
Novel Malolactic Fermentation Starter Formulated Using Native Lactic Acid Bacteria Strains from a Re-Emerging Wine-Growing Region of Argentina—A Pilot Scale Vinification

 ,
,  , ,
, ,
Abstract
1. Introduction
2. Materials and Methods
2.1. LAB Isolation
2.2. Identification and Typing of Strains
2.3. Pilot-Scale Vinification
2.3.1. Preparation of Inocula for Malolactic Fermentation
2.3.2. Fermentation Process
2.3.3. LAB Viability Control
2.3.4. Implantation Capacity of the Inoculated Cultures
2.3.5. Evaluation of L-Malic and Acetic Acids During Malolactic Fermentation (MLF)
2.3.6. Analysis of Aromatic Profiles
2.4. Statistical Analysis
3. Results and Discussion
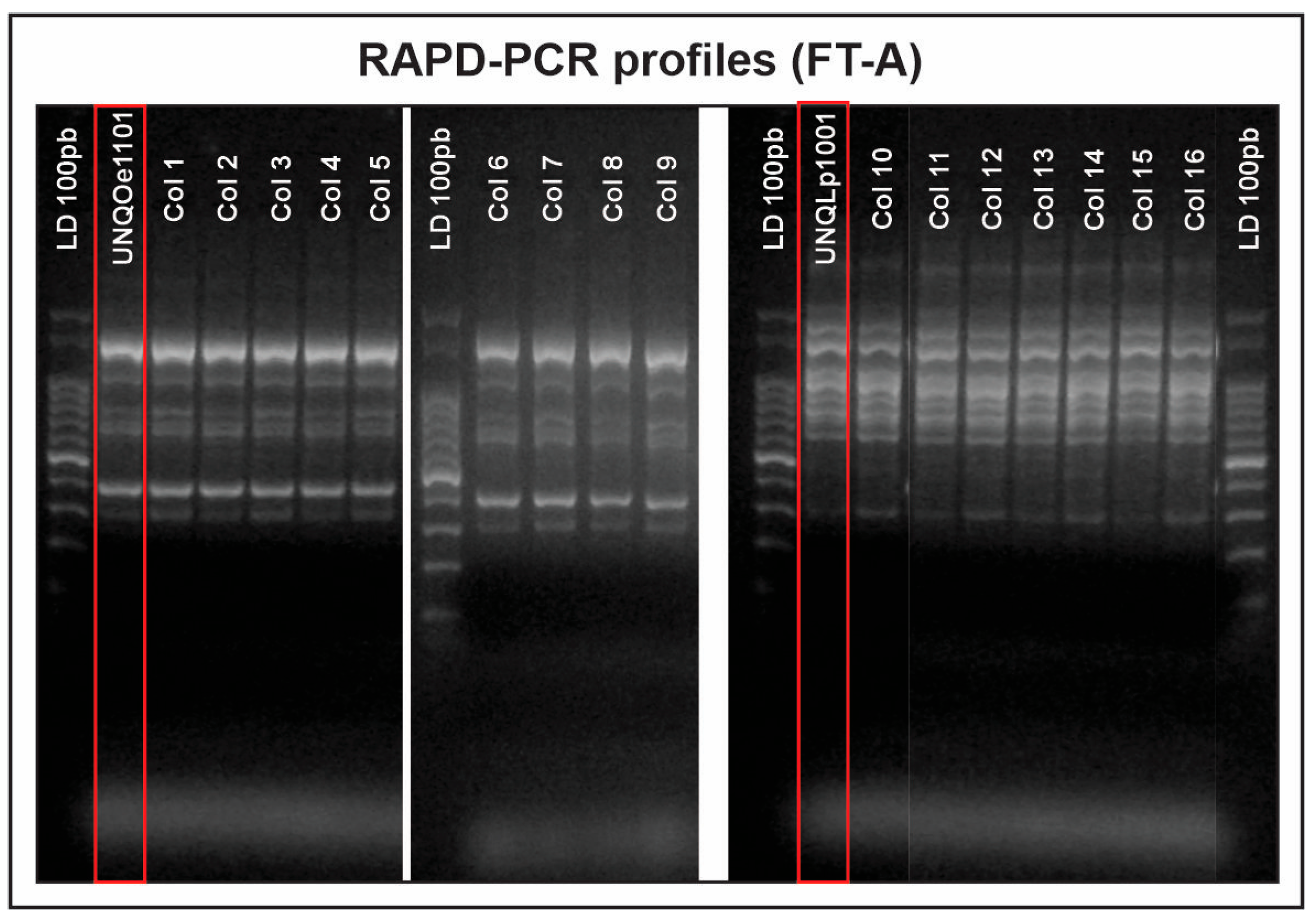
3.1. Isolation, Identification and Typing of Indigenous Strains
3.2. Pilot-Scale Winemaking
3.2.1. Viability Control and Implantation of LAB
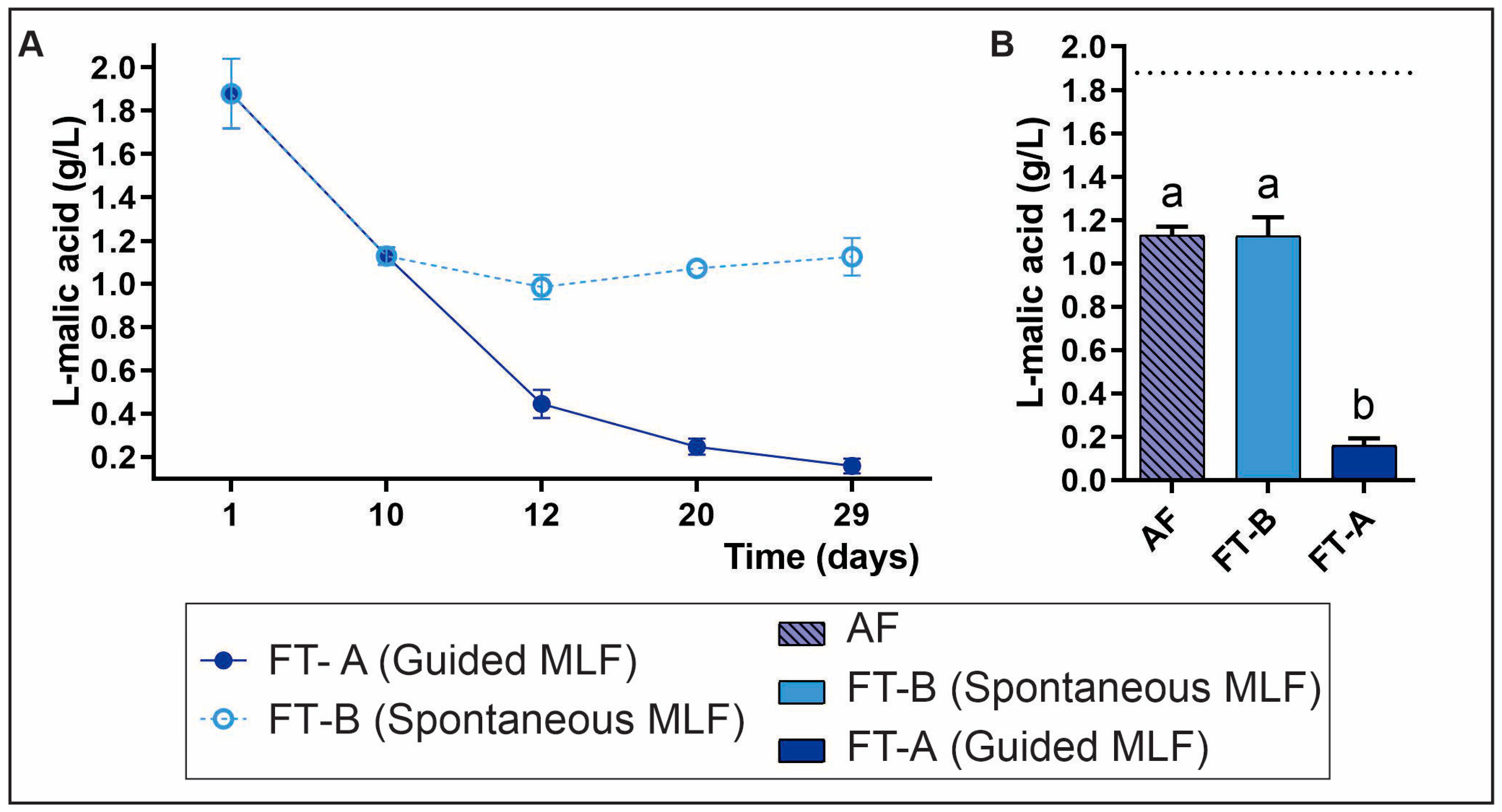
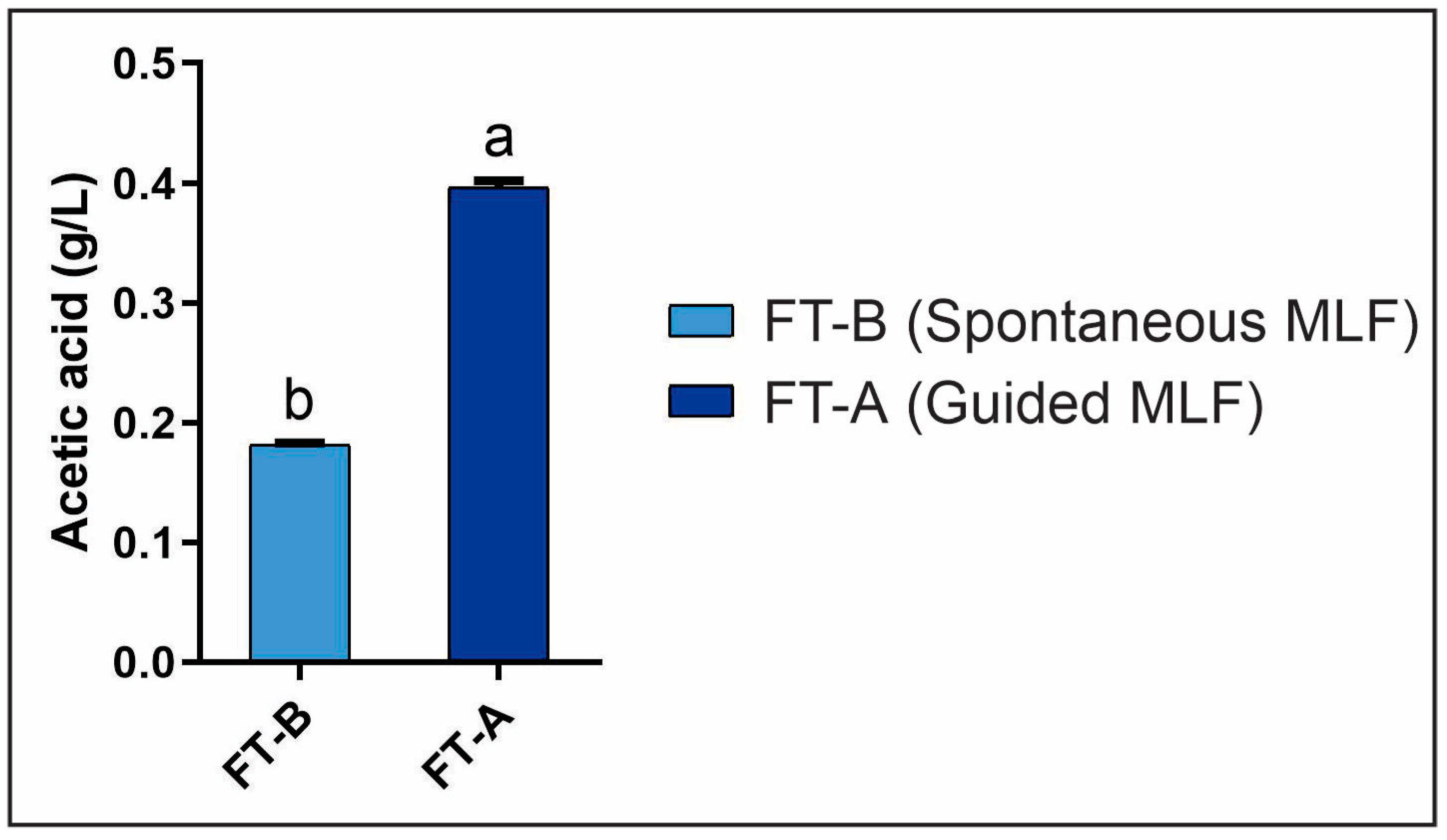
3.2.2. Evaluation of L-Malic and Acetic Acids During MLF
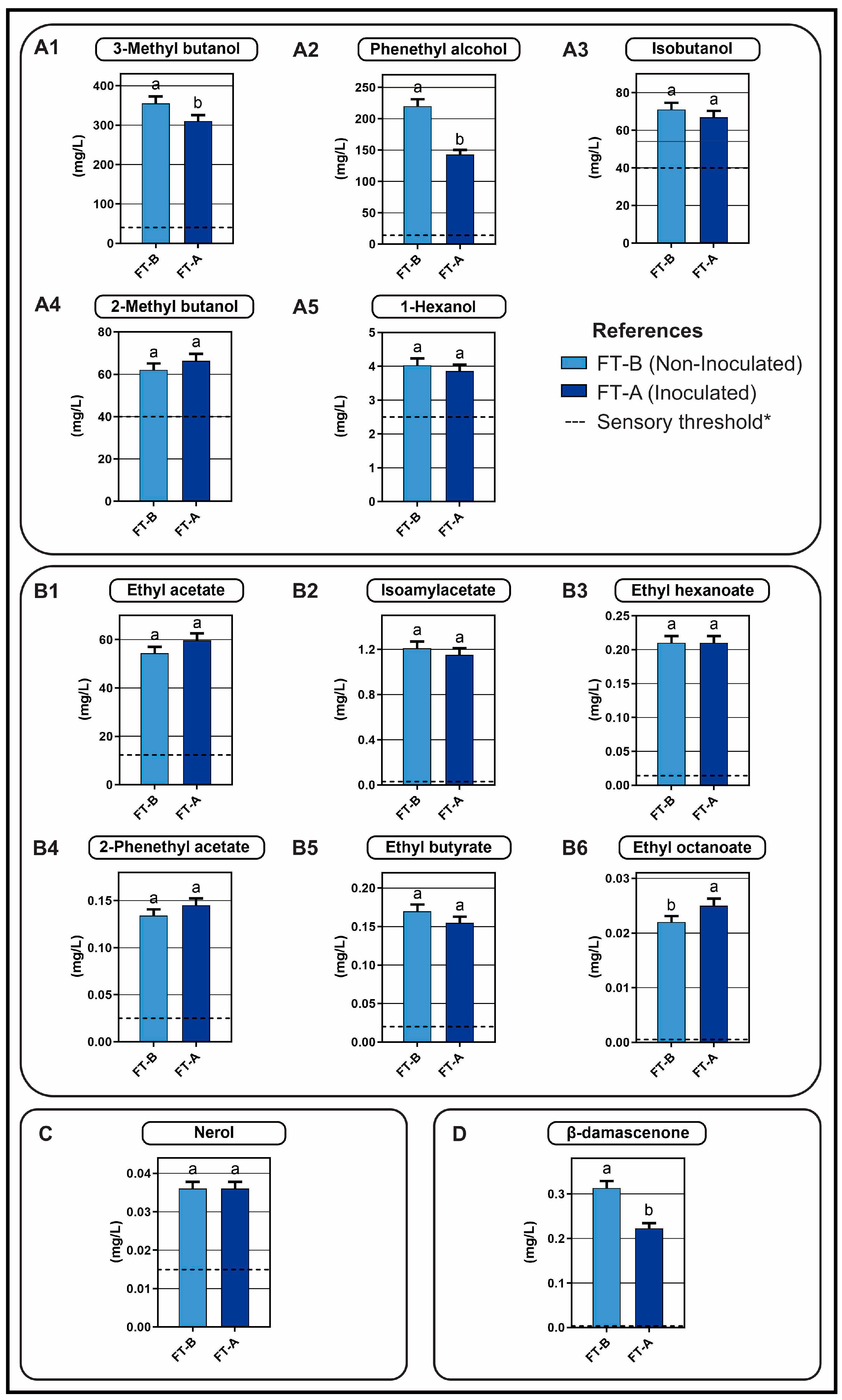
3.2.3. Analysis of the Aromatic Profiles of the Wines Obtained
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| AAB | Acetic acid bacteria |
| AF | Alcoholic fermentation |
| FT | Fermentation tank |
| GC-MS | Gas Chromatography and Mass Spectrometry |
| LAB | Lactic acid bacteria |
| MLF | Malolactic fermentation |
| MLFS | Malolactic fermentation starter |
| NGS | Next-Generation Sequencing |
References
- Swiegers, J.H.; Pretorius, I.S. Yeast modulation of wine flavor. Adv. Appl. Microbiol. 2005, 57, 131–175. [Google Scholar] [CrossRef] [PubMed]
- Lerm, E.; Engelbrecht, L.; Du Toit, M. Selection and Characterisation of Oenococcus oeni and Lactobacillus plantarum South African Wine Isolates for Use as Malolactic Fermentation Starter Cultures; Stellenbosch University: Stellenbosch, South Africa, 2011. [Google Scholar] [CrossRef]
- Paramithiotis, S.; Stasinou, V.; Tzamourani, A.; Kotseridis, Y.; Dimopoulou, M. Malolactic fermentation—Theoretical advances and practical considerations. Fermentation 2022, 8, 521. [Google Scholar] [CrossRef]
