


Building a Hand-Curated ceRNET for Endometrial Cancer, Striving for Clinical as Well as Medicolegal Soundness: A Systematic Review

 ,
,  ,
,  ,
,  and
and 
Abstract
1. Introduction
2. Results
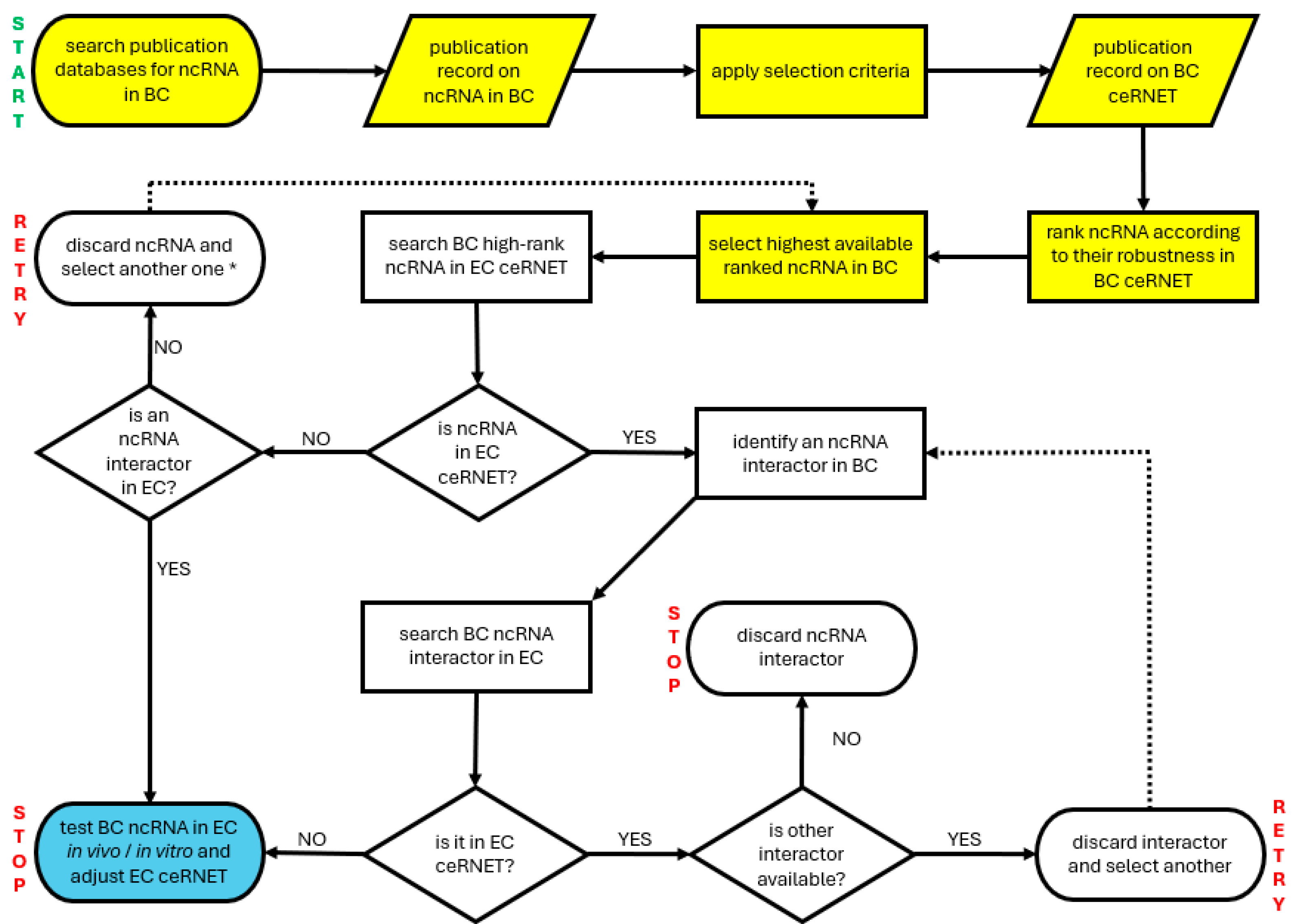
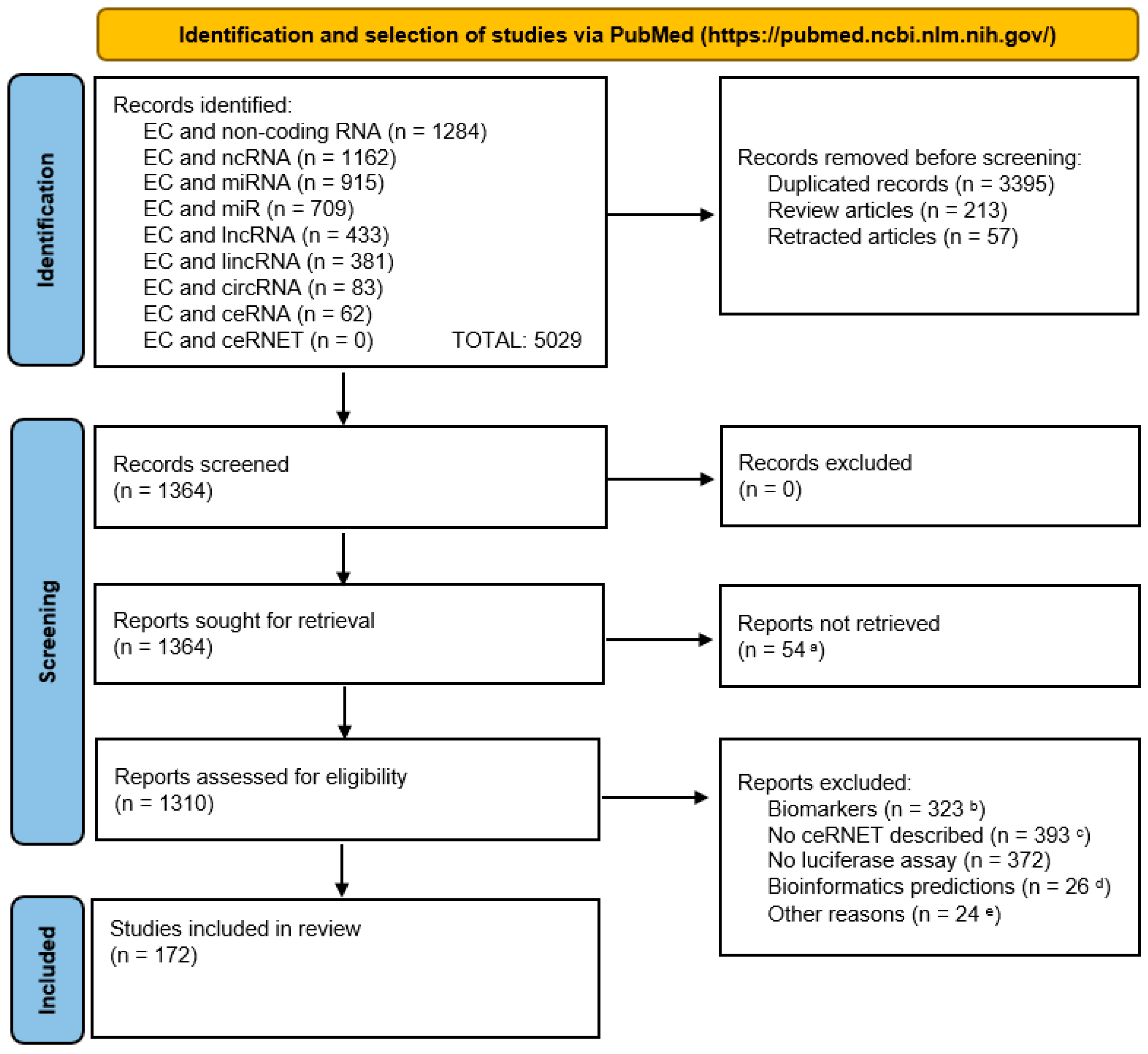
2.1. Data Collection
2.2. Data Assembly
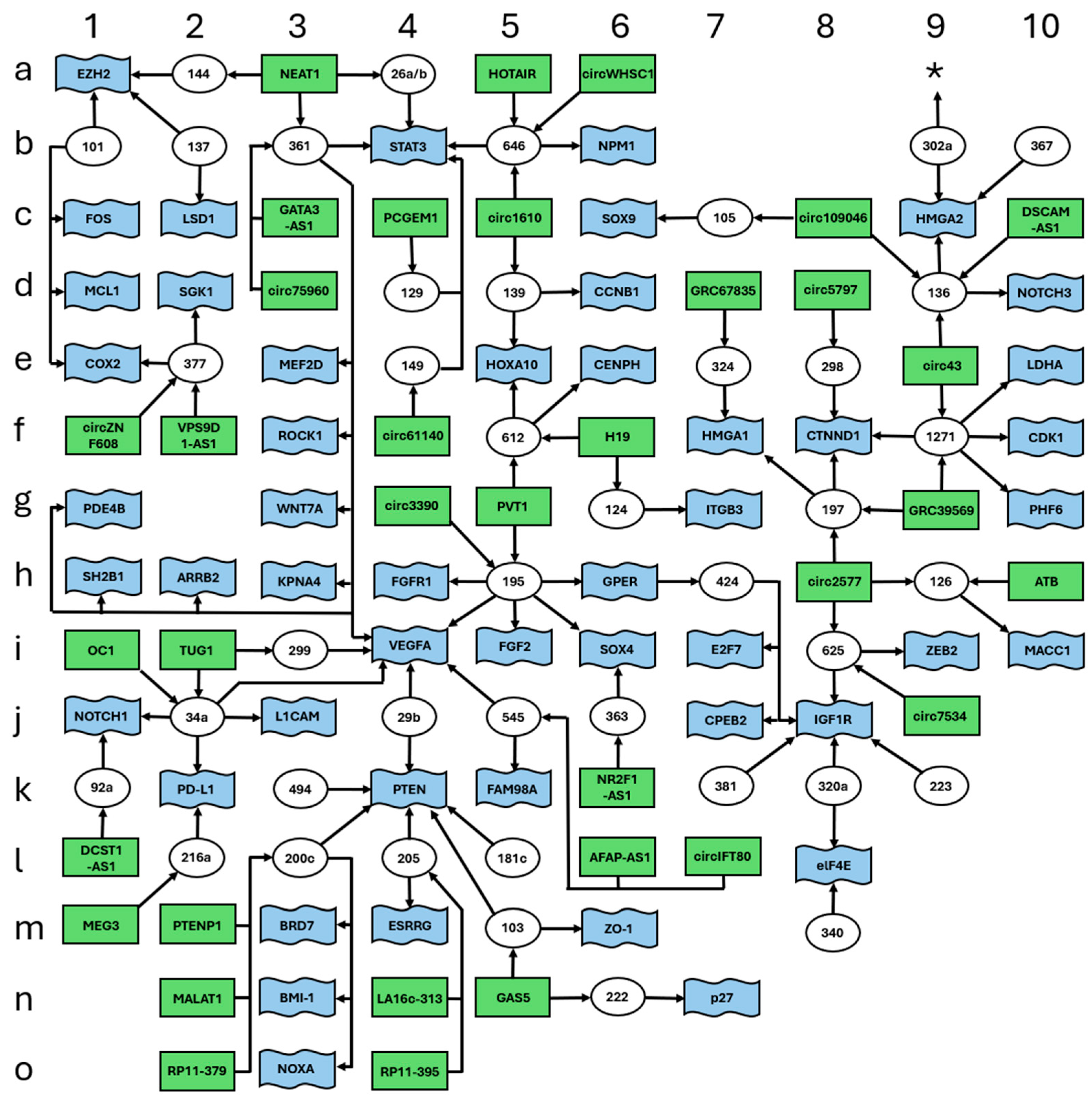
3. Discussion
3.1. General Considerations
3.2. Using Our ceRNET for Investigating Data Robustness
3.3. Using Our ceRNET for Investigating Metabolic Changes in EC
3.4. Using External Data to Expand EC ceRNET
3.5. Automation of ceRNET Construction Through Artificial Intelligence (AI) and Construction of a Multi-Cancer ceRNET
3.6. Importance of EC ceRNET for Personalized Therapy: Implications for Precision Medicine
3.7. Legal and Ethical Aspects of Using ceRNET in Cancer Therapy
Medicolegal Tenability Needs Updated Frames of Reference
4. Materials and Methods
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Belladelli, F.; Montorsi, F.; Martini, A. Metabolic Syndrome, Obesity and Cancer Risk. Curr. Opin. Urol. 2022, 32, 594–597. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.-S.; Scherer, P.E. Obesity, Diabetes, and Increased Cancer Progression. Diabetes Metab. J. 2021, 45, 799–812. [Google Scholar] [CrossRef] [PubMed]
- Ignatov, A.; Ortmann, O. Endocrine Risk Factors of Endometrial Cancer: Polycystic Ovary Syndrome, Oral Contraceptives, Infertility, Tamoxifen. Cancers 2020, 12, 1766. [Google Scholar] [CrossRef]
- Wu, Y.; Sun, W.; Liu, H.; Zhang, D. Age at Menopause and Risk of Developing Endometrial Cancer: A Meta-Analysis. Biomed. Res. Int. 2019, 2019, 8584130. [Google Scholar] [CrossRef]
- Ali, A.T. Reproductive Factors and the Risk of Endometrial Cancer. Int. J. Gynecol. Cancer 2014, 24, 384–393. [Google Scholar] [CrossRef]
- Idos, G.; Valle, L. Lynch Syndrome. In GeneReviews® [Internet]; Adam, M.P., Feldman, J., Mirzaa, G.M., Pagon, R.A., Wallace, S.E., Amemiya, A., Eds.; University of Washington: Seattle, WA, USA, 2004; 1993–2025. [Google Scholar] [PubMed]
- Levine, D.A. The Cancer Genome Atlas Research Network Integrated Genomic Characterization of Endometrial Carcinoma. Nature 2013, 497, 67–73. [Google Scholar] [CrossRef]
- Bianco, B.; Barbosa, C.P.; Trevisan, C.M.; Laganà, A.S.; Montagna, E. Endometrial Cancer: A Genetic Point of View. Transl. Cancer Res. 2020, 9, 7706–7715. [Google Scholar] [CrossRef]
- Bafligil, C.; Thompson, D.J.; Lophatananon, A.; Smith, M.J.; Ryan, N.A.; Naqvi, A.; Evans, D.G.; Crosbie, E.J. Association between Genetic Polymorphisms and Endometrial Cancer Risk: A Systematic Review. J. Med. Genet. 2020, 57, 591–600. [Google Scholar] [CrossRef]
- Benati, M.; Montagnana, M.; Danese, E.; Mazzon, M.; Paviati, E.; Garzon, S.; Laganà, A.S.; Casarin, J.; Giudici, S.; Raffaelli, R.; et al. Aberrant Telomere Length in Circulating Cell-Free DNA as Possible Blood Biomarker with High Diagnostic Performance in Endometrial Cancer. Pathol. Oncol. Res. 2020, 26, 2281–2289. [Google Scholar] [CrossRef]
- Kaneko, E.; Sato, N.; Sugawara, T.; Noto, A.; Takahashi, K.; Makino, K.; Terada, Y. MLH1 Promoter Hypermethylation Predicts Poorer Prognosis in Mismatch Repair Deficiency Endometrial Carcinomas. J. Gynecol. Oncol. 2021, 32, e79. [Google Scholar] [CrossRef] [PubMed]
- Loukovaara, M.; Pasanen, A.; Bützow, R. Mismatch Repair Protein and MLH1 Methylation Status as Predictors of Response to Adjuvant Therapy in Endometrial Cancer. Cancer Med. 2021, 10, 1034–1042. [Google Scholar] [CrossRef]
- Lakhotia, S.C. C-Value Paradox: Genesis in Misconception That Natural Selection Follows Anthropocentric Parameters of ‘Economy’ and ‘Optimum’. BBA Adv. 2023, 4, 100107. [Google Scholar] [CrossRef]
- Choi, I.-Y.; Kwon, E.-C.; Kim, N.-S. The C- and G-Value Paradox with Polyploidy, Repeatomes, Introns, Phenomes and Cell Economy. Genes Genom. 2020, 42, 699–714. [Google Scholar] [CrossRef]
- Singh, R.S.; Gupta, B.P. Genes and Genomes and Unnecessary Complexity in Precision Medicine. NPJ Genom. Med. 2020, 5, 21. [Google Scholar] [CrossRef]
- Palazzo, A.F.; Lee, E.S. Non-Coding RNA: What Is Functional and What Is Junk? Front. Genet. 2015, 6, 2. [Google Scholar] [CrossRef] [PubMed]
- Naeli, P.; Winter, T.; Hackett, A.P.; Alboushi, L.; Jafarnejad, S.M. The Intricate Balance between MicroRNA-Induced MRNA Decay and Translational Repression. FEBS J. 2023, 290, 2508–2524. [Google Scholar] [CrossRef] [PubMed]
- Friedman, R.C.; Farh, K.K.H.; Burge, C.B.; Bartel, D.P. Most Mammalian MRNAs Are Conserved Targets of MicroRNAs. Genome Res. 2009, 19, 92. [Google Scholar] [CrossRef]
- Shu, J.; Resende E Silva, B.V.; Gao, T.; Xu, Z.; Cui, J. Dynamic and Modularized MicroRNA Regulation and Its Implication in Human Cancers. Sci. Rep. 2017, 7, 13356. [Google Scholar] [CrossRef]
- Kumar, S.; Gonzalez, E.A.; Rameshwar, P.; Etchegaray, J.P. Non-Coding RNAs as Mediators of Epigenetic Changes in Malignancies. Cancers 2020, 12, 3657. [Google Scholar] [CrossRef]
- Yang, N.; Liu, K.; Yang, M.; Gao, X. CeRNAs in Cancer: Mechanism and Functions in a Comprehensive Regulatory Network. J. Oncol. 2021, 2021, 1–12. [Google Scholar] [CrossRef]
- Vallone, C.; Rigon, G.; Gulia, C.; Baffa, A.; Votino, R.; Morosetti, G.; Zaami, S.; Briganti, V.; Catania, F.; Gaffi, M.; et al. Non-Coding RNAs and Endometrial Cancer. Genes 2018, 9, 187. [Google Scholar] [CrossRef] [PubMed]
- Xia, L.; Wang, Y.; Meng, Q.; Su, X.; Shen, J.; Wang, J.; He, H.; Wen, B.; Zhang, C.; Xu, M. Integrated Bioinformatic Analysis of a Competing Endogenous RNA Network Reveals a Prognostic Signature in Endometrial Cancer. Front. Oncol. 2019, 9, 448. [Google Scholar] [CrossRef] [PubMed]
