
Differences in the Volatile Profile of Apple Cider Fermented with Schizosaccharomyces pombe and Schizosaccharomyces japonicus

 , , ,
, , ,  ,
,  and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Yeast Strains and Inoculum Preparation
2.2. Cider Fermentation Kinetics and pH Monitoring
2.3. High-Performance Liquid Chromatography (HPLC) Analysis
2.4. Volatile Organic Compound Analysis
2.5. Statistical Analysis
3. Results
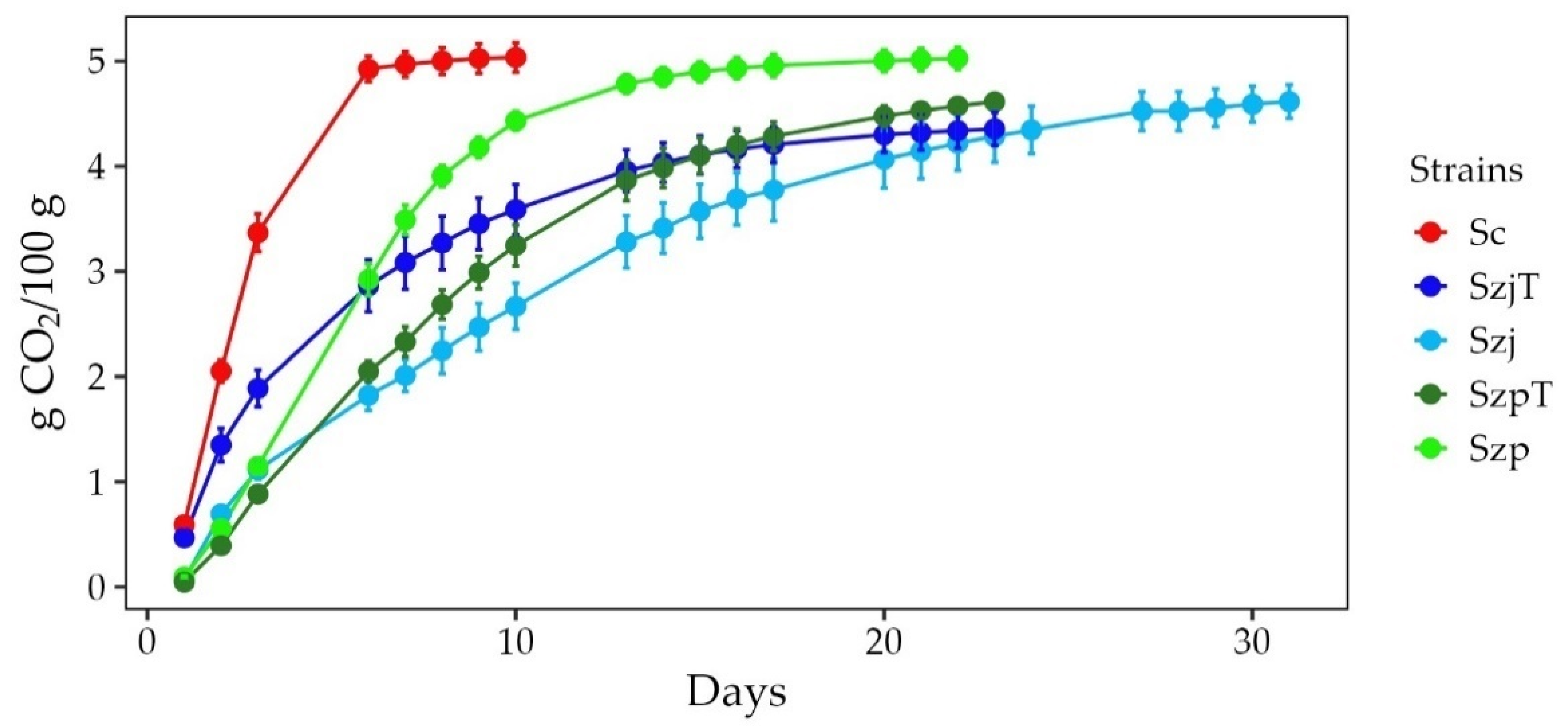
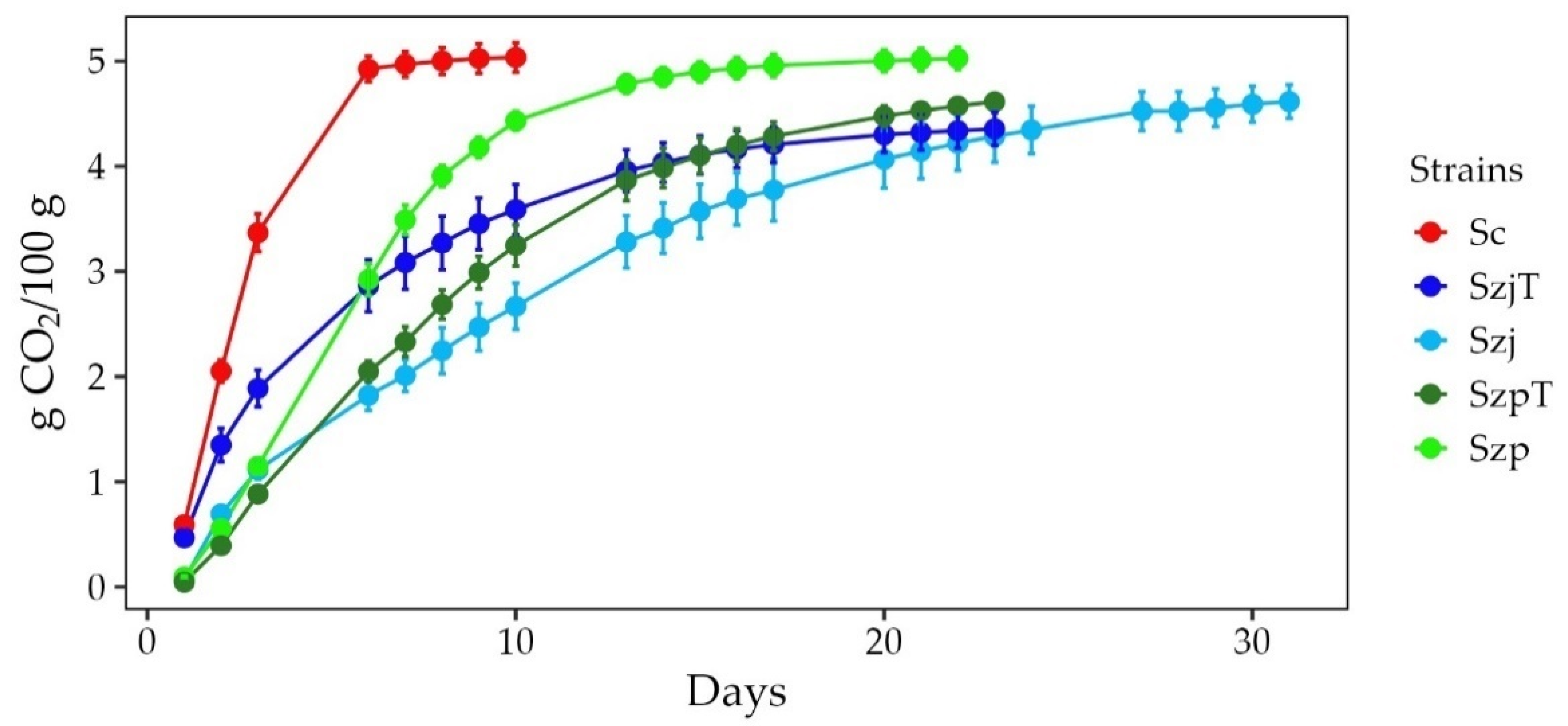
3.1. Fermentations Kinetics
3.2. Fermentation Parameters
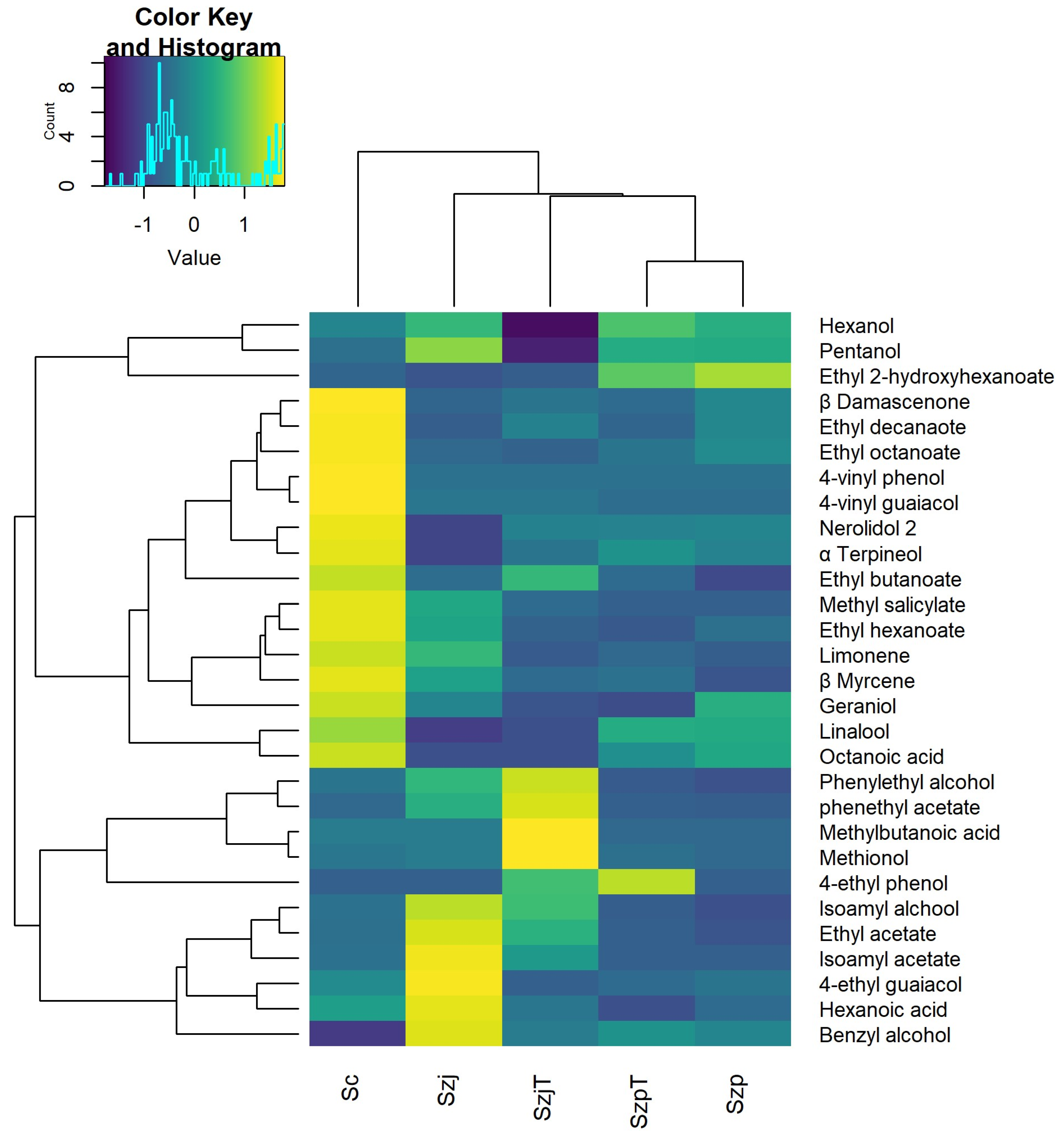
3.3. Volatile Organic Compounds in Apple Cider
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Magalhães, F.; Krogerus, K.; Vidgren, V.; Sandell, M.; Gibson, B. Improved cider fermentation performance and quality with newly generated Saccharomyces cerevisiae × Saccharomyces eubayanus hybrids. J. Ind. Microbiol. Biotechnol. 2017, 44, 1203–1213. [Google Scholar] [CrossRef]
- Calvert, M.D.; Neill, C.L.; Stewart, A.C.; Chang, E.A.B.; Whitehead, S.R.; Lahne, J. Appeal of the apple: Exploring consumer perceptions of hard cider in the Northeast and mid-Atlantic United States. J. Am. Soc. Brew. Chem. 2023. [Google Scholar] [CrossRef]