- Renouf, V.; Vayssieres, L.C.; Claisse, O.; Lonvaud-Funel, A. Genetic and phenotypic evidence for two groups of Oenococcus oeni strains and their prevalence during winemaking. Appl. Microbiol. Biotechnol. 2009, 83, 85–97. [Google Scholar] [CrossRef] [PubMed]
- Manera, C.; Olguin, N.T.; Bravo-Ferrada, B.M.; Tymczyszyn, E.E.; Delfederico, L.; Bibiloni, H.; Caballero, A.C.; Semorile, L.; La Hens, D.V. Survival and implantation of indigenous psychrotrophic Oenococcus oeni strains during malolactic fermentation in a Patagonian Pinot noir wine. LWT 2019, 108, 353–360. [Google Scholar] [CrossRef]
- Marcobal, Á.; Martín-Álvarez, P.J.; Polo, M.C.; Muñoz, R.; Moreno-Arribas, M. Formation of biogenic amines throughout the industrial manufacture of red wine. J. Food Prot. 2006, 69, 397–404. [Google Scholar] [CrossRef]
- Mira de Orduna, R.; Liu, S.Q.; Patchett, M.; Pilone, G. Kinetics of the arginine metabolism of malolactic wine lactic acid bacteria Lactobacillus buchneri CUC-3 and Oenococcus oeni Lo111. J. Appl. Microbiol. 2000, 89, 547–552. [Google Scholar] [CrossRef]
- Virdis, C.; Sumby, K.; Bartowsky, E.; Jiranek, V. Lactic acid bacteria in wine: Technological advances and evaluation of their functional role. Front. Microbiol. 2021, 11, 612118. [Google Scholar] [CrossRef]
- Guilloux-Benatier, M.; Le Fur, Y.; Feuillat, M. Influence of fatty acids on the growth of wine microorganisms Saccharomyces cerevisiae and Oenococcus oeni. J. Ind. Microbiol. Biotechnol. 1998, 20, 144–149. [Google Scholar] [CrossRef]
- International Organisation of Vine and Wine (OIV). State of the World Vitivinicultural Sector in 2019; OIV: Dijon, France, 2021. [Google Scholar]
- Rivas, G.A.; Guillade, A.C.; Semorile, L.C.; Delfederico, L. Influence of climate on soil and wine bacterial diversity on a vineyard in a non-traditional wine region in Argentina. Front. Microbiol. 2021, 12, 726384. [Google Scholar] [CrossRef]
- Rivas, G.A.; Semorile, L.; Delfederico, L. Microbial diversity of the soil, rhizosphere and wine from an emerging wine-producing region of Argentina. LWT 2022, 153, 112429. [Google Scholar] [CrossRef]
- Maicas, S.; González-Cabo, P.; Ferrer, S.; Pardo, I. Production of Oenococcus oeni biomass to induce malolactic fermentation in wine by control of pH and substrate addition. Biotechnol. Lett. 1999, 21, 349–353. [Google Scholar] [CrossRef]
