
Mycoremediation of Synthetic Azo Dyes by White-Rot Fungi Grown on Diary Waste: A Step toward Sustainable and Circular Bioeconomy

 ,
,  ,
,  ,
, 

Abstract
1. Introduction
2. Materials and Methods
2.1. Materials, Microrganisms, and Reagents
2.2. Inoculum Preparation
2.3. Decolorization of Artificial Dye-Baths by White-Rot Fungi
2.4. Lignin Degradation Capacity
2.5. Lignin Analysis
2.6. Decoloration and Lignin Degradation of Pulp and Paper Mill Effluent: Preliminary Tests
2.7. Visible Decolorization Efficiency
3. Results
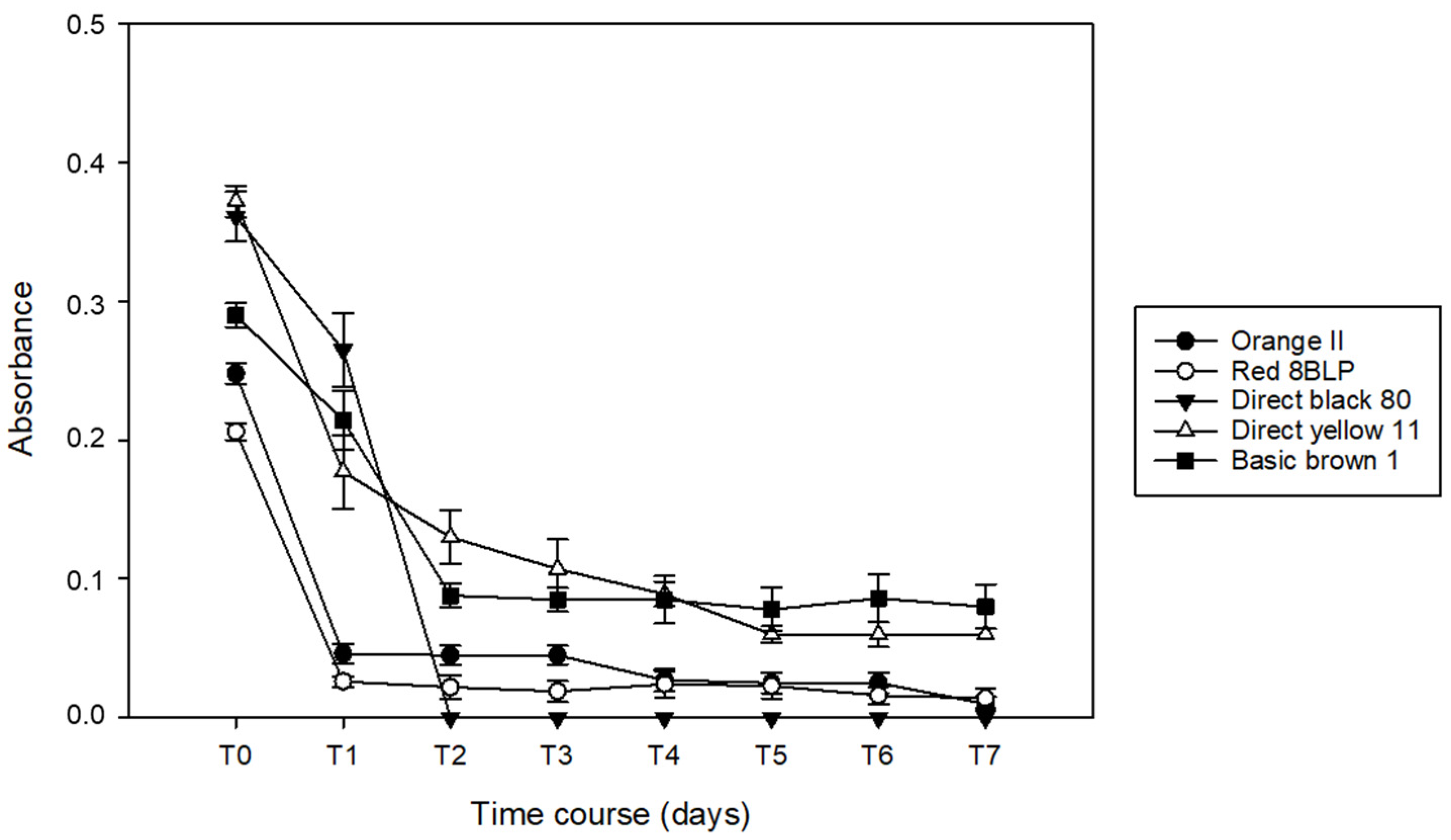
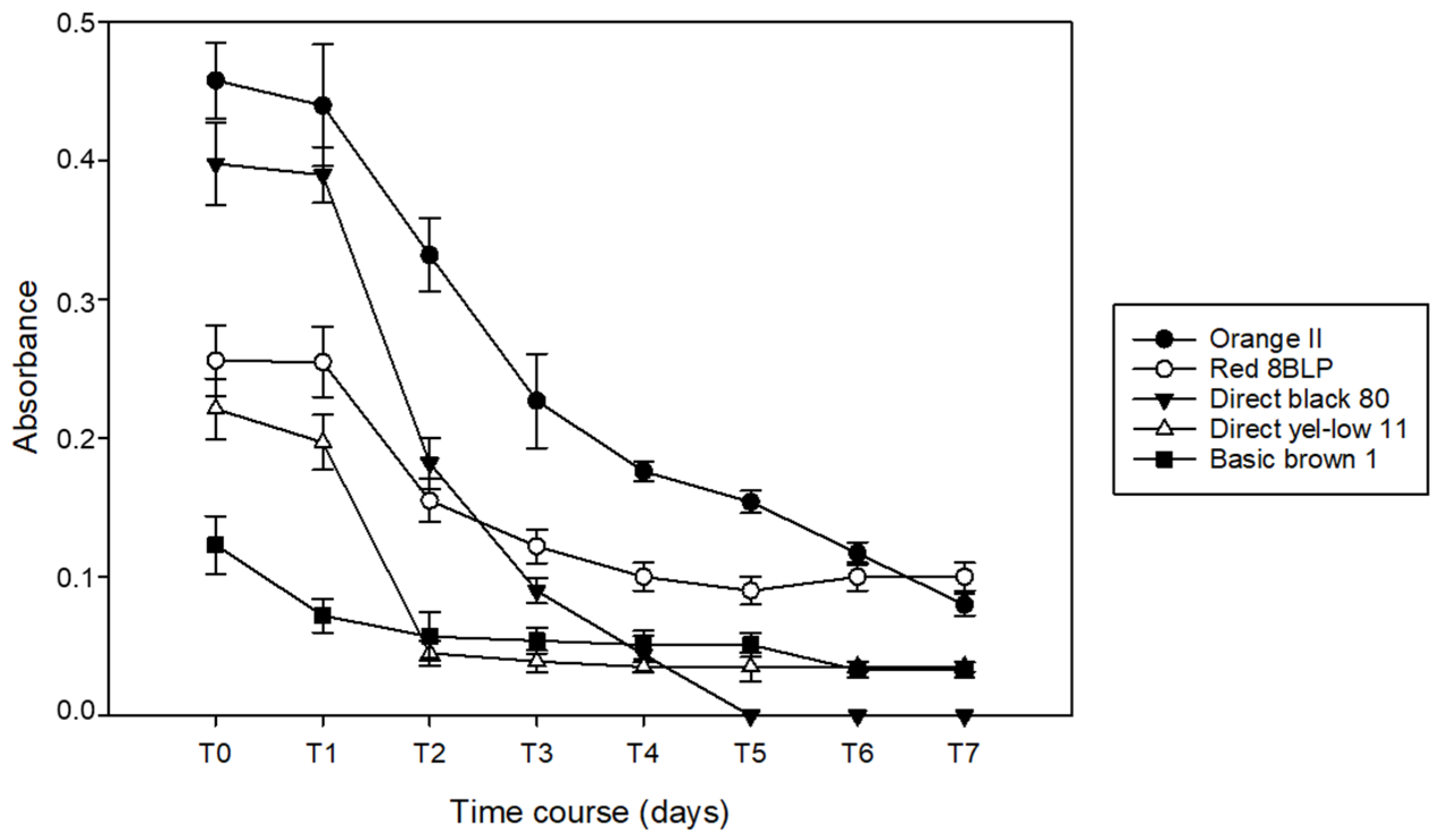
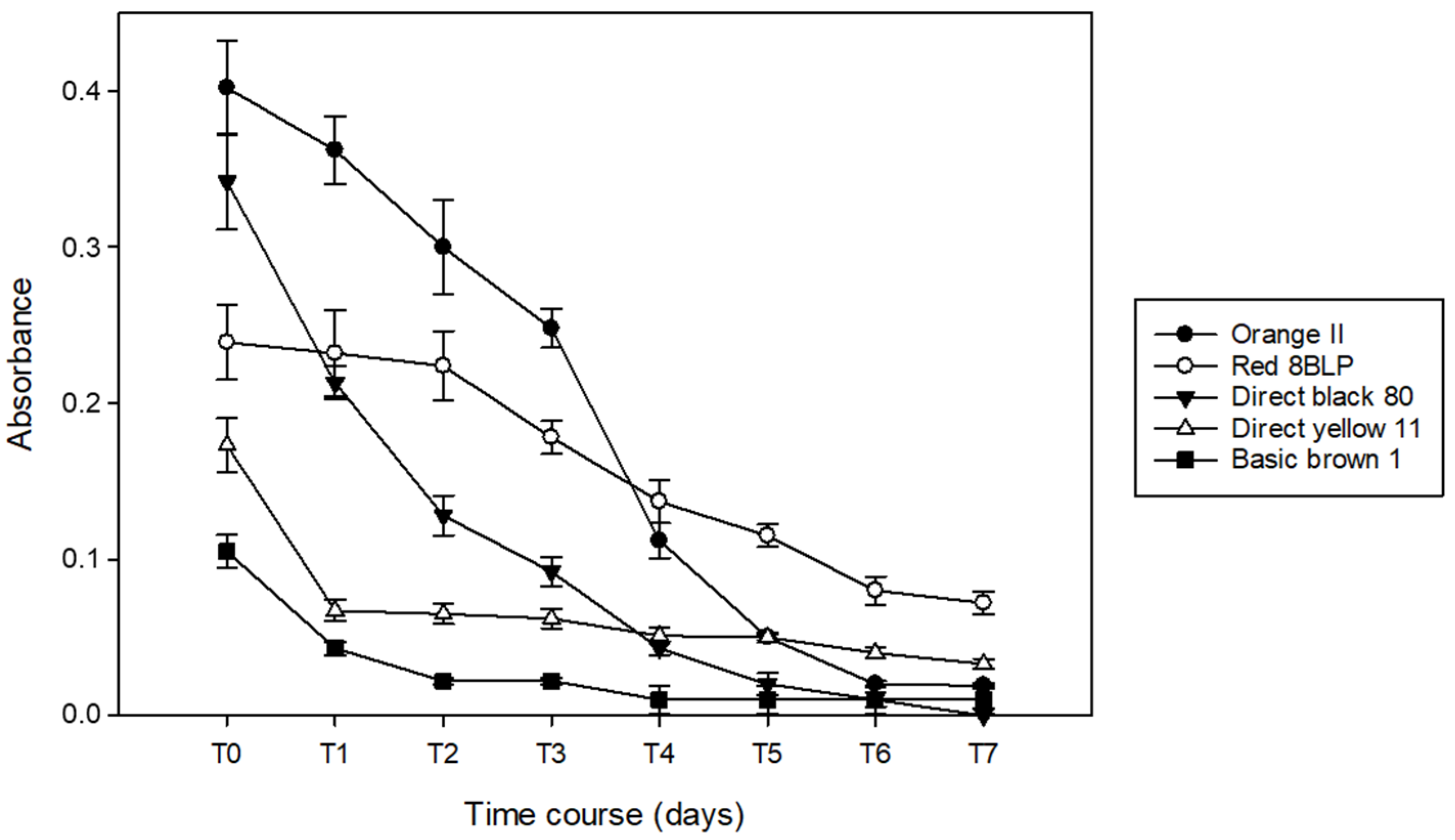
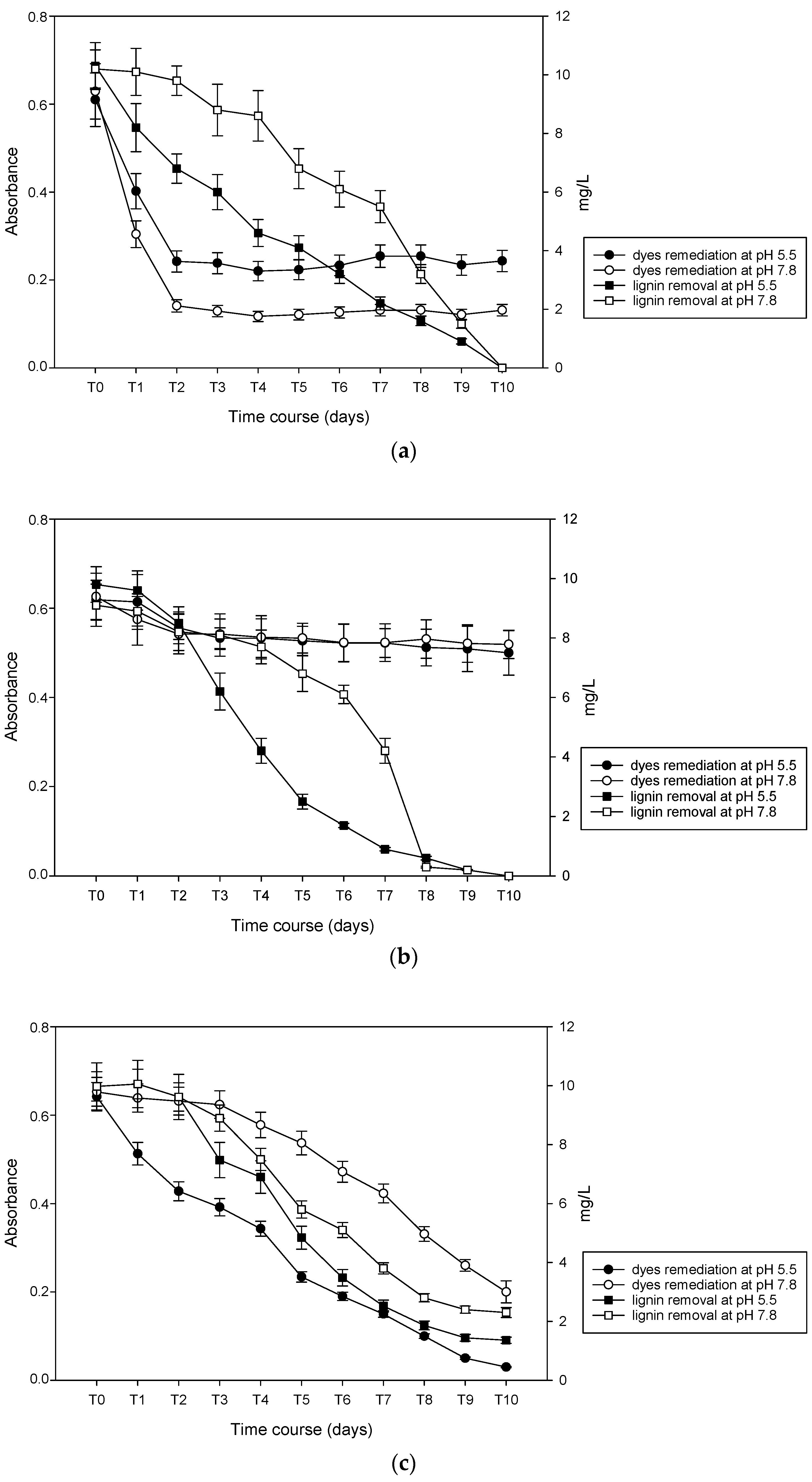
3.1. Decoloration of Artificial Dye-Baths with B. adusta
3.2. Decoloration of Artificial Dye-Baths with P. chrysosporium
3.3. Decoloration of Artifical Dye-Baths with T. versicolor
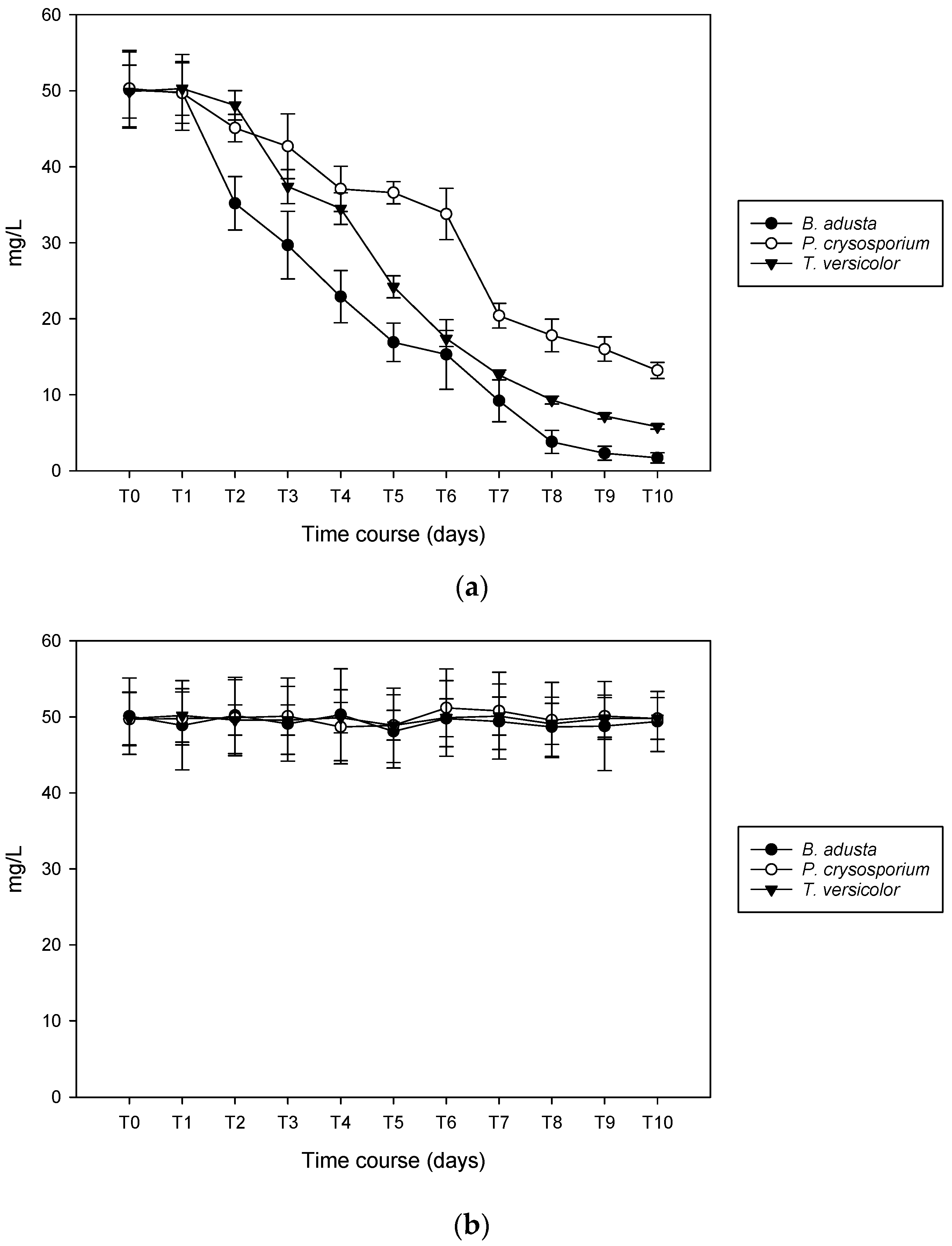
3.4. Lignin Degradation Capacity
3.5. Mycoremediation and Lignin Removal of Colored Pulp and Paper Mill Effluent: Preliminary Test
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Slama, H.B.; Chenari Bouket, A.; Pourhassan, Z.; Alenezi, F.N.; Silini, A.; Cherif-Silini, H.; Oszako, T.; Luptakova, L.; Golińska, P.; Belbahri, L. Diversity of Synthetic Dyes from Textile Industries, Discharge Impacts and Treatment Methods. Appl. Sci. 2021, 11, 6255. [Google Scholar] [CrossRef]