- Becot, F.A.; Bradshaw, T.L.; Conner, D.S. Apple market expansion through value-added hard cider production: Current production and prospects in Vermont. HortTechnology 2016, 26, 220–229. [Google Scholar] [CrossRef]
- Marcotte, B.V.; Verheyde, M.; Pomerleau, S.; Doyen, A.; Couillard, C. Health benefits of apple juice consumption: A review of interventional trials on humans. Nutrients 2022, 14, 821. [Google Scholar] [CrossRef] [PubMed]
- Calugar, P.C.; Coldea, T.E.; Salanță, L.C.; Pop, C.R.; Pasqualone, A.; Burja-Udrea, C.; Zhao, H.; Mudura, E. An overview of the factors influencing apple cider sensory and microbial quality from raw materials to emerging processing technologies. Processes 2021, 9, 502. [Google Scholar] [CrossRef]
- Valles, B.S.; Bedriñana, R.P.; Tascón, N.F.; Simón, A.Q.; Madrera, R.R. Yeast species associated with the spontaneous fermentation of cider. Food Microbiol. 2007, 24, 25–31. [Google Scholar] [CrossRef] [PubMed]
- Cousin, F.J.; Le Guellec, R.; Schlusselhuber, M.; Dalmasso, M.; Laplace, J.M.; Cretenet, M. Microorganisms in fermented apple beverages: Current knowledge and future directions. Microorganisms 2017, 5, 39. [Google Scholar] [CrossRef]
- Wu, Y.; Li, Z.; Zou, S.; Dong, L.; Lin, X.; Chen, Y.; Zhang, S.; Ji, C.; Liang, H. chemical composition and flavor characteristics of cider fermented with Saccharomyces cerevisiae and non-Saccharomyces cerevisiae. Foods 2023, 12, 3565. [Google Scholar] [CrossRef] [PubMed]
