Integrative System Biology Analyses Identify Seven MicroRNAs to Predict Heart Failure


{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
2.1. Analysis of the Protein-MiRNA Network Derived from Post-MI Rats Identified Circulating MiR-21-5p, MiR-23a-3p and MiR-222-3p to Be Associated with LVR after MI
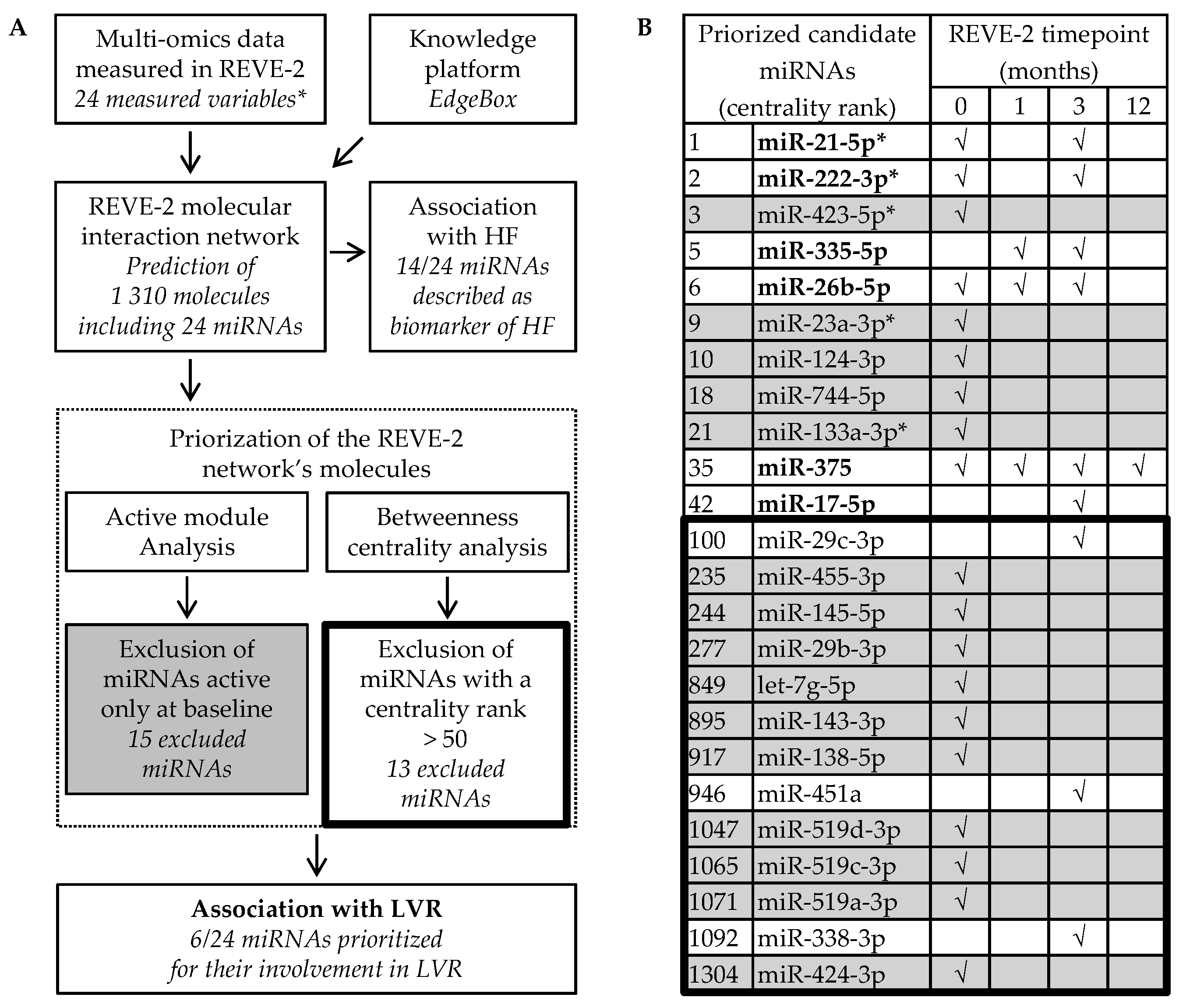
2.2. Analysis of the REVE-2 Network Identified MiR-21-5p, MiR-222-3p, MiR-335-5p, MiR-26b-5p, MiR-375 and MiR-17-5p to Detect LVR after MI
2.3. Gene Ontology Analysis of the 7 MiRNAs Targets Predicted with a High Relation Specificity with Processes Involved in LVR after MI
3. Discussion and Perspectives
4. Methods
4.1. Experimental Model of HF in Rats
4.2. The REVE-2 Study
4.3. Quantification of Candidate MiRNAs
4.4. Functional Analysis of MiRNAs Targets
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Benjamin, E.J.; Blaha, M.J.; Chiuve, S.E.; Cushman, M.; Das, S.R.; Deo, R.; de Ferranti, S.D.; Floyd, J.; Fornage, M.; Gillespie, C.; Isasi, C.R.; et al. Heart Disease and Stroke Statistics—2017 Update: A Report From the American Heart Association. Circulation 2017, 135, e146–e603. [Google Scholar] [CrossRef] [PubMed]
- Savoye, C.; Equine, O.; Tricot, O.; Nugue, O.; Segrestin, B.; Sautière, K.; Elkohen, M.; Pretorian, E.M.; Taghipour, K.; Philias, A.; et al. Left ventricular remodeling after anterior wall acute myocardial infarction in modern clinical practice (from the REmodelage VEntriculaire [REVE] study group). Am. J. Cardiol. 2006, 98, 1144–1149. [Google Scholar] [CrossRef] [PubMed]
- Fertin, M.; Hennache, B.; Hamon, M.; Ennezat, P.V.; Biausque, F.; Elkohen, M.; Nugue, O.; Tricot, O.; Lamblin, N.; Pinet, F.; et al. Usefulness of serial assessment of B-type natriuretic peptide; troponin I; and C-reactive protein to predict left ventricular remodeling after acute myocardial infarction (from the REVE-2 Study). Am. J. Cardiol. 2010, 106, 1410–1416. [Google Scholar] [CrossRef] [PubMed]
- St John Sutton, M.G.; Sharpe, N. Left ventricular remodeling after myocardial infarction pathophysiology and therapy. Circulation 2000, 101, 2981–2988. [Google Scholar] [CrossRef]
- Azuaje, F.J.; Dewey, F.E.; Brutsaert, D.L.; Devaux, Y.; Ashley, E.A.; Wagner, D.R. Systems-based approaches to cardiovascular biomarker discovery. Circ. Cardiovasc. Genet. 2012, 5, 360–367. [Google Scholar] [CrossRef] [PubMed]