- Bravo-Ferrada, B.M.; Hollmann, A.; Delfederico, L.; Valdés La Hens, D.; Caballero, A.; Semorile, L. Patagonian red wines: Selection of Lactobacillus plantarum isolates as potential starter cultures for malolactic fermentation. World J. Microbiol. Biotechnol. 2013, 29, 1537–1549. [Google Scholar] [CrossRef] [PubMed]
- Stenlid, J.; Karlsson, J.-O.; Högberg, N. Intraspecific genetic variation in Heterobasidion annosum revealed by amplification of minisatellite DNA. Mycol. Res. 1994, 98, 57–63. [Google Scholar] [CrossRef]
- Rodas, A.M.; Ferrer, S.; Pardo, I. 16S-ARDRA, a tool for identification of lactic acid bacteria isolated from grape must and wine. Syst. Appl. Microbiol. 2003, 26, 412–422. [Google Scholar] [CrossRef] [PubMed]
- Bravo-Ferrada, B.M.; Tymczyszyn, E.E.; Gómez-Zavaglia, A.; Semorile, L. Effect of acclimation medium on cell viability, membrane integrity and ability to consume malic acid in synthetic wine by oenological Lactobacillus plantarum strains. J. Appl. Microbiol. 2014, 116, 360–367. [Google Scholar] [CrossRef]
- Corral-Lugo, A.; Morales-García, Y.E.; Pazos-Rojas, L.A.; Ramírez-Valverde, A.; Martínez-Contreras, R.D.; Muñoz-Rojas, J. Cuantificación de bacterias cultivables mediante el método de” Goteo en Placa por Sellado (o estampado) Masivo”. Ann. Microbiol. 2012, 14, 147–156. [Google Scholar]
- Aymerich, T.; Martin, B.; Garriga, M.; Vidal-Carou, M.; Bover-Cid, S.; Hugas, M. Safety properties and molecular strain typing of lactic acid bacteria from slightly fermented sausages. J. Appl. Microbiol. 2006, 100, 40–49. [Google Scholar] [CrossRef]
- Brizuela, N.S.; Bravo-Ferrada, B.M.; Pozo-Bayón, M.Á.; Semorile, L.; Tymczyszyn, E.E. Changes in the volatile profile of Pinot noir wines caused by Patagonian Lactobacillus plantarum and Oenococcus oeni strains. Food Res. Int. 2018, 106, 22–28. [Google Scholar] [CrossRef]
- Bartowsky, E.J.; Borneman, A.R. Genomic variations of Oenococcus oeni strains and the potential to impact on malolactic fermentation and aroma compounds in wine. Appl. Microbiol. Biotechnol. 2011, 92, 441–447. [Google Scholar] [CrossRef]