- Piergentili, R.; Zaami, S.; Cavaliere, A.F.; Signore, F.; Scambia, G.; Mattei, A.; Marinelli, E.; Gulia, C.; Perelli, F. Non-Coding Rnas as Prognostic Markers for Endometrial Cancer. Int. J. Mol. Sci. 2021, 22, 3151. [Google Scholar] [CrossRef] [PubMed]
- Zhao, D.; Ren, C.; Yao, Y.; Wang, Q.; Li, F.; Li, Y.; Jiang, A.; Wang, G. Identifying Prognostic Biomarkers in Endometrial Carcinoma Based on ceRNA Network. J. Cell. Biochem. 2020, 121, 2437–2446. [Google Scholar] [CrossRef]
- Cavaliere, A.F.; Perelli, F.; Zaami, S.; Piergentili, R.; Mattei, A.; Vizzielli, G.; Scambia, G.; Straface, G.; Restaino, S.; Signore, F. Towards Personalized Medicine: Non-Coding Rnas and Endometrial Cancer. Healthcare 2021, 9, 965. [Google Scholar] [CrossRef]
- Ravegnini, G.; Gorini, F.; De Crescenzo, E.; De Leo, A.; De Biase, D.; Di Stanislao, M.; Hrelia, P.; Angelini, S.; De Iaco, P.; Perrone, A.M. Can miRNAs Be Useful Biomarkers in Improving Prognostic Stratification in Endometrial Cancer Patients? An Update Review. Int. J. Cancer 2022, 150, 1077–1090. [Google Scholar] [CrossRef]
- Piergentili, R.; Gullo, G.; Basile, G.; Gulia, C.; Porrello, A.; Cucinella, G.; Marinelli, E.; Zaami, S. Circulating MiRNAs as a Tool for Early Diagnosis of Endometrial Cancer—Implications for the Fertility-Sparing Process: Clinical, Biological, and Legal Aspects. Int. J. Mol. Sci. 2023, 24, 11356. [Google Scholar] [CrossRef]
- Liu, Y.; Li, H.; Zhao, C.; Jia, H. MicroRNA-101 Inhibits Angiogenesis via COX-2 in Endometrial Carcinoma. Mol. Cell. Biochem. 2018, 448, 61–69. [Google Scholar] [CrossRef]
- Konno, Y.; Dong, P.; Xiong, Y.; Suzuki, F.; Lu, J.; Cai, M.; Watari, H.; Mitamura, T.; Hosaka, M.; Hanley, S.J.B.; et al. MicroRNA-101 Targets EZH2, MCL-1 and FOS to Suppress Proliferation, Invasion and Stem Cell-like Phenotype of Aggressive Endometrial Cancer Cells. Oncotarget 2014, 5, 6049–6062. [Google Scholar] [CrossRef]
- Wang, C.; Liu, B. MiR-101-3p Induces Autophagy in Endometrial Carcinoma Cells by Targeting EZH2. Arch. Gynecol. Obstet. 2018, 297, 1539–1548. [Google Scholar] [CrossRef] [PubMed]
- Guo, C.; Song, W.Q.; Sun, P.; Jin, L.; Dai, H.Y. LncRNA-GAS5 Induces PTEN Expression through Inhibiting MiR-103 in Endometrial Cancer Cells. J. Biomed. Sci. 2015, 22, 100. [Google Scholar] [CrossRef] [PubMed]
- Du, J.; Zhang, F.; Zhang, L.; Jia, Y.; Chen, H. MicroRNA-103 Regulates the Progression in Endometrial Carcinoma through ZO-1. Int. J. Immunopathol. Pharmacol. 2019, 33. [Google Scholar] [CrossRef]
- Li, Y.; Liu, J.; Piao, J.; Ou, J.; Zhu, X. Circ_0109046 promotes the malignancy of endometrial carcinoma cells through the microRNA-105/SOX9/Wnt/β-catenin axis. IUBMB Life 2021, 73, 159–176. [Google Scholar] [CrossRef] [PubMed]
- Tang, W.; Li, J.; Liu, H.; Zhou, F.; Liu, M. MiR-106a Promotes Tumor Growth, Migration, and Invasion by Targeting BCL2L11 in Human Endometrial Adenocarcinoma. Am. J. Transl. Res. 2017, 9, 4984–4993. [Google Scholar]
- Zhao, Z.-N.; Bai, J.-X.; Zhou, Q.; Yan, B.; Qin, W.-W.; Jia, L.-T.; Meng, Y.-L.; Jin, B.-Q.; Yao, L.-B.; Wang, T.; et al. TSA Suppresses MiR-106b-93-25 Cluster Expression through Downregulation of MYC and Inhibits Proliferation and Induces Apoptosis in Human EMC. PLoS ONE 2012, 7, e45133. [Google Scholar] [CrossRef]
- Bao, W.; Zhang, Y.; Li, S.; Fan, Q.; Qiu, M.; Wang, Y.; Li, Y.; Ji, X.; Yang, Y.; Sang, Z.; et al. MiR-107-5p Promotes Tumor Proliferation and Invasion by Targeting Estrogen Receptor-α in Endometrial Carcinoma. Oncol. Rep. 2018, 41, 1575–1585. [Google Scholar] [CrossRef]
- Chen, H.; Fan, Y.; Xu, W.; Chen, J.; Xu, C.; Wei, X.; Fang, D.; Feng, Y. MiR-10b Inhibits Apoptosis and Promotes Proliferation and Invasion of Endometrial Cancer Cells via Targeting HOXB3. Cancer Biother. Radiopharm. 2016, 31, 225–231. [Google Scholar] [CrossRef]
- Liu, Y.; Chang, Y.; Cai, Y. CircTNFRSF21, a Newly Identified Circular RNA Promotes Endometrial Carcinoma Pathogenesis through Regulating MiR-1227-MAPK13/ATF2 Axis. Aging 2020, 12, 6774–6792. [Google Scholar] [CrossRef]
- Liu, S.; Qiu, J.; Tang, X.; Cui, H.; Zhang, Q.; Yang, Q. LncRNA-H19 Regulates Cell Proliferation and Invasion of Ectopic Endometrium by Targeting ITGB3 via Modulating MiR-124-3p. Exp. Cell. Res. 2019, 381, 215–222. [Google Scholar] [CrossRef]
- Gu, Z.-R.; Liu, W. The LncRNA AL161431.1 Targets MiR-1252-5p and Facilitates Cellular Proliferation and Migration via MAPK Signaling in Endometrial Carcinoma. Eur. Rev. Med. Pharmacol. Sci. 2020, 24, 2294–2302. [Google Scholar] [PubMed]
- Shi, F.; Wang, T.; Liu, Z.; Zhang, Y.; Wang, J.; Zhang, K.; Su, J. LncRNA miR143HG Up-Regulates p53 In Endometrial Carcinoma By Sponging miR-125a. Cancer Manag. Res. 2019, 11, 10117–10123. [Google Scholar] [CrossRef]
- Shang, C.; Ao, C.N.; Cheong, C.C.; Meng, L. Long Non-Coding RNA CDKN2B Antisense RNA 1 Gene Contributes to Paclitaxel Resistance in Endometrial Carcinoma. Front. Oncol. 2019, 9, 27. [Google Scholar] [CrossRef]
- Zheng, X.; Liu, M.; Song, Y.; Feng, C. Long Noncoding RNA-ATB Impairs the Function of Tumor Suppressor MiR-126-Mediated Signals in Endometrial Cancer for Tumor Growth and Metastasis. Cancer Biother. Radiopharm. 2019, 34, 47–55. [Google Scholar] [CrossRef]
- Wang, W.; Jin, W.; Liu, X.; Zheng, L. Circ_0002577/MiR-126-5p/MACC1 Axis Promotes Endometrial Carcinoma Progression by Regulation of Proliferation, Migration, Invasion, and Apoptosis of Endometrial Carcinoma Cells. Arch. Gynecol. Obstet. 2022, 306, 481–491. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Qu, Y.; Li, Y. Over-Expression of MiR-1271 Inhibits Endometrial Cancer Cells Proliferation and Induces Cell Apoptosis by Targeting CDK1. Eur. Rev. Med. Pharmacol. Sci. 2017, 21, 2816–2822. [Google Scholar]
