Abstract
Cellular residues after lipid extraction are the primary waste by-products of microbial polyunsaturated fatty acid production. To mitigate the discharge of cellular residues and reduce the cost of DHA and ARA production, this study examined the utilization of enzymatically hydrolyzed Aurantiochytrium cellular residues (ACRs) and Mortierella alpina cellular residues (MCRs) as nitrogen sources. Results demonstrated that ACRs and MCRs could partially substitute yeast extract (YE) without adverse effects on DHA and ARA fermentation. Moreover, the implementation of a new fermentation medium incorporating ACRs, MCRs, and YE as mixed nitrogen sources resulted in DHA and ARA yields of 17.78 and 5.77 g/L, respectively. These values represented increases of 10.37% and 9.28% compared to traditional cultural methods, while simultaneously reducing the usage of YE by 80% and 60%, respectively. Therefore, the strategy of recycling waste cellular residues presents a novel approach for reducing the costs and environmental impact associated with microbial fermentations.
1. Introduction
Microbial oil, also referred to as single-cell oil (SCO), has emerged as a novel source of oil for various applications, such as food, medical care, and energy, complementing conventional animal and vegetable oils [1]. In recent years, there has been an increased interest in and utilization of polyunsaturated fatty acids (PUFAs) due to their high economic value and wide range of global applications [2]. PUFAs, particularly docosahexaenoic acid (DHA, C22:6, n-3) and arachidonic acid (ARA, C20:4, n-6), play significant roles in the brain and retina development of infants, as well as in improving blood lipid levels and memory in adults [3,4,5].
Aurantiochytrium sp. and Mortierella alpina are the main producers of DHA and ARA, respectively [5,6,7], and have gained commercial importance as alternatives to fish oil and animal liver as sources of these essential PUFAs. However, the industrial production of DHA and ARA through microbial fermentation still faces challenges such as low efficiency, high costs, and unstable oil quality, which result in higher prices that limit widespread adoption and application [8,9]. Among the cost factors, the fermentation medium plays a particularly significant role [10,11,12]. As a result, inexpensive raw materials have been explored as substitutes for traditional costly components like glucose and yeast extract [7,13,14,15].
It is important to note that DHA and ARA-rich oils are intracellular products obtained through cell lysis using methods such as ultrasound, enzymatic, or acid treatments. The subsequent extraction involves the use of organic solvents or centrifugation, leaving behind cellular residues as by-products. In the biofuel industry, it has been reported that approximately 65% of algal residues remain after lipid or oil extraction, posing a significant challenge [16]. For example, the production of 1 ton of DHA-rich oils from Schizochytrium sp. results in 0.75 tons of cellular residues [17], while 1 ton of ARA-rich oils from Mortierella alpina leaves 0.97 tons of cellular residues [18]. Improper disposal of these residues not only leads to environmental pollution but also represents a waste of valuable resources considering their nutrient content, including proteins, polysaccharides, and residual fatty acids. Recent studies have focused on the comprehensive utilization of this residual biomass, such as incorporating it into animal feed to provide proteins and fatty acids [6,19,20] or generating biogas through a biorefinery process [21]. Additionally, algal biomass can serve as organic fertilizer for agriculture [22].
Traditionally, the mycelial residue containing antibiotics was commonly recycled through aerobic composting, anaerobic digestion, or catalytic pyrolysis [23]. These methods have been applied to the disposal of waste microbial cellular residues. However, the DHA producer, Aurantiochytrium sp., has been reported to have a protein content ranging from 15% to 54% under different cultivation conditions [20,24,25,26,27], and it also contains carbohydrates [28]. On the other hand, the ARA producer, Mortierella alpina, has a protein content of 23.2% and is enriched with various vitamins [29]. This makes the cellular residues obtained after oil extraction a potential source of inexpensive raw materials for DHA and ARA production. Therefore, the main focus of this work was to explore the potential of utilizing cellular residues from Aurantiochytrium sp. and Mortierella alpina for the production of PUFAs. By utilizing cellular residues after oil extraction will not only reduce waste discharge and the costs associated with raw materials and waste residue treatment in DHA and ARA production, but it can also have a positive impact on environmental pollution.
In this study, the waste cellular residues of Aurantiochytrium and Mortierella, after oil extraction, were first subjected to enzymatic treatment. Subsequently, these cellular residues were reused as part of the fermentation medium for DHA and ARA production by Aurantiochytrium sp. and Mortierella alpina, respectively. These results provide a novel recycling method for cellular residues that can significantly reduce the cost of PUFA production.
2. Materials and Methods
2.1. Strains
The DHA producer strain, Aurantiochytrium sp. TZ209, used in this study was the same as in our previous research [30] and was stored in 20% (v/v) glycerol at −80 °C. The ARA producer strain, Mortierella alpina Y009, was isolated by our laboratory and is maintained at the Taizhou Key Laboratory of Biomass Functional Materials Development and Application.
2.2. Culture Conditions
2.2.1. Aurantiochytrium Culture and DHA Fermentation
The medium for the culture of Aurantiochytrium was prepared using artificial seawater and consisted of 50 g/L glucose, 10 g/L yeast extract (YE), 15 g/L monosodium glutamate, and 5 g/L corn steep liquor in artificial seawater, which contained: Na2SO4 10, MgSO4 2 (NH4)2SO4 4, KCl 0.5, K2SO4 0.65, CaCl2 0.10, KH2PO4 4, and 2 mL/L of the trace elements solution (containing MnCl2 0.5, ZnSO4 0.5, CoCl2 0.01, Na2MoO4 0.01, CuSO4 0.6, and FeSO4 2.0), and 2 mL/L of vitamins (containing 50 mg/mL thiamine, 50 mg/mL pyridoxine, and 0.25 mg/L cobalamin). The pH of the medium was adjusted to 6.5.
The Aurantiochytrium, preserved in glycerol at −80 °C, was first activated in a seed culture medium containing 20% agar for 72 h. Subsequently, a single colony was transferred into a 250 mL baffled shake-flask containing 50 mL of seed culture medium and cultured in an orbital shaker at 150 rpm and 28 °C for 24 h (one round). For the fed-batch fermentation of DHA, after three rounds of seed culture, seed broth (10%, v/v) was transferred into 500 mL baffled shake-flasks containing 100 mL of culture medium or was transferred into a 5 L bioreactor containing 3 L of culture medium for DHA fermentation. The shake flask fermentation was conducted at 170 rpm and 28 °C for 120 h, while the bioreactor fermentation was conducted at 1 vvm aeration and 28 °C for 120 h. Throughout the entire fed-batch fermentation process, the glucose concentration in the fermentation broth was maintained above 5 g/L by adding a sterilized glucose stock solution with a concentration of 800 g/L.
2.2.2. Mortierella Culture and ARA Fermentation
The seed medium for Mortierella culture consisted of 50 g/L glucose, 8 g/L YE, 2 g/L KH2PO4, 0.4 g/L MgSO4, and 4 g/L NaNO3. The Mortierella strain, preserved on potato dextrose agar (PDA) plates at 4 °C, was transferred into a 250 mL baffled shake-flask containing 50 mL of seed culture medium and incubated in an orbital shaker at 120 rpm and 25 °C for 48 h.
The fermentation medium for ARA fermentation by Mortierella was based on previous studies [31,32] with adjusted compositions. It was composed of 30 g/L glucose, 20 g/L YE, 2 g/L KH2PO4, and 0.4 g/L MgSO4. The pH of the medium was adjusted to 6.5. For the fed-batch fermentation of ARA, the method was the same as that used for DHA production, with an inoculation amount of 10% (v/v) for baffled shake-flasks and the bioreactor. Shake flask fermentation was carried out at 150 rpm and 25 °C for 168 h, while bioreactor fermentation was conducted at 2 vvm aeration and 25 °C for 168 h. Throughout the entire fed-batch fermentation process, the glucose concentration in the fermentation broth was maintained above 5 g/L by adding a sterilized glucose stock solution with a concentration of 800 g/L.
2.3. Preparation of Waste Cellular Residues
2.3.1. Aurantiochytrium Cellular Residues (ACRs)
To obtain ACRs, fed-batch cultures of Aurantiochytrium sp. were conducted in a fermenter for 120 h. After the fermentation period, the cells of Aurantiochytrium sp. were collected by centrifugation at 5000× g to separate them from the fermentation wastewater. The resulting cell pellet was then dissolved in fresh water to achieve a biomass concentration of approximately 100 g/L for subsequent cell lysis. The methodology for cell disruption was described in a previous study [10]. In summary, the temperature of the cell suspension was maintained at 50 °C, and the pH was adjusted to 10.0–11.0 using NaOH. The cell lysis was performed through hydrolysis with 0.5% (w/v) alkaline protease (Wuxi Youpuke Biotechnology Co., Ltd., Wuxi, China) for 8 h. Following the cell lysis, DHA-rich oils were obtained from the oil phase by centrifugation at 95 °C and 8000× g. The aqueous phase and slag phase were collected and stored at 4 °C as nitrogen sources for further experiments.