- Gürses, A.; Açıkyıldız, M.; Güneş, K.; Gürses, M.S. Classification of Dye and Pigments. In Dyes and Pigments; Gürses, A., Açıkyıldız, M., Güneş, K., Gürses, M.S., Eds.; SpringerBriefs in Molecular Science; Springer International Publishing: Cham, Switzerland, 2016; pp. 31–45. ISBN 978-3-319-33892-7. [Google Scholar]
- Tamburini, D.; Breitung, E.; Mori, C.; Kotajima, T.; Clarke, M.L.; McCarthy, B. Exploring the Transition from Natural to Synthetic Dyes in the Production of 19th-Century Central Asian Ikat Textiles. Herit. Sci. 2020, 8, 114. [Google Scholar] [CrossRef]
- Akansha, K.; Kaur, T.; Yadav, A.; Kour, D.; Rai, A.K.; Singh, S.; Mishra, S.; Kumar, L.; Miglani, K.; Singh, K.; et al. Microbe-Mediated Remediation of Dyes: Current Status and Future Challenges. J. Appl. Biol. Biotech 2023, 11, 1–23. [Google Scholar] [CrossRef]
- Omar, A.Z.; El-Rahman, M.A.; Hamed, E.A.; El-Sadany, S.K.; El-atawy, M.A. Synthesis, Spectroscopic Characterization and Dyeing Performance of Novel Bis Azo Dyes Derived from Benzidine. Sci. Rep. 2023, 13, 7826. [Google Scholar] [CrossRef]
- Barciela, P.; Perez-Vazquez, A.; Prieto, M.A. Azo Dyes in the Food Industry: Features, Classification, Toxicity, Alternatives, and Regulation. Food Chem. Toxicol. 2023, 178, 113935. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.; Yang, Z.; Xing, L.; Zhang, X.; Li, X.; Zhang, D. Recent Advances in the Biodegradation of Azo Dyes. World J. Microbiol. Biotechnol. 2021, 37, 137. [Google Scholar] [CrossRef]
- Hashemi, S.H.; Kaykhaii, M. Chapter 15-Azo Dyes: Sources, Occurrence, Toxicity, Sampling, Analysis, and Their Removal Methods. In Emerging Freshwater Pollutants; Dalu, T., Tavengwa, N.T., Eds.; Elsevier: Amsterdam, The Netherlands, 2022; pp. 267–287. ISBN 978-0-12-822850-0. [Google Scholar]