- Tian, Y.; Chen, Y.; Han, A. MiR-1271 Inhibits Cell Proliferation and Metastasis by Targeting LDHA in Endometrial Cancer. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 5648–5656. [Google Scholar]
- Wei, D.; Tian, M.; Fan, W.; Zhong, X.; Wang, S.; Chen, Y.; Zhang, S. Circular RNA Circ_0000043 Promotes Endometrial Carcinoma Progression by Regulating MiR-1271-5p/CTNND1 Axis. Arch. Gynecol. Obstet. 2021, 303, 1075–1087. [Google Scholar] [CrossRef]
- Li, J.; Zhang, Z.; Hu, Y.; Wei, Q.; Shao, X. Circ_0039569 Contributes to the Paclitaxel Resistance of Endometrial Cancer via Targeting MiR-1271-5p/PHF6 Pathway. Anticancer Drugs 2022, 33, 883–892. [Google Scholar] [CrossRef]
- Chen, S.; Liang, Y.; Shen, Y.; Wang, X. LncRNA XIST/MiR-129-2-3p Axis Targets CCP110 to Regulate the Proliferation, Invasion and Migration of Endometrial Cancer Cells. Exp. Ther. Med. 2023, 25, 159. [Google Scholar] [CrossRef]
- Li, Q.; Shen, F.; Zhao, L. The Relationship between LncRNA PCGEM1 and STAT3 during the Occurrence and Development of Endometrial Carcinoma. Biomed. Pharmacother. 2018, 107, 918–928. [Google Scholar] [CrossRef]
- Dong, P.; Karaayvaz, M.; Jia, N.; Kaneuchi, M.; Hamada, J.; Watari, H.; Sudo, S.; Ju, J.; Sakuragi, N. Mutant P53 Gain-of-Function Induces Epithelial–Mesenchymal Transition through Modulation of the MiR-130b–ZEB1 Axis. Oncogene 2013, 32, 3286–3295. [Google Scholar] [CrossRef] [PubMed]
- Niu, Z.; Wang, Y.; Zhang, Y.; Liu, D.; Wang, J.; Shi, T.; Xu, X.; Li, L. MTFR2 Promotes Endometrial Carcinoma Cell Proliferation and Growth via the MiR-132-3p /PI3K/Akt Signaling Pathway. Front. Med. 2024, 11, 1505071. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Liu, T.; Huang, Y. MicroRNA-134 Suppresses Endometrial Cancer Stem Cells by Targeting POGLUT1 and Notch Pathway Proteins. FEBS Lett. 2015, 589, 207–214. [Google Scholar] [CrossRef] [PubMed]
- Zong, Z.; Liu, Y.; Chen, S.; Zhao, Y. Circ_PUM1 Promotes the Development of Endometrial Cancer by Targeting the MiR-136/NOTCH3 Pathway. J. Cell. Mol. Med. 2020, 24, 4127–4135. [Google Scholar] [CrossRef]
- Shi, Y.; Jia, L.; Wen, H. Circ_0109046 Promotes the Progression of Endometrial Cancer via Regulating MiR-136/HMGA2 Axis. Cancer Manag. Res. 2020, 12, 10993–11003. [Google Scholar] [CrossRef]
- Li, L.; Chen, P.; Huang, B.; Cai, P. LncRNA DSCAM-AS1 Facilitates the Progression of Endometrial Cancer via MiR-136-5p. Oncol. Lett. 2021, 22, 825. [Google Scholar] [CrossRef]
- Zhang, W.; Chen, J.-H.; Shan, T.; Aguilera-Barrantes, I.; Wang, L.-S.; Huang, T.H.-M.; Rader, J.S.; Sheng, X.; Huang, Y.-W. MiR-137 Is a Tumor Suppressor in Endometrial Cancer and Is Repressed by DNA Hypermethylation. Lab. Investig. 2018, 98, 1397–1407. [Google Scholar] [CrossRef]
- Gu, X.; Shi, Y.; Dong, M.; Jiang, L.; Yang, J.; Liu, Z. Exosomal Transfer of Tumor-Associated Macrophage-Derived Hsa_circ_0001610 Reduces Radiosensitivity in Endometrial Cancer. Cell. Death Dis. 2021, 12, 818. [Google Scholar] [CrossRef]
- Liu, J.; Li, C.; Jiang, Y.; Wan, Y.; Zhou, S.; Cheng, W. Tumor-Suppressor Role of MiR-139-5p in Endometrial Cancer. Cancer Cell. Int. 2018, 18, 51. [Google Scholar] [CrossRef]
- Dong, P.; Wang, F.; Taheri, M.; Xiong, Y.; Ihira, K.; Kobayashi, N.; Konno, Y.; Yue, J.; Watari, H. Long Non-Coding RNA TMPO-AS1 Promotes GLUT1-Mediated Glycolysis and Paclitaxel Resistance in Endometrial Cancer Cells by Interacting with MiR-140 and MiR-143. Front. Oncol. 2022, 12, 912935. [Google Scholar] [CrossRef] [PubMed]
- Xiong, H.; Zhang, W.; Xie, M.; Chen, R.; Chen, H.; Lin, Q. Long Non-Coding RNA JPX Promotes Endometrial Carcinoma Progression via Janus Kinase 2/Signal Transducer and Activator of Transcription 3. Front. Oncol. 2024, 14, 1340050. [Google Scholar] [CrossRef]
- Cui, Z.; An, X.; Li, J.; Liu, Q.; Liu, W. LncRNA MIR22HG Negatively Regulates MiR-141-3p to Enhance DAPK1 Expression and Inhibits Endometrial Carcinoma Cells Proliferation. Biomed. Pharmacother. 2018, 104, 223–228. [Google Scholar] [CrossRef]
- Chang, L.; Zhang, D.; Shi, H.; Bian, Y.; Guo, R. MiR-143 Inhibits Endometrial Cancer Cell Proliferation and Metastasis by Targeting MAPK1. Oncotarget 2017, 8, 84384–84395. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Ge, L.; Xu, X.-J.; Yang, T.; Yuan, Y.; Ma, X.-L.; Zhang, X.-H. LncRNA NEAT1 Promotes Endometrial Cancer Cell Proliferation, Migration and Invasion by Regulating the MiR-144-3p/EZH2 Axis. Radiol. Oncol. 2019, 53, 434–442. [Google Scholar] [CrossRef]
- Zhou, X.; Gao, Q.; Wang, J.; Zhang, X.; Liu, K.; Duan, Z. Linc-RNA-RoR Acts as a “Sponge” against Mediation of the Differentiation of Endometrial Cancer Stem Cells by MicroRNA-145. Gynecol. Oncol. 2014, 133, 333–339. [Google Scholar] [CrossRef]
- Sun, G.; Tian, J.; Xiao, Y.; Zeng, Y. Circular RNA Circ_0005667 Promotes Cisplatin Resistance of Endometrial Carcinoma Cells by Regulating IGF2BP1 through MiR-145-5p. Anticancer Drugs. 2022, 34, 816–826. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Lu, W.; Qu, J.; Ye, L.; Du, G.; Wan, X. Loss of Exosomal MiR-148b from Cancer-associated Fibroblasts Promotes Endometrial Cancer Cell Invasion and Cancer Metastasis. J. Cell. Physiol. 2019, 234, 2943–2953. [Google Scholar] [CrossRef]
- Liu, Y.; Chang, Y.; Cai, Y. Hsa_circ_0061140 Promotes Endometrial Carcinoma Progression via Regulating MiR-149-5p/STAT3. Gene 2020, 745, 144625. [Google Scholar] [CrossRef]
- Yang, C.; Zhang, X.; Zhou, L.; Wan, Y.; Song, L.; Gu, W.; Liu, R.; Ma, Y.; Meng, H.; Tian, Y.; et al. LncRNA SNHG8 Participates in the Development of Endometrial Carcinoma through Regulating C-MET Expression by MiR-152. Eur. Rev. Med. Pharmacol. Sci. 2018, 22, 1629–1637. [Google Scholar]
- Xie, D.; Liang, Y.; Su, Y.; An, Y.; Qu, P. MiR-152 Inhibits Proliferation of Human Endometrial Cancer Cells via Inducing G2/M Phase Arrest by Suppressing CDC25B Expression. Biomed. Pharmacother. 2018, 99, 299–305. [Google Scholar] [CrossRef]
