

Novel Placenta-Derived Liquid Product Suitable for Cosmetic Application Produced by Fermentation and Digestion of Porcine or Equine Placenta Using Lactic Acid Bacterium Enterococcus faecalis PR31


Abstract
1. Introduction
2. Materials and Methods
2.1. Lactic Acid Bacteria
2.2. The Placenta
2.3. Bacterial Culture and Fermentation
2.4. Cell Proliferation Assay with Normal Human Epidermal Keratinocytes and Dermal Fibroblasts
2.5. Total Nitrogen Concentration
2.6. Size Exclusion Chromatography
2.7. Equilibrium Moisture Content
2.8. Metabolome Analysis
2.9. Analysis of Stratum Corneum Water Content
2.10. Statistics
3. Results
3.1. Digestioin and Solubilization by Fermentation of the Placenta
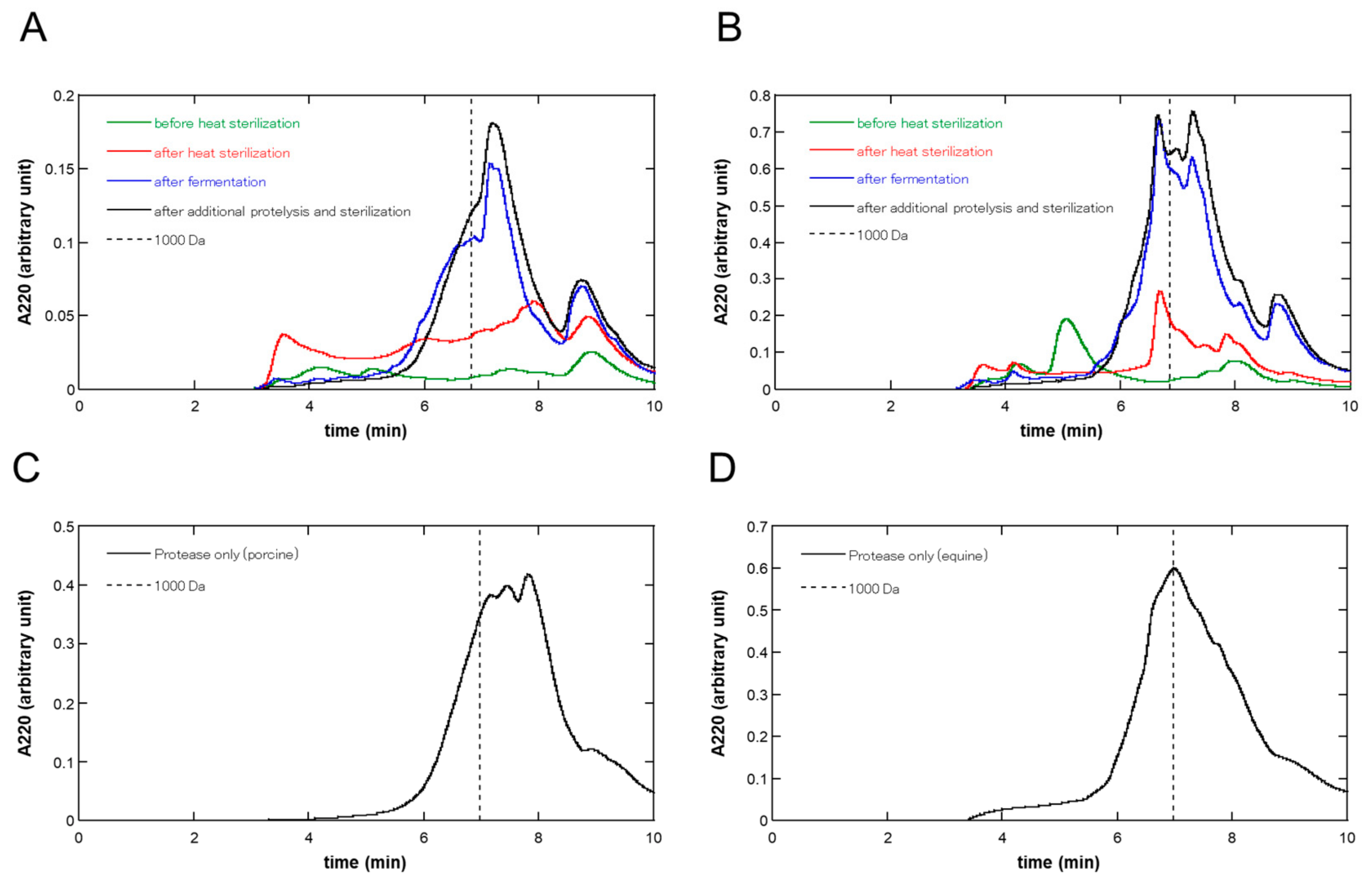
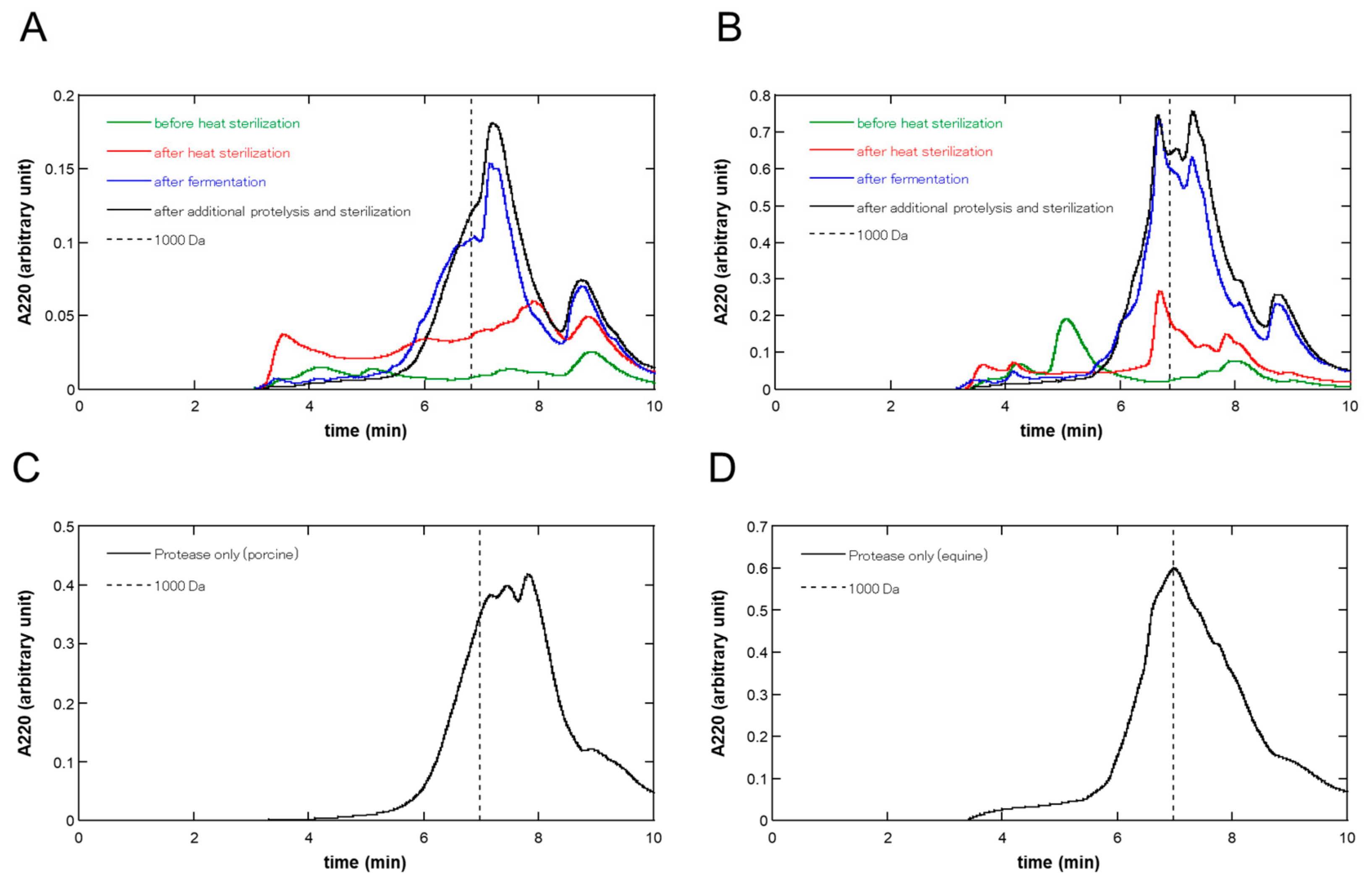
3.2. Molecular Weight Distribution at Each Step of the Fermentation of the Placenta
3.3. Changes in the Amounts of Metabolites Due to Fermentation
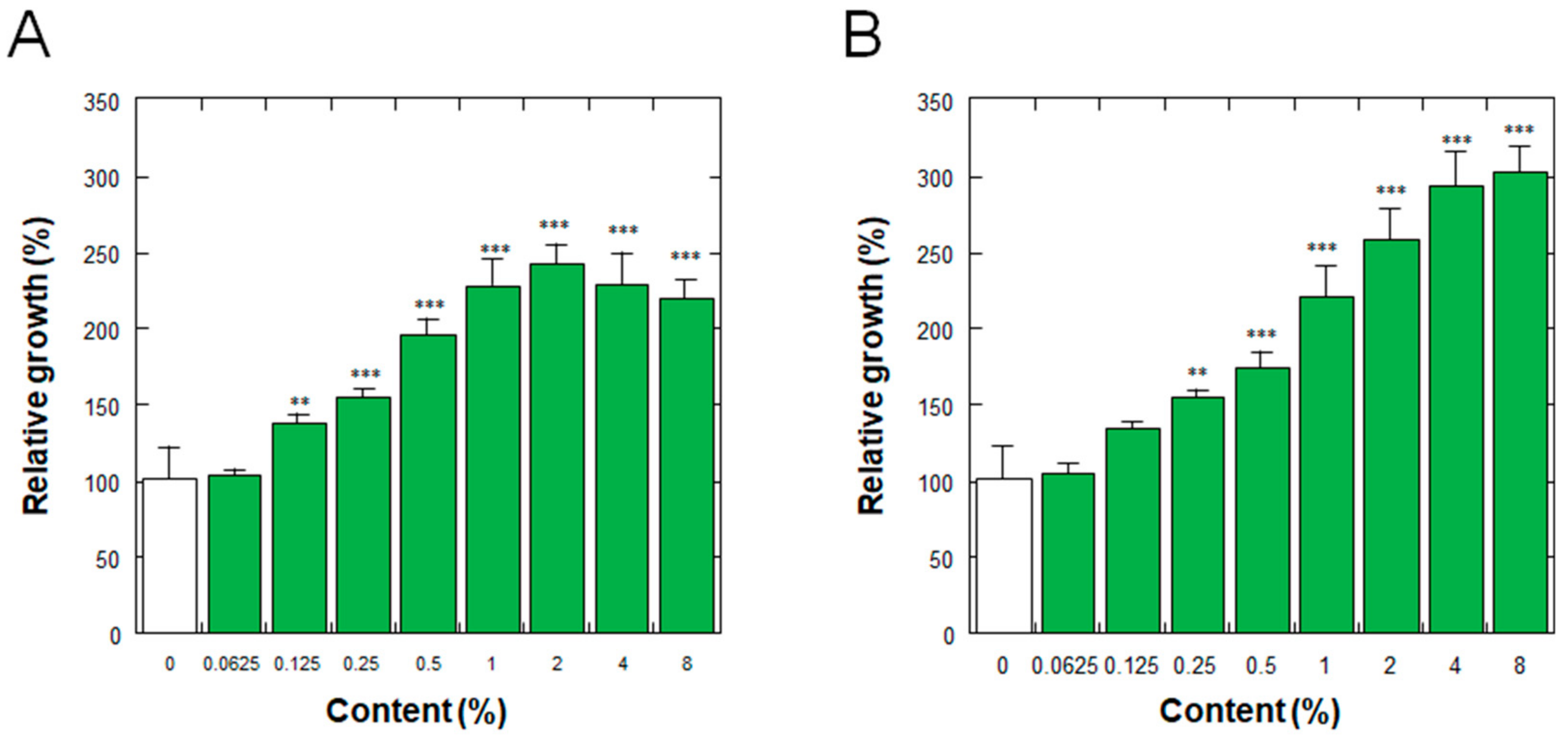
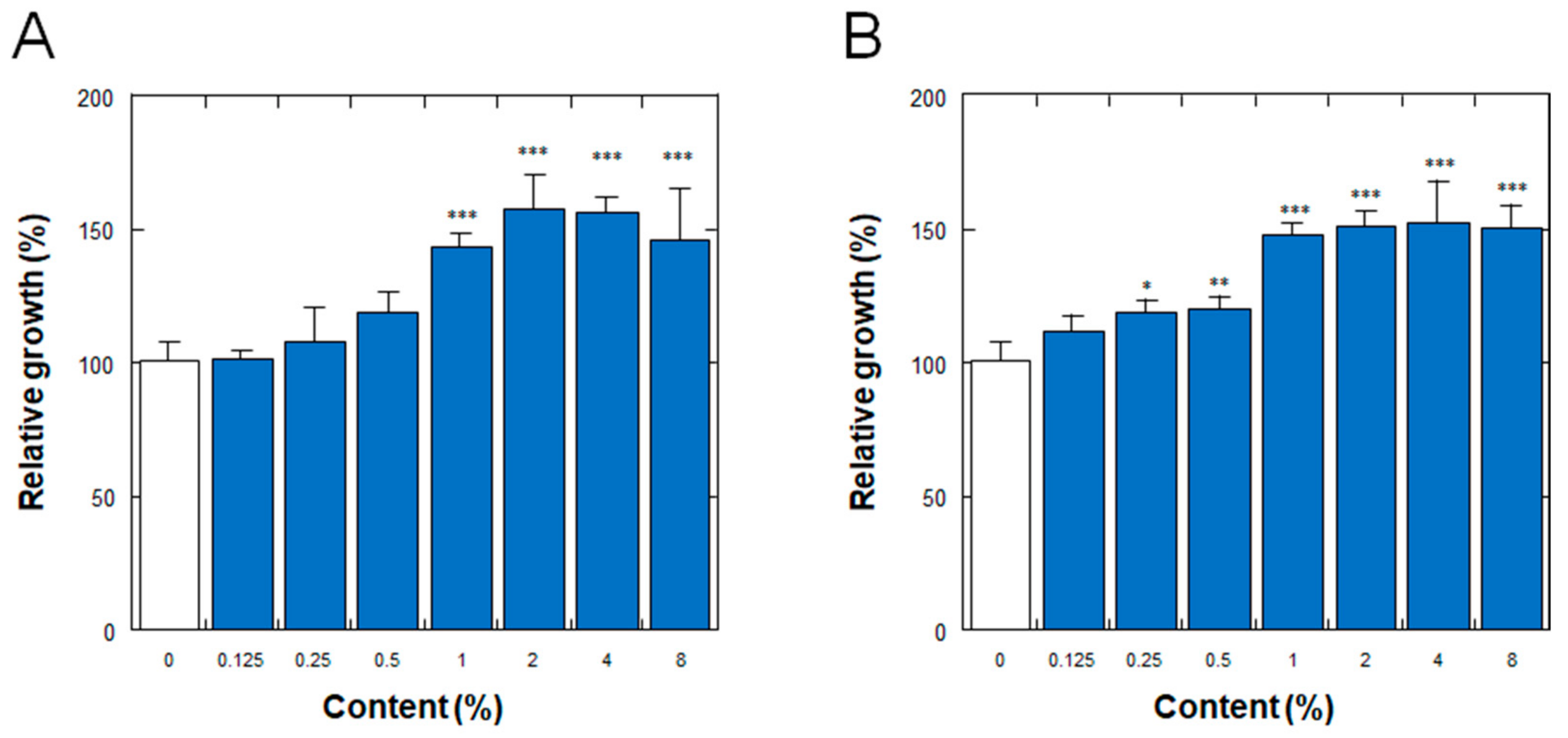
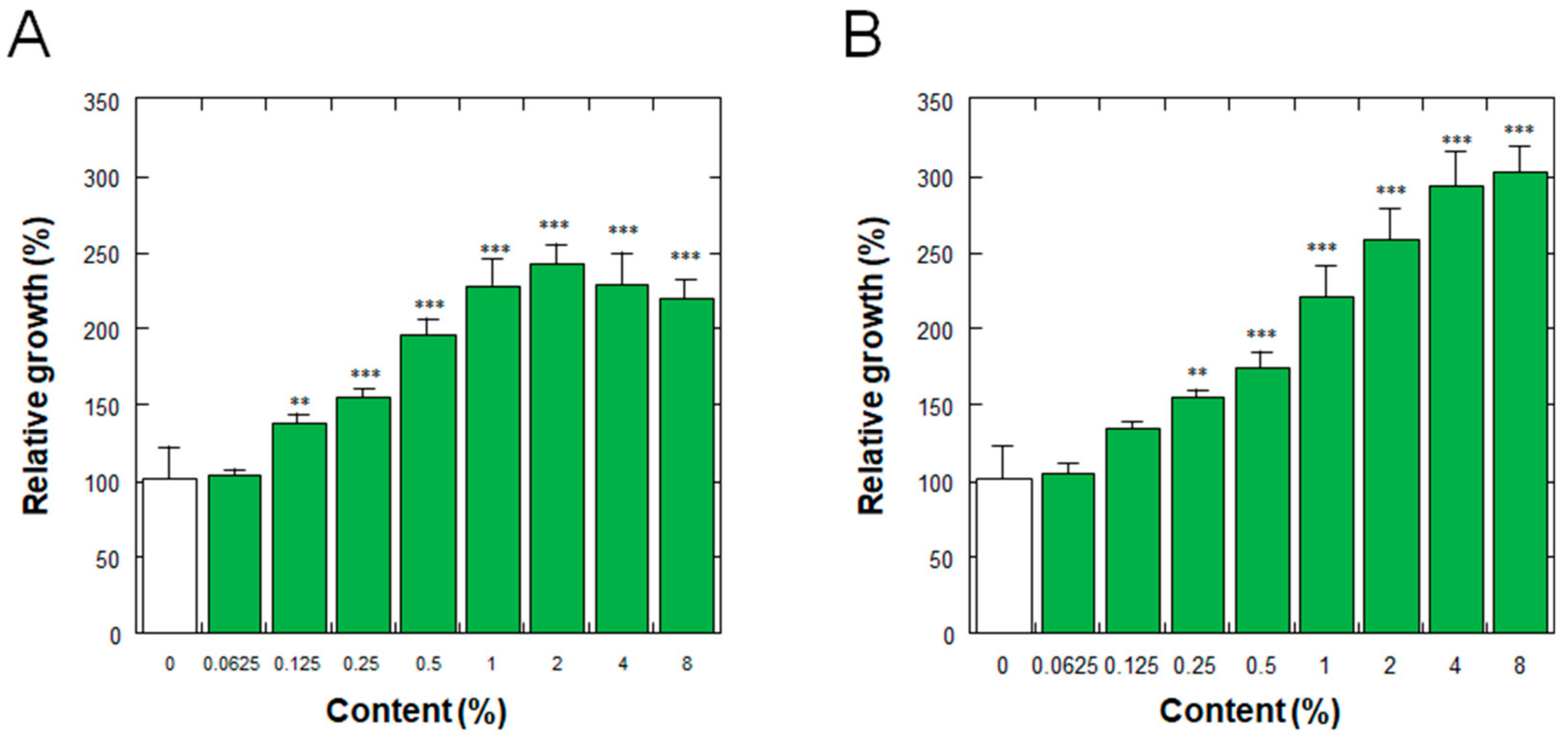
3.4. Effect on the Proliferation of Normal Human Dermal Fibroblasts
3.5. Effect on the Proliferation of Normal Human Epidermal Keratinocytes
3.6. Equilibrium Moisture Content
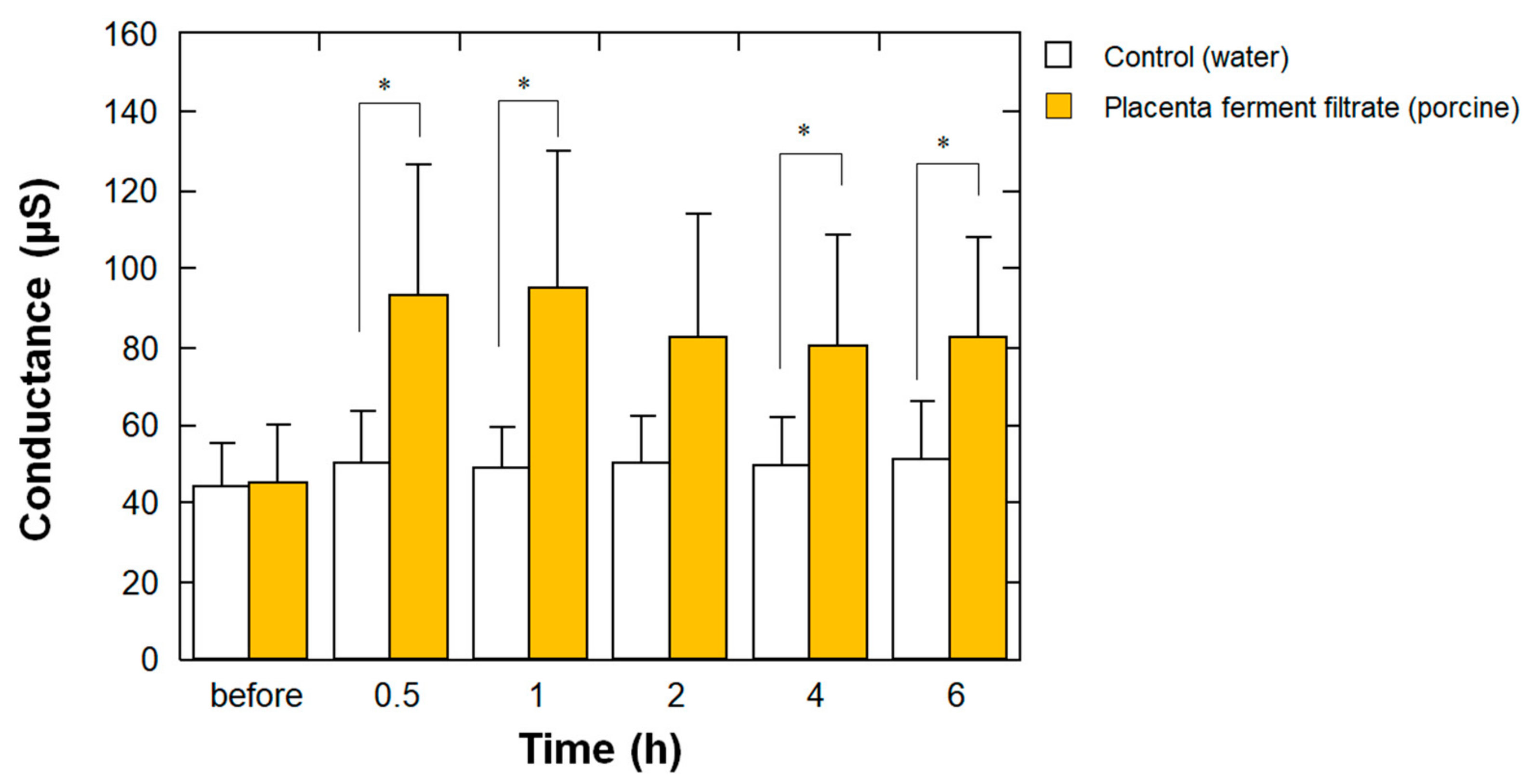
3.7. Stratum Corneum Water Content
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Soykova-Pachnerova, E.; Brutar, V.; Golova, B.; Zvolska, E. Placenta as a lactagogon. Gynaecologia 1954, 138, 617–627. [Google Scholar] [CrossRef]
- Kristal, M.B. Placentophagia: A biobehavioral enigma (or De gustibus non disputandum est). Neurosci. Biobehav. Rev. 1980, 4, 141–150. [Google Scholar] [CrossRef]
- Nunn, J.F. Ancient Gyptian Medicine; University of Oklahoma Press: Norman, OK, USA, 2002. [Google Scholar]
- Silini, A.R.; Cargnoni, A.; Magatti, M.; Pianta, S.; Parolini, O. The Long Path of Human Placenta, and Its Derivatives, in Regenerative Medicine. Front. Bioeng. Biotechnol. 2015, 3, 162. [Google Scholar] [CrossRef]