- Lorenzini, M.; Simonato, B.; Slaghenaufi, D.; Ugliano, M.; Zapparoli, G. Assessment of yeasts for apple juice fermentation and production of cider volatile compounds. LWT-Food Sci. Technol. 2019, 99, 224–230. [Google Scholar] [CrossRef]
- Wei, J.; Zhang, Y.; Yuan, Y.; Dai, L.; Yue, T. Characteristic fruit wine production via reciprocal selection of juice and non-Saccharomyces species. Food Microbiol. 2019, 79, 66–74. [Google Scholar] [CrossRef]
- Gutiérrez, A.; Boekhout, T.; Gojkovic, Z.; Katz, M. Evaluation of non-Saccharomyces yeasts in the fermentation of wine, beer, and cider for the development of new beverages. J. Inst. Brew. 2018, 124, 389–402. [Google Scholar] [CrossRef]
- Satora, P.; Semik-Szczurak, D.; Tarko, T.; Buldys, A. Influence of selected Saccharomyces and Schizosaccharomyces strains and their mixed cultures on chemical composition of apple wines. J. Food Sci. 2018, 83, 424–431. [Google Scholar] [CrossRef] [PubMed]
- He, W.; Liu, S.; Heponiemi, P.; Heinonen, M.; Marsol-Vall, A.; Ma, X.; Yang, B.; Laaksonen, O. Effect of Saccharomyces cerevisiae and Schizosaccharomyces pombe strains on chemical composition and sensory quality of ciders made from Finnish apple cultivars. Food Chem. 2021, 345, 128833. [Google Scholar] [CrossRef] [PubMed]
