
Increase in Fruity Ester Production during Spine Grape Wine Fermentation by Goal-Directed Amino Acid Supplementation


Abstract
:1. Introduction
2. Materials and Methods
2.1. Grape Material
2.2. Chemicals, Standards, and Microorganisms
2.3. Winemaking and Nitrogen Supplementation
2.4. Amino Acid Analysis
2.5. Organic Acid and Glycerol Analyses
2.6. Yeast Assimilable Nitrogen Analysis
2.7. Quantification of Volatiles
2.8. Statistical Analysis
3. Results
3.1. Evolution of Amino Acids and Fruity Esters during Alcoholic Fermentation
3.2. Correlation Analysis and Identification of Goal-Directed Amino Acid
3.3. Modification of Fruity Ester Production under Nitrogen Supplementation
3.4. Analysis of Extracellular Metabolites and Precursors Involved in Fruity Ester Synthesis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bell, S.J.; Henschke, P.A. Implications of nitrogen nutrition for grapes, fermentation and wine. Aust. J. Grape Wine Res. 2005, 11, 242–295. [Google Scholar] [CrossRef]
- Gobert, A.; Tourdot-Marechal, R.; Sparrow, C.; Morge, C.; Alexandre, H. Influence of nitrogen status in wine alcoholic fermentation. Food Microbiol. 2019, 83, 71–85. [Google Scholar] [CrossRef]
- Gutierrez-Gamboa, G.; Carrasco-Quiroz, M.; Martinez-Gil, A.M.; Perez-Alvarez, E.P.; Garde-Cerdan, T.; Moreno-Simunovic, Y. Grape and wine amino acid composition from Carignan noir grapevines growing under rainfed conditions in the Maule Valley, Chile: Effects of location and rootstock. Food Res. Int. 2018, 105, 344–352. [Google Scholar] [CrossRef] [PubMed]
- Vilanova, M.; Ugliano, M.; Varela, C.; Siebert, T.; Pretorius, I.S.; Henschke, P.A. Assimilable nitrogen utilisation and production of volatile and non-volatile compounds in chemically defined medium by Saccharomyces cerevisiae wine yeasts. Appl. Microbiol. Biotechnol 2007, 77, 145–157. [Google Scholar] [CrossRef] [Green Version]
- Lytra, G.; Tempere, S.; Le Floch, A.; de Revel, G.; Barbe, J.C. Study of sensory interactions among red wine fruity esters in a model solution. J. Agric. Food Chem. 2013, 61, 8504–8513. [Google Scholar] [CrossRef] [PubMed]