- Mitra, K.; Carvunis, A.; Ramesh, S.; Ideker, T. Integrative approaches for finding modular structure in biological networks. Nat. Rev. Genet. 2013, 14, 719–732. [Google Scholar] [CrossRef] [PubMed]
- Winter, J.; Jung, S.; Keller, S.; Gregory, R.I.; Diederichs, S. Many roads to maturity: MicroRNA biogenesis pathways and their regulation. Nat. Cell Biol. 2009, 11, 228–234. [Google Scholar] [CrossRef] [PubMed]
- Bang, C.; Batkai, S.; Dangwal, S.; Gupta, S.K.; Foinquinos, A.; Holzmann, A.; Just, A.; Remke, J.; Zimmer, K.; Zeug, A.; et al. Cardiac fibroblast-derived microRNA passenger strand-enriched exosomes mediate cardiomyocyte hypertrophy. J. Clin. Investig. 2014, 124, 2136–2146. [Google Scholar] [CrossRef] [PubMed]
- Tallquist, M.D.; Molkentin, J.D. Redefining the identity of cardiac fibroblasts. Nat. Rev. Cardiol. 2017, 14, 484–491. [Google Scholar] [CrossRef] [PubMed]
- Dubois-Deruy, E.; Cuvelliez, M.; Fiedler, J.; Charrier, H.; Mulder, P.; Hebbar, E.; Pfanne, A.; Beseme, O.; Chwastyniak, M.; Amouyel, P.; et al. MicroRNAs regulating superoxide dismutase 2 are new circulating biomarkers of heart failure. Sci. Rep. 2017, 7, 14747. [Google Scholar] [CrossRef] [PubMed]
- Mulder, P.; Devaux, B.; Richard, V.; Henry, J.; Wimart, M.; Thibout, E.; Mace, B.; Thuillez, C. Early Versus Delayed Angiotensin-Converting Enzyme Inhibition in Experimental Chronic Heart Failure. Effects on Survival; Hemodynamics; and Cardiovascular Remodeling. Circulation 1997, 95, 1314–1319. [Google Scholar] [CrossRef] [PubMed]
- Cieniewski-Bernard, C.; Mulder, P.; Henry, J.P.; Drobecq, H.; Dubois, E.; Pottiez, G.; Thuillez, C.; Amouyel, P.; Richard, V.; Pinet, F. Proteomic analysis of left ventricular remodeling in an experimental model of heart failure. J. Proteome Res. 2008, 7, 5004–5016. [Google Scholar] [CrossRef] [PubMed]
- Dubois, E.; Richard, V.; Mulder, P.; Lamblin, N.; Drobecq, H.; Henry, J.P.; Amouyel, P.; Thuillez, C.; Bauters, C.; Pinet, F. Decreased Serine207 phosphorylation of troponin T as a biomarker for left ventricular remodelling after myocardial infarction. Eur. Heart J. 2011, 32, 115–123. [Google Scholar] [CrossRef] [PubMed]
- Pinet, F.; Cuvelliez, M.; Kelder, T.; Amouyel, P.; Radonjic, M.; Bauters, C. Integrative network analysis reveals time-dependent molecular events underlying left ventricular remodeling in post-myocardial infarction patients. Biochim. Biophys. Acta 2017, 1863, 1445–1453. [Google Scholar] [CrossRef] [PubMed]