- Benito, S. The impacts of Schizosaccharomyces on winemaking. Appl. Microbiol. Biotechnol. 2019, 103, 4291–4312. [Google Scholar] [CrossRef] [PubMed]
- Binati, R.L.; Lemos Junior, W.J.F.; Luzzini, G.; Slaghenaufi, D.; Ugliano, M.; Torriani, S. Contribution of non-Saccharomyces yeasts to wine volatile and sensory diversity: A study on Lachancea thermotolerans, Metschnikowia spp. and Starmerella bacillaris strains isolated in Italy. Int. J. Food Microbiol. 2020, 318, 108470. [Google Scholar] [CrossRef] [PubMed]
- Slaghenaufi, D.; Boscaini, A.; Prandi, A.; Dal Cin, A.; Zandonà, V.; Luzzini, G.; Ugliano, M. Influence of different modalities of grape withering on volatile compounds of young and aged Corvina wines. Molecules 2020, 25, 2141. [Google Scholar] [CrossRef] [PubMed]
- Slaghenaufi, D.; Ugliano, M. Norisoprenoids, sesquiterpenes and terpenoids content of Valpolicella wines during aging: Investigating aroma potential in relationship to evolution of tobacco and balsamic aroma in aged wine. Front. Chem. 2018, 6, 66. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021; Available online: https://www.R-project.org/ (accessed on 18 December 2023).