- Hu, K.; Jin, G.J.; Mei, W.C.; Li, T.; Tao, Y.S. Increase of medium-chain fatty acid ethyl ester content in mixed H. uvarum/S. cerevisiae fermentation leads to wine fruity aroma enhancement. Food Chem. 2018, 239, 495–501. [Google Scholar] [CrossRef]
- Fairbairn, S.; McKinnon, A.; Musarurwa, H.T.; Ferreira, A.C.; Bauer, F.F. The Impact of Single Amino Acids on Growth and Volatile Aroma Production by Saccharomyces cerevisiae Strains. Front. Microbiol. 2017, 8, 2554. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Procopio, S.; Krause, D.; Hofmann, T.; Becker, T. Significant amino acids in aroma compound profiling during yeast fermentation analyzed by PLS regression. LWT-Food Sci. Technol. 2013, 51, 423–432. [Google Scholar] [CrossRef]
- Hernández-Orte, P.; Cacho, J.F.; Ferreira, V. Relationship between varietal amino acid profile of grapes and wine aromatic composition. Experiments with model solutions and chemometric study. J. Agric. Food Chem. 2002, 50, 2891–2899. [Google Scholar] [CrossRef]
- Garde-Cerdán, T.; Martínez-Gil, A.M.; Lorenzo, C.; Lara, J.F.; Pardo, F.; Salinas, M.R. Implications of nitrogen compounds during alcoholic fermentation from some grape varieties at different maturation stages and cultivation systems. Food Chem. 2011, 124, 106–116. [Google Scholar] [CrossRef]
- Torrea, D.; Varela, C.; Ugliano, M.; Ancin-Azpilicueta, C.; Leigh Francis, I.; Henschke, P.A. Comparison of inorganic and organic nitrogen supplementation of grape juice—Effect on volatile composition and aroma profile of a Chardonnay wine fermented with Saccharomyces cerevisiae yeast. Food Chem. 2011, 127, 1072–1083. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.; Chia, J.Y.; Liu, S.Q. Impact of addition of aromatic amino acids on non-volatile and volatile compounds in lychee wine fermented with Saccharomyces cerevisiae MERIT.ferm. Int. J. Food Microbiol. 2014, 170, 12–20. [Google Scholar] [CrossRef]
- Ugliano, M.; Travis, B.; Francis, I.L.; Henschke, P.A. Volatile composition and sensory properties of Shiraz wines as affected by nitrogen supplementation and yeast species: Rationalizing nitrogen modulation of wine aroma. J. Agric. Food Chem. 2010, 58, 12417–12425. [Google Scholar] [CrossRef] [PubMed]