- Barat, T.; Abdollahimajd, F.; Dadkhahfar, S.; Moravvej, H. Evaluation of the efficacy and safety of cow placenta extract lotion versus minoxidil 2% in the treatment of female pattern androgenetic alopecia. Int. J. Womens Dermatol. 2020, 6, 318–321. [Google Scholar] [CrossRef]
- Nagae, M.; Nagata, M.; Teramoto, M.; Yamakawa, M.; Matsuki, T.; Ohnuki, K.; Shimizu, K. Effect of Porcine Placenta Extract Supplement on Skin Condition in Healthy Adult Women: A Randomized, Double-Blind Placebo-Controlled Study. Nutrients 2020, 12, 1671. [Google Scholar] [CrossRef]
- Kitanohara, M.; Yamamoto, T.; Masunaga, S.; Ohishi, M.; Komatsu, Y.; Nagase, M. Effect of porcine placental extract on the mild menopausal symptoms of climacteric women. Climacteric 2017, 20, 144–150. [Google Scholar] [CrossRef] [PubMed]
- Nagase, M.; Watanabe, C.; Kitanohara, M.; Nishiya, M.; Okada, T.; Ohishi, M.; Komatsu, Y. Improvement of Aspects of Subjective Sleep Quality of Healthy Volunteers by Ingestion of Porcine Placental Extract: A Randomized Cross-Over Pilot Study. Front. Nutr. 2020, 7, 550287. [Google Scholar] [CrossRef] [PubMed]
- Nagae, M.; Nishio, T.; Ohnuki, K.; Shimizu, K. Effects of oral administration of equine placental extract supplement on the facial skin of healthy adult women: A randomized, double-blind, placebo-controlled study. Health Sci. Rep. 2022, 5, e522. [Google Scholar] [CrossRef] [PubMed]
- Tebakari, M.; Katoh, S.; Hashimoto, H.; Kawashima, J.; Takano, F. Porcine placenta extract modulates the expression of skin functional proteins in cultured human fibroblasts and keratinocytes. Pharmacometrics 2015, 88, 1–6. [Google Scholar]
- Cho, H.R.; Ryou, J.H.; Lee, J.W.; Lee, M.H. The effects of placental extract on fibroblast proliferation. J. Cosmet. Sci. 2008, 59, 195–202. [Google Scholar] [PubMed]
- Ito, K.; Yamada, R.; Matsumoto, N.; Imamura, T. Evaluation of fibroblast growth factor activity exerted by placental extract used as a cosmetic ingredient. J. Cosmet. Dermatol. 2018, 17, 821–829. [Google Scholar] [CrossRef] [PubMed]
- El-Kilany, E.; Raoof, E. Facial Cosmetics in Ancient Egypt. Egypt. J. Tour. Stud. 2017, 16, 1–19. [Google Scholar]
