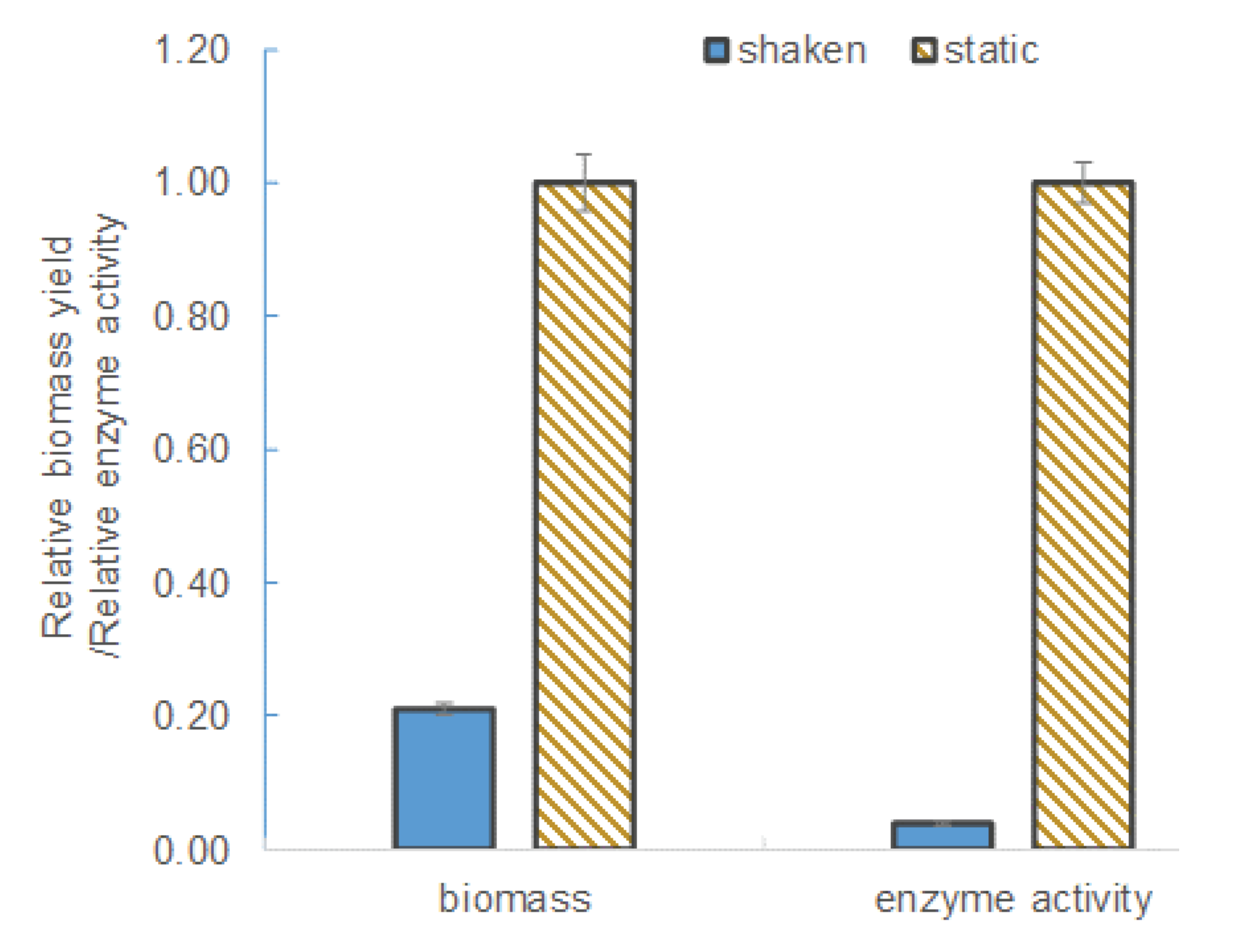
L. leichmannii 313 is classified by ATCC as a microaerophilic organism. Therefore, unlike strict anaerobes, this species is able to survive in the presence of low levels of oxygen. In this study, culture media grown under agitation (shaken at 100 rpm) conditions yielded significantly lower levels of β-galactosidase and biomass (see
Figure 2). The biomass yield and enzyme activity levels in static culture (i.e., those grown without shaking) were ~ 5-fold and 30-fold higher, respectively, than in the shaken cultures. This observation can be attributed to the effect of oxygen or aeration on the growth and metabolism of
L. leichmannii 313. Factors that promote aerobic conditions (including intense agitation) have been shown to significantly inhibit the growth of lactobacilli [
25,
35]. Moreover, previous studies have shown that aerobic conditions inhibit cell growth and the production of certain enzymes (e.g., cell-envelope proteinases) by
L. leichmannii 313 [
36]. This is due to a switch in the usual homolactic fermentation that results in a lower ATP yield per glucose consumed, as well as activation of the enzyme pyruvate oxidase, which converts the oxygen in the media to hydrogen peroxide [
36]. Hydrogen peroxide is a bacteriostatic compound that inhibits cell growth, and this accounts for the low levels of biomass. Static cultures, on the other hand, preserve a near anaerobic condition in the culture, resulting in higher biomass and enzyme yields. The higher biomass yields can be attributed to an efficient utilization of the carbon (i.e., glucose) [
37]. Typically, anaerobic conditions activate the pyruvate-formate lyase system, which, by the actions of acetyl phosphate and acetyl kinase, increases the efficiency of ATP generation per glucose consumed. The efficient sugar utilization and higher levels of ATP produced lead to higher biomass levels [
35,
38]. This phenomenon has been demonstrated in
Streptococcus mutans [
39], as well as in some lactobacilli, such as
L. johnsonii and
L. gasseri [
40],
L. plantarum [
41], and
L. leichmannii 313 [
36]. Based on these findings (shown in
Figure 2), subsequent experiments in this study were carried out under static conditions.