- Hazelwood, L.A.; Daran, J.M.; van Maris, A.J.; Pronk, J.T.; Dickinson, J.R. The Ehrlich pathway for fusel alcohol production: A century of research on Saccharomyces cerevisiae metabolism. Appl. Environ. Microbiol. 2008, 74, 2259–2266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malcorps, P.; Dufour, J.P. Short-chain and medium-chain aliphatic-ester synthesis in Saccharomyces cerevisiae. Eur. J. Biochem. 1992, 210, 1015–1022. [Google Scholar] [CrossRef]
- Crepin, L.; Truong, N.M.; Bloem, A.; Sanchez, I.; Dequin, S.; Camarasa, C. Management of Multiple Nitrogen Sources during Wine Fermentation by Saccharomyces cerevisiae. Appl. Environ. Microbiol. 2017, 83. [Google Scholar] [CrossRef] [Green Version]
- Rollero, S.; Mouret, J.R.; Bloem, A.; Sanchez, I.; Ortiz-Julien, A.; Sablayrolles, J.M.; Dequin, S.; Camarasa, C. Quantitative (13) C-isotope labelling-based analysis to elucidate the influence of environmental parameters on the production of fermentative aromas during wine fermentation. Microb. Biotechnol. 2017, 10, 1649–1662. [Google Scholar] [CrossRef]
- Liu, P.; Wang, Y.; Ye, D.; Duan, L.; Duan, C.; Yan, G. Effect of the addition of branched-chain amino acids to non-limited nitrogen synthetic grape must on volatile compounds and global gene expression during alcoholic fermentation. Aust. J. Grape Wine Res. 2018, 24, 197–205. [Google Scholar] [CrossRef]
- Meng, J.; Xu, T.; Qin, M.; Zhuang, X.; Fang, Y.; Zhang, Z. Phenolic characterization of young wines made from spine grape (Vitis davidii Foex) grown in Chongyi County (China). Food Res. Int. 2012, 49, 664–671. [Google Scholar] [CrossRef]
- Kong, C.; Li, A.; Su, J.; Wang, X.; Chen, C.; Tao, Y. Flavor modification of dry red wine from Chinese spine grape by mixed fermentation with Pichia fermentans and S. cerevisiae. LWT-Food Sci. Technol. 2019, 109, 83–92. [Google Scholar] [CrossRef]
- Antalick, G.; Suklje, K.; Blackman, J.W.; Meeks, C.; Deloire, A.; Schmidtke, L.M. Influence of Grape Composition on Red Wine Ester Profile: Comparison between Cabernet Sauvignon and Shiraz Cultivars from Australian Warm Climate. J. Agric. Food Chem. 2015, 63, 4664–4672. [Google Scholar] [CrossRef]