- Tsuruta, T.; Kozaki, K.; Uesugi, A.; Furuta, M.; Hirasawa, A.; Imoto, I.; Susumu, N.; Aoki, D.; Inazawa, J. MiR-152 Is a Tumor Suppressor MicroRNA That Is Silenced by DNA Hypermethylation in Endometrial Cancer. Cancer Res. 2011, 71, 6450–6462. [Google Scholar] [CrossRef] [PubMed]
- Fang, Q.; Sang, L.; Du, S. Long Noncoding RNA LINC00261 Regulates Endometrial Carcinoma Progression by Modulating MiRNA/FOXO1 Expression. Cell. Biochem. Funct. 2018, 36, 323–330. [Google Scholar] [CrossRef] [PubMed]
- Choi, C.H.; Park, Y.-A.; Choi, J.-J.; Song, T.; Song, S.Y.; Lee, Y.-Y.; Lee, J.-W.; Kim, T.-J.; Kim, B.-G.; Bae, D.-S. Angiotensin II Type I Receptor and MiR-155 in Endometrial Cancers: Synergistic Antiproliferative Effects of Anti-MiR-155 and Losartan on Endometrial Cancer Cells. Gynecol. Oncol. 2012, 126, 124–131. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.-M.; Wan, X.-H.; Sang, G.-Y.; Zhao, J.-D.; Zhu, Q.-Y.; Wang, D.-M. MiR-15a-5p Suppresses Endometrial Cancer Cell Growth via Wnt/β-Catenin Signaling Pathway by Inhibiting WNT3A. Eur. Rev. Med. Pharmacol. Sci. 2017, 21, 4810–4818. [Google Scholar]
- Liu, D.; Bi, X.; Yang, Y. Circular RNA Hsa_circ_0011324 Is Involved in Endometrial Cancer Progression and the Evolution of Its Mechanism. Bioengineered 2022, 13, 7485–7499. [Google Scholar] [CrossRef]
- Dong, P.; Xiong, Y.; Konno, Y.; Ihira, K.; Kobayashi, N.; Yue, J.; Watari, H. Long Non-Coding RNA DLEU2 Drives EMT and Glycolysis in Endometrial Cancer through HK2 by Competitively Binding with MiR-455 and by Modulating the EZH2/MiR-181a Pathway. J. Exp. Clin. Cancer Res. 2021, 40, 216. [Google Scholar] [CrossRef]
- Yu, J.; Jiang, L.; Gao, Y.; Sun, Q.; Liu, B.; Hu, Y.; Han, X. LncRNA CCAT1 Negatively Regulates MiR-181a-5p to Promote Endometrial Carcinoma Cell Proliferation and Migration. Exp. Ther. Med. 2019, 17, 4259–4266. [Google Scholar] [CrossRef]
- Zhuang, L.; Qu, H.; Cong, J.; Dai, H.; Liu, X. MiR-181c Affects Estrogen-Dependent Endometrial Carcinoma Cell Growth by Targeting PTEN. Endocr. J. 2019, 66, 523–533. [Google Scholar] [CrossRef]
- Jia, Y.; Liu, M.; Wang, S. CircRNA Hsa_circRNA_0001776 Inhibits Proliferation and Promotes Apoptosis in Endometrial Cancer via Downregulating LRIG2 by Sponging MiR-182. Cancer Cell. Int. 2020, 20, 412. [Google Scholar] [CrossRef]
- Guo, Y.; Liao, Y.; Jia, C.; Ren, J.; Wang, J.; Li, T. MicroRNA-182 Promotes Tumor Cell Growth by Targeting Transcription Elongation Factor A-like 7 in Endometrial Carcinoma. Cell. Physiol. Biochem. 2013, 32, 581–590. [Google Scholar] [CrossRef] [PubMed]
- Ruan, H.; Liang, X.; Zhao, W.; Ma, L.; Zhao, Y. The Effects of MicroRNA-183 Promots Cell Proliferation and Invasion by Targeting MMP-9 in Endometrial Cancer. Biomed. Pharmacother. 2017, 89, 812–818. [Google Scholar] [CrossRef]
- Chen, Z.; Zhu, Y.; Fan, X.; Liu, Y.; Feng, Q. Decreased Expression of MiR-184 Restrains the Growth and Invasion of Endometrial Carcinoma Cells through CDC25A-Dependent Notch Signaling Pathway. Am. J. Transl. Res. 2019, 11, 755–764. [Google Scholar] [PubMed]
- Yang, C.; Ota-Kurogi, N.; Ikeda, K.; Okumura, T.; Horie-Inoue, K.; Takeda, S.; Inoue, S. MicroRNA-191 Regulates Endometrial Cancer Cell Growth via TET1-Mediated Epigenetic Modulation of APC. J. Biochem. 2020, 168, 7–14. [Google Scholar] [CrossRef]
- Ni, J.; Tian, W.; Liang, S.; Wang, H.; Ren, Y. Promoter Methylation-Mediated Silencing of the MiR-192-5p Promotes Endometrial Cancer Progression by Targeting ALX1. Int. J. Med. Sci. 2021, 18, 2510–2520. [Google Scholar] [CrossRef]
- Deng, J.; Wang, W.; Yu, G.; Ma, X. MicroRNA-195 Inhibits Epithelial-mesenchymal Transition by Targeting G Protein-coupled Estrogen Receptor 1 in Endometrial Carcinoma. Mol. Med. Rep. 2019, 20, 4023–4032. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Dai, L.; Yue, Q.; Wang, H.; Wang, X.U.; Li, Y.; Chen, R. MiR-195 Inhibits Migration, Invasion and Epithelial-Mesenchymal Transition (EMT) of Endometrial Carcinoma Cells by Targeting SOX4. J. Biosci. 2019, 44, 146. [Google Scholar] [CrossRef]
- Ma, J.; Zhao, X.; Shi, L. Circ 003390/Eukaryotic Translation Initiation Factor 4A3 Promoted Cell Migration and Proliferation in Endometrial Cancer via Vascular Endothelial Growth Factor Signaling by MiR-195-5p. Bioengineered 2022, 13, 11958–11972. [Google Scholar] [CrossRef]
- Kong, F.; Ma, J.; Yang, H.; Yang, D.; Wang, C.; Ma, X. Long Non-Coding RNA PVT1 Promotes Malignancy in Human Endometrial Carcinoma Cells through Negative Regulation of MiR-195-5p. Biochim. Biophys. Acta (BBA) Mol. Cell Res. 2018, 1865, 1479–1490. [Google Scholar] [CrossRef]
- Xu, H.; Sun, Y.; Ma, Z.; Xu, X.; Qin, L.; Luo, B. LOC134466 Methylation Promotes Oncogenesis of Endometrial Carcinoma through LOC134466/Hsa-MiR-196a-5p/TAC1 Axis. Aging 2018, 10, 3353–3370. [Google Scholar] [CrossRef]
- Shen, Q.; He, T.; Yuan, H. Hsa_circ_0002577 Promotes Endometrial Carcinoma Progression via Regulating MiR-197/CTNND1 Axis and Activating Wnt/β-Catenin Pathway. Cell Cycle 2019, 18, 1229–1240. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Pan, A.; Zhang, Y.; Li, X. Hsa_circ_0039569 Facilitates the Progression of Endometrial Carcinoma by Targeting the MiR-197/High Mobility Group Protein A1 Axis. Bioengineered 2022, 13, 4212–4225. [Google Scholar] [CrossRef]
- Shi, W.; Wang, X.; Ruan, L.; Fu, J.; Liu, F.; Qu, J. MiR-200a Promotes Epithelial-Mesenchymal Transition of Endometrial Cancer Cells by Negatively Regulating FOXA2 Expression. Pharmazie 2017, 72, 694–699. [Google Scholar] [CrossRef]