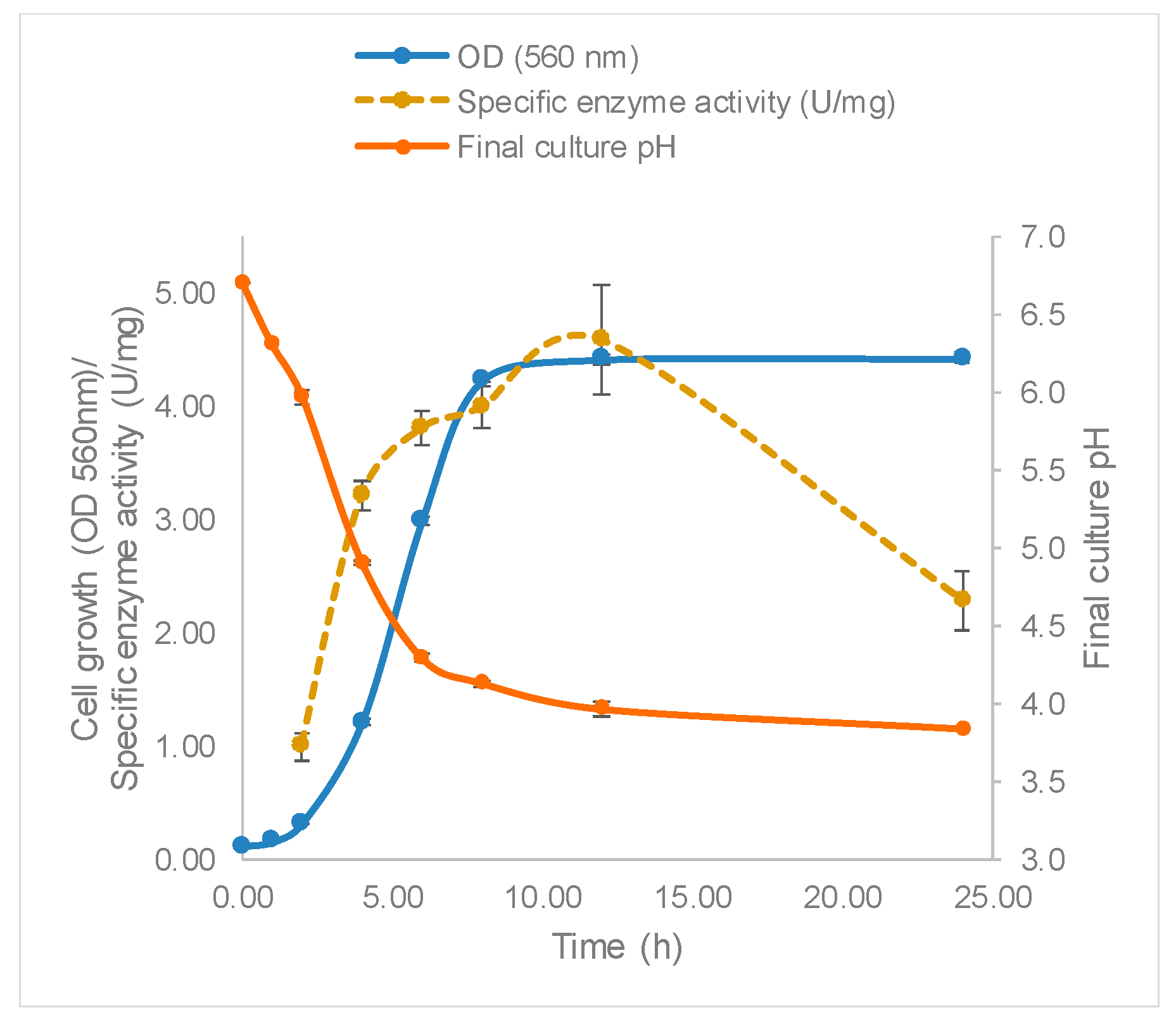
The growth of
L. leichmannii 313 and profile of β-galactosidase production are shown in
Figure 3. The Gompertz model showed that the lag phase was about 2.8 h, after which the bacteria proliferated rapidly at a growth rate of 1.0 per hour, until a maximum OD of 4.5 was attained after 10 h. Accompanying the increasing biomass was the specific enzyme activity, which reached a maximum of 4.58 U/mg after 12 h of growth. This time point that corresponds to the early stationary phase of bacteria growth. The fact that the β-galactosidase levels were highest in the early stationary phase is consistent with reports of other studies involving lactobacilli [
42]. Enzyme activity declined after 12 h, perhaps corresponding to the using up of substrates required for growth, or the production of organic acids leading to the lowering of media pH to levels that inhibit bacteria growth. The pH of the growth medium decreased and reached~3.9 after 24 h (
Figure 3,
Table 5).