- Stewart, S. Cosmetics & Perfumes in the Roman World; Tempus Publishing Limited: Stroud, UK, 2007. [Google Scholar]
- Pérez-Rivero, C.; López-Gómez, J.P. Unlocking the Potential of Fermentation in Cosmetics: A Review. Fermentation 2023, 9, 463. [Google Scholar] [CrossRef]
- Duarte, M.; Oliveira, A.L.; Oliveira, C.; Pintado, M.; Amaro, A.; Madureira, A.R. Current postbiotics in the cosmetic market-an update and development opportunities. Appl. Microbiol. Biotechnol. 2022, 106, 5879–5891. [Google Scholar] [CrossRef] [PubMed]
- Yoon, D.H.; Han, G.Y.; Hwang, S.S.; Lee, D.W.; Kim, J.S.; Kim, K.; Kim, J.; Song, W. The Effect of Fermented Porcine Placental Extract on Fatigue-Related Parameters in Healthy Adults: A Double-Blind, Randomized, Placebo-Controlled Trial. Nutrients 2020, 12, 3086. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.Y.; Han, N.R.; Kim, N.R.; Lee, M.; Kim, J.; Kim, C.J.; Jeong, H.J.; Kim, H.M. Effect of fermented porcine placenta on physical fatigue in mice. Exp. Biol. Med. 2016, 241, 1985–1996. [Google Scholar] [CrossRef]
- Mitsui, Y.; Bagchi, M.; Marone, P.A.; Moriyama, H.; Bagchi, D. Safety and toxicological evaluation of a novel, fermented, peptide-enriched, hydrolyzed swine placenta extract powder. Toxicol. Mech. Methods 2015, 25, 13–20. [Google Scholar] [CrossRef]
- Anderson, A.C.; Jonas, D.; Huber, I.; Karygianni, L.; Wolber, J.; Hellwig, E.; Arweiler, N.; Vach, K.; Wittmer, A.; Al-Ahmad, A. Enterococcus faecalis from Food, Clinical Specimens, and Oral Sites: Prevalence of Virulence Factors in Association with Biofilm Formation. Front. Microbiol. 2015, 6, 1534. [Google Scholar] [CrossRef]
- Strzelecki, J.; Hryniewicz, W.; Sadowy, E. Gelatinase-associated phenotypes and genotypes among clinical isolates of Enterococcus faecalis in Poland. Pol. J. Microbiol. 2011, 60, 287–292. [Google Scholar] [CrossRef]
- Eaton, T.J.; Gasson, M.J. Molecular screening of Enterococcus virulence determinants and potential for genetic exchange between food and medical isolates. Appl. Environ. Microbiol. 2001, 67, 1628–1635. [Google Scholar] [CrossRef]