- Kuosmanen, S.M.; Hartikainen, J.; Hippelainen, M.; Kokki, H.; Levonen, A.L.; Tavi, P. MicroRNA profiling of pericardial fluid samples from patients with heart failure. PLoS ONE 2015, 10, e0119646. [Google Scholar] [CrossRef] [PubMed]
- Cortez-Dias, N.; Costa, M.C.; Carrilho-Ferreira, P.; Silva, D.; Jorge, C.; Calisto, C.; Pessoa, T.; Martins, S.R.; de Sousa, J.C.; da Silva, P.C.; et al. Circulating miR-122-5p/miR-133b Ratio Is a Specific Early Prognostic Biomarker in Acute Myocardial Infarction. Circ. J. 2016, 80, 2183–2191. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Meng, H.; Jiang, C.; Yang, S.; Cui, F.; Yang, P. Differential microRNA Expression and Regulation in the Rat Model of Post-Infarction Heart Failure. PLoS ONE 2016, 11, e0160920. [Google Scholar] [CrossRef] [PubMed]
- Marques, F.Z.; Vizi, D.; Khammy, O.; Mariani, J.A.; Kaye, D.M. The transcardiac gradient of cardio-microRNAs in the failing heart. Eur. J. Heart Fail 2016, 18, 1000–1008. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Cheng, Y.J.; Sara, J.D.; Liu, L.J.; Liu, L.P.; Zhao, X.; Gao, H. Circulating MicroRNA-145 is Associated with Acute Myocardial Infarction and Heart Failure. Chin. Med. J. (Engl.) 2017, 130, 51–56. [Google Scholar] [CrossRef] [PubMed]
- Van Boven, N.; Kardys, I.; van Vark, L.C.; Akkerhuis, K.M.; de Ronde, M.W.J.; Khan, M.A.F.; Merkus, D.; Liu, Z.; Voors, A.A.; Asselbergs, F.W.; et al. Serially measured circulating microRNAs and adverse clinical outcomes in patients with acute heart failure. Eur. J. Heart Fail 2018, 20, 89–96. [Google Scholar] [CrossRef] [PubMed]
- Jiao, M.; You, H.Z.; Yang, X.Y.; Yuan, H.; Li, Y.L.; Liu, W.X.; Jin, M.; Du, J. Circulating microRNA signature for the diagnosis of childhood dilated cardiomyopathy. Sci. Rep. 2018, 8, 724. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Liu, J.; Xu, B.; Liu, Y.L.; Liu, Z. Reduced exosome miR-425 and miR-744 in the plasma represents the progression of fibrosis and heart failure. Kaohsiung J. Med. Sci. 2018, 34, 626–633. [Google Scholar] [CrossRef] [PubMed]
- Jakob, P.; Kacprowski, T.; Briand-Schumacher, S.; Heg, D.; Klingenberg, R.; Stahli, B.E.; Jaguszewski, M.; Rodondi, N.; Nanchen, D.; Raber, L.; et al. Profiling and validation of circulating microRNAs for cardiovascular events in patients presenting with ST-segment elevation myocardial infarction. Eur Heart J 2017, 38, 511–515. [Google Scholar] [CrossRef] [PubMed]