3.1.1. Independent Factor Analysis
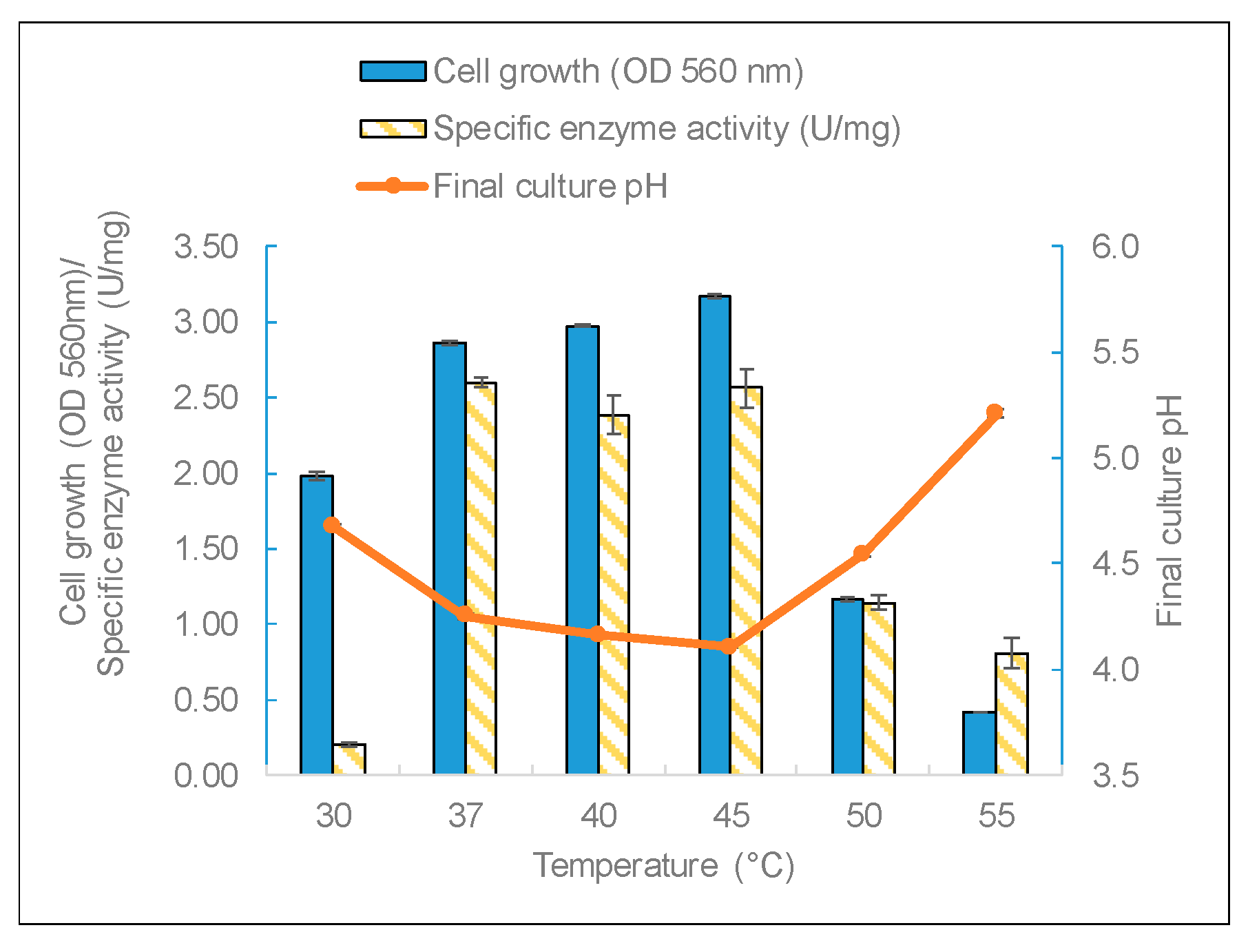
The three factors of temperature, pH, and carbon source were studied to identify the most important growth conditions that affect enzyme production. Temperature is a critical factor for bacteria growth.
L. leichmannii 313 had almost the same yield of biomass and enzymes between 37 and 45 °C (
Figure 4). The biomass yields (measured by OD at 560 nm) and final pH corroborated the results of the specific enzyme yields; that is to say, cell growth in the mesophilic temperature range of 37 to 45 °C recorded significantly higher levels of biomass (and a lower final pH) than at temperatures below 37 °C and above 45 °C. This indicates that temperature has a growth-dependent effect on β-galactosidase production by
L. leichmannii 313. Additionally, compared with other lactic acid bacteria, such as
Bifidobacterium longum [
43],
L. leichmannii 313 has a wider range of temperatures for β-galactosidase production. This optimum growth range is to be expected for a mesophilic lactobacillus. Moreover, considering that most dairy products are processed in this temperature range, it is an indication that
L. leichmannii 313 has the potential to be used as a starter that can withstand conditions employed in the processing of dairy products. The sharp decline in enzyme activity over 45 °C could be due to cell growth impairment at high temperatures. This is demonstrated by the low OD values and the corresponding high final culture pH. Moreover, temperatures below 37 °C were not optimum for the production and specific activity of β-galactosidase by