- Petruzzi, L.; Capozzi, V.; Berbegal, C.; Corbo, M.R.; Bevilacqua, A.; Spano, G.; Sinigaglia, M. Microbial resources and enological significance: Opportunities and benefits. Front. Microbiol. 2017, 8, 995. [Google Scholar] [CrossRef]
- Brizuela, N.S.; Franco-Luesma, E.; Bravo-Ferrada, B.M.; Pérez-Jiménez, M.; Semorile, L.; Tymczyszyn, E.E.; Pozo-Bayon, M. Influence of Patagonian Lactiplantibacillus plantarum and Oenococcus oeni strains on sensory perception of Pinot Noir wine after malolactic fermentation. Aust. J. Grape Wine Res. 2021, 27, 118–127. [Google Scholar] [CrossRef]
- Diez-Ozaeta, I.; Lavilla, M.; Amárita, F. Technological characterisation of potential malolactic starters from Rioja Alavesa winemaking region. LWT 2020, 134, 109916. [Google Scholar] [CrossRef]
- Capozzi, V.; Di Toro, M.R.; Grieco, F.; Michelotti, V.; Salma, M.; Lamontanara, A.; Russo, P.; Orrù, L.; Alexandre, H.; Spano, G. Viable But Not Culturable (VBNC) state of Brettanomyces bruxellensis in wine: New insights on molecular basis of VBNC behaviour using a transcriptomic approach. Food Microbiol. 2016, 59, 196–204. [Google Scholar] [CrossRef] [PubMed]
- Kioroglou, D. Analysis of Microbial Populations in Wines Through NGS Methodologies. Ph.D. Thesis, Universitat Rovira i Virgili, Tarragona, Spain, 2020. [Google Scholar]
- Millet, V.; Lonvaud-Funel, A. The viable but non-culturable state of wine micro-organisms during storage. Lett. Appl. Microbiol. 2000, 30, 136–141. [Google Scholar] [CrossRef]
- Bravo-Ferrada, B.M.; Brizuela, N.; Gerbino, E.; Gómez-Zavaglia, A.; Semorile, L.; Tymczyszyn, E.E. Effect of protective agents and previous acclimation on ethanol resistance of frozen and freeze-dried Lactobacillus plantarum strains. Cryobiology 2015, 71, 522–528. [Google Scholar] [CrossRef]
- Brizuela, N.S.; Bravo-Ferrada, B.M.; La Hens, D.V.; Hollmann, A.; Delfederico, L.; Caballero, A.; Tymczyszyn, E.E.; Semorile, L. Comparative vinification assays with selected Patagonian strains of Oenococcus oeni and Lactobacillus plantarum. LWT 2017, 77, 348–355. [Google Scholar] [CrossRef]
- Chu-Ky, S.; Tourdot-Marechal, R.; Marechal, P.-A.; Guzzo, J. Combined cold, acid, ethanol shocks in Oenococcus oeni: Effects on membrane fluidity and cell viability. Biochim. Biophys. Acta (BBA) Biomembr. 2005, 1717, 118–124. [Google Scholar] [CrossRef]