- Chung, K.-T. Azo Dyes and Human Health: A Review. J. Environ. Sci. Health Part C 2016, 34, 233–261. [Google Scholar] [CrossRef]
- Waghmode, T.R.; Kurade, M.B.; Sapkal, R.T.; Bhosale, C.H.; Jeon, B.-H.; Govindwar, S.P. Sequential Photocatalysis and Biological Treatment for the Enhanced Degradation of the Persistent Azo Dye Methyl Red. J. Hazard. Mater. 2019, 371, 115–122. [Google Scholar] [CrossRef]
- Islam, T.; Repon, M.R.; Islam, T.; Sarwar, Z.; Rahman, M.M. Impact of Textile Dyes on Health and Ecosystem: A Review of Structure, Causes, and Potential Solutions. Environ. Sci. Pollut. Res. 2023, 30, 9207–9242. [Google Scholar] [CrossRef]
- Samchetshabam, G.; Hussan, A.; Gon Choudhury, T.; Gita, S.; Soholars, P.; Hussan, A. Impact of Textile Dyes Waste on Aquatic Environments and Its Treatment. Environ. Ecol. 2017, 35, 2349–2353. [Google Scholar]
- Saha, J.K.; Selladurai, R.; Coumar, M.V.; Dotaniya, M.L.; Kundu, S.; Patra, A.K. Impact of Different Developmental Projects on Soil Fertility. In Soil Pollution-An Emerging Threat to Agriculture; Saha, J.K., Selladurai, R., Coumar, M.V., Dotaniya, M.L., Kundu, S., Patra, A.K., Eds.; Environmental Chemistry for a Sustainable World; Springer: Singapore, 2017; pp. 251–269. ISBN 978-981-10-4274-4. [Google Scholar]
- Kumari, R.; Sunil, D.; Ningthoujam, R.S.; Kumar, N.A. Azodyes as Markers for Tumor Hypoxia Imaging and Therapy: An up-to-Date Review. Chem.-Biol. Interact. 2019, 307, 91–104. [Google Scholar] [CrossRef]