L. leichmannii 313.
Biomass and β-galactosidase production by
L. leichmannii 313 were significantly affected by the extracellular pH. This is to be expected since many enzymatic processes and the transport of components across cell membranes depend on the culture pH [
44,
45]. For example, the transport of sugars, such as lactose and galactose, by the action of lactose permease work via a symport mechanism where protons and sugar molecules are simultaneously translocated across the cell membrane [
46]. This suggests that a lower pH (i.e., abundance of protons) will favor the effective transport and metabolism of sugars. pH has also been shown to play a crucial role in the metabolism and microbial diversity of gut bacteria, and cultures grown at a lower starting pH (i.e., pH 6 rather than 6.9) experienced large pH drops at the end of the growth phase [
47]. In this study,
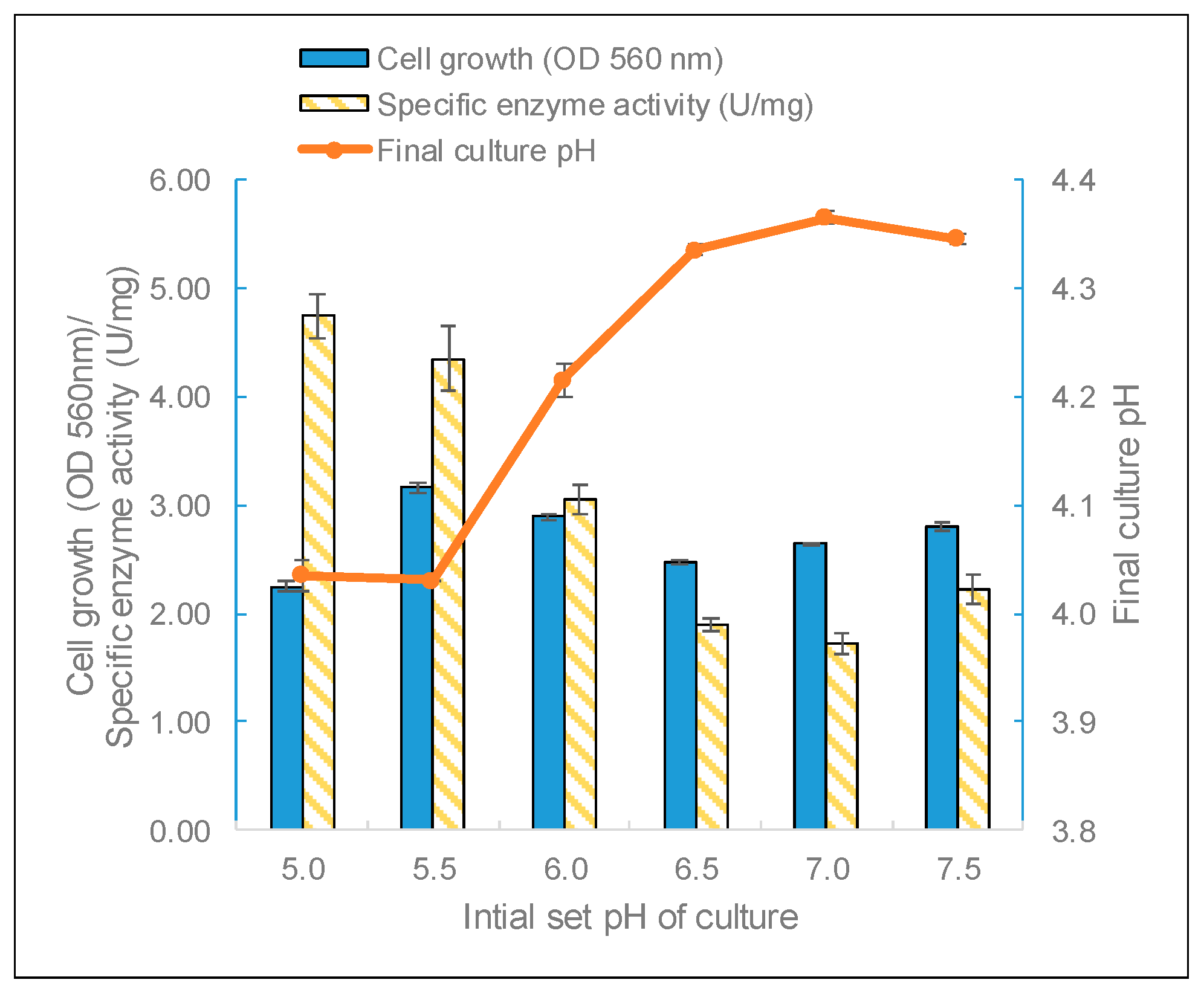
L. leichmannii 313 had the highest biomass yield when grown at a starting pH of 5.5, but the highest enzyme activity was observed for cultures grown at a starting pH of 5.0 (
Figure 5). Moreover, although
L. leichmannii 313 was cultured in different initial pH values (pH 5 to 7.5), the final pH values of cultures were very similar (between 4.0 and 4.3) (
Figure 4).
L. leichmannii 313 is a lactic acid bacterium, and a suitable pH for its cell growth is between 5.5 and 7.0 [
36]. However, in most bacteria, the optimum pH for cell growth and that for enzyme production could be different [
48], as has been observed in the growth and cell envelope proteinase production profile of
L. leichmannii 313 [
37]. Furthermore, another study involving the expression of recombinant β-galactosidase in
L. reuteri showed that pH was one of the most important factors for the production of β-galactosidase in this lactobacillus [
49].