2.3.2. Mortierella Cellular Residues (MCRs)
After 168 h of ARA fermentation, Mortierella cells were subjected to a pasteurized step at 65 °C to inactivate lipase enzymes. Then, the cells were collected by filtration to separate them from the fermentation wastewater. The resulting mycelia were dried and disrupted using a grinder. To extract ARA-rich oils, a Soxhlet apparatus was employed, with hexane serving as the extractant. The fungal cellular residues, after oil extraction, were dried in an oven at 60 °C and subsequently crushed to a granular size of 80 mesh using a grinder. The granulated residues were resuspended in fresh water at a ratio of 1:10 (w/v), and the pH was adjusted to 5.5 for enzymatic hydrolysis. This hydrolysis step was conducted at 150 rpm and 50 °C for 6 h, utilizing 0.2% (w/v) chitinase (Jiangsu Tonglou Biotechnology Co., Ltd., Lianyungang, China) and 0.3% cellulose (Qingdao Haiweisen Biotechnology Co., Ltd., Qingdao, China) as enzymes. Following the enzymatic hydrolysis, the processed cellular residues were further treated with alkaline protease, following the same procedure as described for the Aurantiochytrium cellular residues mentioned earlier. This treatment resulted in an enzyme-treated Mortierella cellular residues (MCRs) solution, which was stored at 4 °C as a nitrogen source for subsequent experiments.
2.4. Analytical Methods
2.4.1. Total Nitrogen, Glucose, and Cell Dry Weight
The total nitrogen of waste cellular residues was determined using the Kjeldahl method. The concentration of glucose in the fermentation broth was measured using a biosensor equipped with a glucose oxidase electrode. The cell dry weight (CDW) of Aurantiochytrium sp. was measured the same as in our previous study [30]. The CDW of Mortierella was measured by filtration through filter paper to remove the fermentation broth. The mycelia were then washed three times with distilled water and dried at 60 °C until a constant weight was achieved.
2.4.2. Total Lipids, DHA, and ARA Content
In our previous study [33], we described the extraction of total lipids (TL) from DHA fermentation by Aurantiochytrium sp. Now, for the extraction of TL from ARA fermentation by Mortierella, the dry biomass was initially ground into a fine powder (60 mesh), and lipids from 2 g of the powder were extracted using a Soxhlet apparatus. The extraction process involved using 100 mL of hexane as the extractant and maintaining the extraction temperature at 75 °C for 8 h. The DHA and ARA content were determined using a GC-2030 system (Shimadzu, Kyoto, Japan) following the fatty acid methylation and analysis methods described in our previous study [30].
3. Results and Discussion
3.1. Nutrient Composition of Cellular Residues from PUFA Fermentation
In order to evaluate the potential utilization of ACRs and MCRs as partial substitutes for YE in the fermentation medium for DHA and ARA production, we conducted an analysis of the nutrient composition of these cellular residues, as presented in Table 1.

Table 1.
Nutrient components of Aurantiochytrium cellular residues and Mortierella cellular residues.
The dry biomass of Aurantiochytrium comprises both lipid and non-lipid components [33]. During the lipid extraction process, the non-lipid fraction of biomass was subjected to enzymolysis using alkaline protease to break down complex proteins into simpler peptides or amino acids. From Table 1, it can be observed that ACRs have a total nitrogen (TN) content of 6.37 g/L and a free ammonia (FA) content of 2.44 g/L. These values are equivalent to 54.26 and 48.61 g/L of YE (Angle LM808, Yichang, China), which has the TN and FA values of 11.74% and 5.02%, respectively.
Mucoromycota fungi, such as Mortierella, have the natural ability to produce chitosan, which typically constitutes 1% to 10% of the cell dry weight [34]. Notably, chitin/chitosan makes up more than 40% of the cell wall of Mortierella alpina [35]. This characteristic led to the initial treatment of MCRs with chitinase following the lipid extraction process. In the past, Mortierella cellular residues were typically regarded as waste and disposed of or sent to landfills, resulting in the squandering of resources and environmental pollution. However, recent studies have focused on utilizing these cellular residues as fertilizers or feed materials [36] due to their protein content and remaining lipid components. After undergoing further enzymatic treatment with alkaline protease, the TN and FN values of the MCRs were found to be 10.43 g/L and 3.50 g/L, respectively.
In addition, both ACRs and MCRs were found to contain significant amounts of phosphorus (P), potassium (K), and other minerals that are essential inorganic salts required in the fermentation medium, as shown in Table 1. These chemical analysis results suggest that the ACRs and MCRs, obtained after undergoing multiple enzyme treatments, could potentially be used as alternative nitrogen sources to partially replace YE in fermentation media for the production of PUFAs. To assess the feasibility of incorporating these novel nitrogen sources into the fermentation process, different proportions of YE were substituted with the treated cellular residues. The resulting media were then tested in laboratory-scale shake flask fermentations of Aurantiochytrium and Mortierella.
3.2. Aurantiochytrium Cellular Residues as a Nitrogen Source for DHA and ARA Production
It is well known that YE is commonly used as a nitrogen source in DHA and ARA production by Aurantiochytrium sp. and Mortierella alpina [37,38]. To investigate the potential use of ACRs as an alternative nitrogen source to replace YE, the effects of different ratios of YE replacement with ACRs on DHA and ARA fermentation were examined. Importantly, the initial total nitrogen content of each fermentation medium was kept at the same value based on a TN value of 10 g/L of YE.
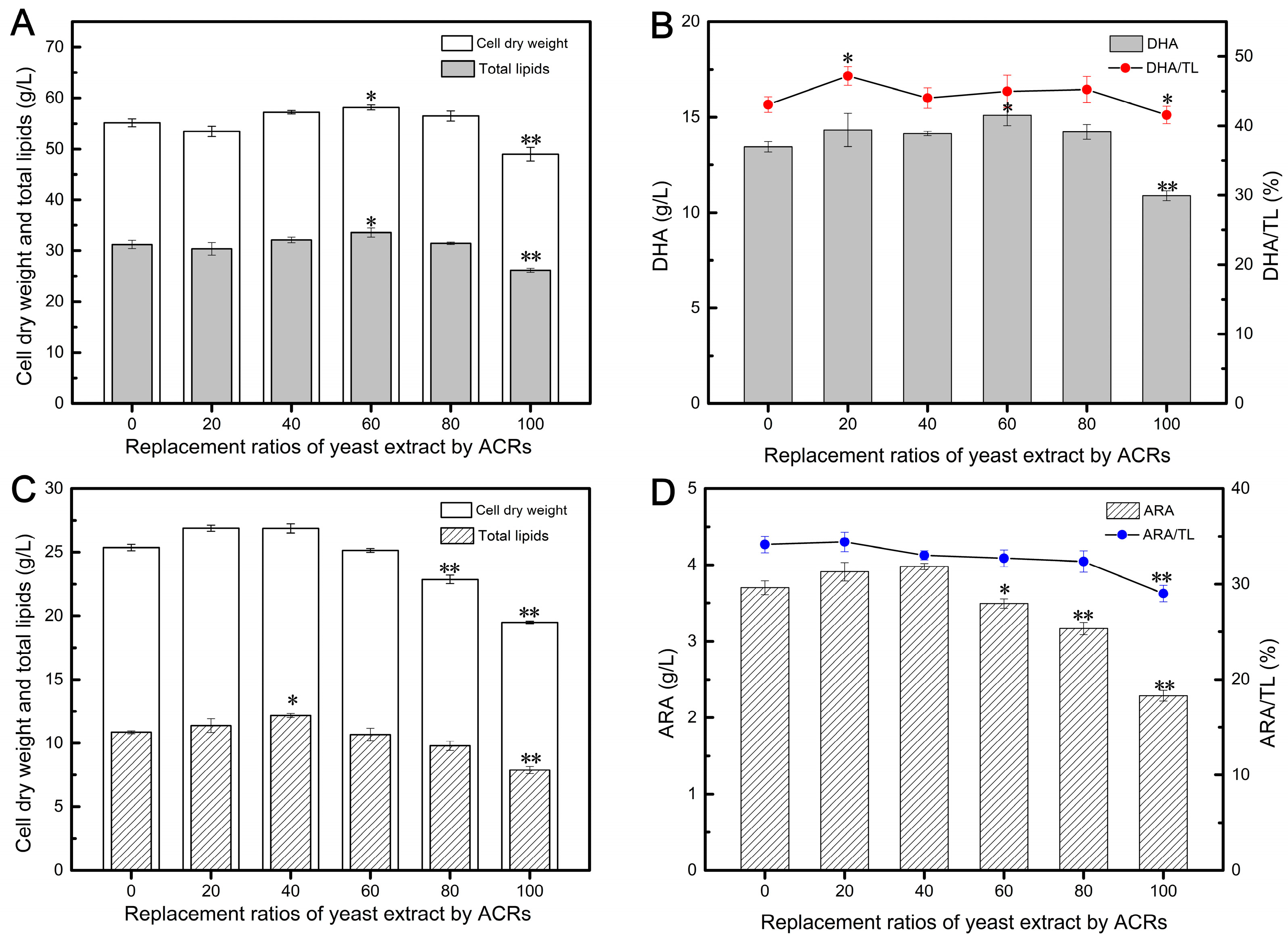
Figure 1A illustrates the varying effects of different ACR addition ratios on the biomass of Aurantiochytrium. Interestingly, the application of ACRs did not significantly impact the final CDW when the replacement ratios were in the range of 20–80%. These ratios yielded similar results to the control group, suggesting that enzymatically-treated ACRs could be partially recycled as a substitute for YE in Aurantiochytrium cultivation. Notably, replacing 60% of YE with ACRs resulted in the highest CDW of 58.16 g/L, surpassing the value obtained with 100% YE. This finding suggests that ACRs may contain easily accessible nitrogen sources derived from the enzymatic hydrolysis of proteins, which could be more conducive to promoting cell growth. However, complete replacement of YE with ACRs (100% replacement) resulted in a significant decrease in biomass, reaching only 43.58 g/L. This decrease may be attributed to the presence of self-generated fermentation inhibitors and the absence of essential cell growth factors commonly found in YE [10].