- Dai, Y.; Xia, W.; Song, T.; Su, X.; Li, J.; Li, S.; Chen, Y.; Wang, W.; Ding, H.; Liu, X.; et al. MicroRNA-200b Is Overexpressed in Endometrial Adenocarcinomas and Enhances MMP2 Activity by Downregulating TIMP2 in Human Endometrial Cancer Cell Line HEC-1A Cells. Nucleic Acid Ther. 2013, 23, 29–34. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Zhang, C.; Chen, R.; Xiong, H.; Qiu, F.; Liu, S.; Zhang, M.; Wang, F.; Wang, Y.; Zhou, X.; et al. Disrupting MALAT1/MiR-200c Sponge Decreases Invasion and Migration in Endometrioid Endometrial Carcinoma. Cancer Lett. 2016, 383, 28–40. [Google Scholar] [CrossRef] [PubMed]
- Chen, R.; Zhang, M.; Liu, W.; Chen, H.; Cai, T.; Xiong, H.; Sheng, X.; Liu, S.; Peng, J.; Wang, F.; et al. Estrogen Affects the Negative Feedback Loop of PTENP1-MiR200c to Inhibit PTEN Expression in the Development of Endometrioid Endometrial Carcinoma. Cell. Death Dis. 2018, 10, 4. [Google Scholar] [CrossRef]
- Li, F.; Liang, A.; Lv, Y.; Liu, G.; Jiang, A.; Liu, P. MicroRNA-200c Inhibits Epithelial-Mesenchymal Transition by Targeting the BMI-1 Gene Through the Phospho-AKT Pathway in Endometrial Carcinoma Cells In Vitro. Med. Sci. Monit. 2017, 23, 5139–5149. [Google Scholar] [CrossRef]
- Park, Y.-A.; Lee, J.-W.; Choi, J.-J.; Jeon, H.-K.; Cho, Y.; Choi, C.; Kim, T.-J.; Lee, N.W.; Kim, B.-G.; Bae, D.-S. The Interactions between MicroRNA-200c and BRD7 in Endometrial Carcinoma. Gynecol. Oncol. 2012, 124, 125–133. [Google Scholar] [CrossRef]
- Xin, W.; Gao, X.; Zhao, P.; Wang, T.; Ding, X.; Wu, Q.; Hua, K. Long Non-Coding RNA RP11-379k17.4 Derived MicroRNA-200c-3p Modulates Human Endometrial Cancer by Targeting Noxa. J. Cancer 2021, 12, 2268–2274. [Google Scholar] [CrossRef]
- Chung, T.K.H.; Lau, T.S.; Cheung, T.H.; Yim, S.F.; Lo, K.W.K.; Siu, N.S.S.; Chan, L.K.Y.; Yu, M.Y.; Kwong, J.; Doran, G.; et al. Dysregulation of MicroRNA-204 Mediates Migration and Invasion of Endometrial Cancer by Regulating FOXC1. Int. J. Cancer. 2012, 130, 1036–1045. [Google Scholar] [CrossRef]
- Jing, S.; Feng, Y.; He, X.L.; Wang, Y. Effects of LncRNA-UCA1 Targeting MiR-204-5p on the Proliferation, Migration, Apoptosis and Immune Escape of Endometrial Carcinoma Cells. Zhonghua Zhong Liu Za Zhi 2023, 45, 56–63. [Google Scholar] [CrossRef] [PubMed]
- Bao, W.; Wang, H.-H.; Tian, F.-J.; He, X.-Y.; Qiu, M.-T.; Wang, J.-Y.; Zhang, H.-J.; Wang, L.-H.; Wan, X.-P. A TrkB–STAT3–MiR-204-5p Regulatory Circuitry Controls Proliferation and Invasion of Endometrial Carcinoma Cells. Mol. Cancer 2013, 12, 155. [Google Scholar] [CrossRef] [PubMed]
- Su, N.; Qiu, H.; Chen, Y.; Yang, T.; Yan, Q.; Wan, X. MiR-205 Promotes Tumor Proliferation and Invasion through Targeting ESRRG in Endometrial Carcinoma. Oncol. Rep. 2013, 29, 2297–2302. [Google Scholar] [CrossRef]
- Zhang, G.; Hou, X.; Li, Y.; Zhao, M. MiR-205 Inhibits Cell Apoptosis by Targeting Phosphatase and Tensin Homolog Deleted on Chromosome Ten in Endometrial Cancer Ishikawa Cells. BMC Cancer 2014, 14, 440. [Google Scholar] [CrossRef]
- Xin, W.; Zhao, S.; Han, X.; Zhao, P.; Yu, H.; Gao, X.; Li, P.; Wu, Q.; Ding, J.; Hua, K. LncRNA LA16c-313D11.11 Modulates the Development of Endometrial Cancer by Binding to and Inhibiting MicroRNA-205-5p Function and Indirectly Increasing PTEN Activity. Int. J. Oncol. 2020, 57, 355–363. [Google Scholar] [CrossRef]
- Xin, W.; Gao, X.; Zhao, S.; Zhao, P.; Yu, H.; Wu, Q.; Hua, K. LncRNA RP11-395G23.3 Suppresses the Endometrial Cancer Progression via Regulating MicroRNA-205-5p/PTEN Axis. Am. J. Transl. Res. 2020, 12, 4422–4433. [Google Scholar] [PubMed]
- Zheng, Y.; Yang, X.; Wang, C.; Zhang, S.; Wang, Z.; Li, M.; Wang, Y.; Wang, X.; Yang, X. HDAC6, Modulated by MiR-206, Promotes Endometrial Cancer Progression through the PTEN/AKT/MTOR Pathway. Sci. Rep. 2020, 10, 3576. [Google Scholar] [CrossRef]
- Dai, Z.; Luo, H.; Chen, J.; Li, L. MiR-210-3p Accelerates Tumor-Relevant Cell Functions of Endometrial Carcinoma by Repressing RUNX1T1. Mutat. Res. Fundam. Mol. Mech. Mutagen. 2022, 825, 111793. [Google Scholar] [CrossRef]
- Sun, J.; Gao, S.; Lu, C. Knockdown of Differentiation Antagonizing Non-Protein Coding RNA Exerts Anti-Tumor Effect by up-Regulating MiR-214 in Endometrial Carcinoma. Mol. Cell. Biochem. 2019, 460, 9–15. [Google Scholar] [CrossRef]
- Wang, C.; Li, Q.; He, Y. MicroRNA-21-5p Promotes Epithelial to Mesenchymal Transition by Targeting SRY-box 17 in Endometrial Cancer. Oncol. Rep. 2020, 43, 1897–1905. [Google Scholar] [CrossRef]
- Xu, D.; Dong, P.; Xiong, Y.; Chen, R.; Konno, Y.; Ihira, K.; Yue, J.; Watari, H. PD-L1 Is a Tumor Suppressor in Aggressive Endometrial Cancer Cells and Its Expression Is Regulated by MiR-216a and LncRNA MEG3. Front. Cell. Dev. Biol. 2020, 8, 598205. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Huang, M.; You, J.; Lin, Y.; Huang, Q.; He, C. Circular RNA Hsa_circ_0023404 Promotes the Proliferation, Migration and Invasion in Endometrial Cancer Cells through Regulating MiR-217/MAPK1 Axis. Eur. J. Med. Res. 2022, 27, 242. [Google Scholar] [CrossRef] [PubMed]
- Li, X.-C.; Hai, J.-J.; Tan, Y.-J.; Yue, Q.-F.; Liu, L.-J. MiR-218 Suppresses Metastasis and Invasion of Endometrial Cancer via Negatively Regulating ADD2. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 1408–1417. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Yu, Z.; Meng, X.; Zhou, S.; Xiao, S.; Li, X.; Liu, S.; Yu, P. Long Noncoding RNA GAS5 Impairs the Proliferation and Invasion of Endometrial Carcinoma Induced by High Glucose via Targeting MiR-222-3p/P27. Am. J. Transl. Res. 2019, 11, 2413–2421. [Google Scholar]
- Huang, K.; Dong, X.; Sui, C.; Hu, D.; Xiong, T.; Liao, S.; Zhang, H. MiR-223 Suppresses Endometrial Carcinoma Cells Proliferation by Targeting IGF-1R. Am. J. Transl. Res. 2014, 6, 841–849. [Google Scholar]