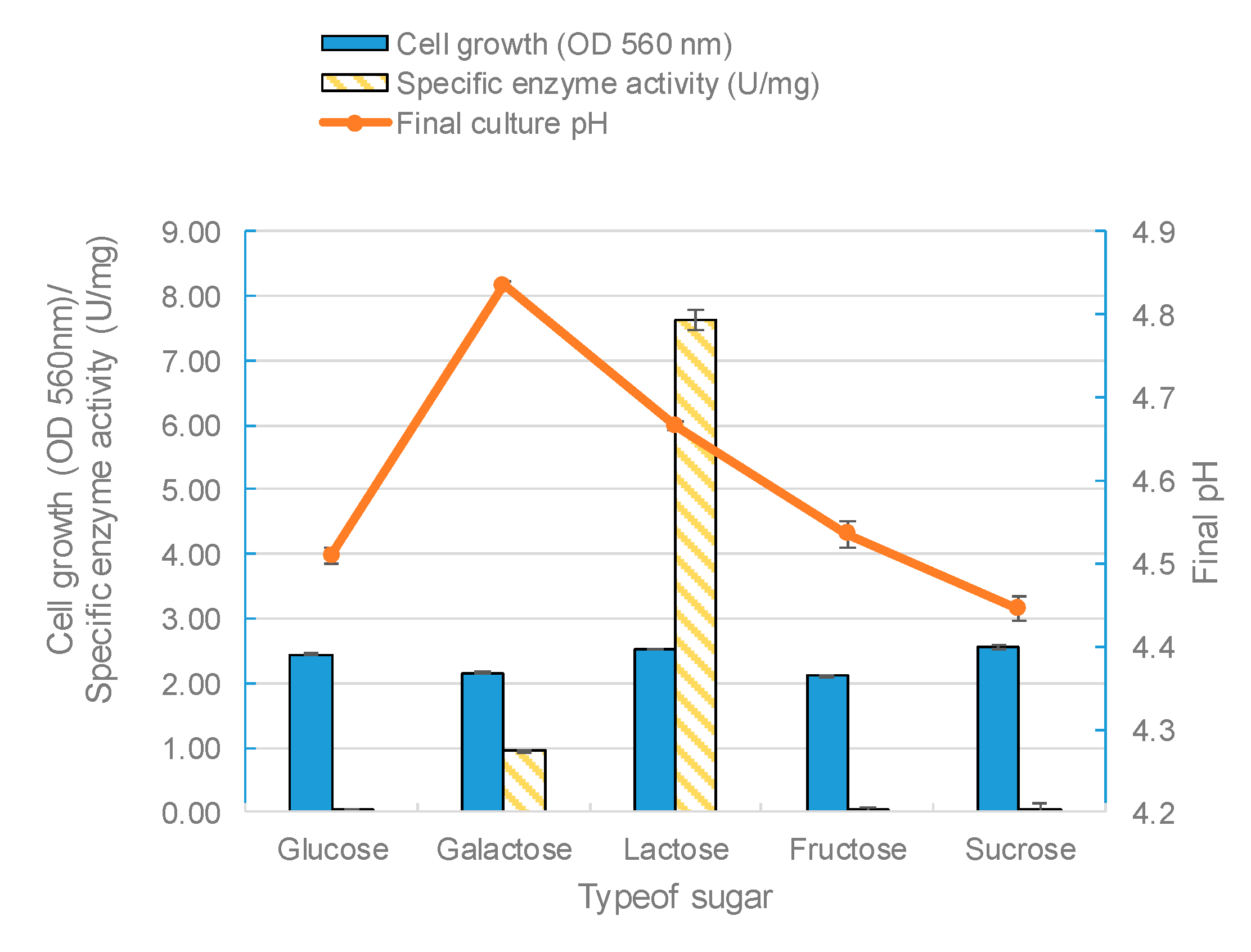
The effect of the carbon source on enzyme biosynthesis depends on the microbial species being studied [
50]. In fact, the carbon substrate type and concentration are two of the most important factors for the production of β-galactosidase in lactobacilli [
49]. In this study, five kinds of sugars (glucose, galactose, lactose, fructose, and sucrose) were used as carbon sources in the growth medium to find the optimum sugar for β-galactosidase synthesis. The modified MRS broth was prepared using each carbon substrate at a 2% (
w/
v) concentration. All sugars used could support the growth of
L. leichmannii 313. However, lactose as the sole carbon significantly promoted the activity of β-galactosidase. This was followed by galactose, with glucose giving the lowest enzyme activity (
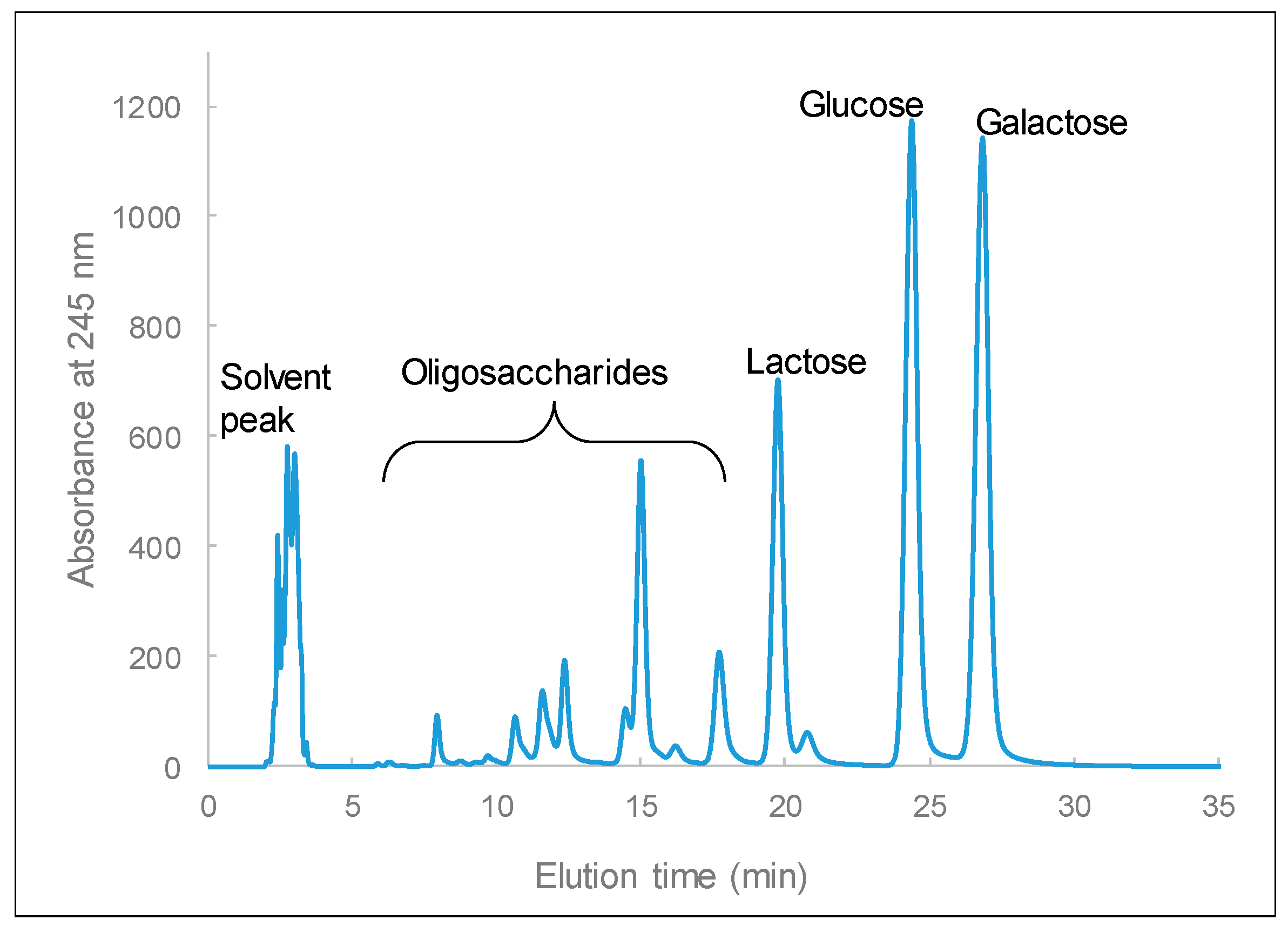
Figure 6). These results are compatible with previous studies [
43,
50], and also those of
Kluyveromyces fragilis and
Rhizomucor sp [
15,
51]. Furthermore, our results showed that fructose and sucrose, when used as sole carbon sources, generated biomasses with low specific enzyme activities. Lactose was the optimum carbon source that stimulated the profuse synthesis of β-galactosidase. This is because lactose (particularly in the form of the allolactose isomer) promotes the expression of the
