Enhanced Production and Quantitative Evaluation of Nigericin from the Algerian Soil-Living Streptomyces youssoufiensis SF10 Strain

 and
and 
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Isolation and Identification of the Bacterial Strain
2.2. Crude Extracts
2.3. Culture Conditions of the Bacterial Strain Used to Maximize Nigericin Production
2.3.1. Selection of the Fermentation Process
2.3.2. Carbon Sources, pH Values, Presence of FeSO4 and Co-Culture Conditions
2.4. Calibration Curve Construction, Evaluation of the Detection Limit and Quantitative Analysis of Nigericin Produced under Different Culture Conditions
2.5. Phenylacetic Acid from Fermentation in Streptomyces sp. co-Culture
2.5.1. NMR and Mass Spectrometric Analysis
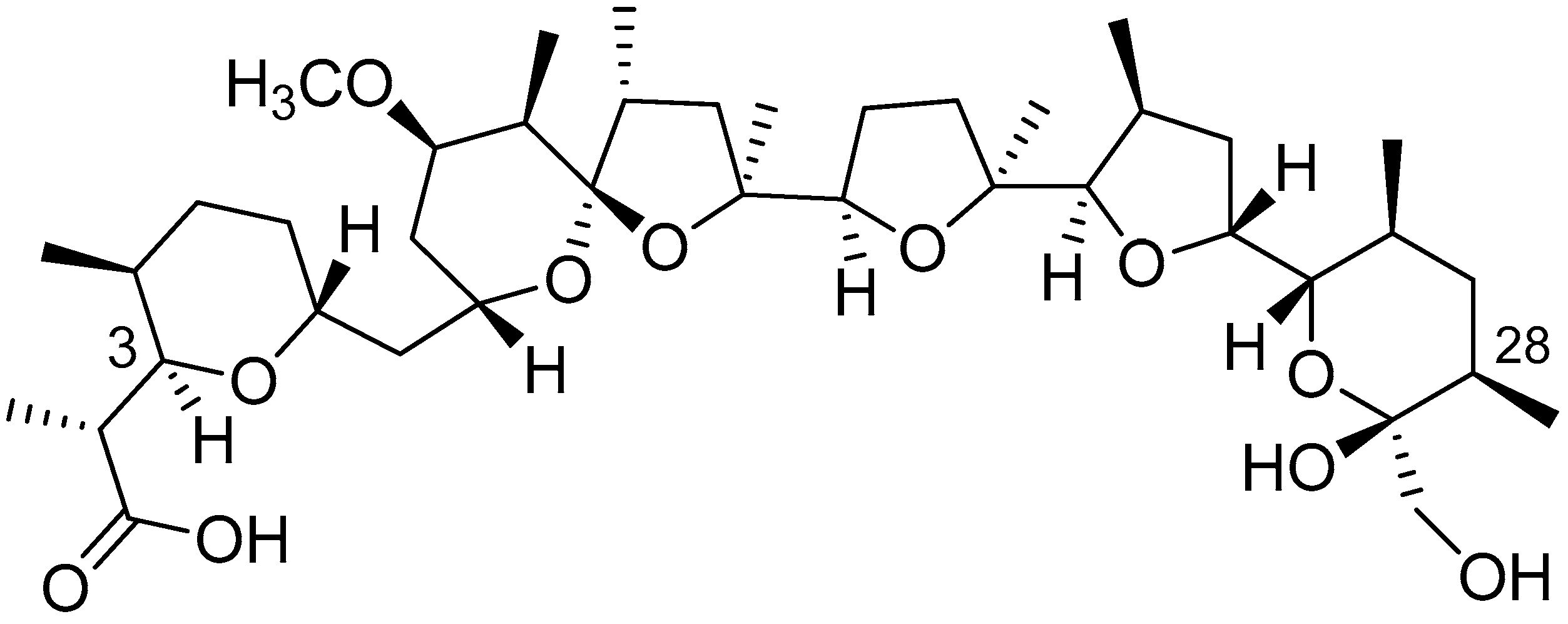
2.5.2. Isolation and Structural Characterization of the Metabolite
3. Results and Discussion
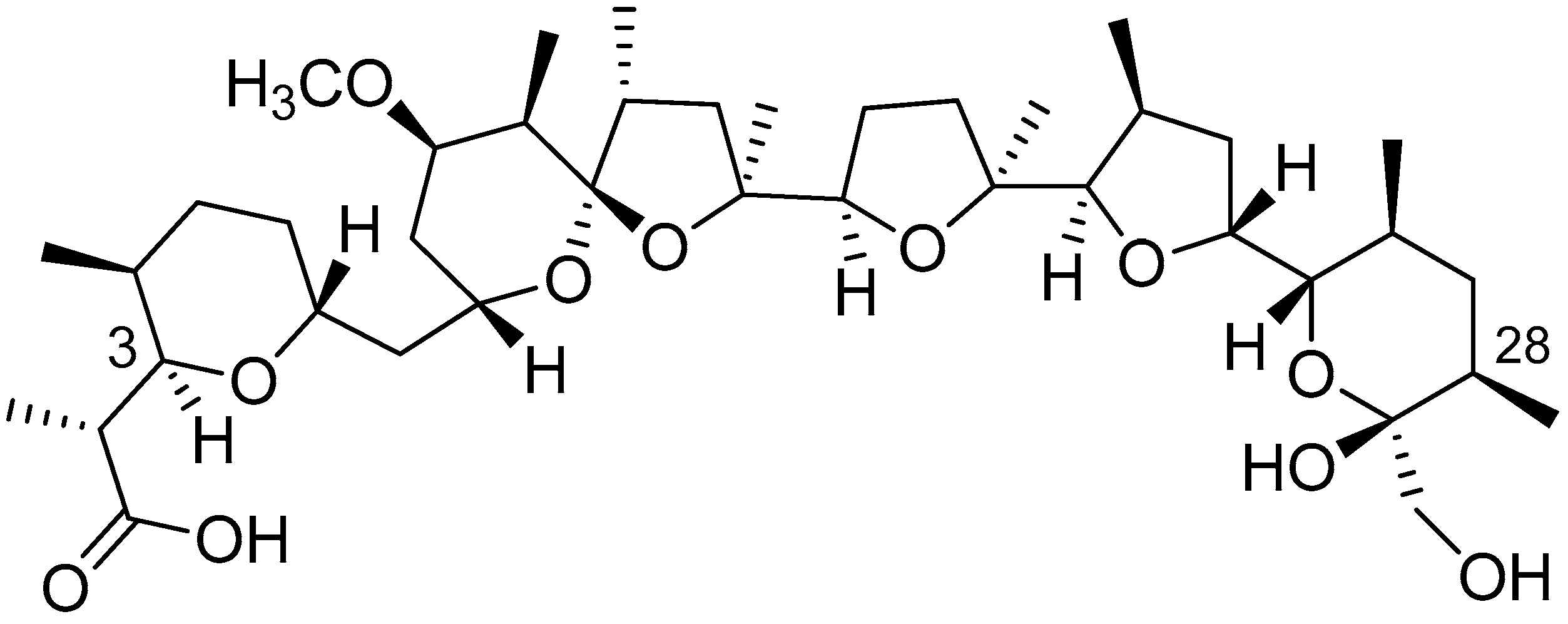
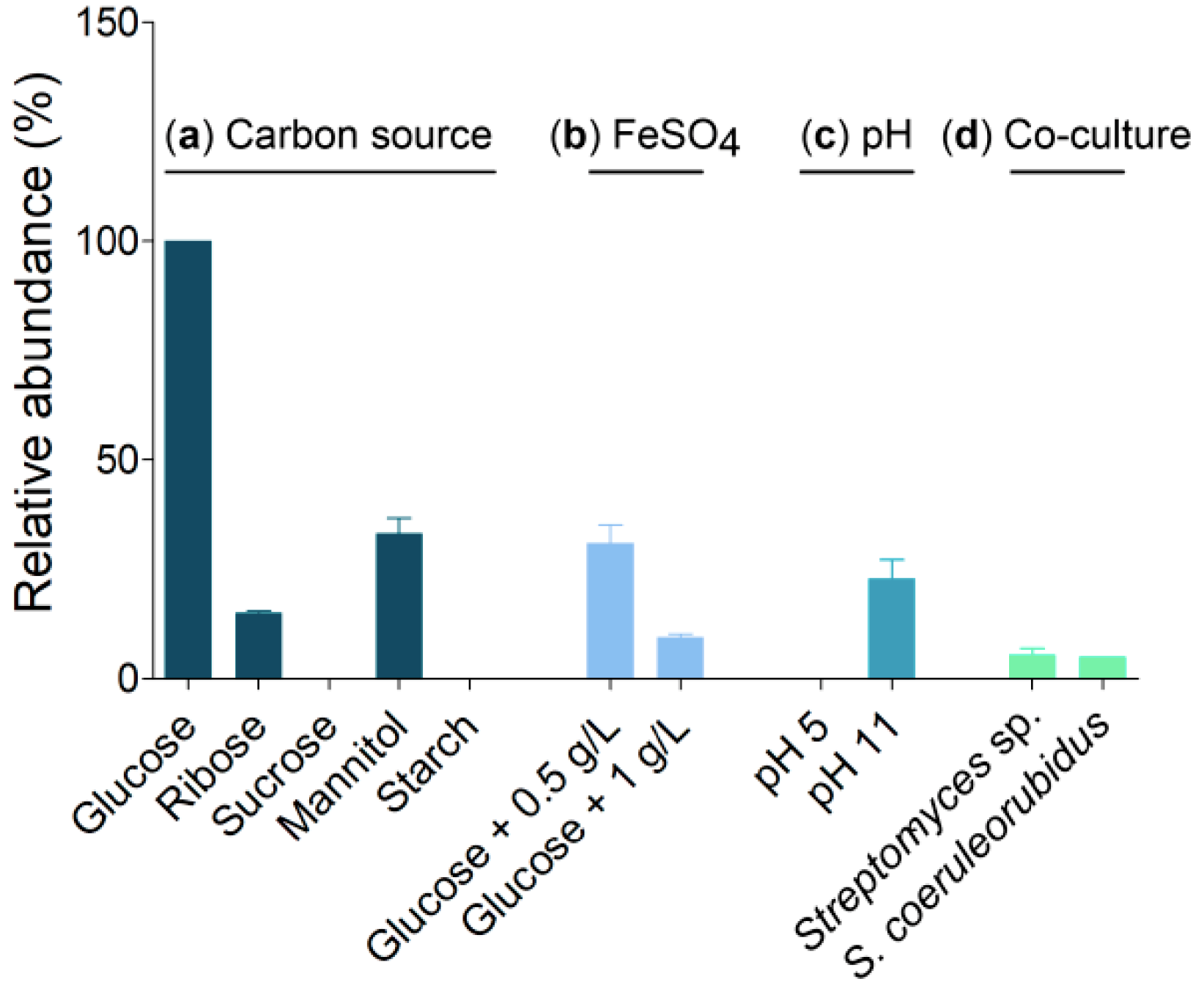
3.1. Metabolite Production under Different Culture Conditions
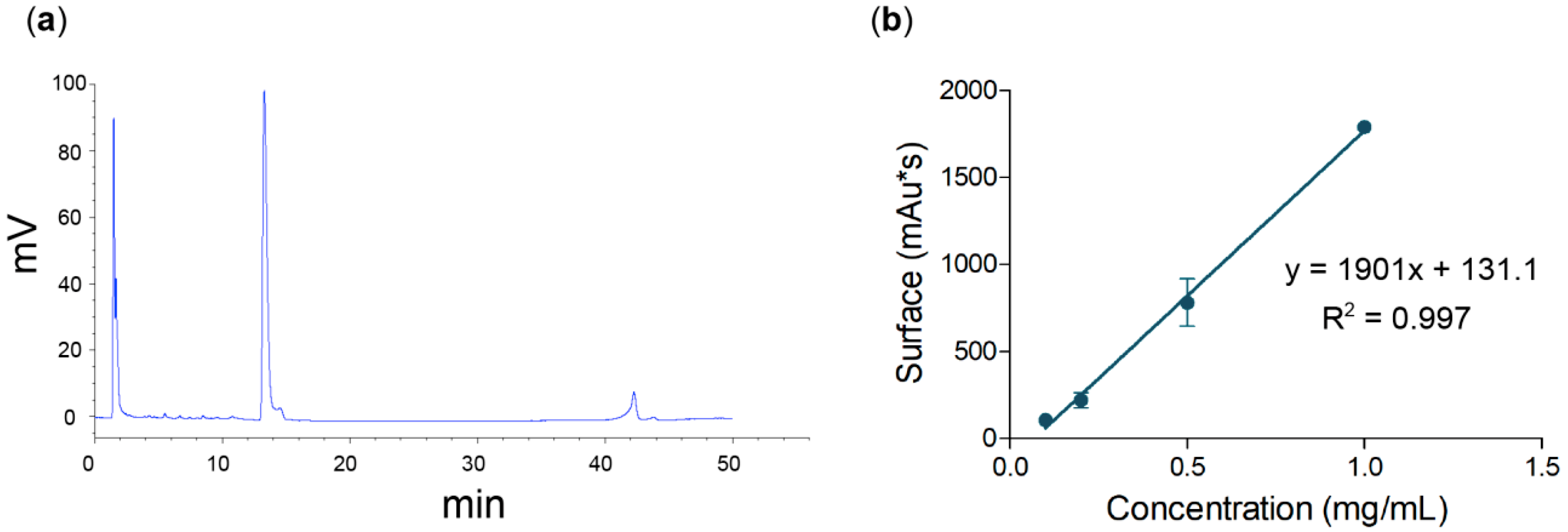
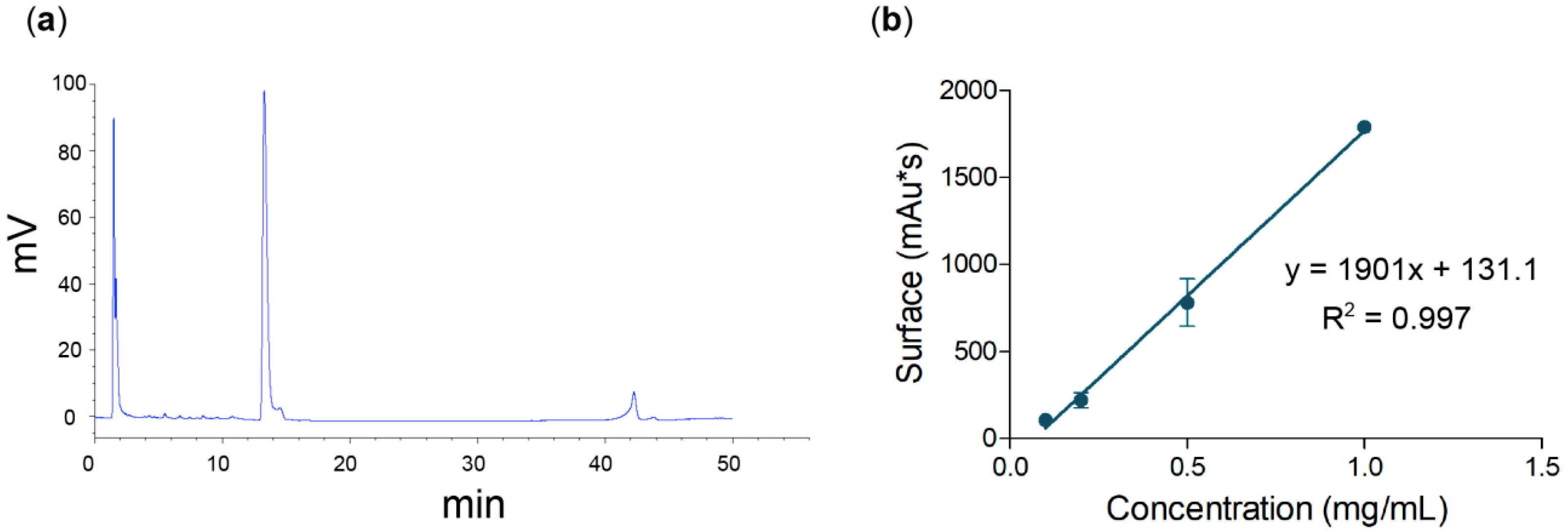
3.2. Quantitative Method for Nigericin Evaluation
3.3. Comparative Analysis of Nigericin Production
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Mohammadipanah, F.; Wink, J. Actinobacteria from arid and desert habitats: Diversity and biological activity. Front. Microbiol. 2016, 6, 1541. [Google Scholar] [CrossRef] [PubMed]
- Bamdad, M.; David, L.; Groliére, C.A. Epinigericin toxicity towards Tetrahymena pyriformis GL; changes in cell volume and intracellular pH. Appl. Microbiol. Biotechnol. 1995, 44, 206–209. [Google Scholar] [CrossRef] [PubMed]