- Ohashi, Y.; Hirayama, A.; Ishikawa, T.; Nakamura, S.; Shimizu, K.; Ueno, Y.; Tomita, M.; Soga, T. Depiction of metabolome changes in histidine-starved Escherichia coli by CE-TOFMS. Mol. Biosyst. 2008, 4, 135–147. [Google Scholar] [CrossRef]
- Ooga, T.; Sato, H.; Nagashima, A.; Sasaki, K.; Tomita, M.; Soga, T.; Ohashi, Y. Metabolomic anatomy of an animal model revealing homeostatic imbalances in dyslipidaemia. Mol. Biosyst. 2011, 7, 1217–1223. [Google Scholar] [CrossRef]
- Sugimoto, M.; Wong, D.T.; Hirayama, A.; Soga, T.; Tomita, M. Capillary electrophoresis mass spectrometry-based saliva metabolomics identified oral, breast and pancreatic cancer-specific profiles. Metabolomics 2010, 6, 78–95. [Google Scholar] [CrossRef]
- Tagami, H.; Ohi, M.; Iwatsuki, K.; Kanamaru, Y.; Yamada, M.; Ichijo, B. Evaluation of the skin surface hydration in vivo by electrical measurement. J. Investig. Dermatol. 1980, 75, 500–507. [Google Scholar] [CrossRef] [PubMed]
- Hashimoto-Kumasaka, K.; Takahashi, K.; Tagami, H. Electrical measurement of the water content of the stratum corneum in vivo and in vitro under various conditions: Comparison between skin surface hygrometer and corneometer in evaluation of the skin surface hydration state. Acta Derm. Venereol. 1993, 73, 335–339. [Google Scholar] [CrossRef] [PubMed]
- Choi, H.S.; Ha, K.Y.; Xu, X.Y.; Kang, H.C.; Kim, H.; Kim, Y.J. Off-Flavor Removal from Sheep Placenta via Fermentation with Novel Yeast Strain Brettanomyces deamine kh3 Isolated from Traditional Apple Vinegar. Molecules 2021, 26, 5835. [Google Scholar] [CrossRef]
- Xu, L.; Nagata, N.; Nagashimada, M.; Zhuge, F.; Ni, Y.; Chen, G.; Kamei, J.; Ishikawa, H.; Komatsu, Y.; Kaneko, S.; et al. A porcine placental extract prevents steatohepatitis by suppressing activation of macrophages and stellate cells in mice. Oncotarget 2018, 9, 15047–15060. [Google Scholar] [CrossRef]
- Ando, Y.; Sato, F.; Fukunaga, H.; Iwasaki, Y.; Chiba, Y.; Tebakari, M.; Daigo, Y.; Kawashima, J.; Kamei, J. Placental extract suppresses differentiation of 3T3-L1 preadipocytes to mature adipocytes via accelerated activation of p38 MAPK during the early phase of adipogenesis. Nutr. Metab. 2019, 16, 32. [Google Scholar] [CrossRef] [PubMed]