- Shang, C.; Lang, B.; Ao, C.N.; Meng, L. Long Non-Coding RNA Tumor Suppressor Candidate 7 Advances Chemotherapy Sensitivity of Endometrial Carcinoma through Targeted Silencing of MiR-23b. Tumor Biol. 2017, 39, 101042831770788. [Google Scholar] [CrossRef]
- Lu, M.; Ding, N.; Zhuang, S.; Li, Y. LINC01410/MiR-23c/CHD7 Functions as a CeRNA Network to Affect the Prognosis of Patients with Endometrial Cancer and Strengthen the Malignant Properties of Endometrial Cancer Cells. Mol. Cell. Biochem. 2020, 469, 9–19. [Google Scholar] [CrossRef]
- Li, S.; Shan, Y.; Li, X.; Zhang, C.; Wei, S.; Dai, S.; Zhao, R.; Zhao, X.; Zhao, L.; Shan, B. LncRNA SNHG5 Modulates Endometrial Cancer Progression via the MiR-25-3p/BTG2 Axis. J. Oncol. 2019, 2019, 7024675. [Google Scholar] [CrossRef]
- Fan, J.-T.; Zhou, Z.-Y.; Luo, Y.-L.; Luo, Q.; Chen, S.-B.; Zhao, J.-C.; Chen, Q.-R. Exosomal LncRNA NEAT1 from Cancer-Associated Fibroblasts Facilitates Endometrial Cancer Progression via MiR-26a/b-5p-Mediated STAT3/YKL-40 Signaling Pathway. Neoplasia 2021, 23, 692–703. [Google Scholar] [CrossRef]
- Che, X.; Jian, F.; Chen, C.; Liu, C.; Liu, G.; Feng, W. PCOS Serum-Derived Exosomal MiR-27a-5p Stimulates Endometrial Cancer Cells Migration and Invasion. J. Mol. Endocrinol. 2020, 64, 1–12. [Google Scholar] [CrossRef]
- Fan, L.; Wang, C.; Zhan, P.; Liu, Y. LncRNA RBAT1 Reduces Chemosensitivity of Cancer Cells to Carboplatin/Paclitaxel by Sponging MiR-27b in Endometrial Carcinoma. J. Ovarian Res. 2023, 16, 147. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Hu, J.; Yu, T.; You, S.; Zhang, Y.; Hu, L. MiR-27b-3p/MARCH7 Regulates Invasion and Metastasis of Endometrial Cancer Cells through Snail-Mediated Pathway. Acta Biochim. Biophys. Sin 2019, 51, 492–500. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Xing, G.; Liu, S.; Li, B.; He, Y.; Wang, F. LncRNA LOXL1-AS1 Promotes Endometrial Cancer Progression by Sponging MiR-28-5p to Upregulate RAP1B Expression. Biomed. Pharmacother. 2020, 125, 109839. [Google Scholar] [CrossRef]
- Liu, Y.; Yuan, H.; He, T. Downregulated Circular RNA Hsa_circ_0005797 Inhibits Endometrial Cancer by Modulating MicroRNA-298/Catenin Delta 1 Signaling. Bioengineered 2022, 13, 4634–4645. [Google Scholar] [CrossRef]
- Liu, L.; Chen, X.; Zhang, Y.; Hu, Y.; Shen, X.; Zhu, W. Long Non-Coding RNA TUG1 Promotes Endometrial Cancer Development via Inhibiting MiR-299 and MiR-34a-5p. Oncotarget 2017, 8, 31386–31394. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Xiao, J.; Ma, C. Circular RNA FOXO3 Regulates Endometrial Carcinoma Progression through the MicroRNA-29a-3p/HDAC4 Axis. Am. J. Transl. Res. 2022, 14, 8572–8587. [Google Scholar] [PubMed]
- Kong, J.; He, X.; Wang, Y.; Li, J. Effect of MicroRNA-29b on Proliferation, Migration, and Invasion of Endometrial Cancer Cells. J. Int. Med. Res. 2019, 47, 3803–3817. [Google Scholar] [CrossRef]
- Chen, H.-X.; Xu, X.-X.; Tan, B.-Z.; Zhang, Z.; Zhou, X.-D. MicroRNA-29b Inhibits Angiogenesis by Targeting VEGFA through the MAPK/ERK and PI3K/Akt Signaling Pathways in Endometrial Carcinoma. Cell. Physiol. Biochem. 2017, 41, 933–946. [Google Scholar] [CrossRef]
- Pan, X.; Li, D.; Huo, J.; Kong, F.; Yang, H.; Ma, X. LINC01016 Promotes the Malignant Phenotype of Endometrial Cancer Cells by Regulating the MiR-302a-3p/MiR-3130-3p/NFYA/SATB1 Axis. Cell. Death Dis. 2018, 9, 303. [Google Scholar] [CrossRef]
- Ma, J.; Li, D.; Kong, F.-F.; Yang, D.; Yang, H.; Ma, X.-X. MiR-302a-5p/367-3p-HMGA2 Axis Regulates Malignant Processes during Endometrial Cancer Development. J. Exp. Clin. Cancer Res. 2018, 37, 19. [Google Scholar] [CrossRef]
- Hu, Y. MicroRNA-30c Negatively Regulates Endometrial Cancer Cells by Targeting Metastasis-Associated Gene-1. Oncol. Rep. 2011, 27, 807–812. [Google Scholar] [CrossRef] [PubMed]
- Shu, S.; Liu, X.; Xu, M.; Gao, X.; Chen, S.; Zhang, L.; Li, R. MicroRNA-320a Acts as a Tumor Suppressor in Endometrial Carcinoma by Targeting IGF-1R. Int. J. Mol. Med. 2019, 43, 1505–1512. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Li, R.; Li, Y.; Yu, X.; Sun, Q.; Li, A.; Kong, Y. EIF4E-related MiR-320a and MiR-340-5p Inhibit Endometrial Carcinoma Cell Metastatic Capability by Preventing TGF-β1-induced Epithelial-mesenchymal Transition. Oncol. Rep. 2019, 43, 447–460. [Google Scholar] [CrossRef]
- Liu, Y.; Chang, Y.; Cai, Y. Circ_0067835 Sponges MiR-324-5p to Induce HMGA1 Expression in Endometrial Carcinoma Cells. J. Cell. Mol. Med. 2020, 24, 13927–13937. [Google Scholar] [CrossRef]
- Gao, Y.; Qian, H.; Tang, X.; Du, X.; Wang, G.; Zhang, H.; Ye, F.; Liu, T. Superparamagnetic Iron Oxide Nanoparticle-Mediated Expression of miR-326 Inhibits Human Endometrial Carcinoma Stem Cell Growth. Int. J. Nanomed. 2019, 14, 2719–2731. [Google Scholar] [CrossRef]
- Liu, W.; Zhang, B.; Xu, N.; Wang, M.-J.; Liu, Q. MiR-326 Regulates EMT and Metastasis of Endometrial Cancer through Targeting TWIST1. Eur. Rev. Med. Pharmacol. Sci. 2017, 21, 3787–3793. [Google Scholar]
- Wang, R.; Zhang, C.; Guan, W.; Yang, Q. MiRNA-329-3p Suppresses Proliferation and Metastasis of Endometrial Carcinoma through Downregulating E2F1. Neoplasma 2023, 70, 566–579. [Google Scholar] [CrossRef]
- Dou, X.; Chen, X.; Zhou, Q.; Wen, M.; Zhang, S.; Zhang, S. MiR-335 Modulates Numb Alternative Splicing via Targeting RBM10 in Endometrial Cancer. Kaohsiung J. Med. Sci. 2020, 36, 171–177. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Chang, Y.; Cai, Y. Inhibition of Lnc-OC1 Induced Cell Apoptosis and Decreased Cell Viability by Releasing MiR-34a and Inhibiting PD-L1 in Endometrial Carcinoma. Reprod. Sci. 2020, 27, 1848–1856. [Google Scholar] [CrossRef]
- Schirmer, U.; Doberstein, K.; Rupp, A.-K.; Bretz, N.P.; Wuttig, D.; Kiefel, H.; Breunig, C.; Fiegl, H.; Müller-Holzner, E.; Zeillinger, R.; et al. Role of MiR-34a as a Suppressor of L1CAM in Endometrial Carcinoma. Oncotarget 2014, 5, 462–472. [Google Scholar] [CrossRef]