- Smith, L.; Hong, H.; Spencer, J.B.; Leadaly, P.F. Analysis of specific mutants in the lasalocid gene cluster: Evidence for enzymatic catalysis of a disfavoured polyether ring closure. ChemBioChem 2008, 9, 2967–2975. [Google Scholar] [CrossRef] [PubMed]
- Elibol, M.; Mavituna, F. Effect of sucrose on actinorhodin production by Streptomyces coelicolor A3(2). Proc. Biochem. 1998, 33, 307–311. [Google Scholar] [CrossRef]
- Djinni, I.; Defant, A.; Kecha, M.; Mancini, I. Metabolite profile of marine-derived endophytic Streptomyces sundarbansensis WR1L1S8 by liquid chromatography–mass spectrometry and evaluation of culture conditions on antibacterial activity and mycelial growth. J. Appl. Microbiol. 2013, 116, 39–50. [Google Scholar] [CrossRef]
- Jounsbu, E.; McIntyre, M.; Nielsen, J. The influence of carbon sources and morphology on nystatin production by Streptomyces noursei. J. Biotechnol. 2002, 95, 133–144. [Google Scholar] [CrossRef]
- Dashti, Y.; Grkovic, T.; Abdelmohsen, U.R.; Hentschel, U.J.; Quinn, R. Production of induced secondary metabolites by a co-culture of sponge-associated Actinomycetes, Actinokineospora sp. EG49 and Nocardiopsis sp. RV163. Mar. Drugs 2014, 12, 3046–3059. [Google Scholar] [CrossRef]
- Elibol, M. Optimization of medium composition for actinorhodin production by Streptomyces coelicolor A3(2) with response surface methodology. Process. Biochem. 2004, 39, 1057–1062. [Google Scholar] [CrossRef]