- Siddiqui, S.I.; Allehyani, E.S.; Al-Harbi, S.A.; Hasan, Z.; Abomuti, M.A.; Rajor, H.K.; Oh, S. Investigation of Congo Red Toxicity towards Different Living Organisms: A Review. Processes 2023, 11, 807. [Google Scholar] [CrossRef]
- Donkadokula, N.Y.; Kola, A.K.; Naz, I.; Saroj, D. A Review on Advanced Physico-Chemical and Biological Textile Dye Wastewater Treatment Techniques. Rev. Environ. Sci. Biotechnol. 2020, 19, 543–560. [Google Scholar] [CrossRef]
- Vishnu, D.; Dhandapani, B.; Authilingam, S.; Sivakumar, S.V. A Comprehensive Review of Effective Adsorbents Used for the Removal of Dyes from Wastewater. Curr. Anal. Chem. 2022, 18, 255–268. [Google Scholar] [CrossRef]
- Asaithambi, P.; Yesuf, M.B.; Govindarajan, R.; Hariharan, N.M.; Thangavelu, P.; Alemayehu, E. A Review of Hybrid Process Development Based on Electrochemical and Advanced Oxidation Processes for the Treatment of Industrial Wastewater. Int. J. Chem. Eng. 2022, 2022, e1105376. [Google Scholar] [CrossRef]
- Piaskowski, K.; Świderska-Dąbrowska, R.; Zarzycki, P.K. Dye Removal from Water and Wastewater Using Various Physical, Chemical, and Biological Processes. J. AOAC Int. 2018, 101, 1371–1384. [Google Scholar] [CrossRef]
- Katheresan, V.; Kansedo, J.; Lau, S.Y. Efficiency of Various Recent Wastewater Dye Removal Methods: A Review. J. Environ. Chem. Eng. 2018, 6, 4676–4697. [Google Scholar] [CrossRef]