lacZ gene by binding to the Lac repressor protein, preventing this protein from engaging the operator region of the
lac operon [
52,
53]. In most
L. delbrueckii species, the expression of β-galactosidase has been shown to be constitutive [
54]. However, in
L. delbrueckii subsp.
lactis strains (which includes
L.
leichmannii 313), β-galactosidase production is inducible, through the presence of lactose (and to a lesser extent galactose) in the growth media [
55]. Glucose, on the other hand, inhibits the expression of β-galactosidase via catabolite repression mechanisms. During catabolite repression, the presence of glucose leads to the production of lower levels of cyclic adenosine monophosphates (cAMPs) and activation of catabolite activator proteins that bind to and turn off the
lac operon [
56,
57].
3.1.2. Results of Factor Screening Experiments
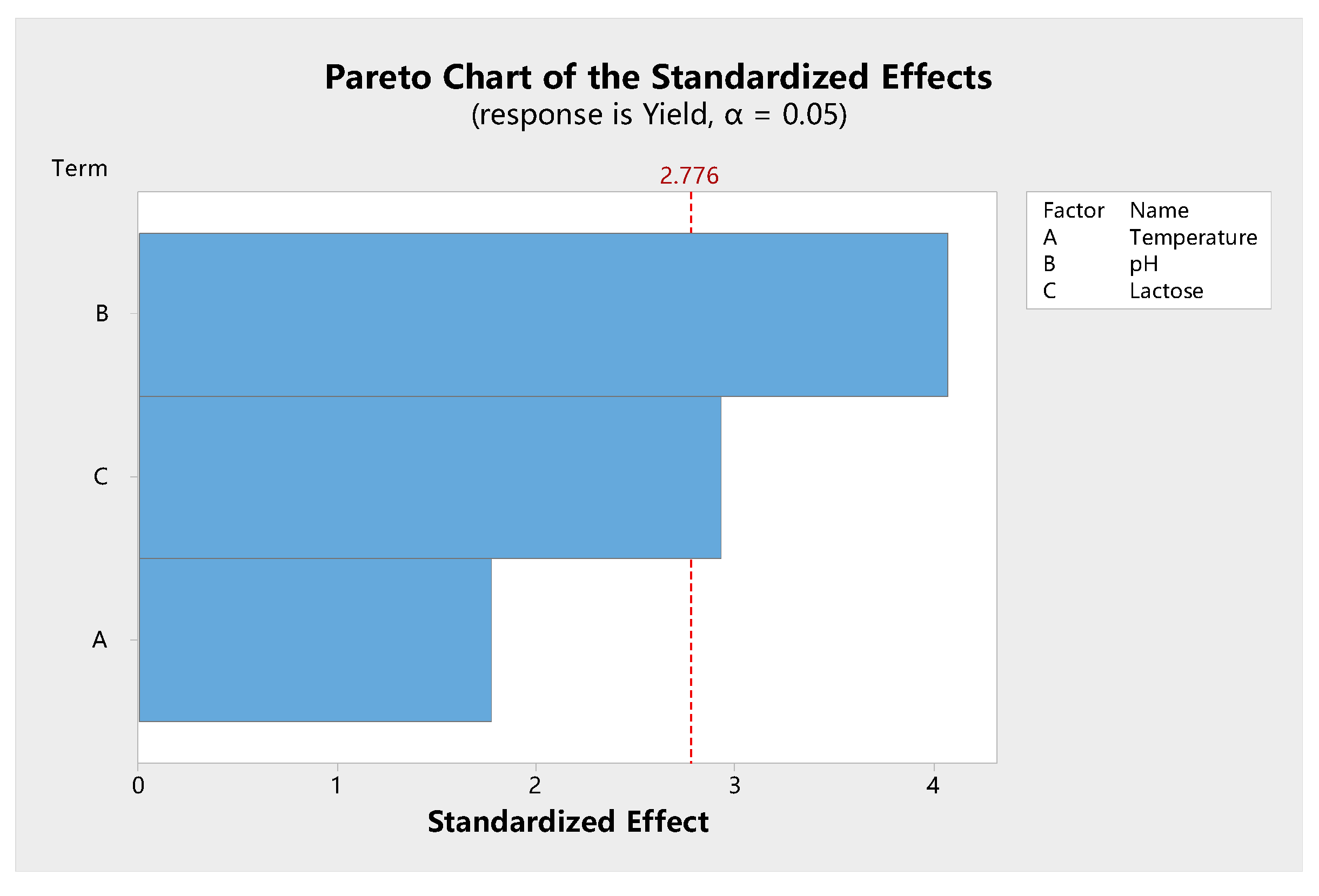
Shown in
Figure 7 is the Pareto chart of the effect and level of significance of various factors that influence β-galactosidase synthesis. The Pareto chart showed that the temperature, initial pH, and lactose concentration had a positive effect on specific enzyme activity. However, temperature did not cause a significant effect. The trends for pH and carbon source were different from those of the independent factor analysis. For example, the trend for lactose concentration agreed with those of other studies that have shown that an increased production/activity of β-galactosidase is observed as the concentration of lactose in the medium is increased up to 2.5% [
