |||||
| EA2 | 54.3 Aa | 48.8 b | 47.3 b | 55.4 a | 51.5 | |||||
| Non-protein nitrogen (% TN) | CK | 48.1 B | 47.6 | 49.1 | 48.0 | 48.2 | 0.58 | NS | NS | ** |
| EA1 | 54.7 A | 43.7 | 50.6 | 48.7 | 48.9 | |||||
| EA2 | 45.7 Bb | 51.2 a | 52.7 a | 44.6 b | 48.5 | |||||
| Ammonium nitrogen (% TN) | CK | 2.61 d | 5.07 Ac | 6.31 b | 9.05 a | 5.76 A | 0.14 | ** | ** | NS |
| EA1 | 1.97 d | 3.59 Bc | 5.62 b | 8.6 a | 4.95 B | |||||
| EA2 | 2.19 d | 2.99 Bc | 4.55 b | 7.54 a | 4.51 B | |||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zou, X.; Chen, D.; Lv, H.; Zhang, Q.; Zheng, P. Effect of Ellagic Acid on Fermentation Quality and Bacterial Community of Stylo Silage. Fermentation 2021, 7, 256. https://doi.org/10.3390/fermentation7040256
Zou X, Chen D, Lv H, Zhang Q, Zheng P. Effect of Ellagic Acid on Fermentation Quality and Bacterial Community of Stylo Silage. Fermentation. 2021; 7(4):256. https://doi.org/10.3390/fermentation7040256
Chicago/Turabian StyleZou, Xuan, Dandan Chen, Hongjian Lv, Qing Zhang, and Peng Zheng. 2021. "Effect of Ellagic Acid on Fermentation Quality and Bacterial Community of Stylo Silage" Fermentation 7, no. 4: 256. https://doi.org/10.3390/fermentation7040256
APA StyleZou, X., Chen, D., Lv, H., Zhang, Q., & Zheng, P. (2021). Effect of Ellagic Acid on Fermentation Quality and Bacterial Community of Stylo Silage. Fermentation, 7(4), 256. https://doi.org/10.3390/fermentation7040256