- Calleja, G.B.; Johnson, B.F. Temperature sensitivity of flocculation induction, conjugation and sporulation in fission yeast. Antoni Van Leeuwenhoek 1979, 45, 391–400. [Google Scholar] [CrossRef]
- Nešpor, J.; Karabín, M.; Štulíková, K.; Dostálek, P. An HS-SPME-GC-MS method for profiling volatile compounds as related to technology used in cider production. Molecules 2019, 24, 2117. [Google Scholar] [CrossRef]
- Way, M.L.; Jones, J.E.; Longo, R.; Dambergs, R.G.; Swarts, N.D. A preliminary study of yeast strain influence on chemical and sensory characteristics of apple cider. Fermentation 2022, 8, 455. [Google Scholar] [CrossRef]
- European Cider and Fruit Wine Association. 2018. Available online: https://food.ec.europa.eu/system/files/2018-03/fs_labelling-nutrition_legis_alcohol-self-regulatory-proposal_cider_en.pdf (accessed on 18 December 2023).
- Domizio, P.; Lencioni, L.; Calamao, L.; Portaro, L.; Bisson, L.F. Evaluation of the yeast Schizosaccharomyces japonicus for use in wine production. Am. J. Enol. Vitic. 2018, 69, 266–277. [Google Scholar] [CrossRef]
- Zhao, X.; Procopio, S.; Becker, T. Flavor impacts of glycerol in the processing of yeast fermented beverages: Review. J. Food Sci. Technol. 2015, 52, 7588–7598. [Google Scholar] [CrossRef] [PubMed]
- Duvnjak, Z.; Turcotte, G.; Duan, Z.D. Production and consumption of sorbitol and fructose by Saccharomyces cerevisiae ATCC 36859. J. Chem. Technol. Biotechnol. 1991, 52, 527–537. [Google Scholar] [CrossRef]
- Grembecka, M. Sugar alcohols-their role in the modern world of sweeteners: A review. Eur. Food Res. Technol. 2015, 241, 1–14. [Google Scholar] [CrossRef]
- Verstrepen, K.J.; Van Laere, S.D.; Vanderhaegen, B.M.; Derdelinckx, G.; Dufour, J.P.; Pretorius, I.S.; Winderickx, J.; Thevelein, J.M.; Delvaux, F.R. Expression levels of the yeast alcohol acetyltransferase genes ATF1, Lg-ATF1, and ATF2 control the formation of a broad range of volatile esters. Appl. Environ. Microbiol. 2003, 69, 5228–5237. [Google Scholar] [CrossRef]
- Peng, B.; Li, F.; Cui, L.; Guo, Y. Effects of fermentation temperature on key aroma compounds and sensory properties of apple wine. J. Food Sci. 2015, 80, 2937–2943. [Google Scholar] [CrossRef] [PubMed]
- Satora, P.; Tarko, T.; Duda-Chodak, A.; Sroka, P.; Tuszynski, T.; Czepielik, M. Influence of prefermentative treatments and fermentation on the antioxidant and volatile profiles of apple wines. J. Agric. Food Chem. 2009, 57, 11209–11217. [Google Scholar] [CrossRef]
- Wang, L.P.; Xu, Y.; Zhao, G.A.; Li, J.M. Rapid analysis of flavor volatile in apple wine using headspace solid-phase microextraction. J. Inst. Brew. 2004, 110, 57–65. [Google Scholar] [CrossRef]
- Carpena, M.; Fraga-Corral, M.; Otero, P.; Nogueira, R.A.; Garcia-Oliveira, P.; Prieto, M.A.; Simal-Gandara, J. Secondary aroma: Influence of wine microorganisms in their aroma profile. Foods 2021, 10, 51. [Google Scholar] [CrossRef]
- Lilly, M.; Lambrechts, M.G.; Pretorius, I.S. Effect of increased yeast alcohol acetyltransferase activity on flavor profiles of wine and distillates. Appl. Environ. Microbiol. 2000, 66, 744–753. [Google Scholar] [CrossRef]