- Takahashi, H.; Hashimoto, Y. Formaldehyde-Mediated Modification of Natural Deoxyguanosine with Amines: One-Pot Cyclization as a Molecular Model for Genotoxicity. Bioorg. Med. Chem. Lett. 2001, 11, 729–731. [Google Scholar] [CrossRef]
- Covino, S.; Stella, T.; Cajthaml, T. Mycoremediation of Organic Pollutants: Principles, Opportunities, and Pitfalls. In Fungal Applications in Sustainable Environmental Biotechnology; Purchase, D., Ed.; Fungal Biology; Springer International Publishing: Cham, Switzerland, 2016; pp. 185–231. ISBN 978-3-319-42852-9. [Google Scholar]
- Ajaz, M.; Shakeel, S.; Rehman, A. Microbial Use for Azo Dye Degradation—A Strategy for Dye Bioremediation. Int. Microbiol. 2020, 23, 149–159. [Google Scholar] [CrossRef]
- Pinheiro, L.R.S.; Gradíssimo, D.G.; Xavier, L.P.; Santos, A.V. Degradation of Azo Dyes: Bacterial Potential for Bioremediation. Sustainability 2022, 14, 1510. [Google Scholar] [CrossRef]
- Noman, E.; Al-Gheethi, A.; Mohamed, R.M.S.R.; Talip, B.A. Myco-Remediation of Xenobiotic Organic Compounds for a Sustainable Environment: A Critical Review. Top. Curr. Chem. 2019, 377, 17. [Google Scholar] [CrossRef]
- Rajhans, G.; Barik, A.; Sen, S.K.; Raut, S. Degradation of Dyes by Fungi: An Insight into Mycoremediation. BioTechnologia 2021, 102, 445–455. [Google Scholar] [CrossRef]
- Zhuo, R.; Fan, F. A Comprehensive Insight into the Application of White Rot Fungi and Their Lignocellulolytic Enzymes in the Removal of Organic Pollutants. Sci. Total Environ. 2021, 778, 146132. [Google Scholar] [CrossRef]