- Zhu, Z.; Chen, S.; Su, J.; Tao, Y. Correlation Analysis between Amino Acids and Fruity Esters during Spine Grape Fermentation. Sci. Agric. Sin. 2020, 53, 2272–2284. [Google Scholar] [CrossRef]
- Li, N.; Liu, Y.; Zhao, Y.; Zheng, X.; Lu, J.; Liang, Y. Simultaneous HPLC determination of amino acids in tea infusion coupled to pre-column derivatization with 2, 4-dinitrofluorobenzene. Food Anal. Methods 2016, 9, 1307–1314. [Google Scholar] [CrossRef]
- Zoecklein, B.; Fugelsang, K.; Gump, B.; Nury, F. Wine Analysis and Production; Chapman & Hall. Inc.: New York, NY, USA, 1995. [Google Scholar]
- Hu, K.; Jin, G.J.; Xu, Y.H.; Xue, S.J.; Qiao, S.J.; Teng, Y.X.; Tao, Y.S. Enhancing wine ester biosynthesis in mixed Hanseniaspora uvarum/Saccharomyces cerevisiae fermentation by nitrogen nutrient addition. Food Res. Int. 2019, 123, 559–566. [Google Scholar] [CrossRef] [PubMed]
- Huang, Z.; Ough, C.S. Amino acid profiles of commercial grape juices and wines. Am. J. Enol. Vitic. 1991, 42, 261–267. [Google Scholar]
- Roca-Mesa, H.; Sendra, S.; Mas, A.; Beltran, G.; Torija, M.J. Nitrogen Preferences during Alcoholic Fermentation of Different Non-Saccharomyces Yeasts of Oenological Interest. Microorganisms 2020, 8, 157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, N.; Li, L.; Deng, X.; Song, L. Changes of free amino acids during wine fermentation. China Brew. 2011, 1, 28–33. [Google Scholar] [CrossRef]
- Braus, G.H. Aromatic amino acid biosynthesis in the yeast Saccharomyces cerevisiae: A model system for the regulation of a eukaryotic biosynthetic pathway. Microbiol. Rev. 1991, 55, 349–370. [Google Scholar] [CrossRef]
- Beltran, G.; Esteve-Zarzoso, B.; Rozès, N.; Mas, A.; Guillamón, J.M. Influence of the timing of nitrogen additions during synthetic grape must fermentations on fermentation kinetics and nitrogen consumption. J. Agric. Food Chem. 2005, 53, 996–1002. [Google Scholar] [CrossRef]
- Ebeler, S.E.; Thorngate, J.H. Wine chemistry and flavor: Looking into the crystal glass. J. Agric. Food Chem. 2009, 57, 8098–8108. [Google Scholar] [CrossRef]
- Meng, J.; Xu, T.; Song, C.; Li, X.; Yue, T.; Qin, M.; Fang, Y.; Zhang, Z.; Xi, Z. Characteristic free aromatic components of nine clones of spine grape (Vitis davidii Foex) from Zhongfang County (China). Food Res. Int. 2013, 54, 1795–1800. [Google Scholar] [CrossRef]