- Ando, Y.; Odawara, E.; Sakai, H.; Sato, F.; Kamei, J. Placental extract suppresses lipid droplet accumulation by autophagy during the differentiation of adipose-derived mesenchymal stromal/stem cells into mature adipocytes. BMC Res. Notes 2023, 16, 338. [Google Scholar] [CrossRef]
- O’Keefe, E.J.; Chiu, M.L. Stimulation of thymidine incorporation in keratinocytes by insulin, epidermal growth factor, and placental extract: Comparison with cell number to assess growth. J. Investig. Dermatol. 1988, 90, 2–7. [Google Scholar] [CrossRef]
- O’Keefe, E.J.; Payne, R.E.; Russell, N. Keratinocyte growth-promoting activity from human placenta. J. Cell Physiol. 1985, 124, 439–445. [Google Scholar] [CrossRef] [PubMed]
- Rawlings, A.V.; Matts, P.J. Stratum corneum moisturization at the molecular level: An update in relation to the dry skin cycle. J. Investig. Dermatol. 2005, 124, 1099–1110. [Google Scholar] [CrossRef] [PubMed]
- Sandilands, A.; Sutherland, C.; Irvine, A.D.; McLean, W.H. Filaggrin in the frontline: Role in skin barrier function and disease. J. Cell Sci. 2009, 122, 1285–1294. [Google Scholar] [CrossRef]
- Mlitz, V.; Latreille, J.; Gardinier, S.; Jdid, R.; Drouault, Y.; Hufnagl, P.; Eckhart, L.; Guinot, C.; Tschachler, E. Impact of filaggrin mutations on Raman spectra and biophysical properties of the stratum corneum in mild to moderate atopic dermatitis. J. Eur. Acad. Dermatol. Venereol. 2012, 26, 983–990. [Google Scholar] [CrossRef]
- Caspers, P.J.; Lucassen, G.W.; Carter, E.A.; Bruining, H.A.; Puppels, G.J. In vivo confocal Raman microspectroscopy of the skin: Noninvasive determination of molecular concentration profiles. J. Investig. Dermatol. 2001, 116, 434–442. [Google Scholar] [CrossRef]
- Haddad, A.; Mohiuddin, S.S. Biochemistry, Citric Acid Cycle; StatPearls: Tampa, FL, USA, 2023. Available online: https://www.ncbi.nlm.nih.gov/books/NBK541072/ (accessed on 11 December 2023).