- Wang, Z.; Wang, W.; Huang, K.; Wang, Y.; Li, J.; Yang, X. MicroRNA-34a Inhibits Cells Proliferation and Invasion by Downregulating Notch1 in Endometrial Cancer. Oncotarget 2017, 8, 111258–111270. [Google Scholar] [CrossRef]
- Li, F.; Chen, H.; Huang, Y.; Zhang, Q.; Xue, J.; Liu, Z.; Zheng, F. MiR-34c Plays a Role of Tumor Suppressor in HEC-1-B Cells by Targeting E2F3 Protein. Oncol. Rep. 2015, 33, 3069–3074. [Google Scholar] [CrossRef] [PubMed]
- Jiang, L.; Meng, W.; Zeng, J.; Hu, H.; Lu, L. MiR-34c Oligonucleotide Enhances Chemosensitivity of Ishikawa Cell to Cisplatin by Inducing Apoptosis. Cell. Biol. Int. 2013, 37, 577–583. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Yuan, S.; Liu, X.; Huang, Y.; Qiu, P.; Gao, J.; Deng, G. LncRNA GATA3-AS1 Promoted Invasion and Migration in Human Endometrial Carcinoma by Regulating the MiR-361/ARRB2 Axis. J. Mol. Med. 2022, 100, 1271–1286. [Google Scholar] [CrossRef]
- Dong, P.; Xiong, Y.; Yue, J.; Xu, D.; Ihira, K.; Konno, Y.; Kobayashi, N.; Todo, Y.; Watari, H. Long Noncoding RNA NEAT1 Drives Aggressive Endometrial Cancer Progression via MiR-361-Regulated Networks Involving STAT3 and Tumor Microenvironment-Related Genes. J. Exp. Clin. Cancer Res. 2019, 38, 295. [Google Scholar] [CrossRef]
- Wu, B.; Ren, A.; Tian, Y.; Huang, R. Hsa_circ_0075960 Serves as a Sponge for MiR-361-3p/SH2B1 in Endometrial Carcinoma. Technol. Cancer Res. Treat. 2020, 19, 1533033820983079. [Google Scholar] [CrossRef]
- Wang, L.; Zhao, S.; Mingxin, Y.U. LncRNA NR2F1-AS1 Is Involved in the Progression of Endometrial Cancer by Sponging MiR-363 to Target SOX4. Pharmazie 2019, 74, 295–300. [Google Scholar] [CrossRef]
- Liu, B.-L.; Sun, K.-X.; Zong, Z.-H.; Chen, S.; Zhao, Y. MicroRNA-372 Inhibits Endometrial Carcinoma Development by Targeting the Expression of the Ras Homolog Gene Family Member C (RhoC). Oncotarget 2016, 7, 6649–6664. [Google Scholar] [CrossRef]
- Liang, H.; Liu, Y.; Fu, L.; Li, L.; Gong, N. Berberine Inhibits the Development of Endometrial Cancer through Circ_ZNF608/MiR-377-3p/COX2 Axis. Autoimmunity 2022, 55, 485–495. [Google Scholar] [CrossRef]
- Peng, T.; Zhou, Y.; Zhou, J.; Zhou, Y.; Li, X.; Ouyang, Q. Long Non-coding RNA VPS9D1-AS1Enhances Proliferation, Invasion, and Epithelial-mesenchymal Transition in Endometrial Cancer via miR-377-3p/ SGK1. Kaohsiung J. Med. Sci. 2022, 38, 1048–1059. [Google Scholar] [CrossRef]
- Zhang, C.; Wang, R.; Li, M.; Yang, Q. Long Non-coding RNA BLACAT2/MiR-378a-3p/YY1 Feedback Loop Promotes the Proliferation, Migration and Invasion of Uterine Corpus Endometrial Carcinoma. Oncol. Rep. 2023, 49, 108. [Google Scholar] [CrossRef] [PubMed]
- Tu, C.; Wang, F.; Wan, J. MicroRNA-381 Inhibits Cell Proliferation and Invasion in Endometrial Carcinoma by Targeting the IGF-1R. Mol. Med. Rep. 2017, 17, 4090–4098. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Wang, B.; Wu, L. MicroRNA-409 May Function as a Tumor Suppressor in Endometrial Carcinoma Cells by Targeting Smad2. Mol. Med. Rep. 2018, 19, 622–628. [Google Scholar] [CrossRef] [PubMed]
- Lu, K.; Lin, J.; Jiang, J. Osthole Inhibited Cell Proliferation and Induced Cell Apoptosis through Decreasing CPEB2 Expression via Up-Regulating MiR-424 in Endometrial Carcinoma. J. Recept. Signal Transduct. 2020, 40, 89–96. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Qiu, X.-M.; Li, Q.-H.; Wang, X.-Y.; Li, L.; Xu, M.; Dong, M.; Xiao, Y.-B. MicroRNA-424 May Function as a Tumor Suppressor in Endometrial Carcinoma Cells by Targeting E2F7. Oncol. Rep. 2015, 33, 2354–2360. [Google Scholar] [CrossRef]
- Zhang, H.; Wang, X.; Chen, Z.; Wang, W. MicroRNA-424 Suppresses Estradiol-Induced Cell Proliferation via Targeting GPER in Endometrial Cancer Cells. Cell. Mol. Biol. 2015, 61, 96–101. [Google Scholar] [PubMed]
- Shu, S.; Liu, X.; Xu, M.; Gao, X.; Fan, J.; Liu, H.; Li, R. MicroRNA-424 Regulates Epithelial-mesenchymal Transition of Endometrial Carcinoma by Directly Targeting Insulin-like Growth Factor 1 Receptor. J. Cell. Biochem. 2019, 120, 2171–2179. [Google Scholar] [CrossRef]
- Cai, P.; Wu, M.; Zhang, B.; Wu, S.; Wei, H.; Wei, L. Long Non-coding RNA SNHG12 Regulates Cell Proliferation, Invasion and Migration in Endometrial Cancer by Targeting MiR-4429. Mol. Med. Rep. 2020, 22, 2842–2850. [Google Scholar] [CrossRef]
- Gao, X.; Yu, L.; Zhang, J.; Xue, P. Silencing of Long Non-Coding RNA LINC01106 Suppresses the Proliferation, Migration and Invasion of Endometrial Cancer Cells Through Regulating the MiR-449a/MET Axis. Onco. Targets Ther. 2020, 13, 9643–9655. [Google Scholar] [CrossRef]
- Ye, W.; Xue, J.; Zhang, Q.; Li, F.; Zhang, W.; Chen, H.; Huang, Y.; Zheng, F. MiR-449a Functions as a Tumor Suppressor in Endometrial Cancer by Targeting CDC25A. Oncol. Rep. 2014, 32, 1193–1199. [Google Scholar] [CrossRef]
- Wu, A.-Y.; Hu, Y.; Cang, W.; Li, D.; Wang, W.-J.; Tian, Q.; Gu, L.-Y.; Zhang, N.; Ji, F.; Qiu, L.-H. Suppressive Effect of MicroRNA-449a on the NDRG1/PTEN/AKT Axis Regulates Endometrial Cancer Growth and Metastasis. Exp. Cell. Res. 2019, 382, 111468. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Wu, A.-Y.; Xu, C.; Song, K.-Q.; Wang, W.-J.; Yin, X.; Di, W.; Hong, Z.-B.; Qiu, L.-H. MicroRNA-449a Inhibits Tumor Metastasis through AKT/ERK1/2 Inactivation by Targeting Steroid Receptor Coactivator (SRC) in Endometrial Cancer. J. Cancer 2019, 10, 547–555. [Google Scholar] [CrossRef] [PubMed]
- Zuo, X.; Li, W.; Yan, X.; Ma, T.; Ren, Y.; Hua, M.; Yang, H.; Wu, H.; Zhu, H. Long Non-coding RNA LINC01224 Promotes Cell Proliferation and Inhibits Apoptosis by Regulating AKT3 Expression via Targeting MiR-485-5p in Endometrial Carcinoma. Oncol. Rep. 2021, 46, 186. [Google Scholar] [CrossRef]