- Martínez-Pinilla, O.; Guadalupe, Z.; Hernández, Z.; Ayestarán, B. Amino acids and biogenic amines in red varietal wines: The role of grape variety, malolactic fermentation and vintage. Eur. Food Res. Technol. 2013, 237, 887–895. [Google Scholar] [CrossRef]
- Garde-Cerdán, T.; Ancín-Azpilicueta, C. Effect of the addition of different quantities of amino acids to nitrogen-deficient must on the formation of esters, alcohols, and acids during wine alcoholic fermentation. LWT-Food Sci. Technol. 2008, 41, 501–510. [Google Scholar] [CrossRef]
- Hernández-Orte, P.; Ibarz, M.; Cacho, J.; Ferreira, V. Effect of the addition of ammonium and amino acids to musts of Airen variety on aromatic composition and sensory properties of the obtained wine. Food Chem. 2005, 89, 163–174. [Google Scholar] [CrossRef]
- Frick, O.; Wittmann, C. Characterization of the metabolic shift between oxidative and fermentative growth in Saccharomyces cerevisiae by comparative 13 C flux analysis. Microb. Cell Fact. 2005, 4, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Saerens, S.M.; Delvaux, F.R.; Verstrepen, K.J.; Thevelein, J.M. Production and biological function of volatile esters in Saccharomyces cerevisiae. Microb. Biotechnol. 2010, 3, 165–177. [Google Scholar] [CrossRef] [Green Version]
- Boutou, S.; Chatonnet, P. Rapid headspace solid-phase microextraction/gas chromatographic/mass spectrometric assay for the quantitative determination of some of the main odorants causing off-flavours in wine. J. Chromatogr. A 2007, 1141, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Magasanik, B.; Kaiser, C.A. Nitrogen regulation in Saccharomyces cerevisiae. Gene 2002, 290, 1–18. [Google Scholar] [CrossRef]
- Bloem, A.; Sanchez, I.; Dequin, S.; Camarasa, C. Metabolic Impact of Redox Cofactor Perturbations on the Formation of Aroma Compounds in Saccharomyces cerevisiae. Appl. Environ. Microbiol. 2016, 82, 174–183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saerens, S.; Delvaux, F.; Verstrepen, K.; Van Dijck, P.; Thevelein, J.; Delvaux, F. Parameters affecting ethyl ester production by Saccharomyces cerevisiae during fermentation. Appl. Environ. Microbiol. 