- Da Silveira, M.G.; Golovina, E.A.; Hoekstra, F.A.; Rombouts, F.M.; Abee, T. Membrane fluidity adjustments in ethanol-stressed Oenococcus oeni cells. Appl. Environ. Microbiol. 2003, 69, 5826–5832. [Google Scholar] [CrossRef]
- van Wyk, N.; Scansani, S.; Beisert, B.; Brezina, S.; Fritsch, S.; Semmler, H.; Pretorius, I.S.; Rauhut, D.; von Wallbrunn, C. The use of Hanseniaspora occidentalis in a sequential must inoculation to reduce the malic acid content of wine. Appl. Sci. 2022, 12, 6919. [Google Scholar] [CrossRef]
- Vicente, J.; Kelanne, N.; Navascués, E.; Calderón, F.; Santos, A.; Marquina, D.; Yang, B.; Benito, S. Combined use of Schizosaccharomyces pombe and a Lachancea thermotolerans strain with a high malic acid consumption ability for wine production. Fermentation 2023, 9, 165. [Google Scholar] [CrossRef]
- Bravo-Ferrada, B.M.; Hollmann, A.; Brizuela, N.; La Hens, D.V.; Tymczyszyn, E.; Semorile, L. Growth and consumption of L-malic acid in wine-like medium by acclimated and non-acclimated cultures of Patagonian Oenococcus oeni strains. Folia Microbiol. 2016, 61, 365–373. [Google Scholar] [CrossRef] [PubMed]
- Comitini, F.; Ciani, M. The inhibitory activity of wine yeast starters on malolactic bacteria. Ann. Microbiol. 2007, 57, 61–66. [Google Scholar] [CrossRef]
- Instituto Nacional de Vitivinicultura (INV); Secretaría de Agricultura; Ganadería y Pesca; Ministerio de Economía. Legislación y Avisos Oficiales; RESOL-2018-2-APN-INV, MPYT12; INV: Mendoza, Argentina, 2021. [Google Scholar]
- Cibrario, A.; Peanne, C.; Lailheugue, M.; Campbell-Sills, H.; Dols-Lafargue, M. Carbohydrate metabolism in Oenococcus oeni: A genomic insight. BMC Genom. 2016, 17, 984. [Google Scholar] [CrossRef]
- du Toit, M.; Engelbrecht, L.; Lerm, E.; Krieger-Weber, S. Lactobacillus: The next generation of malolactic fermentation starter cultures—An overview. Food Bioprocess Technol. 2011, 4, 876–906. [Google Scholar] [CrossRef]
- Pretorius, N. Evaluation of Citrate Metabolism in Oenococcus oeni and Lactobacillus plantarum. Ph.D. Thesis, Stellenbosch University, Stellenbosch, South Africa, 2016. [Google Scholar]