- Singh, V.; Haque, S.; Niwas, R.; Srivastava, A.; Pasupuleti, M.; Tripathi, C.K. Strategies for fermentation medium optimization: An in-depth review. Front. Microbiol. 2017, 7, 2087. [Google Scholar] [CrossRef]
- Subramaniyam, R.; Vimala, R. Solid state and submerged fermentation for the production of bioactive substances: A comparative study. Int. J. Sci. Nat. 2012, 3, 480–486. [Google Scholar]
- Hölker, U.; Lenz, J. Solid-state fermentation—Are there any biotechnological advantages? Curr. Opin. Microbiol. 2005, 8, 301–306. [Google Scholar] [CrossRef] [PubMed]
- Rueda, B.; Miguelez, E.M.; Hardisson, C.; Manzanal, M.B. Mycelial differentiation and spore formation by Streptomyces brasiliensis in submerged culture. Can. J. Microbiol. 2001, 47, 1042–1047. [Google Scholar] [CrossRef] [PubMed]
- Shu, C.H.; Lung, M.Y. Effect of the pH on the production and molecular weight distribution of exopolysaccharide by antrodia camphorate in batch cultures. Process Biochem. 2004, 39, 931–937. [Google Scholar] [CrossRef]
- Harvey, M.B.; Mironenko, T.; Sun, Y.; Hong, H.; Deng, Z.; Leadlay, F.P.; Weissman, J.K.; Haydock, S.F. Insights into polyether biosynthesis from analysis of the nigericin biosynthetic gene cluster in Streptomyces sp. DSM 4137. Chem. Biol. 2007, 14, 703–714. [Google Scholar] [CrossRef] [PubMed]
- Taechowisan, T.; Chanaphat, S.; Ruensamran, W.; Phutdhawong, W.S. Antibacterial activity of 1-methyl ester-nigericin from Streptomycxes hygroscopicus BR10: An endophyte in Alpinia galanga. J. Appl. Pharm. Sci. 2013, 3, 104–109. [Google Scholar] [CrossRef]