2008, 74, 454–461. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Q.; Wang, X.; Xu, Y. Palladium-Catalyzed Tandem Reactions of β-(2-Bromophenyl)-α,β-Unsaturated Carbonyl Compounds with 2-Hydroxyphenylboronic Acid: A New Route to Benzo[c]chromenes. Eur. J. Org. Chem. 2012, 2012, 1112–1114. [Google Scholar] [CrossRef]
- Mason, A.B.; Dufour, J.P. Alcohol acetyltransferases and the significance of ester synthesis in yeast. Yeast 2000, 16, 1287–1298. [Google Scholar] [CrossRef]
- Mauricio, J.C.; Valero, E.; Millán, C.; Ortega, J.M. Changes in nitrogen compounds in must and wine during fermentation and biological aging by flor yeasts. J. Agric. Food Chem. 2001, 49, 3310–3315. [Google Scholar] [CrossRef] [PubMed]
- Farina, L.; Medina, K.; Urruty, M.; Boido, E.; Dellacassa, E.; Carrau, F. Redox effect on volatile compound formation in wine during fermentation by Saccharomyces cerevisiae. Food Chem. 2012, 134, 933–939. [Google Scholar] [CrossRef]
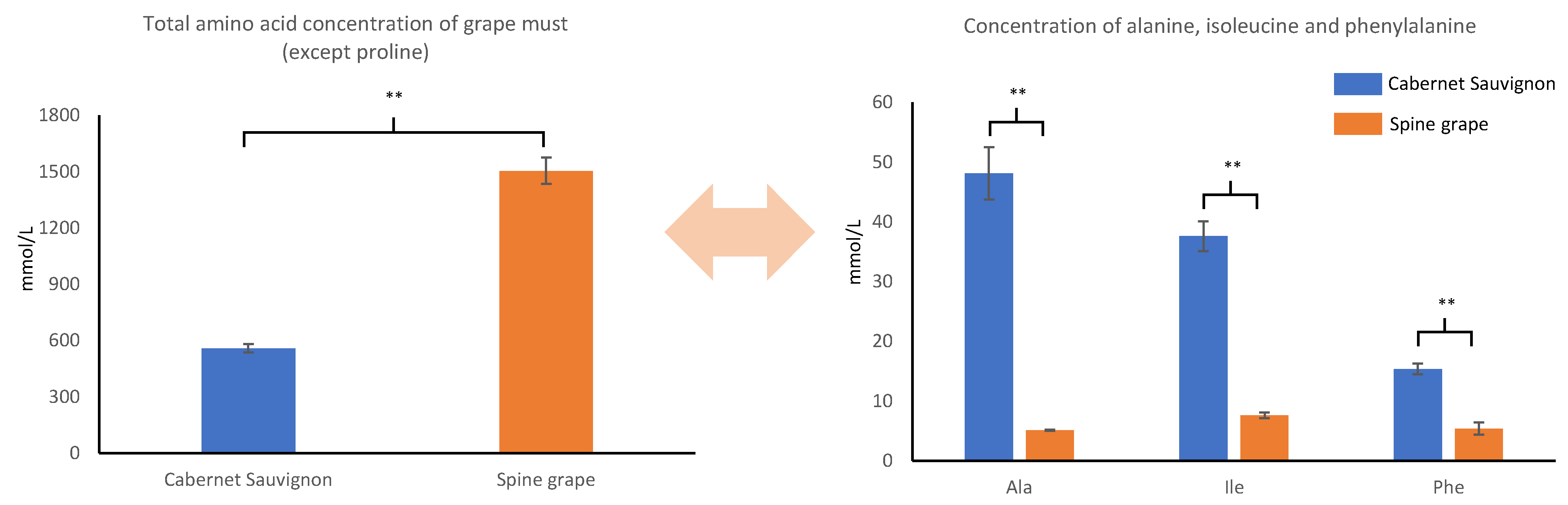

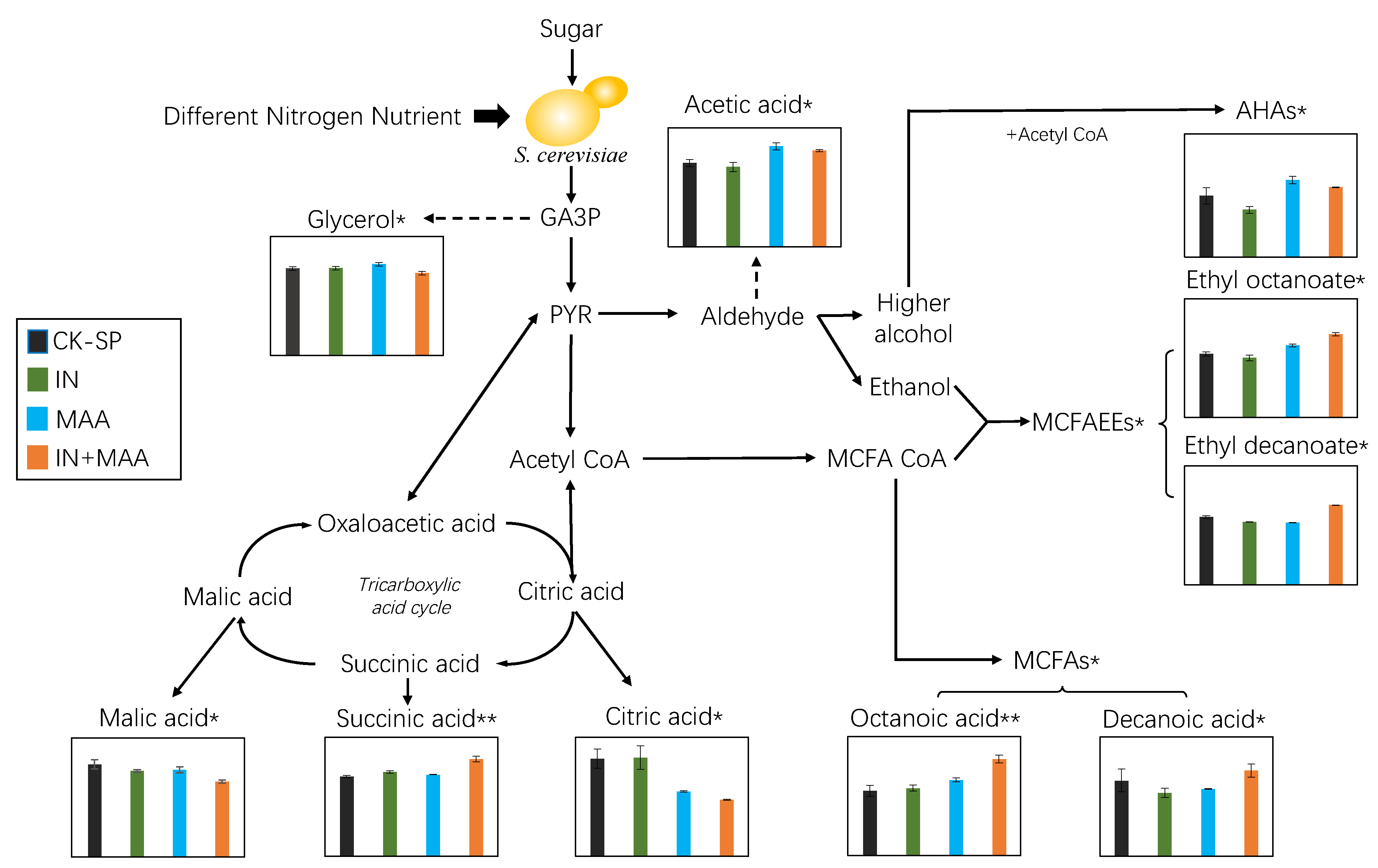

{kind=link}
{kind=link}
{kind=link}
| Supplement | Composition |
|---|---|
| IN | DAP, 236 mg/L |
| AA-Ala | Alanine, 318 mg/L |
| AA-Phe | Phenylalanine, 589 mg/L |
| AA-Ile | Isoleucine, 468 mg/L |
| MAA | Alanine, 163 mg/L; phenylalanine, 71 mg/L; isoleucine, 170 mg/L |
| IN+MAA | Alanine, 39 mg/L; phenylalanine, 17 mg/L; isoleucine, 39 mg/L; DAP, 178 mg/L |
| Amino Acid | AHAs | SCFAEEs | MCFAEEs |
|---|---|---|---|
| Total AAs | −0.274 | −0.011 | −0.075 |
| Arg | −0.185 | −0.010 | 0.031 |
| Asp | −0.186 | 0.080 | −0.002 |
| Glu | 0.038 | 0.123 | 0.143 |
| GABA | −0.164 | 0.149 | −0.044 |
| Leu | −0.143 | 0.011 | −0.002 |
| Met | −0.101 | 0.095 | 0.072 |
| Trp | −0.123 | 0.004 | 0.024 |
| Val | −0.136 | −0.064 | 0.045 |
| Ala | −0.251 | −0.156 | −0.022 |
| Ile | −0.099 | −0.020 | −0.157 |
| Phe | −0.406 | −0.266 | −0.237 |
| Compounds | CK-CS | CK-SP | IN | AA-Ala | AA-Phe | AA-Ile | MAA | IN+MAA |
|---|---|---|---|---|---|---|---|---|
| Ethyl acetate (μg/L) | 30,905 ± 1083b | 21,973 ± 436cd | 23,043 ± 1397cd | 25,454 ± 5621bc | 20,857 ± 2039cd | 18,409 ± 2093d | 45,946 ± 899a | 28,962 ± 2052b |