- Howe, J.; Costantino, R.; Slominski, A. On the putative mechanism of induction and regulation of melanogenesis by L-tyrosine. Acta Derm. Venereol. 1991, 71, 150–152. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Step for Analysis | Porcine | Equine | ||||
|---|---|---|---|---|---|---|
| Nitrogen Concentration (1) (mg/mL) | Solubilized (2) (%) | Step Solubilization (3) (%) | Nitrogen Concentration (1) (mg/mL) | Solubilized (2) (%) | Step Solubilization (2) (%) | |
| 1. Before heat pasteurization | 0.41 | 20.1 | 20.1 | 0.33 | 19.2 | 19.2 |
| 2. After heat pasteurization | 1.23 | 59.6 | 39.5 | 0.69 | 40.1 | 20.9 |
| 3. After fermentation | 1.79 | 87.0 | 27.4 | 1.57 | 91.3 | 51.2 |
| 4. After additional proteolysis and sterilization | 2.06 | 100.0 | 13.0 | 1.72 | 100.0 | 8.7 |
| 5. Without fermentation | 2.28 | 110.6 | - | 2.08 | 120.8 | - |
| Step for Analysis | Porcine | Equine | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| ≤1000 Da (1) (%) | >1000 Da (2) (%) | Total Area (3) | Total area (4) (%) | Step Increase (5) (%) | ≤1000 Da (1) (%) | >1000 Da (2) (%) | Total Area (3) | Total Area (4) (%) | Step Increase (5) (%) | |
| 1. Before heat pasteurization | 54.0 | 46.0 | 86.3 | 22.1 | 22.1 | 33.9 | 65.8 | 373.9 | 20.9 | 20.9 |
| 2. After heat pasteurization | 54.8 | 45.2 | 255.1 | 65.4 | 43.3 | 51.7 | 47.7 | 576.6 | 32.3 | 11.3 |
| 3. After fermentation | 65.8 | 34.2 | 356.6 | 91.4 | 26.0 | 62.5 | 37.0 | 1525.9 | 85.4 | 53.1 |
| 4. After additional proteolysis and sterilization | 74.0 | 26.0 | 390.0 | 100.0 | 8.6 | 67.9 | 31.6 | 1786.2 | 100.0 | 14.6 |
| Compound Name | m/z | MT (min) | Relative Area (4) | Concentration (µM) (5) | Fold Amount in Ferment (6) | ||
|---|---|---|---|---|---|---|---|
| Ferment Filtrate | Proteolysate Filtrate | Ferment Filtrate | Proteolysate Filtrate | ||||
| Lactic acid | 89.024 | 8.70 | 5.0 × 100 | 1.8 × 10−2 | 412,140 | 1452 | 283.8 |
| Ornithine | 133.097 | 5.53 | 6.8 × 10−2 | 8.3 × 10−3 | 1002 | 122 | 8.2 |
| Tyramine | 138.091 | 6.69 | 4.9 × 10−2 | 4.6 × 10−4 | 538 | 5.06 | 106.3 |
| 2-Phenylethylamine | 122.096 | 6.26 | 4.1 × 10−2 | N.D. | - | - | 1< |
| 2-Hydroxy-4-methylvaleric acid (1) 2-Hydroxy-3-methylvaleric acid (1) | 131.071 | 7.33 | 3.3 × 10−2 | 2.2 × 10−4 | - | - | 150.7 |
| Pyruvic acid | 87.009 | 9.90 | 2.0 × 10−2 | 5.8 × 10−4 | 2322 | 68.3 | 34.0 |
| Citrulline | 176.103 | 8.93 | 1.2 × 10−2 | 2.8 × 10−3 | 217 | 51 | 4.2 |
| 3-Amino-2-piperidone | 115.086 | 6.16 | 9.0 × 10−3 | 6.4 × 10−4 | - | - | 14.1 |
| Succinic acid | 117.019 | 13.90 | 4.3 × 10−3 | 1.1 × 10−4 | 292 | 7.26 | 40.2 |
| Hydroxyproline | 132.065 | 9.72 | 3.7 × 10−3 | 3.9 × 10−4 | 61.6 | 6.59 | 9.4 |
| Glycerophosphocholine | 258.110 | 17.38 | 3.4 × 10−3 | 3.9 × 10−4 | - | - | 8.6 |
| β-Hydroxyisovaleric acid (2) 2-Hydroxyvaleric acid (2) 2-Hydroxyisovaleric acid (2) | 117.056 | 7.62 | 2.9 × 10−3 | 2.8 × 10−5 | - | - | 104.7 |
| N5-Ethylglutamine | 175.108 | 9.18 | 2.9 × 10−3 | N.D. | - | - | 1< |
| Propionic acid | 73.029 | 8.83 | 2.4 × 10−3 | N.D. | - | - | 1< |
| Butyric acid Isobutyric acid | 87.045 | 8.04 | 1.0 × 10−3 | N.D. | - | - | 1< |
| Glucose 6-phosphate | 259.023 | 8.02 | 8.6 × 10−4 | N.D. | 68.6 | N.D. | 1< |
| Tropic acid (3) 3-(2-Hydroxyphenyl)propionic acid (3) | 165.056 | 7.19 | 8.0 × 10−4 | 4.3 × 10−5 | - | - | 18.7 |
| Gluconic acid | 195.051 | 6.83 | 5.9 × 10−4 | 1.4 × 10−4 | 40 | 10 | 4.2 |
| N-Acetylserine | 148.060 | 18.56 | 5.4 × 10−4 | 1.0 × 10−4 | - | - | 5.3 |
| N1,N12-Diacetylspermine | 287.244 | 6.10 | 4.4 × 10−4 | 4.9 × 10−6 | - | - | 90.3 |
| N-Acetylhistidine | 198.087 | 7.91 | 4.2 × 10−4 | 1.6 × 10−5 | - | - | 25.7 |
| N-Lactoylphenylalanine | 236.094 | 6.56 | 3.7 × 10−4 | N.D. | - | - | 1< |
| N-Acetylaspartic acid | 174.041 | 10.92 | 3.6 × 10−4 | 2.4 × 10−5 | - | - | 15.2 |