- Gumila, C.; Ancelin, M.L.; Delort, A.M.; Jeminet, G.; Vialj, H. Characterization of the potent in vitro and in vivo antimalarial activities of ionophore compounds. Antimicrob. Agents Chemother. 1997, 41, 523–529. [Google Scholar] [CrossRef] [PubMed]
- Bilen, S.; Biswas, G.; Otsuyama, S.; Kono, T.; Sakai, M.; Hikima, J.I. Inflammatory responses in the Japanese pufferfish (Takifugu rubripes) head kidney cells stimulated with an inflammasome-inducing agent, nigericin. Dev. Comp. Immunol. 2014, 46, 222–230. [Google Scholar] [CrossRef]
- Kapoor, A.; He, R.; Venkatadri, R.; Forman, M.; Arav-Boger, R. Wnt modulating agents inhibit human cytomegalovirus replication. Antimicrob. Agents Chemother. 2013, 57, 2761–2767. [Google Scholar] [CrossRef]
- Zhou, H.M.; Dong, T.T.; Feng, L.L.; Feng, B.; Zhao, H.C.; Fan, X.K.; Zheng, M.H. Suppression of colorectal cancer metatstasis by nigericin through inihition of epithelial-mesenchymal transition. World J. Gastroenterol. 2012, 18, 2640–2648. [Google Scholar] [CrossRef]
- Deng, C.C.; Liang, Y.; Wu, M.S.; Feng, F.T.; Hu, W.R.; Chen, L.Z.; Feng, Q.S.; Bei, J.X.; Zeng, Y.X. Nigericin selectively targets cancer stem cells in nasopharyngeal carcinoma. Int. J. Biochem. Cell Biol. 2013, 45, 1997–2006. [Google Scholar] [CrossRef]
- Hegazy, A.M.; Yamada, D.; Kobayashi, M.; Kohno, S.; Ueno, M.; Ali, M.A.E.; Ohta, K.; Tadokoro, Y.; Ino, Y.; Todo, T.; et al. Therapeutic strategy for targeting aggressive malignant gliomas by disrupting their energy balance. J. Biol. Chem. 2016, 291, 21496–21509. [Google Scholar] [CrossRef] [PubMed]