| AHAs (μg/L) | 1573 ± 14a | 745 ± 99cde | 574 ± 41e | 698 ± 62de | 1346 ± 97b | 1161 ± 83b | 937 ± 44c | 847 ± 4cd |
| Isobutyl acetate | 41 ± 2c | 29 ± 2c | 76 ± 4b | 112 ± 18a | 74 ± 5b | 110 ± 12a | 119 ± 15a | 86 ± 4b |
| Isoamyl acetate | 1346 ± 7a | 584 ± 87cd | 411 ± 31e | 505 ± 39de | 508 ± 55de | 964 ± 66b | 689 ± 22c | 634 ± 0de |
| Hexyl acetate | 54 ± 1a | 36 ± 7bcd | 33 ± 4cd | 32 ± 1cd | 38 ± 1bcd | 31 ± 0d | 39 ± 1bc | 42 ± 0b |
| β-Phenylethyl acetate | 132 ± 4b | 96 ± 3c | 54 ± 2d | 49 ± 4d | 726 ± 36a | 56 ± 5d | 90 ± 6c | 85 ± 0c |
| SCFAEEs (μg/L) | 140 ± 3b | 54 ± 1c | 38 ± 30c | 250 ± 57a | 210 ± 8a | 212 ± 39a | 106 ± 9bc | 232 ± 26a |
| Ethyl butyrate | 140 ± 3 | 54 ± 1 | 38 ± 30 | 250 ± 57 | 210 ± 8 | 212 ± 39 | 106 ± 9 | 232 ± 26 |
| MCFAEEs (μg/L) | 2101 ± 43b | 1867 ± 63c | 1858 ± 75c | 2290 ± 242ab | 2423 ± 220a | 1954 ± 97bc | 2125 ± 39b | 2485 ± 65a |
| Ethyl hexanoate | 817 ± 2a | 576 ± 24d | 575 ± 39d | 695 ± 77bc | 739 ± 63ab | 668 ± 34bcd | 691 ± 22bc | 762 ± 42ab |
| Ethyl octanoate | 754 ± 1c | 751 ± 27c | 753 ± 35c | 934 ± 88b | 1056 ± 79a | 847 ± 34bc | 910 ± 17b | 1050 ± 21a |
| Ethyl decanoate | 530 ± 40ab | 540 ± 12ab | 530 ± 1ab | 661 ± 77a | 628 ± 78a | 439 ± 29b | 524 ± 0ab | 673 ± 2a |
| Higher alcohols (mg/L) | 972.15 ± 26.45a | 597.29 ± 54.96cd | 471.12 ± 29.12d | 604.04 ± 93.42cd | 796.24 ± 53.59b | 994.98 ± 64.11a | 707.40 ± 19.11bc | 558.89 ± 19.84cd |
| Isobutanol | 39.15 ± 2.52a | 27.10 ± 2.87bc | 20.48 ± 1.57c | 42.75 ± 7.03a | 23.20 ± 1.19bc | 41.03 ± 2.52a | 43.75 ± 1.31a | 30.14 ± 2.09b |
| Isopentanol | 800.05 ± 19.86a | 502.08 ± 48.40bc | 398.67 ± 24.57c | 500.95 ± 78.78bc | 436.37 ± 34.42c | 890.35 ± 56.28a | 577.80 ± 12.87b | 477.30 ± 16.68bc |
| 1-hexanol | 4.91 ± 0.15a | 2.26 ± 0.30d | 3.03 ± 0.15c | 2.99 ± 0.43c | 3.02 ± 0.22c | 1.66 ± 0.09e | 3.09 ± 0.13c | 3.63 ± 0.19b |
| 2-phenylethanol | 128.04 ± 3.92b | 65.85 ± 3.39cde | 48.94 ± 2.83e | 57.35 ± 7.18de | 333.65 ± 17.76a | 61.94 ± 5.22de | 82.76 ± 4.80c | 47.82 ± 0.88cd |
| Volatile acids (mg/L) | 7.70 ± 0.53d | 7.74 ± 0.58d | 8.08 ± 0.50cd | 13.37 ± 0.97a | 11.75 ± 0.47b | 8.39 ± 1.02cd | 9.28 ± 0.37c | 11.22 ± 0.30b |