50] or 4.0% [
43]. In this study, temperature had no significant effect on enzyme activity (
p = 0.056). Our independent factor analysis also showed that there was no significant difference for enzyme activity between 37 and 45 °C. This indicates that
L. leichmannii 313 has a wide range of temperatures for β-galactosidase production, compared with other lactobacilli [
43]. In subsequent experiments, 37 °C was used for bacteria growth, as it is a common temperature for the growth of most microorganisms.
The results of Plackett–Burman design for two levels of independent factors is shown in
Table 6, and the predicted enzyme activity is based on Equation (3). An analysis of variance for these results (see
Table 7) shows that the model had a good fit to the experimental data (
R2 = 0.87).
3.1.3. Central Composite Design (CCD) and Response Surface Methodology (RSM) Analysis
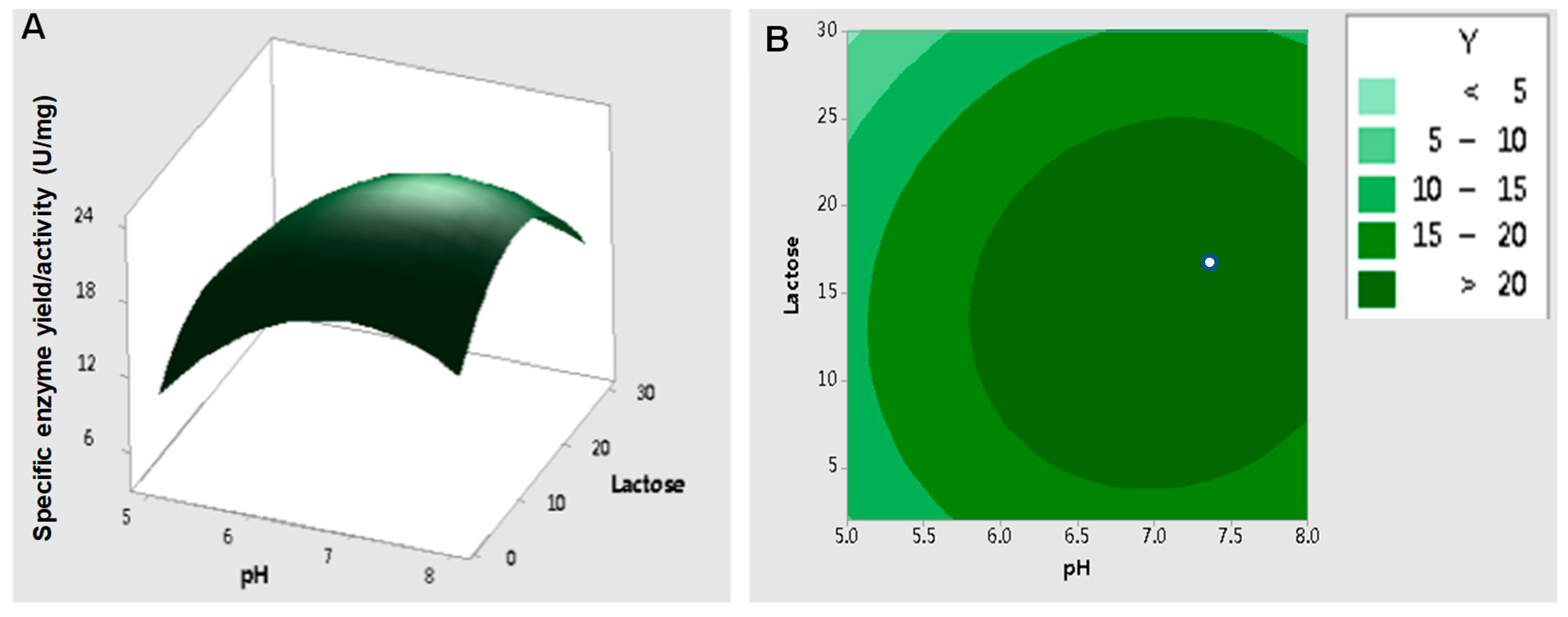
Central composite design (CCD) is one of the most frequently used approaches for RSM design employed to determine the key factors and interactive components in a system [
14]. In this case, it was critical to set the center point of the variables to make sure that no convergence appeared, since the converged values may have been out of range [