- Yakisich, J.S.; Azad, N.; Kaushik, V.; O’Doherty, G.A.; Iyer, A.K.V. Nigericin decreases viability of multidrug-resistant cancer cells and lung tumorspheres and potentiates the effects of cardiac glycosides. Tumor Biol. 2017, 39. [Google Scholar] [CrossRef] [PubMed]
- Leulmi, N.; Sighel, D.; Defant, A.; Khenaka, K.; Boulahrouf, A.; Mancini, I. Nigericin and grisorixin methyl ester from the Algerian soil-living Streptomyces youssoufiensis SF10 strain: A computational study on their epimeric structures and evaluation of glioblastoma stem cells growth inhibition. Nat. Prod. Res. 2018, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Manteca, A.; Alvarez, R.; Salazar, N.; Yague, P.; Sanchez, J. Mycelium differentiation and antibiotic production in submerged culture of Streptomyces celicolor. Appl. Environ. Microbiol. 2008, 74, 3877–3886. [Google Scholar] [CrossRef] [PubMed]
- Hwang, B.K.; Lim, S.W.; Kim, B.S.; Lee, J.Y. Isolation and in vivo and in vitro antifungal activity of phenylacetic acid and sodium phenylacetate from Streptomyces humidus. Appl. Environ. Microbiol. 2001, 67, 3739–3745. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.; Cho, J.Y.; Kuk, J.H.; Moon, J.H.; Cho, J.; Kim, Y.C.; Park, K.H. Identification and antimicrobial activity of phenylacetic produced by Bacillus licheniformis isolated from fermented soybean, Chungkook-Jang. Curr. Microbiol. 2004, 48, 312–317. [Google Scholar] [CrossRef] [PubMed]
- Crevelin, E.J.; Crotti, A.E.M.; Zucchi, T.D.; Melo, I.S.; Moraes, L.A.B. Dereplication of Streptomyces sp. AMC 23 polyether ionophore antibiotics by accurate mass electrospray tandem mass spectrometry. J. Mass Spectrom. 2014, 49, 1117–1126. [Google Scholar] [CrossRef]