- Fang, L.; Ellims, A.H.; Moore, X.; White, D.A.; Taylor, A.J.; Chin-Dusting, J.; Dart, A.M. Circulating microRNAs as biomarkers for diffuse myocardial fibrosis in patients with hypertrophic cardiomyopathy. J. Transl. Med. 2015, 13, 314. [Google Scholar] [CrossRef] [PubMed]
- Bindea, G.; Mlecnik, B.; Hackl, H.; Charoentong, P.; Tosolini, M.; Kirilovsky, A.; Fridman, W.H.; Pagès, F.; Trajanoski, Z.; Galon, J. ClueGO: A Cytoscape plug-in to decipher functionally grouped gene ontology and pathway annotation networks. Bioinformatics 2009, 25, 1091–1093. [Google Scholar] [CrossRef] [PubMed]
- Bindea, G.; Galon, J.; Mlecnik, B. CluePedia Cytoscape plugin: Pathway insights using integrated experimental and in silico data. Bioinformatics 2013, 29, 661–663. [Google Scholar] [CrossRef] [PubMed]
- Bray, D. Limits of computational biology. In Silico Biol. 2015, 12, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Oulas, A.; Minadakis, G.; Zachariou, M.; Sokratous, K.; Bourdakou, M.M.; Spyrou, G.M. Systems Bioinformatics: Increasing precision of computational diagnostics and therapeutics through network-based approaches. Brief. Bioinform. 2017, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Vegter, E.L.; Ovchinnikova, E.S.; Sillj, H.H.W.; Meems, L.M.G.; van der Pol, A.; van der Velde, A.R.; Berezikov, E.; Voors, A.A.; Boer, R.A.D.; van der Meer, P. Rodent heart failure models do not reflect the human circulating microRNA signature in heart failure. PLoS ONE 2017, 12, e0177242. [Google Scholar] [CrossRef] [PubMed]
- Sarrion, I.; Milian, L.; Juan, G.; Ramon, M.; Furest, I.; Carda, C.; Cortijo Gimeno, J.; Mata Roig, M. Role of circulating miRNAs as biomarkers in idiopathic pulmonary arterial hypertension: Possible relevance of miR-23a. Oxid. Med. Cell. Longev. 2015, 2015, 792846. [Google Scholar] [CrossRef] [PubMed]
- Long, B.; Gan, T.Y.; Zhang, R.C.; Zhang, Y.H. miR-23a Regulates Cardiomyocyte Apoptosis by Targeting Manganese Superoxide Dismutase. Mol. Cells 2017, 40, 542–549. [Google Scholar] [CrossRef] [PubMed]
- Verjans, R.; Peters, T.; Beaumont, F.J.; van Leeuwen, R.; van Herwaarden, T.; Verhesen, W.; Munts, C.; Bijnen, M.; Henkens, M.; Diez, J.; et al. MicroRNA-221/222 Family Counteracts Myocardial Fibrosis in Pressure Overload-Induced Heart Failure. Hypertension 2018, 71, 280–288. [Google Scholar] [CrossRef] [PubMed]