| AMP | 346.056 | 7.62 | 3.4 × 10−4 | 1.7 × 10−5 | 22.3 | 1.12 | 19.8 |
| UMP | 323.030 | 7.97 | 3.2 × 10−4 | 2.8 × 10−5 | 23.6 | 2.05 | 11.5 |
| Terephthalic acid | 165.019 | 12.00 | 3.0 × 10−4 | 2.1 × 10−5 | - | - | 14.3 |
| O-Succinylhomoserine | 218.067 | 9.53 | 2.8 × 10−4 | N.D. | - | - | 1< |
| Ethanolamine phosphate | 140.012 | 6.73 | 2.7 × 10−4 | 5.9 × 10−6 | - | - | 46.1 |
| N1,N8-Diacetylspermidine | 230.186 | 8.23 | 2.4 × 10−4 | 5.1 × 10−5 | - | - | 4.7 |
| Homoserine | 120.066 | 8.15 | 2.3 × 10−4 | 3.3 × 10−5 | 3.85 | 0.551 | 7.0 |
| GMP | 362.050 | 7.53 | 2.3 × 10−4 | N.D. | 20.3 | N.D. | 1< |
| N-Acetylputrescine | 131.118 | 6.83 | 2.2 × 10−4 | 2.5 × 10−5 | - | - | 8.8 |
| Spermidine | 146.165 | 3.70 | 2.2 × 10−4 | N.D. | 2.70 | N.D. | 1< |
| Fructose 6-phosphate | 259.023 | 8.11 | 2.2 × 10−4 | N.D. | 15.7 | N.D. | 1< |
| CMP | 322.045 | 7.84 | 2.0 × 10−4 | 2.7 × 10−5 | 21.5 | 2.92 | 7.4 |
| N-Acetylthreonine | 160.062 | 7.02 | 2.0 × 10−4 | 3.1 × 10−5 | - | - | 6.4 |
| N1-Acetylspermine | 245.233 | 4.54 | 2.0 × 10−4 | N.D. | - | - | 1< |
| Nicotinamide | 123.055 | 6.03 | 1.9 × 10−4 | 1.9 × 10−5 | - | - | 9.9 |
| Homocystine | 269.062 | 7.80 | 1.2 × 10−4 | N.D. | - | - | 1< |
| Mevalonic acid | 147.066 | 7.16 | 1.0 × 10−4 | 7.6 × 10−6 | - | - | 13.6 |
| Compound Name | m/z | MT (min) | Relative Area (4) | Concentration (µM) (5) | Fold Amount in Ferment (6) | ||
|---|---|---|---|---|---|---|---|
| Ferment Filtrate | Proteolysate Filtrate | Ferment Filtrate | Proteolysate Filtrate | ||||
| Lactic acid | 89.024 | 8.70 | 2.3 × 100 | 8.0 × 10−3 | 188,642 | 651 | 289.8 |
| Ornithine | 133.097 | 5.53 | 4.3 × 10−2 | 8.1 × 10−4 | 637 | 12.0 | 52.9 |
| Tyramine | 138.091 | 6.69 | 3.5 × 10−2 | 1.2 × 10−3 | 388 | 12.9 | 30.2 |
| 2-Phenylethylamine | 122.096 | 6.26 | 3.1 × 10−2 | N.D. | - | - | 1< |
| Pyruvic acid | 87.009 | 9.90 | 2.1 × 10−2 | 3.4 × 10−4 | 2454 | 40.3 | 61.0 |
| 2-Hydroxy-4-methylvaleric acid (1) 2-Hydroxy-3-methylvaleric acid (1) | 131.071 | 7.33 | 1.5 × 10−2 | 1.3 × 10−5 | - | - | 1114.1 |
| 3-Amino-2-piperidone | 115.086 | 6.16 | 7.8 × 10−3 | 1.4 × 10−4 | - | - | 53.9 |
| Citrulline | 176.103 | 8.93 | 7.6 × 10−3 | 4.0 × 10−4 | 138 | 7.28 | 18.9 |
| Hydroxyproline | 132.065 | 9.72 | 3.6 × 10−3 | 5.9 × 10−4 | 60.1 | 9.8 | 6.1 |
| Glyceric acid | 105.019 | 8.37 | 2.8 × 10−3 | 9.3 × 10−5 | - | - | 30.5 |
| N5-Ethylglutamine | 175.108 | 9.18 | 2.8 × 10−3 | N.D. | - | - | 1< |
| β-Hydroxyisovaleric acid (2) 2-Hydroxyvaleric acid (2) 2-Hydroxyisovaleric acid (2) | 117.056 | 7.62 | 2.4 × 10−3 | 7.8 × 10−6 | - | - | 312.2 |
| Succinic acid | 117.019 | 13.90 | 1.2 × 10−3 | N.D. | 79.8 | N.D. | 1< |
| Threonic acid | 135.030 | 7.62 | 1.0 × 10−3 | 1.8 × 10−4 | - | - | 5.4 |
| Gluconic acid | 195.051 | 6.83 | 8.5 × 10−4 | 7.4 × 10−5 | 58.4 | 5.10 | 11.5 |
| Tropic acid3-(2-Hydroxyphenyl)propionic acid | 165.056 | 7.19 | 7.9 × 10−4 | 4.0 × 10−5 | - | - | 19.7 |
| N-Acetylserine | 148.060 | 18.56 | 7.9 × 10−4 | 6.6 × 10−5 | - | - | 12.0 |
| Mevalonic acid | 147.066 | 7.16 | 5.7 × 10−4 | N.D. | - | - | 1< |
| Mevalolactone | 131.070 | 17.84 | 5.1 × 10−4 | N.D. | - | - | 1< |
| Gluconolactone | 179.055 | 18.36 | 4.0 × 10−4 | 4.6 × 10−5 | - | - | 8.8 |
| Terephthalic acid | 165.019 | 12.00 | 3.4 × 10−4 | 2.4 × 10−5 | - | - | 13.8 |
| 3,4-Dihydroxyhydrocinnamic acid (3) Homovanillic acid (3) Hydroxyphenyllactic acid (3) | 181.051 | 6.93 | 3.3 × 10−4 | 1.9 × 10−5 | - | - | 17.8 |
| N-Acetylaspartic acid | 174.041 | 10.92 | 3.0 × 10−4 | N.D. | - | - | 1< |
| N1,N12-Diacetylspermine | 287.244 | 6.10 | 3.0 × 10−4 | N.D. | - | - | 1< |
| Glucose 6-phosphate | 259.023 | 8.02 | 3.0 × 10−4 | 8.8 × 10−6 | 23.8 | 0.70 | 33.9 |
| UMP | 323.030 | 7.97 | 2.6 × 10−4 | 1.8 × 10−5 | 19.1 | 1.31 | 14.6 |
| N-Acetylthreonine | 160.062 | 7.02 | 1.8 × 10−4 | 1.1 × 10−5 | - | - | 15.6 |
| AMP | 346.056 | 7.62 | 1.8 × 10−4 | 1.1 × 10−5 | 11.6 | 0.72 | 16.1 |