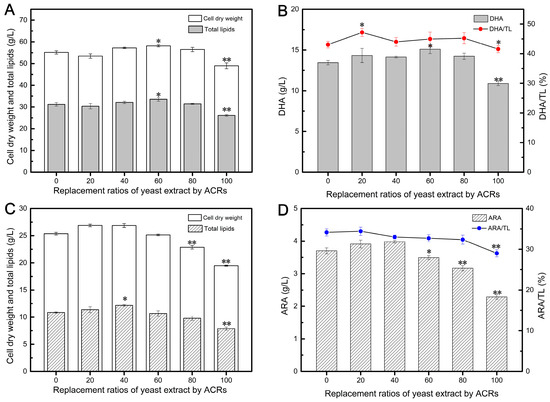
Figure 1.
Effects of different replacement ratios of yeast extract by ACRs on DHA (A,B) and ARA (C,D) fermentation parameters. Values are represented as the mean ± standard deviations of three independent experiments. The statistical significance between the experimental group and the control group was presented by a t-test. * p < 0.05, ** p < 0.01.
Previous studies have demonstrated that cell growth and lipid accumulation in Aurantiochytrium are not synchronized. The synthesis of DHA-rich oils often begins in response to external stress conditions, such as the depletion of nitrogen sources [30,39]. Therefore, promoting the growth of highly vital cells in the early stage is beneficial for adapting to adverse environments and facilitating lipid accumulation in the later stage [40]. As depicted in Figure 1A, the TL content followed a similar trend to biomass accumulation, and replacing 20–80% of YE with ACRs did not have an impact on lipid synthesis during the oil accumulation phase. Interestingly, the TL content gradually increased with the proportion of ACRs, from 20% to 60%, suggesting that some nitrogen sources derived from enzymatically treated cellular residues contributed to lipid accumulation. Similarly, during the phase of oil accumulation, replacing 20–80% of YE with ACRs did not impact DHA synthesis (Figure 1B). In fact, the DHA content after 120 h of fermentation increased from 13.45 to 15.10 g/L at ACR replacement ratios of 0% to 60%. However, completely replacing YE with ACRs resulted in a substantial decrease in DHA production (Figure 1B), indicating that a certain amount of YE was essential for DHA synthesis. The highest DHA yield was achieved when replacing 60% of YE with ACRs, although an Although the 80% replacement group resulted in a DHA yield of 14.23 g/L, lower than the group replacing 60% of YE, it was still 5.80% higher than the control group with 100% YE, demonstrating the suitability of using enzymatically treated cellular residues of Aurantiochytrium as nitrogen sources for DHA production.
Additionally, the applicability of ACRs as a nitrogen source for ARA production by Mortierella was also tested (Figure 1C,D). When YE was the sole nitrogen source, the CDW reached 25.36 g/L. Low substitution ratios (20–40%) of ACRs did not impact normal cell growth of Mortierella, and the CDW increased from 25.33 to 26.88 g/L with ACR replacement ratios ranging from 0% to 40% (Figure 1C). However, higher substitution ratios resulted in a significant decrease in biomass accumulation, which could be attributed to the specific amino acids found in each nitrogen source [41]. As YE usage decreased, the fermentation broth became less conducive to cell growth. It is worth noting that YE contains molecules such as guanine, guanosine-5′-triphosphate, vitamins, and growth factors, which play crucial roles in the growth and metabolism of microbial cells [42].
The accumulation of ARA-rich oils is also dependent on nitrogen consumption in the culture system, which influences the distribution of carbon flux towards lipid synthesis [43]. In this study, the culture supplemented with 100% YE produced 10.85 g/L of TL, and the addition of a small amount of ACRs did not lead to a reduction in lipid accumulation. On the contrary, the culture with 40% of YE replaced by ACRs achieved a higher TL content, reaching 12.06 g/L, possibly as a result of enhanced mycelium growth (Figure 1C). Previous studies have analyzed the impact of different nitrogen sources on ARA synthesis [44,45,46]. Typically, YE is regarded as the ideal organic nitrogen source for ARA production by Mortierella and has been demonstrated to be the most effective nitrogen source for ARA yield [9,47]. However, in this study, we found that a mixture of YE and ACRs was more conducive to ARA synthesis, resulting in an ARA yield of 3.98 g/L when 40% of YE was replaced by ACRs, representing a 7.576% improvement compared to 100% YE (Figure 1D), with an ARA/TL ratio of 33.00%. This conclusion aligns with a previous report that used corn steep liquor and YE as a combined nitrogen source to improve ARA production [44].
These results demonstrate that ACRs, obtained after enzymatic hydrolysis, can be reused as a nitrogen source to partially replace YE in DHA and ARA production. In fact, inexpensive ACRs were able to replace 80% and 40% of YE in the fermentation of DHA and ARA, respectively, highlighting their potential to be utilized as a new raw material.
3.3. Mortierella Cellular Residues as a Nitrogen Source for DHA and ARA Production
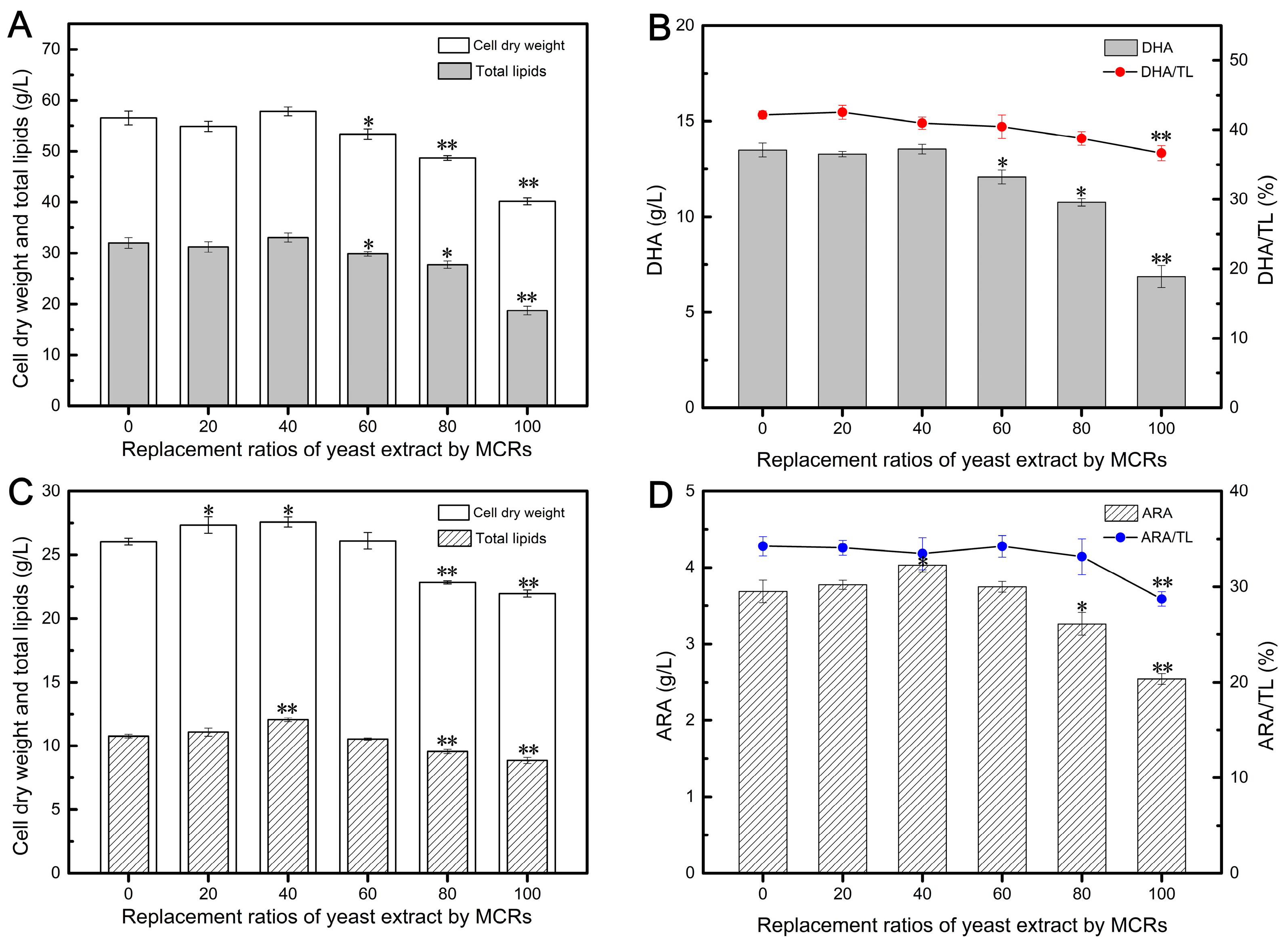
After 120 h of fermentation with Aurantiochytrium, it was evident that replacing 20–60% of YE with MCRs resulted in comparable biomass levels to the control group (Figure 2A), indicating that low replacement ratios of YE with MCRs did not affect the cell growth of Aurantiochytrium. Interestingly, the fermentation medium with a 40% replacement of MCRs achieved the highest CDW of 57.81 g/L. This can be attributed to the presence of proteins, carbohydrates, and vitamins in the enzymatically treated cellular residues, which may promote cell growth [29,35]. However, further increasing the replacement ratio had a negative impact on cell growth. This could be due to the challenge of hydrolyzing the proteins present in Mortierella cellular residues, as they are mainly distributed in mycelial biomass and have a complex structure [48], leading to incomplete protein hydrolysis and the presence of insoluble residues in MCRs.