- Peinado, R.A.; Moreno, J.; Bueno, J.E.; Moreno, J.A.; Mauricio, J.C. Comparative study of aromatic compounds in two young white wines subjected to pre-fermentative cryomaceration. Food Chem. 2004, 84, 585–590. [Google Scholar] [CrossRef]
- Baron, M.; Prusova, B.; Tomaskova, L.; Kumsta, M.; Sochor, J. Terpene content of wine from the aromatic grape variety ‘Irsai Oliver’ (Vitis vinifera L.) depends on maceration time. Open Life Sci. 2017, 12, 42–50. [Google Scholar] [CrossRef]
- Fariña, L.; Villar, V.; Ares, G.; Carrau, F.; Dellacassa, E.; Boido, E. Volatile composition and aroma profile of Uruguayan Tannat wines. Food Res. Int. 2015, 69, 244–255. [Google Scholar] [CrossRef]
- Santos, J.P.; Lozano, J.; Aleixandre, M.; Arroyo, T.; Cabellos, J.M.; Gil, M.; del Carmen Horrillo, M. Threshold detection of aromatic compounds in wine with an electronic nose and a human sensory panel. Talanta 2010, 80, 1899–1906. [Google Scholar] [CrossRef]
- Synos, K.; Reynolds, A.; Bowen, A. Effect of yeast strain on aroma compounds in Cabernet franc icewines. LWT Food Sci. Technol. 2015, 64, 227–235. [Google Scholar] [CrossRef]
- Rodriguez-Campos, J.; Escalona-Buendía, H.; Orozco-Avila, I.; Lugo-Cervantes, E.; Jaramillo-Flores, M.E. Dynamics of volatile and non-volatile compounds in cocoa (Theobroma cacao L.) during fermentation and drying processes using principal components analysis. Food Res. Int. 2011, 44, 250–258. [Google Scholar] [CrossRef]
- Lambrechts, M.; Pretorius, I. Yeast and its importance to wine aroma. S. Afr. J. Enol. Vitic. 2000, 21, 97–129. [Google Scholar] [CrossRef]
- Amerine, M.; Berg, H.; Kunkee, R.; Ough, C.; Singleton, V.; Webb, A. The Technology of Wine Making, 4th ed.; AVI Publishing Co.: Westport, CT, USA, 1980; pp. 50–75. [Google Scholar]
- Catania, C.; Avagnina, S. Curso Superior de Degustación de Vinos; INTA EEA: Mendoza, Argentina; INTA: Ciudad de Buenos Aires, Argentina, 2007. [Google Scholar]
- King, E.S.; Stoumen, M.; Buscema, F.; Hjelmeland, A.K.; Ebeler, S.E.; Heymann, H.; Boulton, R.B. Regional sensory and chemical characteristics of Malbec wines from Mendoza and California. Food Chem. 2014, 143, 256–267. [Google Scholar] [CrossRef]