- Shahab, N.; Kamaruddin, K.; Platt, J.; Butler, P.R.; Olivier, S.G.; Hobbs, G. Cell physiology and antibiotic production of Streptomyces coelicolor grown on solid medium. Biotechnol. Lett. 1994, 16, 1015–1020. [Google Scholar] [CrossRef]
- Darakchieva, M.; Ratchev, R.; Sachanekova, M.; Tzvetkova, R.; Gameyska, Y.O.; Popov, K.; Roussev, P.; Schlegel, R.; Kleinwachter, W.; Thrum, H. Polyether antibiotic complex produced by Streptomyces hygroscopicus IM-110-81. Biotechnol. Bioind. 1987, 2, 15–17. [Google Scholar] [CrossRef]
- Zhou, J.; He, H.; Wang, X.; Lu, J.; Zhou, X.; Cai, M.; Zhang, Y. Optimization of nutrients for dinactin production by a marine Streptomyces sp. from the high latitude Arctic. Biotechnol. Bioprocess Eng. 2015, 20, 725–732. [Google Scholar] [CrossRef]
- Gesheva, V.; Ivanova, V.; Gesheva, R. Effects of nutrients on the production of AK-111-81 macrolide antibiotic by Streptomyces hygroscopicus. Microbiol. Res. 2005, 160, 243–248. [Google Scholar] [CrossRef] [PubMed]
- Dauphin, G.; David, L.; Kergomard, A.; Sainz, C. Nigericic acids, two new chemical derivatives of nigericin. Agric. Biol. Chem. 1989, 53, 2497–2498. [Google Scholar] [CrossRef]
- Harned, R.L.; Hidy, P.H.; Corum, C.J.; Jones, K.L. Nigericin a new crystalline antibiotic from an unidentified Streptomyces. Antibiot. Chemother. 1951, 1, 594–596. [Google Scholar]