| Homoserine | 120.066 | 8.15 | 1.6 × 10−4 | N.D. | 2.64 | N.D. | 1< |
| N-Acetylhistidine | 198.087 | 7.91 | 1.6 × 10−4 | N.D. | - | - | 1< |
| Homocystine | 269.062 | 7.80 | 1.4 × 10−4 | N.D. | - | - | 1< |
| 2-Aminobutyric acid | 104.071 | 7.79 | 1.4 × 10−4 | N.D. | - | - | 1< |
| N-Acetylputrescine | 131.118 | 6.83 | 1.3 × 10−4 | 2.1 × 10−5 | - | - | 6.6 |
| Argininic acid | 176.103 | 7.39 | 1.3 × 10−4 | N.D. | - | - | 1< |
| N-Lactoylphenylalanine | 236.094 | 6.56 | 1.3 × 10−4 | 6.6 × 10−6 | - | - | 19.4 |
| Compound Name | m/z | MT (min) | Relative Area (1) | Concentration (µM) (2) | Fold Amount in Ferment (3) | ||
|---|---|---|---|---|---|---|---|
| Ferment Filtrate | Proteolysate Filtrate | Ferment Filtrate | Proteolysate Filtrate | ||||
| Arg | 175.119 | 5.77 | 2.5 × 10−2 | 1.0 × 10−1 | 361 | 1455 | 0.248 |
| Tyr | 182.081 | 9.16 | 6.2 × 10−3 | 4.7 × 10−2 | 107 | 808 | 0.133 |
| Guanosine | 284.099 | 10.22 | 5.8 × 10−4 | 7.6 × 10−3 | 11.3 | 148 | 0.076 |
| Cytidine | 244.093 | 7.80 | 5.3 × 10−4 | 5.5 × 10−3 | 10.2 | 106 | 0.096 |
| Inosine | 269.088 | 15.75 | 1.2 × 10−4 | 3.6 × 10−3 | 2.99 | 88.3 | 0.034 |
| Thymidine | 243.098 | 17.79 | 1.6 × 10−4 | 2.1 × 10−3 | 7.73 | 97.9 | 0.079 |
| 2′-Deoxycytidine | 228.098 | 7.61 | 6.1 × 10−5 | 2.0 × 10−3 | - | - | 0.031 |
| Uridine | 245.078 | 17.77 | 4.0 × 10−4 | 2.0 × 10−3 | 19.9 | 98.4 | 0.202 |
| 2′-Deoxyguanosine | 268.104 | 9.37 | 6.9 × 10−5 | 1.2 × 10−3 | - | - | 0.056 |
| Cystine | 241.031 | 8.90 | 1.6 × 10−4 | 9.5 × 10−4 | - | - | 0.165 |
| 2′-Deoxyinosine | 253.094 | 14.68 | N.D. | 1.7 × 10−4 | - | - | <1 |
| 3′-CMP | 322.045 | 7.98 | N.D. | 1.3 × 10−4 | - | - | <1 |
| 5-Methyl-2′-deoxycytidine | 242.113 | 7.86 | N.D. | 1.3 × 10−4 | - | - | <1 |
| Compound Name | m/z | MT (min) | Relative Area (1) | Concentration (µM) (2) | Fold Amount in Ferment (3) | ||
|---|---|---|---|---|---|---|---|
| Ferment Filtrate | Proteolysate Filtrate | Ferment Filtrate | Proteolysate Filtrate | ||||
| Arg | 175.119 | 5.77 | 1.4 × 10−2 | 6.0 × 10−2 | 195 | 866 | 0.225 |
| Tyr | 182.081 | 9.16 | 1.1 × 10−3 | 1.5 × 10−2 | 19.5 | 260 | 0.075 |
| Guanosine | 284.099 | 10.22 | 1.1 × 10−4 | 6.3 × 10−4 | 2.11 | 12.3 | 0.172 |
| Cystine | 241.031 | 8.90 | 5.0 × 10−5 | 2.2 × 10−4 | - | - | 0.224 |
| Sample | EMC (%) |
|---|---|
| Porcine placenta ferment filtrate | 80.3 ± 2.1 |
| Equine placenta ferment filtrate | 79.3 ± 3.0 |
| Porcine placenta proteolysate filtrate | 34.5 ± 1.9 |
| Equine placenta proteolysate filtrate | 38.0 ± 0.9 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Matsunaga, K.; Komatsu, Y. Novel Placenta-Derived Liquid Product Suitable for Cosmetic Application Produced by Fermentation and Digestion of Porcine or Equine Placenta Using Lactic Acid Bacterium Enterococcus faecalis PR31. Fermentation 2024, 10, 89. https://doi.org/10.3390/fermentation10020089
Matsunaga K, Komatsu Y. Novel Placenta-Derived Liquid Product Suitable for Cosmetic Application Produced by Fermentation and Digestion of Porcine or Equine Placenta Using Lactic Acid Bacterium Enterococcus faecalis PR31. Fermentation. 2024; 10(2):89. https://doi.org/10.3390/fermentation10020089
Chicago/Turabian StyleMatsunaga, Kanako, and Yasuhiko Komatsu. 2024. "Novel Placenta-Derived Liquid Product Suitable for Cosmetic Application Produced by Fermentation and Digestion of Porcine or Equine Placenta Using Lactic Acid Bacterium Enterococcus faecalis PR31" Fermentation 10, no. 2: 89. https://doi.org/10.3390/fermentation10020089
APA StyleMatsunaga, K., & Komatsu, Y. (2024). Novel Placenta-Derived Liquid Product Suitable for Cosmetic Application Produced by Fermentation and Digestion of Porcine or Equine Placenta Using Lactic Acid Bacterium Enterococcus faecalis PR31. Fermentation, 10(2), 89. https://doi.org/10.3390/fermentation10020089