- Goldner, M.C.; Zamora, M.C.; Di Leo Lira, P.; Gianninoto, H.; Bandoni, A. Effect of ethanol level in the perception of aroma attributes and the detection of volatile compounds in red wine. J. Sens. Stud. 2009, 24, 243–257. [Google Scholar] [CrossRef]
- Pereira, V.; Albuquerque, F.; Ferreira, A.; Cacho, J.; Marques, J. Evolution of 5-hydroxymethylfurfural (HMF) and furfural (F) in fortified wines submitted to overheating conditions. Food Res. Int. 2011, 44, 71–76. [Google Scholar] [CrossRef]
- Hale, M.D.; Mccafferty, K.; Larmie, E.; Newton, J.; Swan, J.S. The influence of oak seasoning and toasting parameters on the composition and quality of wine. Am. J. Enol. Vitic. 1999, 50, 495–502. [Google Scholar] [CrossRef]
- Delgado de la Torre, M.P.; Priego-Capote, F.; Luque de Castro, M.a.D. Evaluation of the composition of vine shoots and oak chips for oenological purposes by superheated liquid extraction and high-resolution liquid chromatography–time-of-flight/mass spectrometry analysis. J. Agric. Food Chem. 2012, 60, 3409–3417. [Google Scholar] [CrossRef]
- Waterhouse, A.L.; Sacks, G.L.; Jeffery, D.W. Understanding Wine Chemistry; John Wiley & Sons: Hoboken, NJ, USA, 2024. [Google Scholar]
- Maarman, B.C. Interaction Between Wine Yeast and Malolactic Bacteria and the Impact on Wine Aroma and Flavour. Ph.D. Thesis, Stellenbosch University, Stellenbosch, South Africa, 2014. [Google Scholar]
- Guerrini, L.; Masella, P.; Angeloni, G.; Sacconi, A.; Calamai, L.; Parenti, A. Effects of a small increase in carbon dioxide pressure during fermentation on wine aroma. Foods 2020, 9, 1496. [Google Scholar] [CrossRef]
- Pineau, B.; Barbe, J.-C.; Van Leeuwen, C.; Dubourdieu, D. Examples of perceptive interactions involved in specific “red-” and “black-berry” aromas in red wines. J. Agric. Food Chem. 2009, 57, 3702–3708. [Google Scholar] [CrossRef]
- Gammacurta, M.; Lytra, G.; Marchal, A.; Marchand, S.; Barbe, J.C.; Moine, V.; de Revel, G. Influence of lactic acid bacteria strains on ester concentrations in red wines: Specific impact on branched hydroxylated compounds. Food Chem. 2018, 239, 252–259. [Google Scholar] [CrossRef]