- David, L.; Leal Ayala, H.; Tabet, J.C. Abierixin, a new polyether antibiotic. Production, structural determination and biological activities. J. Antibiot. 1985, 31, 1655–1663. [Google Scholar] [CrossRef]
- Xiao, L.; Huang, W.Y. Improvement of fermentation conditions for nigericin produced by Streptomyces hygroscopicus NND-52. Zhongguo Yiyao Gongye Zazhi (Chin. J. Pharm.) 2002, 33, 375–377. [Google Scholar]
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Leulmi, N.; Sighel, D.; Defant, A.; Khenaka, K.; Boulahrouf, A.; Mancini, I. Enhanced Production and Quantitative Evaluation of Nigericin from the Algerian Soil-Living Streptomyces youssoufiensis SF10 Strain. Fermentation 2019, 5, 13. https://doi.org/10.3390/fermentation5010013
Leulmi N, Sighel D, Defant A, Khenaka K, Boulahrouf A, Mancini I. Enhanced Production and Quantitative Evaluation of Nigericin from the Algerian Soil-Living Streptomyces youssoufiensis SF10 Strain. Fermentation. 2019; 5(1):13. https://doi.org/10.3390/fermentation5010013
Chicago/Turabian StyleLeulmi, Nassima, Denise Sighel, Andrea Defant, Karima Khenaka, Abderrahmane Boulahrouf, and Ines Mancini. 2019. "Enhanced Production and Quantitative Evaluation of Nigericin from the Algerian Soil-Living Streptomyces youssoufiensis SF10 Strain" Fermentation 5, no. 1: 13. https://doi.org/10.3390/fermentation5010013
APA StyleLeulmi, N., Sighel, D., Defant, A., Khenaka, K., Boulahrouf, A., & Mancini, I. (2019). Enhanced Production and Quantitative Evaluation of Nigericin from the Algerian Soil-Living Streptomyces youssoufiensis SF10 Strain. Fermentation, 5(1), 13. https://doi.org/10.3390/fermentation5010013