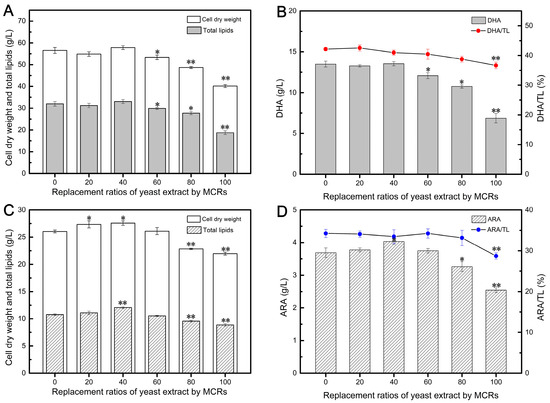
Figure 2.
Effects of different replacement ratios of yeast extract by MCRs on DHA (A,B) and ARA (C,D) fermentation parameters. Values are represented as the mean ± standard deviations of three independent experiments. The statistical significance between the experimental group and the control group was presented by a t-test. * p < 0.05, ** p < 0.01.
Similarly, replacing 20–40% of YE with MCRs yielded TL yields of 32.07 and 33.05 g/L, respectively, which were comparable to the control group using 100% YE (Figure 2A). Previous studies have demonstrated that the composition of amino acids in nitrogen sources can influence lipid accumulation, with low glutamate (Glu) content and high arginine (Arg) and proline (Pro) concentrations promoting lipid accumulation [10]. However, this effect depends on the suitability of nitrogen sources. As Figure 2A depicts, when the replacement exceeded 40%, the cells were unable to effectively utilize the nitrogen source components provided by MCRs, resulting in a decrease in TL yield. It is worth noting that Aurantiochytrium synthesizes DHA through Polyketide Synthase (PKS)-like machinery, which is an anaerobic pathway [49]. Therefore, conditions with low dissolved oxygen levels promote DHA accumulation. In this study, cultures with a 20–40% replacement of MCRs achieved a final DHA content of approximately 13.5 g/L (Figure 2B), similar to the control group, indicating that the PKS pathway functioned normally. However, higher MCR replacement ratios significantly inhibited DHA synthesis, and complete substitution of YE with MCRs resulted in only 6.87 g/L of DHA, which was 49.07% lower than the control group. This may be attributed to the inhibition of the PKS pathway due to insufficient or unsuitable nitrogen sources.
The impact of substituting YE with MCRs on ARA fermentation is shown in Figure 2C,D. Similar to its application in DHA fermentation, enzymatically-treated MCRs could replace up to 60% of YE without affecting cell growth. Furthermore, the highest CDW of 27.57 g/L was achieved in the 40% replacement group, representing a 5.88% increase compared to the control group, thus indicating the feasibility of utilizing MCRs for Mortierella cell growth after enzymatic treatment (Figure 2C). However, replacing over 80% of YE with MCRs resulted in a gradual decrease in CDW, possibly due to a lack of essential growth factors such as amino acids, vitamins, and nucleotides provided by YE [50]. A previous study demonstrated the successful replacement of 100% of YE and mineral salts in the culture medium using the fungal extract of Mucor indicus [51]. However, this difference may be attributed to specific processing methods employed for MCRs. In this study, even after undergoing multi-step enzymatic treatment, the MCRs still retained some insoluble residues.
The final TL yield followed a similar trend as cell biomass since high cell density is a prerequisite for effective lipid accumulation. Low replacement of YE with MCRs did not have an impact on lipid accumulation in Mortierella (Figure 2C). As expected, higher fungal biomass concentrations resulted in higher lipid yields at the end of fermentation, with the highest lipid content of 12.05 g/L achieved with a 40% replacement of MCRs. The production of high-quality, ARA-rich oil not only requires a high lipid yield but also necessitates a high ARA content in total fatty acids. Studies have reported that the ARA synthesis pathway in Mortierella involves desaturation and elongation steps [52]. Figure 2D shows that the highest ARA yield of 4.03 g/L was obtained in the 40% ACR replacement group. Interestingly, further addition of MCRs (60%) resulted in an ARA/TL ratio of 34.23%, similar to that of the control group, resulting in a final ARA content of 3.75 g/L, which was similar to the value of the control group. This indicates that 60% replacement of MCRs did not have an inhibitory effect on ARA accumulation. However, the ARA/TL ratios obtained with 80% and 100% replacement of YE with ACRs were 33.14% and 28.71%, respectively, suggesting that even a small amount of YE is necessary for efficient ARA biosynthesis.