- Chen, S.; Zhu, M.; Guo, X.; Yang, B.; Zhuo, R. Coupling of Fenton Reaction and White Rot Fungi for the Degradation of Organic Pollutants. Ecotoxicol. Environ. Saf. 2023, 254, 114697. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Zhang, X.; Zhang, M.; Zhu, Y.; Zhuo, R. Removal of Heavy-Metal Pollutants by White Rot Fungi: Mechanisms, Achievements, and Perspectives. J. Clean. Prod. 2022, 354, 131681. [Google Scholar] [CrossRef]
- Costa, S.; Dedola, D.G.; Pellizzari, S.; Blo, R.; Rugiero, I.; Pedrini, P.; Tamburini, E. Lignin Biodegradation in Pulp-and-Paper Mill Wastewater by Selected White Rot Fungi. Water 2017, 9, 935. [Google Scholar] [CrossRef]
- Schwanninger, M.; Hinterstoisser, B. Klason Lignin: Modifications to Improve the Precision of the Standardized Determination. Holzforschung 2002, 56, 161–166. [Google Scholar] [CrossRef]
- López, M.J.; Guisado, G.; Vargas-García, M.C.; Suárez-Estrella, F.; Moreno, J. Decolorization of Industrial Dyes by Ligninolytic Microorganisms Isolated from Composting Environment. Enzym. Microb. Technol. 2006, 40, 42–45. [Google Scholar] [CrossRef]
- Sanz, J.L.; Köchling, T. Next-Generation Sequencing and Waste/Wastewater Treatment: A Comprehensive Overview. Rev. Environ. Sci. Bio/Technol. 2019, 18, 635–680. [Google Scholar] [CrossRef]
- FAOSTAT: Forestry Production and Trade-Google Scholar. Available online: https://scholar.google.com/scholar_lookup?title=FAOSTAT%20-%20Forestry%20Production%20and%20Trade&publication_year=2021&author=FAO%20(Food%20and%20Agriculture%20Organization%20of%20the%20United%20Nations (accessed on 3 November 2023).