| Hexanoic acid | 4.24 ± 0.26d | 4.15 ± 0.27d | 4.51 ± 0.34cd | 7.43 ± 0.70a | 5.76 ± 0.28b | 4.64 ± 0.22cd | 5.29 ± 0.26bc | 6.14 ± 0.09b |
| Octanoic acid | 3.27 ± 0.36c | 3.44 ± 0.28c | 3.41 ± 0.15c | 5.70 ± 0.26a | 5.74 ± 0.18a | 3.58 ± 0.78c | 3.82 ± 0.10c | 4.86 ± 0.20b |
| Decanoic acid | 0.19 ± 0.01bc | 0.18 ± 0.03bc | 0.16 ± 0.01c | 0.24 ± 0.01a | 0.25 ± 0.01a | 0.17 ± 0.02c | 0.17 ± 0.01bc | 0.22 ± 0.01ab |
| Compounds | CK-CS | CK-SP | IN | AA-Ala | AA-Phe | AA-Ile | MAA | IN+MAA |
|---|---|---|---|---|---|---|---|---|
| Glycerol | 7.47 ± 0.12 | 7.26 ± 0.22b | 7.36 ± 0.28ab | 7.65 ± 0.24 | 7.63 ± 0.19 | 7.76 ± 0.26 | 7.73 ± 0.30a | 6.98 ± 0.15c |
| Malic acid | 2.51 ± 0.14 | 0.78 ± 0.04a | 0.72 ± 0.01a | 0.65 ± 0.01 | 0.71 ± 0.01 | 0.67 ± 0.01 | 0.73 ± 0.02a | 0.64 ± 0.01b |
| Acetic acid | 1.41 ± 0.01 | 0.71 ± 0.04b | 0.67 ± 0.03b | 0.77 ± 0.07 | 0.66 ± 0.03 | 0.74 ± 0.01 | 0.85 ± 0.03a | 0.81 ± 0.01a |
| Citric acid | 0.70 ± 0.06 | 0.33 ± 0.08a | 0.33 ± 0.04a | 0.25 ± 0.01 | 0.22 ± 0.01 | 0.25 ± 0.01 | 0.22 ± 0.01ab | 0.19 ± 0.01b |
| Succinic acid | 4.61 ± 0.10 | 1.01 ± 0.02b | 1.06 ± 0.02b | 0.92 ± 0.01 | 1.11 ± 0.01 | 0.92 ± 0.04 | 1.03 ± 0.01b | 1.22 ± 0.04a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhu, Z.; Hu, K.; Chen, S.; Xiong, S.; Tao, Y. Increase in Fruity Ester Production during Spine Grape Wine Fermentation by Goal-Directed Amino Acid Supplementation. Fermentation 2021, 7, 231. https://doi.org/10.3390/fermentation7040231
Zhu Z, Hu K, Chen S, Xiong S, Tao Y. Increase in Fruity Ester Production during Spine Grape Wine Fermentation by Goal-Directed Amino Acid Supplementation. Fermentation. 2021; 7(4):231. https://doi.org/10.3390/fermentation7040231
Chicago/Turabian StyleZhu, Zijian, Kai Hu, Siyu Chen, Sirui Xiong, and Yongsheng Tao. 2021. "Increase in Fruity Ester Production during Spine Grape Wine Fermentation by Goal-Directed Amino Acid Supplementation" Fermentation 7, no. 4: 231. https://doi.org/10.3390/fermentation7040231
APA StyleZhu, Z., Hu, K., Chen, S., Xiong, S., & Tao, Y. (2021). Increase in Fruity Ester Production during Spine Grape Wine Fermentation by Goal-Directed Amino Acid Supplementation. Fermentation, 7(4), 231. https://doi.org/10.3390/fermentation7040231