These results demonstrate that MCRs obtained after multi-step enzymatic hydrolysis can be utilized as a novel nitrogen source for DHA and ARA fermentation. With this approach, 40% and 60% of YE can be replaced by MCRs in DHA and ARA fermentations, respectively, without significant differences compared to traditional fermentations. Furthermore, MCRs derived from Mortierella have the advantage of being free from antibiotic residues, unlike residues from antibiotic-producing fungi, which can be challenging [23]. Therefore, MCRs have the potential to serve as a novel nitrogen source, similar to ACRs.
3.4. Fermentation of PUFAs Using Waste Cellular Residues
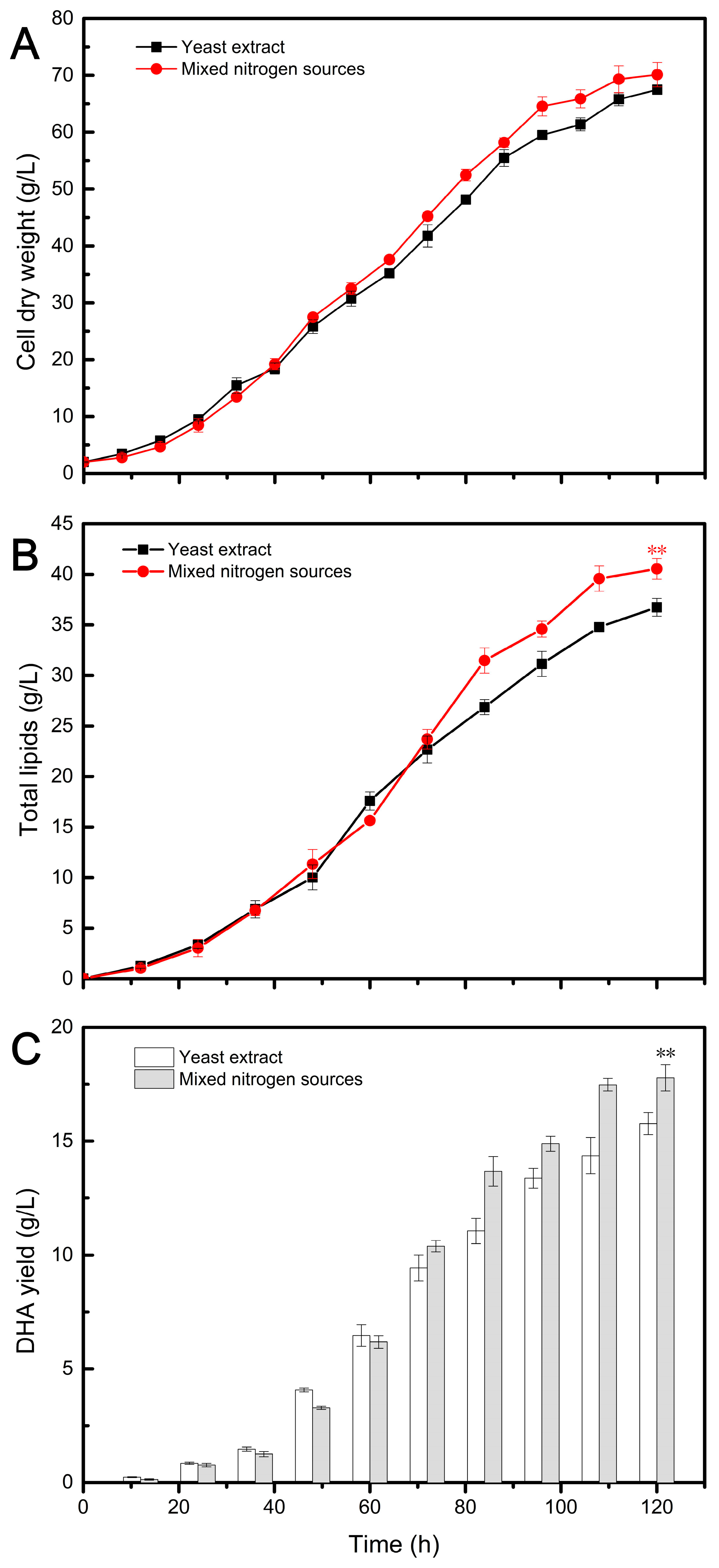
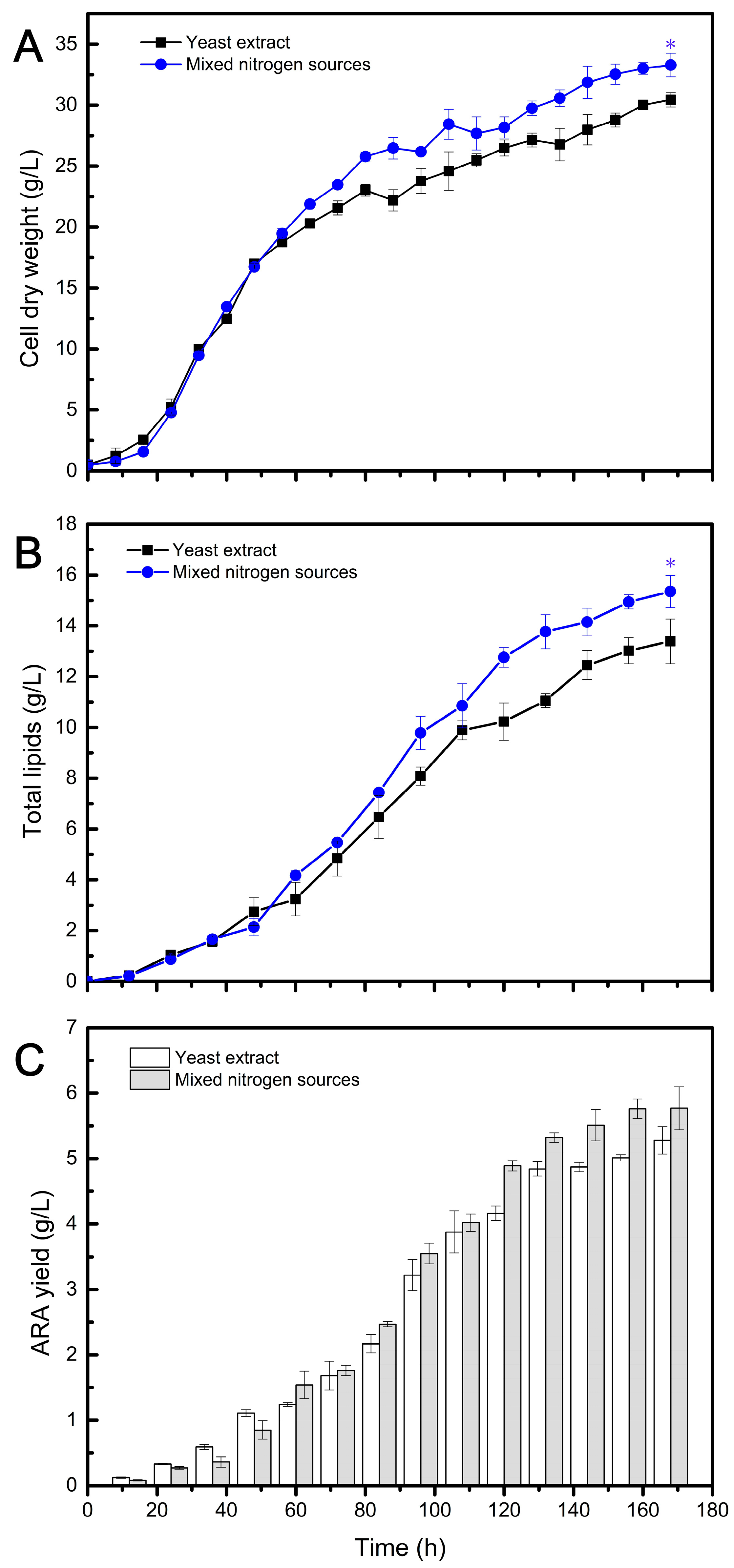
Having established that recycling waste cellular residues as a partial replacement for YE did not impact DHA and ARA fermentation, the study applied a mixed nitrogen source (MNS) composed of ACRs, MCRs, and YE in DHA and ARA fermentation. Based on previous results, where 80% ACRs and 40% MCRs had no effect on DHA yield, the ratio of ACRs and MCRs was set at 80:40. Considering that at least 20% of YE was required for DHA fermentation, the final usage ratio of the three nitrogen sources (ACRs, MCRs, and YE) for DHA fermentation was set at 53.33:26.67:20. Similarly, 40% ACRs and 60% MCRs had no effect on ARA yield, so the ratio of ACRs and MCRs was set at 40:60. Taking into account that at least 40% of YE was necessary for ARA fermentation, the final usage ratio of the three nitrogen sources for ARA fermentation was set at 24:36:40. The experiments were conducted in a 5 L bioreactor, and the results are shown in Figure 3 and Figure 4 and summarized in Table 2 and Table 3.
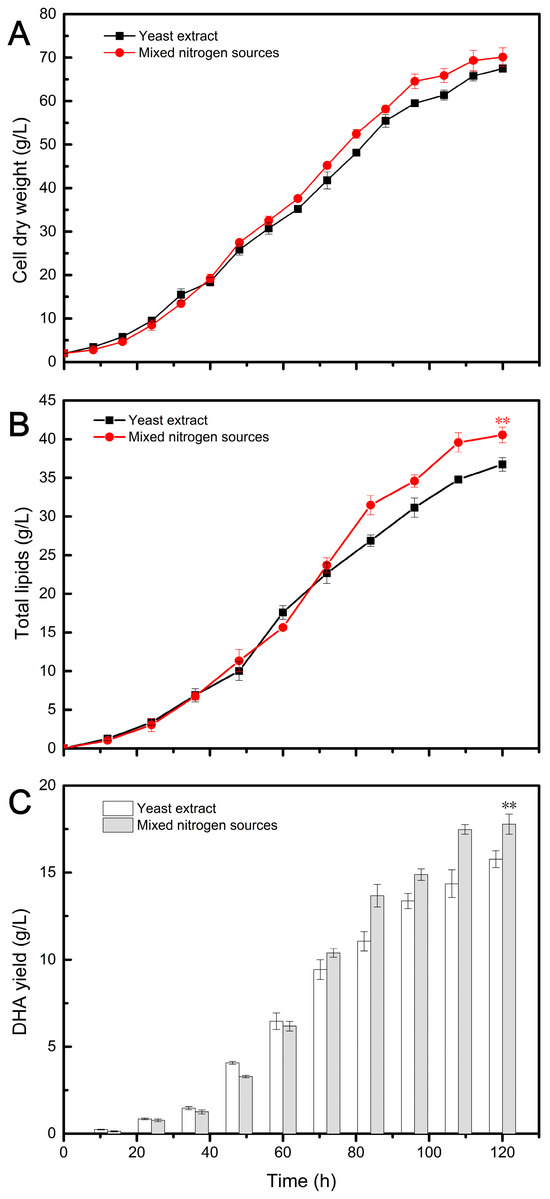
Figure 3.
Fermentation profiles of DHA production by Aurantiochytrium sp. TZ209 using different nitrogen sources. The statistical significance between the final results was presented by a t-test. ** p < 0.01.
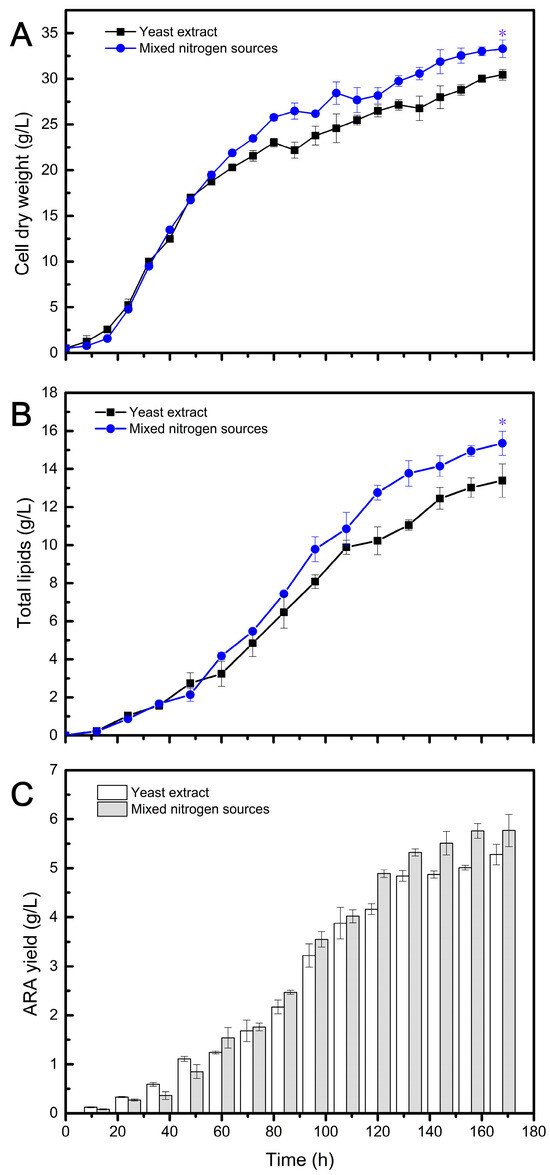
Figure 4.
Fermentation profiles of ARA production by Mortierella alpina (Y009) using different nitrogen sources. The statistical significance between the final results was presented by a t-test. * p < 0.05.
Table 2.
Comparison of DHA fermentation performances using mixed nitrogen sources medium and yeast extract source medium.
Table 3.
Comparison of ARA fermentation performances using mixed nitrogen source medium and yeast extract source medium.
In Figure 3A, the cell growth using mixed nitrogen sources (ACRs, MCRs, and YE) exhibited a similar pattern to that observed with YE alone in the initial 48 h. Previous studies have reported a lack of synchronization between cell growth and lipid synthesis in Aurantiochytrium [30], with the accumulation of biomass in the early stage primarily consisting of non-lipid biomass [33]. Our results indicate that the utilization of MNS did not affect the normal cell growth of Aurantiochytrium compared to YE. However, subsequent differences observed may be related to TL accumulation (Figure 3B). The use of MNS appears to be more conducive to lipid synthesis, as evidenced by higher TL levels observed in the MNS group compared to the YE group during the oil accumulation stage. While nitrogen limitation promotes lipid accumulation, excessive nitrogen limitation actually reduces lipid productivity due to decreased biomass. Therefore, a nitrogen-feeding strategy was employed to alleviate the inhibitory effects of high nitrogen limits [53]. In this study, it is possible that some slow-release nitrogen sources present in the mixed nitrogen had a similar effect as the nitrogen feeding strategy. The DHA yield followed a similar trend to TL, as indicated in Figure 3C. The DHA content obtained from the MNS group was higher than that from the control group from 72 h until the end of fermentation. This may be attributed to the accumulation of TL in cells, as the DHA/TL ratio was similar in both groups (Table 2).