58]. Factor values of pH 6.5 and 15 g/L lactose were set as the center points based on the results of Plackett–Burman analysis. These values were assumed to be close to the optimum point for enzyme activity for RSM analysis. A three-dimensional response surface plot and contour plot were used to display the effects of pH and lactose concentration on the specific β-galactosidase activity (
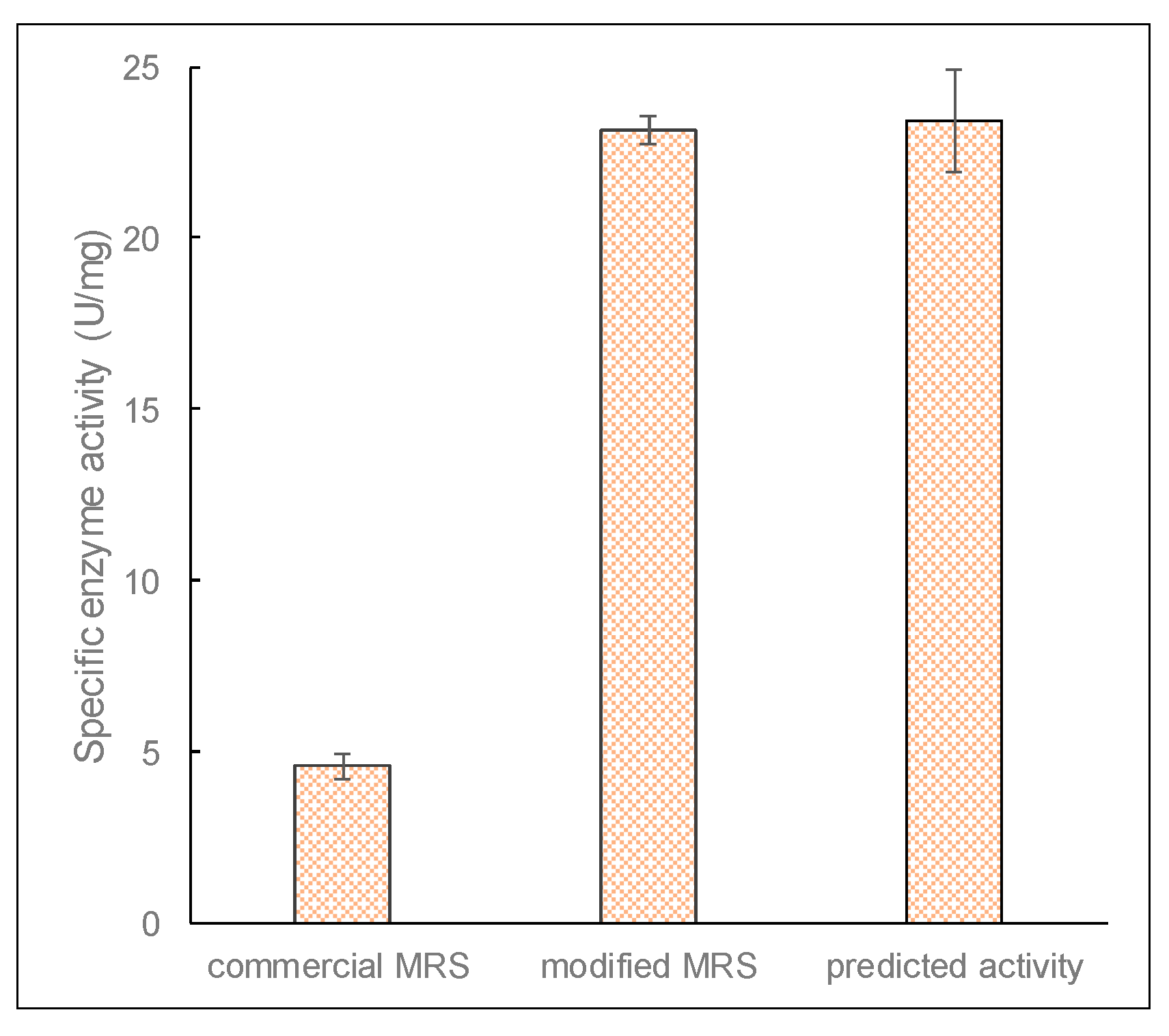
Figure 8). The predicted optimum pH and lactose concentration were 7.06 and 15.3 g/L, respectively, and the corresponding maximum specific enzyme activity of β-galactosidase was 23.4 ± 1.49 U/mg.
Table 8 shows the specific enzyme activity and model predictions for the central composite design for each experimental run. Moreover, a regression analysis of data produced the following quadratic polynomial equation (Equation (4)):
where
Y is the specific enzyme activity (predicted) of β-galactosidase in U/mg, and pH and lactose are the values of the pH and concentration of lactose, respectively. As shown in
Table 9, the coefficient of determination (
R2) was 0.98 when the goodness of fit of the model was tested. ANOVA for the CCD showed a very high fitting of experimental data to the model (
R2 = 0.99).













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}