- Etschmann, M.; Bluemke, W.; Sell, D.; Schrader, J. Biotechnological production of 2-phenylethanol. Appl. Microbiol. Biotechnol. 2002, 59, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Hazelwood, A.A.; Daran, J.M.; van Maris, A.J.A.; Pronk, J.T.; Dickinson, R. Ehrlich pathway for fusel alcohol production: A century of research on Saccharomyces cerevisiae metabolism. Appl. Environ. Microbiol. 2008, 74, 2259–2266. [Google Scholar] [CrossRef] [PubMed]
- Portaro, L.; Maioli, F.; Canuti, V.; Picchi, M.; Lencioni, L.; Mannazzu, I.; Domizio, P. Schizosaccharomyces japonicus/Saccharomyces cerevisiae mixed starter cultures: New perspectives for the improvement of Sangiovese aroma, taste, and color stability. LWT-Food Sci. Technol. 2022, 156, 113009. [Google Scholar] [CrossRef]
- Česnik, U.; Martelanc, M.; Øvsthus, I.; Radovanović Vukajlović, T.; Hosseini, A.; Mozetič Vodopivec, B.; Butinar, L. Functional characterization of Saccharomyces yeasts from cider produced in Hardanger. Fermentation 2023, 9, 824. [Google Scholar] [CrossRef]
- Yu, W.; Zhu, Y.; Zhu, R.; Bai, J.; Qiu, J.; Wu, Y.; Zhong, K.; Gao, H. Insight into the characteristics of cider fermented by single and co-culture with Saccharomyces cerevisiae and Schizosaccharomyces pombe based on metabolomic and transcriptomic approaches. LWT-Food Sci. Technol. 2022, 163, 113538. [Google Scholar] [CrossRef]
- Ribéreau-Gayon, P.; Boidron, J.N.; Terrier, A. Aroma of Muscat grape varieties. J. Agric. Food Chem. 1975, 23, 1042–1047. [Google Scholar] [CrossRef]
- Bingman, M.T.; Stellick, C.E.; Pelkey, J.P.; Scott, J.M.; Cole, C.A. Monitoring cider aroma development throughout the fermentation process by Headspace Solid Phase Microextraction (HS-SPME) Gas Chromatography–Mass Spectrometry (GC-MS) analysis. Beverages 2020, 6, 40. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
| Species | Strain | Strain Designation | Origin and Isolation Source |
|---|---|---|---|
| Schizosaccharomyces pombe | NCAIM Y01474T | SzpT | NCAIM culture collection (Budapest, Hungary), isolated from Arak (a fermented beverage) macerate |
| Schizosaccharomyces pombe | SPBS | Szp | Agricultural and Food Sciences Department of Bologna University culture collection (Bologna, Italy), isolated from fruit syrup |
| Schizosaccharomyces japonicus | DBVPG 6274T | SzjT | DBVPG culture collection (Perugia, Italy), isolated from strawberry wine |
| Schizosaccharomyces japonicus | M23B | Szj | VUCC-DBT culture collection (Verona, Italy), isolated from grape must (Rovereto, TN, Italy) |
| Saccharomyces cerevisiae | EC1118 | Sc | Lallemand Inc. (Montreal, QC, Canada), isolated from French sparkling wine |
| Compound (g/L) | Apple Juice | Strain | ||||
|---|---|---|---|---|---|---|
| SzpT | Szp | SzjT | Szj | Sc | ||
| Fructose | 91.14 | N.D. | N.D. | N.D. | N.D. | N.D. |
| Glucose | 40.42 | N.D. | N.D. | N.D. | 0.17 ± 0.09 | N.D. |
| Sucrose | 13.96 | N.D. | N.D. | N.D. | N.D. | N.D. |
| Citrate | N.D. | 0.34 ± 0.01 a | 0.37 ± 0.03 a | N.D. | N.D. | N.D. |