Figure 4 presents the data for CDW, lipid content, and ARA production between the MNS and YE conditions. During the first 48 h of fermentation, the CDW obtained from both nitrogen sources did not exhibit significant differences, indicating a period of rapid cell growth. However, differences between the two conditions started to emerge after 48 h and continued until the end of fermentation. After 168 h, the biomass obtained from the MNS reached 33.29 g/L, whereas the control group exhibited slower growth with a final CDW of 30.44 g/L (Figure 4A). Generally, in the early stage of fermentation, there is primarily rapid growth of fungal cells without significant lipid accumulation. The TL content did not change significantly in the first 48 h under both fermentation conditions, and the lipids synthesized at this stage mainly consisted of functional membrane lipids. However, after this period, oils started to accumulate within the cells. As depicted in Figure 4B, the trend of TL content under both conditions closely resembled that of CDW. This is because ARA-rich oils are intracellular products that are coupled with biomass. The variation in ARA yield is shown in Figure 4C, and it is evident that the MNS group achieved a higher ARA yield during the oil accumulation stage, despite having a lower ARA/TL ratio compared to the YE group (Table 3). This may be due to the presence of insoluble substances that hinder the transfer of oxygen and nutrients. Nevertheless, the final ARA yield was higher than that obtained from YE.
The results of the novel fermentation using MNS and the traditional fermentation using YE are summarized in Table 2 and Table 3. As shown in Table 2, all fermentation indices obtained from the novel fermentation were higher compared to those in the traditional fermentation. Particularly noteworthy is the DHA yield of 17.78 g/L obtained from the novel recycled medium, which was 12.75% higher than that obtained with the traditional medium. This demonstrates that the MNS composed of ACRs (63%), MCRs (26%), and YE (20%) had a positive effect on the accumulation of DHA oil in Aurantiochytrium. Additionally, this MNS formulation could save 80% of the expensive YE used in traditional fermentation. Similarly, the application of combined nitrogen sources promoted ARA production in Mortierella, resulting in a 9.36% increase in CDW, a 14.64% increase in TL, and a 9.8% increase in ARA yield compared to using YE alone as the nitrogen source (Table 3). Furthermore, this strategy reduced the use of expensive YE by 60%. These findings highlight the benefits of using MNS and mixed nitrogen sources for improving the production of DHA and ARA while also reducing the reliance on expensive YE.
3.5. Nitrogen Source Cost Analysis
The calculation for using MNS and YE as nitrogen sources in the production of DHA and ARA can be observed in Table 4 and Table 5. The enzymes employed in this study, including alkaline protease, chitinase, and cellulase, are commercially accessible enzymatic products. The price of alkaline protease is 6.26 USD per kilogram; chitinase costs 82.04 USD per kilogram; and cellulase is priced at 6.26 USD per kilogram. Additionally, the YE utilized in this study is also a commercially available product and is priced at 5.70 USD per kilogram. These prices serve as an estimation of the expenses associated with employing these enzymatic products and YE during the fermentation process, based on an exchange rate of 1 USD = 7.19 RMB as of January 2024.
Table 4.
Cost calculation of conventional fermentation using yeast extract and the novel fermentation using mixed nitrogen sources in DHA fermentation.
Table 5.
Cost calculation of conventional fermentation using yeast extract and the novel fermentation using mixed nitrogen sources in ARA fermentation.
For DHA fermentation, in the case of 1 ton of culture medium, the conventional fermentation method consumes 10 kg of YE with a nitrogen content of 11.74%. This means that YE provides a total of 1174 g of nitrogen. In the new fermentation method, while maintaining the same total nitrogen conditions, the nitrogen sources consist of ACRs, MCRs, and YE in a ratio of 53.33:26.67:20. Consequently, the required amount of ACRs in the new culture medium is 98.29 L, MCRs are 30.02 L, and YE is 2 kg. These values are calculated based on the TN content of 6.37 g/L, 10.43 g/L, and 11.74%, respectively. The necessary enzyme amounts for ACRs are 0.49145 kg of alkaline protease. For MCRs, the required enzyme amounts are as follows: 0.1501 kg of alkaline protease, 0.09006 kg of cellulase, and 0.0600 kg of chitinase. These values are calculated based on usage percentages of 0.5%, 0.3%, and 0.2% of 30.02 L MCRs, respectively. It should be noted that, in practical application, it is necessary to concentrate solutions of ACRs and MCRs according to the dosage. Consequently, the total amounts of enzymes used are 0.6415 kg of alkaline protease, 0.0901 kg of cellulase, and 0.0600 kg of chitinase. The total cost of enzymes is calculated by multiplying their unit prices by the quantities required: 0.6415 × 6.26 + 0.0901 × 6.26 + 0.0600 × 82.04 = 9.50 USD. The total cost of YE is calculated as 2 × 5.70 = 11.40 USD. Therefore, the total cost of enzymes and YE for the new fermentation method is 9.50 + 11.40 = 20.90 USD. In contrast, under traditional cultural conditions, the total cost amounts to 57 USD (10 × 5.70 USD for YE).
Therefore, the cost of nitrogen sources for producing 1 kg of DHA is calculated to be 3.61 USD for traditional fermentation and 1.18 USD for novel fermentation. This represents a reduction of 67.31% in the cost of DHA production using this recycling fermentation strategy (Table 4).
Similarly, for ARA fermentation, the total cost of the nitrogen source is calculated to be 65.73 USD under the new culture conditions, compared to 114 USD under the traditional culture conditions. This recycling fermentation strategy results in a 47.24% reduction in the cost of ARA production (Table 5).
The economic viability of PUFA production by Aurantiochytrium sp. and Mortierella alpina is greatly influenced by the costs associated with the fermentation of raw materials. To address this issue, previous studies have explored the use of low-cost resources and corresponding fermentation strategies, such as soybean meal [54], corn steep liquor [44], sugarcane bagasse [55], corn, rice, and potatoes [56], cane molasses [40], tofu whey wastewater [12], and alanine mother liquor [57]. According to reports, utilizing food industry by-products and waste streams as fermentation resources can result in a significant reduction of 38% in the unit production cost of DHA [58]. In this work, ACRs and MCRs after enzymic treatment were used as nitrogen sources, partially replacing YE. The combination of these mixed nitrogen sources with ACRs, MCRs, and YE provided favorable conditions for cell growth, lipid synthesis, and PUFA production, resulting in a significant decrease in production costs. In conclusion, considering the importance of substrate cost for the economical production of PUFAs and the environmental pollution caused by waste cellular residues from microbial fermentation, this study successfully implemented a recycling strategy for waste cellular residues in single-cell oil production. This not only lowers costs but also reduces environmental pollution. Furthermore, this strategy has the potential to be applied to the fermentation of other biological products.
4. Conclusions
In this study, cellular residues from Aurantiochytrium sp. and Mortierella alpina were collected after lipid extraction and underwent enzymatic hydrolysis. The resulting ACRs and MCRs were then recycled as a novel nitrogen source to partially replace yeast extract in the DHA and ARA fermentation mediums. By using ACRs and MCRs in combination with a small amount of yeast extract as mixed nitrogen sources, it was observed that PUFA production increased compared to using 100% yeast extract. Furthermore, this approach led to substantial cost savings, with the nitrogen source costs for DHA and ARA fermentation being reduced by 67.31% and 47.24%, respectively. Notably, this waste recycling method not only contributes to cost savings but also presents a new approach to mitigate environmental pollution in PUFA production. By repurposing and reusing cellular residues, this strategy helps reduce waste and promote sustainability in the production process.
Author Contributions
Conceptualization, F.-W.Y.; methodology, F.-W.Y., J.H., C.-T.Z. and W.-L.Z.; validation, L.-F.Y. and Y.-Y.Z.; formal analysis, F.-W.Y. and X.L.; investigation, J.H. and C.-T.Z.; resources, F.-W.Y.; data curation, X.-L.S.; writing—original draft preparation, F.-W.Y.; writing—review and editing, F.-W.Y. and Y.-Q.F.; visualization, F.-W.Y.; supervision, Y.-Q.F.; project administration, Y.-Q.F.; funding acquisition, F.-W.Y. and Y.-Q.F. All authors have read and agreed to the published version of the manuscript.