- Echezonachi, S.O. Chapter 16-The Role of White Rot Fungi in Bioremediation. In Microbes and Microbial Biotechnology for Green Remediation; Malik, J.A., Ed.; Elsevier: Amsterdam, The Netherlands, 2022; pp. 305–320. ISBN 978-0-323-90452-0. [Google Scholar]
- Kumar, A.; Chandra, R. Ligninolytic Enzymes and Its Mechanisms for Degradation of Lignocellulosic Waste in Environment. Heliyon 2020, 6, e03170. [Google Scholar] [CrossRef]
- Sun, S.; Liu, P.; Ullah, M. Efficient Azo Dye Biodecolorization System Using Lignin-Co-Cultured White-Rot Fungus. J. Fungi 2023, 9, 91. [Google Scholar] [CrossRef] [PubMed]
- Cui, M.-H.; Liu, W.-Z.; Tang, Z.-E.; Cui, D. Recent Advancements in Azo Dye Decolorization in Bio-Electrochemical Systems (BESs): Insights into Decolorization Mechanism and Practical Application. Water Res. 2021, 203, 117512. [Google Scholar] [CrossRef]
- Moreira, P.R.; Almeida-Vara, E.; Sena-Martins, G.; Polónia, I.; Xavier Malcata, F.; Cardoso Duarte, J. Decolourisation of Remazol Brilliant Blue R via a Novel Bjerkandera Sp. Strain. J. Biotechnol. 2001, 89, 107–111. [Google Scholar] [CrossRef] [PubMed]
- Choi, Y.-S.; Seo, J.-Y.; Lee, H.; Yoo, J.; Jung, J.; Kim, J.-J.; Kim, G.-H. Decolorization and Detoxification of Wastewater Containing Industrial Dyes by Bjerkandera adusta KUC9065. Water Air Soil. Pollut. 2013, 225, 1801. [Google Scholar] [CrossRef]
- Riegas-Villalobos, A.; Martínez-Morales, F.; Tinoco-Valencia, R.; Serrano-Carreón, L.; Bertrand, B.; Trejo-Hernández, M.R. Efficient Removal of Azo-Dye Orange II by Fungal Biomass Absorption and Laccase Enzymatic Treatment. 3 Biotech 2020, 10, 146. [Google Scholar] [CrossRef]
- Feijoo, G.; Moreira, M.T.; Roca, E.; Lema, J.M. Use of Cheese Whey as a Substrate to Produce Manganese Peroxidase by Bjerkandera Sp BOS55. J. Ind. Microbiol. Biotech 1999, 23, 86–90. [Google Scholar] [CrossRef]
- Nawaz, M.; Gunasekaran, M. Effect of Peat Extract on the Hydrolytic Enzymes of Phanerochaete chrysosporium. Resour. Conserv. Recycl. 1988, 1, 197–205. [Google Scholar] [CrossRef]
- Singh, G.; Dwivedi, S.K.; Mishra, J. Role of Fungal Enzymes in the Removal of Azo Dyes. In Microbial Enzymes: Roles and Applications in Industries; Arora, N.K., Mishra, J., Mishra, V., Eds.; Microorganisms for Sustainability; Springer: Singapore, 2020; pp. 231–257. ISBN 9789811517105. [Google Scholar]
- Qi-he, C.; Krügener, S.; Hirth, T.; Rupp, S.; Zibek, S. Co-Cultured Production of Lignin-Modifying Enzymes with White-Rot Fungi. Appl. Biochem. Biotechnol. 2011, 165, 700–718. [Google Scholar] [CrossRef]
- Fackler, K.; Gradinger, C.; Hinterstoisser, B.; Messner, K.; Schwanninger, M. Lignin Degradation by White Rot Fungi on Spruce Wood Shavings during Short-Time Solid-State Fermentations Monitored by near Infrared Spectroscopy. Enzym. Microb. Technol. 2006, 39, 1476–1483. [Google Scholar] [CrossRef]
- Korniłłowicz-Kowalska, T.; Rybczyńska-Tkaczyk, K. Growth Conditions, Physiological Properties, and Selection of Optimal Parameters of Biodegradation of Anticancer Drug Daunomycin in Industrial Effluents by Bjerkandera adusta CCBAS930. Int. Microbiol. 2020, 23, 287–301. [Google Scholar] [CrossRef] [PubMed]