| Malate | 3.25 | 0.80 ± 0.01 b | 0.79 ± 0.02 b | 0.69 ± 0.01 c | 0.11 ± 0.02 d | 3.22 ± 0.02 a |
| Glycerol | N.D. | 3.89 ± 0.18 c | 4.01 ± 0.10 c | 7.21 ± 0.53 b | 8.46 ± 0.57 a | 3.10 ± 0.27 c |
| Sorbitol | N.D. | 5.89 ± 1.66 a | 5.57 ± 1.50 a | 4.39 ± 1.03 a | 6.56 ± 0.65 a | 6.41 ± 1.20 a |
| Acetate | N.D. | 0.28 ± 0.02 | N.D. | N.D. | N.D. | N.D. |
| Ethanol (v/v, %) | N.D. | 6.93 ± 0.35 ab | 7.16 ± 0.24 ab | 6.07 ± 0.32 c | 6.53 ± 0.11 bc | 7.27 ± 0.27 a |
| pH | 3.70 | 3.27 ± 0.05 c | 3.54 ± 0.13 bc | 3.73 ± 0.04 b | 4.42 ± 0.21 a | 3.67 ± 0.01 b |
| Compounds | OT ‡ | Strain | ||||
|---|---|---|---|---|---|---|
| SzpT | Szp | Szjt | Szj | Sc | ||
| Esters | ||||||
| Ethyl acetate | 12 mg/L | 2.71 | 1.47 | 15.08 | 26.29 | 5.27 |
| Ethyl butanoate | 20 µg/L | 6.27 | 5.22 | 9.14 | 6.39 | 11.29 |
| Ethyl hexanoate | 14 µg/L | 9.13 | 12.26 | 10.63 | 19.77 | 32.05 |
| Ethyl decanoate | 200 µg/L | 0.25 | 0.56 | 0.51 | 0.17 | 1.76 |
| Isoamyl acetate | 30 µg/L | 6.12 | 4.83 | 72.23 | 199.8 | 25.81 |
| 2-phenethyl acetate | 108 µg/L | 1.29 | 1.21 | 10.87 | 6.1 | 1.82 |
| Alcohols | ||||||
| Isoamyl alcohol | 30 mg/L | 1.42 | 0.83 | 5.92 | 8.26 | 2.37 |
| Phenylethyl alcohol | 390 µg/L | 7.38 | 6.99 | 15.89 | 12.42 | 8.8 |
| Fatty acids | ||||||
| Hexanoic acid | 420 µg/L | 4.29 | 6.48 | 7.39 | 19.14 | 10.74 |
| Octanoic acid | 500 µg/L | 21.61 | 28.07 | 4.08 | 4.29 | 50.1 |
| 3-Methylbutanoic acid | 250 µg/L | 0.07 | 0.14 | 9.45 | 1.2 | 1.22 |
| Norisoprenoids | ||||||
| β-Damascenone | 0.05 µg/L | 14.4 | 20.8 | 16.33 | 13.1 | 49.73 |
| Benzenoids | ||||||
| 4-vinyl guaiacol | 40 µg/L | 0.08 | 0.1 | 0.29 | 0.33 | 3.76 |
| 4-vinyl phenol | 180 µg/L | 0.02 | 0.02 | 0.03 | 0.03 | 2.09 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ferremi Leali, N.; Salvetti, E.; Luzzini, G.; Salini, A.; Slaghenaufi, D.; Fusco, S.; Ugliano, M.; Torriani, S.; Binati, R.L. Differences in the Volatile Profile of Apple Cider Fermented with Schizosaccharomyces pombe and Schizosaccharomyces japonicus. Fermentation 2024, 10, 128. https://doi.org/10.3390/fermentation10030128
Ferremi Leali N, Salvetti E, Luzzini G, Salini A, Slaghenaufi D, Fusco S, Ugliano M, Torriani S, Binati RL. Differences in the Volatile Profile of Apple Cider Fermented with Schizosaccharomyces pombe and Schizosaccharomyces japonicus. Fermentation. 2024; 10(3):128. https://doi.org/10.3390/fermentation10030128
Chicago/Turabian StyleFerremi Leali, Nicola, Elisa Salvetti, Giovanni Luzzini, Andrea Salini, Davide Slaghenaufi, Salvatore Fusco, Maurizio Ugliano, Sandra Torriani, and Renato L. Binati. 2024. "Differences in the Volatile Profile of Apple Cider Fermented with Schizosaccharomyces pombe and Schizosaccharomyces japonicus" Fermentation 10, no. 3: 128. https://doi.org/10.3390/fermentation10030128
APA StyleFerremi Leali, N., Salvetti, E., Luzzini, G., Salini, A., Slaghenaufi, D., Fusco, S., Ugliano, M., Torriani, S., & Binati, R. L. (2024). Differences in the Volatile Profile of Apple Cider Fermented with Schizosaccharomyces pombe and Schizosaccharomyces japonicus. Fermentation, 10(3), 128. https://doi.org/10.3390/fermentation10030128