Funding
This work was funded by the Taizhou Scientific and Technological Project (numbers 22gyb06 and 22gya05) and the National College Students Innovation and Entrepreneurship Training Program (number 202110350022).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data are contained within the article.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Ratledge, C. Fatty acid biosynthesis in microorganisms being used for Single Cell Oil production. Biochimie 2004, 86, 807–815. [Google Scholar] [CrossRef]
- Ji, X.-J.; Ledesma-Amaro, R. Microbial Lipid Biotechnology to Produce Polyunsaturated Fatty Acids. Trends Biotechnol. 2020, 38, 832–834. [Google Scholar] [CrossRef]
- Campoy, C.; Chisaguano Tonato, A.M.; de la Garza Puente, A.; Seenz de Pipaon, M.; Verduci, E.; Koletzko, B.; Gonzalez Casanova, I.; Larque, E.; Valenzuela, R.; Moreno Villares, J.M.; et al. Controversy about the critical role of long-chain pol-yunsaturated fatty acids, arachidonic acid (ARA) and docosahexaenoic acid (DHA), during infancy. Nutr. Hosp. 2021, 38, 1101–1112. [Google Scholar]
- Von Schacky, C. Importance of EPA and DHA Blood Levels in Brain Structure and Function. Nutrients 2021, 13, 1074. [Google Scholar] [CrossRef] [PubMed]
- Mironov, A.A.; Nemashkalov, V.A.; Stepanova, N.N.; Kamzolova, S.V.; Rymowicz, W.; Morgunov, I.G. The Effect of pH and Temperature on Arachidonic Acid Production by Glycerol-Grown Mortierella alpina NRRL-A-10995. Fermentation 2018, 4, 17. [Google Scholar] [CrossRef]
- Chang, L.; Chen, H.; Tang, X.; Zhao, J.; Zhang, H.; Chen, Y.Q.; Chen, W. Advances in improving the biotechnological application of oleaginous fungus Mortierella alpina. Appl. Microbiol. Biotechnol. 2021, 105, 6275–6289. [Google Scholar] [CrossRef]
- Xu, X.; Huang, C.; Xu, Z.; Xu, H.; Wang, Z.; Yu, X. The strategies to reduce cost and improve productivity in DHA production by Aurantiochytrium sp.: From biochemical to genetic respects. Appl. Microbiol. Biotechnol. 2020, 104, 9433–9447. [Google Scholar] [CrossRef]
- Kothri, M.; Mavrommati, M.; Elazzazy, A.M.; Baeshen, M.N.; Moussa, T.A.A.; Aggelis, G. Microbial sources of polyun-saturated fatty acids (PUFAs) and the prospect of organic residues and wastes as growth media for PUFA-producing microorganisms. FEMS Microbiol. Lett. 2020, 367, fnaa028. [Google Scholar] [CrossRef]
- Chen, Y.-H.; Ong, C.-C.; Lin, T.-Y. Effect of Sea Salt and Taro Waste on Fungal Mortierella alpina Cultivation for Arachidonic Acid-Rich Lipid Production. Fermentation 2022, 8, 81. [Google Scholar] [CrossRef]
- Yin, F.-W.; Guo, D.-S.; Ren, L.-J.; Ji, X.-J.; Huang, H. Development of a method for the valorization of fermentation wastewater and algal-residue extract in docosahexaenoic acid production by Schizochytrium sp. Bioresour. Technol. 2018, 266, 482–487. [Google Scholar] [CrossRef]
- Park, W.-K.; Moon, M.; Shin, S.-E.; Cho, J.M.; Suh, W.I.; Chang, Y.K.; Lee, B. Economical DHA (Docosahexaenoic acid) production from Aurantiochytrium sp. KRS101 using orange peel extract and low cost nitrogen sources. Algal Res. Biomass Biofuels Bioprod. 2018, 29, 71–79. [Google Scholar] [CrossRef]
- Wang, S.-K.; Wang, X.; Tian, Y.-T.; Cui, Y.-H. Nutrient recovery from tofu whey wastewater for the economical production of docosahexaenoic acid by Schizochytrium sp. S31. Sci. Total. Environ. 2020, 710, 136448. [Google Scholar] [CrossRef]
- Goyzueta-Mamani, L.D.; de Carvalho, J.C.; Magalhaes, A.I., Jr.; Soccol, C.R. Production of arachidonic acid by Mortierella alpine using wastes from potato chips industry. J. Appl. Microbiol. 2021, 130, 1592–1601. [Google Scholar] [CrossRef]
- Asimakopoulou, G.; Karnaouri, A.; Staikos, S.; Stefanidis, S.D.; Kalogiannis, K.G.; Lappas, A.A.; Topakas, E. Production of Omega-3 Fatty Acids from the Microalga Crypthecodinium cohnii by Utilizing Both Pentose and Hexose Sugars from Agricultural Residues. Fermentation 2021, 7, 219. [Google Scholar] [CrossRef]
- Yin, F.-W.; Sun, X.-L.; Zheng, W.-L.; Yin, L.-F.; Luo, X.; Zhang, Y.-Y.; Wang, Y.-F.; Fu, Y.-Q. Development of a Strategy for L-Lactic Acid Production by Rhizopus oryzae Using Zizania latifolia Waste and Cane Molasses as Carbon Sources. Molecules 2023, 28, 6234. [Google Scholar] [CrossRef]
- Kumar, M.; Sun, Y.; Rathour, R.; Pandey, A.; Thakur, I.S.; Tsang, D.C. Algae as potential feedstock for the production of biofuels and value-added products: Opportunities and challenges. Sci. Total. Environ. 2020, 716, 137116. [Google Scholar] [CrossRef] [PubMed]
- Guo, D.-S.; Tong, L.-L.; Ji, X.-J.; Ren, L.-J.; Ding, Q.-Q. Development of a Strategy to Improve the Stability of Culture Envi-ronment for Docosahexaenoic Acid Fermentation by Schizochytrium sp. Appl. Biochem. Biotechnol. 2020, 192, 881–894. [Google Scholar] [CrossRef]
- Shi, K.; Gao, Z.; Lin, L.; Wang, W.-J.; Shi, X.-Q.; Yu, X.; Song, P.; Ren, L.-J.; Huang, H.; Ji, X.-J. Manipulating the generation of reactive oxygen species through intermittent hypoxic stress for enhanced accumulation of arachidonic acid-rich lipids. Chem. Eng. Sci. 2018, 186, 36–43. [Google Scholar] [CrossRef]
- Ammar, E.M.; Arora, N.; Philippidis, G.P. The Prospects of Agricultural and Food Residue Hydrolysates for Sustainable Production of Algal Products. Energies 2020, 13, 6427. [Google Scholar] [CrossRef]
- Jaseera, K.V.; Ebeneezar, S.; Sayooj, P.; Nair, A.V.; Kaladharan, P. Dietary supplementation of microalgae, Aurantiochytrium sp. and co-feeding with Artemia enhances the growth, stress tolerance and survival in Penaeus monodon (Fabricius, 1798) post larvae. Aquaculture 2021, 533, 736176. [Google Scholar]
- Uggetti, E.; Sialve, B.; Trably, E.; Steyer, J.-P. Integrating microalgae production with anaerobic digestion: A biorefinery approach. Biofuels Bioprod. Biorefining-Biofpr 2014, 8, 516–529. [Google Scholar] [CrossRef]
- Karthik, V.; Kumar, P.S.; Vo, D.-V.N.; Sindhu, J.; Sneka, D.; Subhashini, B.; Saravanan, K.; Jeyanthi, J. Hydrothermal pro-duction of algal biochar for environmental and fertilizer applications: A review. Environ. Chem. Lett. 2021, 19, 1025–1042. [Google Scholar] [CrossRef]
- Hui, X.; Fang, W.; Wang, G.; Liu, H.; Dai, X. Waste recycling of antibiotic mycelial residue: The feasible harmless treatment and source control of antibiotic resistance. J. Clean. Prod. 2023, 401, 136786. [Google Scholar] [CrossRef]
- Nobrega, R.O.; Batista, R.O.; Correa, C.F.; Mattioni, B.; Filer, K.; Pettigrew, J.E.; Fracalossi, D.M. Dietary supplementation of Aurantiochytrium sp. meal, a docosahexaenoic-acid source, promotes growth of Nile tilapia at a suboptimal low temperature. Aquaculture 2019, 507, 500–509. [Google Scholar] [CrossRef]
- Reboleira, J.; Felix, R.; Felix, C.; de Melo, M.M.R.; Silva, C.M.; Saraiva, J.A.; Bandarra, N.M.; Teixeira, B.; Mendes, R.; Paulo, M.C.; et al. Evaluating the Potential of the Defatted By-Product of Aurantiochytrium sp. Industrial Cultivation as a Functional Food. Foods 2021, 10, 3058. [Google Scholar] [CrossRef]
- Trovao, M.; Pereira, H.; Costa, M.; Machado, A.; Barros, A.; Soares, M.; Carvalho, B.; Silva, J.T.; Varela, J.; Silva, J. Lab-Scale Optimization of Aurantiochytrium sp. Culture Medium for Improved Growth and DHA Production. Appl. Sci. 2020, 10, 2500. [Google Scholar] [CrossRef]
- Guimaraes, A.M.; Schleder, D.D.; Nagata, M.; Nobrega, R.O.; Fracalossi, D.M.; Seiffert, W.Q.; Vieira, F.D.N. Aurantiochytrium sp. meal can replace fish oil in practical diets for the juvenile Pacific white shrimp. Aquac. Nutr. 2019, 25, 798–807. [Google Scholar] [CrossRef]
- Yu, X.-J.; Huang, C.-Y.; Chen, H.; Wang, D.-S.; Chen, J.-L.; Li, H.-J.; Liu, X.-Y.; Wang, Z.; Sun, J.; Wang, Z.-P. High-Throughput Biochemical Fingerprinting of Oleaginous Aurantiochytrium sp. Strains by Fourier Transform Infrared Spectroscopy (FT-IR) for Lipid and Carbohydrate Productions. Molecules 2019, 24, 1593. [Google Scholar] [CrossRef]