- Sumby, K.M.; Jiranek, V.; Grbin, P.R. Ester synthesis and hydrolysis in an aqueous environment, and strain specific changes during malolactic fermentation in wine with Oenococcus oeni. Food Chem. 2013, 141, 1673–1680. [Google Scholar] [CrossRef]
- Antalick, G.; Perello, M.-C.; de Revel, G. Characterization of fruity aroma modifications in red wines during malolactic fermentation. J. Agric. Food Chem. 2012, 60, 12371–12383. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Aromatic Compound Type | FT-B (Spontaneous MLF) | FT-A (Guided MLF) (UNQLP1001 + UNQOe1101) |
|---|---|---|
| Alcohols | 92.38% | 90.06% |
| Esters | 7.57% | 9.90% |
| Terpenes | 0.01% | 0.01% |
| Aldehydes and Ketones | 0.04% | 0.03% |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rivas, G.A.; Flores, N.E.; Brizuela, N.S.; Guillade, A.C.; Semorile, L.C.; Delfederico, L. Novel Malolactic Fermentation Starter Formulated Using Native Lactic Acid Bacteria Strains from a Re-Emerging Wine-Growing Region of Argentina—A Pilot Scale Vinification. Fermentation 2025, 11, 140. https://doi.org/10.3390/fermentation11030140
Rivas GA, Flores NE, Brizuela NS, Guillade AC, Semorile LC, Delfederico L. Novel Malolactic Fermentation Starter Formulated Using Native Lactic Acid Bacteria Strains from a Re-Emerging Wine-Growing Region of Argentina—A Pilot Scale Vinification. Fermentation. 2025; 11(3):140. https://doi.org/10.3390/fermentation11030140
Chicago/Turabian StyleRivas, Gabriel Alejandro, Naiquen Elizabeth Flores, Natalia Soledad Brizuela, Andrea Cecilia Guillade, Liliana Carmen Semorile, and Lucrecia Delfederico. 2025. "Novel Malolactic Fermentation Starter Formulated Using Native Lactic Acid Bacteria Strains from a Re-Emerging Wine-Growing Region of Argentina—A Pilot Scale Vinification" Fermentation 11, no. 3: 140. https://doi.org/10.3390/fermentation11030140
APA StyleRivas, G. A., Flores, N. E., Brizuela, N. S., Guillade, A. C., Semorile, L. C., & Delfederico, L. (2025). Novel Malolactic Fermentation Starter Formulated Using Native Lactic Acid Bacteria Strains from a Re-Emerging Wine-Growing Region of Argentina—A Pilot Scale Vinification. Fermentation, 11(3), 140. https://doi.org/10.3390/fermentation11030140