- Allen-Rhoades, W.; Kurenbekova, L.; Satterfield, L.; Parikh, N.; Fuja, D.; Shuck, R.L.; Rainusso, N.; Trucco, M.; Barkauskas, D.A.; Jo, E.; et al. Cross-species identification of a plasma microRNA signature for detection, therapeutic monitoring; and prognosis in osteosarcoma. Cancer Med. 2015, 4, 977–988. [Google Scholar] [CrossRef] [PubMed]
- Kocijan, R.; Muschitz, C.; Geiger, E.; Skalicky, S.; Baierl, A.; Dormann, R.; Plachel, F.; Feichtinger, X.; Heimel, P.; Fahrleitner-Pammer, A.; et al. Circulating microRNA signatures in patients with idiopathic and postmenopausal osteoporosis and fragility fractures. J. Clin. Endocrinol. Metab. 2016, 101, 4125–4134. [Google Scholar] [CrossRef] [PubMed]
- Cao, L.; Kong, L.P.; Yu, Z.B.; Han, S.P.; Bai, Y.F.; Zhu, J.; Hu, X.; Zhu, C.; Zhu, S.; Guo, X.R. microRNA expression profiling of the developing mouse heart. Int. J. Mol. Med. 2012, 30, 1095–1104. [Google Scholar] [CrossRef] [PubMed]
- Zhu, S.; Cao, L.; Zhu, J.; Kong, L.; Jin, J.; Qian, L.; Zhu, C.; Hu, X.; Li, M.; Guo, X.; et al. Identification of maternal serum microRNAs as novel non-invasive biomarkers for prenatal detection of fetal congenital heart defects. Clin. Chim. Acta 2013, 424, 66–72. [Google Scholar] [CrossRef] [PubMed]



© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Charrier, H.; Cuvelliez, M.; Dubois-Deruy, E.; Mulder, P.; Richard, V.; Bauters, C.; Pinet, F. Integrative System Biology Analyses Identify Seven MicroRNAs to Predict Heart Failure. Non-Coding RNA 2019, 5, 22. https://doi.org/10.3390/ncrna5010022
Charrier H, Cuvelliez M, Dubois-Deruy E, Mulder P, Richard V, Bauters C, Pinet F. Integrative System Biology Analyses Identify Seven MicroRNAs to Predict Heart Failure. Non-Coding RNA. 2019; 5(1):22. https://doi.org/10.3390/ncrna5010022
Chicago/Turabian StyleCharrier, Henri, Marie Cuvelliez, Emilie Dubois-Deruy, Paul Mulder, Vincent Richard, Christophe Bauters, and Florence Pinet. 2019. "Integrative System Biology Analyses Identify Seven MicroRNAs to Predict Heart Failure" Non-Coding RNA 5, no. 1: 22. https://doi.org/10.3390/ncrna5010022
APA StyleCharrier, H., Cuvelliez, M., Dubois-Deruy, E., Mulder, P., Richard, V., Bauters, C., & Pinet, F. (2019). Integrative System Biology Analyses Identify Seven MicroRNAs to Predict Heart Failure. Non-Coding RNA, 5(1), 22. https://doi.org/10.3390/ncrna5010022