- Madhushika, H.G.; Ariyadasa, T.U.; Gunawardena, S.H.P. Textile Dye Removal from Industrial Wastewater by Biological Methods and Impact on Environment. In Environmental Degradation: Challenges and Strategies for Mitigation; Singh, V.P., Yadav, S., Yadav, K.K., Yadava, R.N., Eds.; Water Science and Technology Library; Springer International Publishing: Cham, Switzerland, 2022; pp. 181–210. ISBN 978-3-030-95542-7. [Google Scholar]
- Wang, Y.; Chen, S.; Zhou, J.; Fan, X.; He, L.; Fan, G. Enhanced Degradation Capability of White-Rot Fungi after Short-Term Pre-Exposure to Silver Ion: Performance and Selectively Antimicrobial Mechanisms. Sci. Total Environ. 2022, 818, 151672. [Google Scholar] [CrossRef] [PubMed]
- Sen, S.K.; Raut, S.; Bandyopadhyay, P.; Raut, S. Fungal Decolouration and Degradation of Azo Dyes: A Review. Fungal Biol. Rev. 2016, 30, 112–133. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Dye | Molecular Structure | IUPAC Name | Molecular Weight | Dye Classification | Maximum Absorption Wavelength |
|---|---|---|---|---|---|
| Orange II | 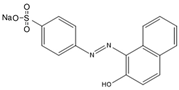 | 4-(2-Hydroxy-1-naphthylazo) benzene sulfonic acid sodium salt | 350.32 | Single azo class | 412 nm |
| Red 8BLP | 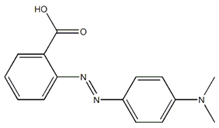 | 2-[[4-(dimethylamino)phenyl]diazenyl]benzoic acid | 269.29 | Single azo class | 422 nm |
| Direct yellow 11 | 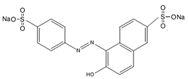 | 6-Hydroxy-5-((4-sulphophenyl) azo) naphthalene-2-sulphonic acid | 408.4 | Single azo class | 411 nm |
| Basic brown 1 | 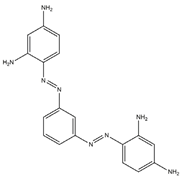 | 4,4′-1,3-Phenylenebis (azo)-bisbenzene-1,3-diamine | 346.38 | Double azo class | 461 nm |
| Direct black 80 | 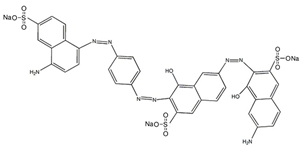 | 1,3,6-Naphthalenetrisulfonic acid, 7-((2-((aminocarbonyl) amino)-4-((4-fluoro-6-((2-methylphenyl) amino)-1,3,5-triazin-2-yl) amino)phenyl)azo)-trisodium salt | 908.78 | Tris azo class | 599 nm |
| White Rot Fungi | Orange II | Red 8BLP | Direct Yellow 11 | Basic Brown 1 | Direct Black 80 | AIR | ASL |
|---|---|---|---|---|---|---|---|
| B. adusta | C | C | NC | NC | C | C | - |
| P. chrysosporium | C | NC | NC | NC | NC | NC | - |
| T. versicolor | C | NC | NC | NC | C | NC | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gugel, I.; Summa, D.; Costa, S.; Manfredini, S.; Vertuani, S.; Marchetti, F.; Tamburini, E. Mycoremediation of Synthetic Azo Dyes by White-Rot Fungi Grown on Diary Waste: A Step toward Sustainable and Circular Bioeconomy. Fermentation 2024, 10, 80. https://doi.org/10.3390/fermentation10020080
Gugel I, Summa D, Costa S, Manfredini S, Vertuani S, Marchetti F, Tamburini E. Mycoremediation of Synthetic Azo Dyes by White-Rot Fungi Grown on Diary Waste: A Step toward Sustainable and Circular Bioeconomy. Fermentation. 2024; 10(2):80. https://doi.org/10.3390/fermentation10020080
Chicago/Turabian StyleGugel, Irene, Daniela Summa, Stefania Costa, Stefano Manfredini, Silvia Vertuani, Filippo Marchetti, and Elena Tamburini. 2024. "Mycoremediation of Synthetic Azo Dyes by White-Rot Fungi Grown on Diary Waste: A Step toward Sustainable and Circular Bioeconomy" Fermentation 10, no. 2: 80. https://doi.org/10.3390/fermentation10020080
APA StyleGugel, I., Summa, D., Costa, S., Manfredini, S., Vertuani, S., Marchetti, F., & Tamburini, E. (2024). Mycoremediation of Synthetic Azo Dyes by White-Rot Fungi Grown on Diary Waste: A Step toward Sustainable and Circular Bioeconomy. Fermentation, 10(2), 80. https://doi.org/10.3390/fermentation10020080