- Shinmen, Y.; Kawashima, H.; Shimizu, S.; Yamada, H. Concentration of eicosapentaenoic acid and docosahexaenoic acid in an arachidonic acid-producing fungus, Mortierella alpina 1S-4, grown with fish oil. Appl. Microbiol. Biotechnol. 1992, 38, 301–304. [Google Scholar] [CrossRef]
- Yin, F.-W.; Zhan, C.-T.; Huang, J.; Sun, X.-L.; Yin, L.-F.; Zheng, W.-L.; Luo, X.; Zhang, Y.-Y.; Fu, Y.-Q. Efficient Co-production of Docosahexaenoic Acid Oil and Carotenoids in Aurantiochytrium sp. Using a Light Intensity Gradient Strategy. Appl. Biochem. Biotechnol. 2022, 195, 623–638. [Google Scholar] [CrossRef]
- Ji, X.-J.; Zhang, A.-H.; Nie, Z.-K.; Wu, W.-J.; Ren, L.-J.; Huang, H. Efficient arachidonic acid-rich oil production by Mortierella alpina through a repeated fed-batch fermentation strategy. Bioresour. Technol. 2014, 170, 356–360. [Google Scholar] [CrossRef]
- Xiangyu, L.; Chao, Y.; Jianming, Y.; Zhiming, W.; Shuhuan, L. An Online Respiratory Quotient-Feedback Strategy of Feeding Yeast Extract for Efficient Arachidonic Acid Production by Mortierella alpina. Front. Bioeng. Biotechnol. 2017, 5, 83. [Google Scholar]
- Yin, F.-W.; Zhang, Y.-T.; Jiang, J.-Y.; Guo, D.-S.; Gao, S.; Gao, Z. Efficient docosahexaenoic acid production by Schizochytrium sp. via a two-phase pH control strategy using ammonia and citric acid as pH regulators. Process. Biochem. 2019, 77, 1–7. [Google Scholar] [CrossRef]
- Dzurendova, S.; Losada, C.B.; Dupuy-Galet, B.X.; Fjær, K.; Shapaval, V. Mucoromycota fungi as powerful cell factories for modern biorefinery. Appl. Microbiol. Biotechnol. 2022, 106, 101–115. [Google Scholar] [CrossRef]
- Campos-Takaki, G.M.; Dietrich, S.M.C. Characterization of Cell Walls from Mucoralean Fungi by Biochemical Composition, Transmission Electron Microscopy and X-Ray Microanalysis. In Current Research Topics in Applied Microbiology and Microbial Biotechnology; World Scientific: Singapore, 2009; pp. 121–125. [Google Scholar]
- Glencross, B.; Huyben, D.; Schrama, J. The Application of Single-Cell Ingredients in Aquaculture Feeds—A Review. Fishes 2020, 5, 22. [Google Scholar] [CrossRef]
- Yen, S.-W.; Nagarajan, D.; Chen, W.-H.; Lee, D.-J.; Chang, J.-S. Fermentative production of astaxanthin from sorghum distillery residue by an indigenous Aurantiochytrium sp. CJ6 strain using a continuous-feeding fed-batch process. Bioresour. Technol. 2023, 376, 128817. [Google Scholar] [CrossRef]
- Chang, L.; Lu, H.; Chen, H.; Tang, X.; Zhao, J.; Zhang, H.; Chen, Y.Q.; Chen, W. Lipid metabolism research in oleaginous fungus Mortierella alpina: Current progress and future prospects. Biotechnol. Adv. 2022, 54, 107794. [Google Scholar] [CrossRef]
- Prabhakaran, P.; Raethong, N.; Thananusak, R.; Nazir, M.Y.M.; Sapkaew, C.; Soommat, P.; Kingkaw, A.; Hamid, A.A.; Vongsangnak, W.; Song, Y. Revealing holistic metabolic responses associated with lipid and docosahexaenoic acid (DHA) production in Aurantiochytrium sp. SW1. Biochim. Et Biophys. Acta (BBA) Mol. Cell Biol. Lipids 2023, 1868, 159306. [Google Scholar] [CrossRef]
- Yin, F.-W.; Zhu, S.-Y.; Guo, D.-S.; Ren, L.-J.; Ji, X.-J.; Huang, H.; Gao, Z. Development of a strategy for the production of do-cosahexaenoic acid by Schizochytrium sp. from cane molasses and algae-residue. Bioresour. Technol. 2019, 271, 118–124. [Google Scholar] [CrossRef]
- Koizumi, K.; Higashiyama, K.; Park, E.Y. Effects of amino acid on morphological development and nucleus formation of arachidonic acid-producing filamentous micro-organism, Mortierella alpina. J. Appl. Microbiol. 2006, 100, 885–892. [Google Scholar] [CrossRef]
- Valdebenito, D.; Urrutia, S.; Leyton, A.; Chisti, Y.; Asenjo, J.A.; Shene, C. Nitrogen sources affect the long-chain polyunsatu-rated fatty acids content in Thraustochytrium sp. RT2316-16. Mar. Drugs 2022, 21, 15. [Google Scholar] [CrossRef]
- Lu, H.; Chen, H.; Tang, X.; Yang, Q.; Zhang, H.; Chen, Y.Q.; Chen, W. Time-resolved multi-omics analysis reveals the role of nutrient stress-induced resource reallocation for TAG accumulation in oleaginous fungus Mortierella alpina. Biotechnol. Biofuels 2020, 13, 117. [Google Scholar] [CrossRef]
- Ling, X.-P.; Zeng, S.-Y.; Chen, C.-X.; Liu, X.-T.; Lu, Y.-H. Enhanced arachidonic acid production using a bioreactor culture of Mortierella alpina with a combined organic nitrogen source. Bioresour. Bioprocess. 2016, 3, 43. [Google Scholar] [CrossRef]
- Yu, Y.; Zhang, L.; Li, T.; Wu, N.; Jiang, L.; Ji, X.; Huang, H. How nitrogen sources influence Mortierella alpina aging: From the lipid droplet proteome to the whole-cell proteome and metabolome. J. Proteom. 2018, 179, 140–149. [Google Scholar] [CrossRef]
- Lu, J.; Peng, C.; Ji, X.-J.; You, J.; Cong, L.; Ouyang, P.; Huang, H. Fermentation Characteristics of Mortierella alpina in Response to Different Nitrogen Sources. Appl. Biochem. Biotechnol. 2011, 164, 979–990. [Google Scholar] [CrossRef] [PubMed]
- Nisha, A.; Venkateswaran, G. Effect of Culture Variables on Mycelial Arachidonic acid Production by Mortierella alpina. Food Bioprocess Technol. 2008, 4, 232–240. [Google Scholar] [CrossRef]
- Li, C.; Zhang, G.; Zhang, Z.; Ma, D.; Xu, G. Alkaline thermal pretreatment at mild temperatures for biogas production from anaerobic digestion of antibiotic mycelial residue. Bioresour. Technol. 2016, 208, 49–57. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Zhu, X.; Ye, H.; Wen, Y.; Sen, B.; Wang, G. Low dissolved oxygen supply functions as a global regulator of the growth and metabolism of Aurantiochytrium sp. PKU#Mn16 in the early stages of docosahexaenoic acid fermentation. Microb. Cell Factories 2023, 22, 52. [Google Scholar]
- Tao, Z.; Yuan, H.; Liu, M.; Liu, Q.; Zhang, S.; Liu, H.; Jiang, Y.; Huang, D.; Wang, T. Yeast Extract: Characteristics, Production, Applications and Future Perspectives. J. Microbiol. Biotechnol. 2023, 33, 151. [Google Scholar] [CrossRef]
- Asachi, R.; Karimi, K. Enhanced ethanol and chitosan production from wheat straw by Mucor indicus with minimal nutrient consumption. Process. Biochem. 2013, 48, 1524–1531. [Google Scholar] [CrossRef]
- Zhang, H.; Cui, Q.; Song, X. Research advances on arachidonic acid production by fermentation and genetic modification of Mortierella alpina. World J. Microbiol. Biotechnol. 2021, 37, 4. [Google Scholar] [CrossRef]
- Liu, L.; Wang, F.; Yang, J.; Li, X.; Cui, J.; Liu, J.; Shi, M.; Wang, K.; Chen, L.; Zhang, W. Nitrogen Feeding Strategies and Metabolomic Analysis To Alleviate High-Nitrogen Inhibition on Docosahexaenoic Acid Production in Crypthecodinium cohnii. J. Agric. Food Chem. 2018, 66, 10640–10650. [Google Scholar] [CrossRef]
- Song, X.; Zang, X.; Zhang, X. Production of High Docosahexaenoic Acid by Schizochytrium sp. Using Low-cost Raw Materials from Food Industry. J. Oleo Sci. 2015, 64, 197–204. [Google Scholar] [CrossRef]
- Cai, G.; Moghaddam, L.; O’Hara, I.M.; Zhang, Z. Microbial oil production from acidified glycerol pretreated sugarcane bagasse by Mortierella isabellina. RSC Adv. 2019, 9, 2539–2550. [Google Scholar] [CrossRef]
- Samadlouie, H.R.; Nurmohamadi, S.; Moradpoor, F.; Gharanjik, S. Effect of low-cost substrate on the fatty acid profiles of Mortierella alpina CBS 754.68 and Wickerhamomyces siamensis SAKSG. Biotechnol. Biotechnol. Equip. 2018, 32, 1228–1235. [Google Scholar] [CrossRef]
- Xu, J.; Zhu, Y.; Li, H.; Chen, L.; Chen, W.; Cui, M.; Han, L.; Hou, W.; Li, D. Alanine mother liquor as a nitrogen source for docosahexaenoic acid production by Schizochytrium sp. B4D1. Electron. J. Biotechnol. 2018, 35, 10–17. [Google Scholar] [CrossRef]
- Russo, G.L.; Langellotti, A.L.; Sacchi, R.; Masi, P. Techno-economic assessment of DHA-rich Aurantiochytrium sp. production using food industry by-products and waste streams as alternative growth media. Bioresour. Technol. Rep. 2022, 18, 100997. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).