- Zheng, X.; Xu, K.; Zhu, L.; Mao, M.; Zhang, F.; Cui, L. MiR-486-5p Act as a Biomarker in Endometrial Carcinoma: Promotes Cell Proliferation, Migration, Invasion by Targeting MARK1. Onco. Targets Ther. 2020, 13, 4843–4853. [Google Scholar] [CrossRef] [PubMed]
- Shao, W.; Li, Y.; Chen, F.; Jia, H.; Jia, J.; Fu, Y. Long Non-Coding RNA DLEU1 Contributes to the Development of Endometrial Cancer by Sponging MiR-490 to Regulate SP1 Expression. Pharmazie 2018, 73, 379–385. [Google Scholar] [CrossRef]
- Zhang, G.; Ma, A.; Jin, Y.; Pan, G.; Wang, C. LncRNA SNHG16 Induced by TFAP2A Modulates Glycolysis and Proliferation of Endometrial Carcinoma through MiR-490-3p/HK2 Axis. Am. J. Transl. Res. 2019, 11, 7137–7145. [Google Scholar]
- Zhu, L.; Wang, X.; Wang, T.; Zhu, W.; Zhou, X. MiR-494-3p Promotes the Progression of Endometrial Cancer by Regulating the PTEN/PI3K/AKT Pathway. Mol. Med. Rep. 2018, 19, 581–588. [Google Scholar] [CrossRef]
- Tan, A.; Luo, R.; Ruan, P. MiR-495 Promotes Apoptosis and Inhibits Proliferation in Endometrial Cells via Targeting PIK3R1. Pathol. Res. Pract. 2019, 215, 594–599. [Google Scholar] [CrossRef]
- He, Y.; Xu, S.; Qi, Y.; Tian, J.; Xu, F. Long Noncoding RNA SNHG25 Promotes the Malignancy of Endometrial Cancer by Sponging MicroRNA-497-5p and Increasing FASN Expression. J. Ovarian Res. 2021, 14, 163. [Google Scholar] [CrossRef]
- Xing, Y.; Sun, X.; Li, F.; Jiang, X.; Jiang, A.; Li, X.; Lv, R.; Shao, L. Long Non-Coding RNA (LncRNA) HOXB-AS3 Promotes Cell Proliferation and Inhibits Apoptosis by Regulating ADAM9 Expression through Targeting MiR-498-5p in Endometrial Carcinoma. J. Int. Med. Res. 2021, 49, 03000605211013548. [Google Scholar] [CrossRef]
- Chen, S.; Sun, K.-X.; Liu, B.-L.; Zong, Z.-H.; Zhao, Y. MicroRNA-505 Functions as a Tumor Suppressor in Endometrial Cancer by Targeting TGF-α. Mol. Cancer 2016, 15, 11. [Google Scholar] [CrossRef]
- Yuan, S.; Zheng, P.; Sun, X.; Zeng, J.; Cao, W.; Gao, W.; Wang, Y.; Wang, L. Hsa_Circ_0001860 Promotes Smad7 to Enhance MPA Resistance in Endometrial Cancer via MiR-520h. Front. Cell. Dev. Biol. 2021, 9, 738189. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Han, Y.; Zhang, X.; Xiao, N.; Jiang, T.; Zhu, S.; Wang, E.; Chen, C. MiR-522 Facilitates the Prosperities of Endometrial Carcinoma Cells by Directly Binding to Monoamine Oxidase, B. Kaohsiung J. Med. Sci. 2019, 35, 598–606. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Wang, M.; Lv, X.; Zhou, J.; Gao, L. CircCCL22 Regulates CDC25A via Sponging MiR-543 and Promotes Proliferation and Metastasis in Endometrial Cancer. Mol. Biotechnol. 2024, 66, 2522–2531. [Google Scholar] [CrossRef] [PubMed]
- Bing, L.; Hong, C.; Li-Xin, S.; Wei, G. MicroRNA-543 Suppresses Endometrial Cancer Oncogenicity via Targeting FAK and TWIST1 Expression. Arch. Gynecol. Obstet. 2014, 290, 533–541. [Google Scholar] [CrossRef]
- Zhong, Y.; Wang, Y.; Dang, H.; Wu, X. LncRNA AFAP1-AS1 Contributes to the Progression of Endometrial Carcinoma by Regulating MiR-545-3p/VEGFA Pathway. Mol. Cell. Probes 2020, 53, 101606. [Google Scholar] [CrossRef]
- Wang, N.; Guo, Y.; Song, L.; Tong, T.; Fan, X. Circular RNA Intraflagellar Transport 80 Facilitates Endometrial Cancer Progression through Modulating MiR-545-3p/FAM98A Signaling. J. Gynecol. Oncol. 2022, 33, e2. [Google Scholar] [CrossRef]
- Wang, Y.; Dong, L.; Liu, Y. Targeting Thyroid Receptor Interacting Protein 6 by MicroRNA-589-5p Inhibits Cell Proliferation, Migration, and Invasion in Endometrial Carcinoma. Cancer Biother. Radiopharm. 2019, 34, 529–536. [Google Scholar] [CrossRef]
- Shi, Y.; Zha, J.; Zuo, M.; Yan, Q.; Song, H. Long Noncoding RNA CHL1-AS1 Promotes Cell Proliferation and Migration by Sponging MiR-6076 to Regulate CHL1 Expression in Endometrial Cancer. J. Cell. Biochem. 2020, 121, 2655–2663. [Google Scholar] [CrossRef]
- Zhang, L.; Wang, D.-L.; Yu, P. LncRNA H19 Regulates the Expression of Its Target Gene HOXA10 in Endometrial Carcinoma through Competing with MiR-612. Eur. Rev. Med. Pharmacol. Sci. 2018, 22, 4820–4827. [Google Scholar] [CrossRef]
- Cong, R.; Kong, F.; Ma, J.; Li, Q.; Yang, H.; Ma, X. The PVT1/MiR-612/CENP-H/CDK1 Axis Promotes Malignant Progression of Advanced Endometrial Cancer. Am. J. Cancer Res. 2021, 11, 1480–1502. [Google Scholar] [PubMed]
- Wu, X.; Cai, D.; Zhang, F.; Li, M.; Wan, Q. Long Noncoding RNA TUSC7 Inhibits Cell Proliferation, Migration and Invasion by Regulating SOCS4 (SOCS5) Expression through Targeting MiR-616 in Endometrial Carcinoma. Life Sci. 2019, 231, 116549. [Google Scholar] [CrossRef] [PubMed]
- Yi, H.; Han, Y.; Li, S. Oncogenic Circular RNA Circ_0007534 Contributes to Paclitaxel Resistance in Endometrial Cancer by Sponging MiR-625 and Promoting ZEB2 Expression. Front. Oncol. 2022, 12, 985470. [Google Scholar] [CrossRef]
- Wang, Y.; Yin, L.; Sun, X. CircRNA Hsa_circ_0002577 Accelerates Endometrial Cancer Progression through Activating IGF1R/PI3K/Akt Pathway. J. Exp. Clin. Cancer Res. 2020, 39, 169. [Google Scholar] [CrossRef]
- Li, X.; Liu, Y. Hsa_circ_0000437 Inhibits the Development of Endometrial Carcinoma through MiR-626/CDKN1B Axis. Protein Pept. Lett. 2022, 29, 611–620. [Google Scholar] [CrossRef]
- Li, Y.; Huo, J.; He, J.; Ma, X. LncRNA MONC Suppresses the Malignant Phenotype of Endometrial Cancer Stem Cells and Endometrial Carcinoma Cells by Regulating the MiR-636/GLCE Axis. Cancer Cell. Int. 2021, 21, 331. [Google Scholar] [CrossRef]
- Ni, J.; Liang, S.; Shan, B.; Tian, W.; Wang, H.; Ren, Y. Methylation-associated Silencing of MiR-638 Promotes Endometrial Carcinoma Progression by Targeting MEF2C. Int. J. Mol. Med. 2020, 45, 1753–1770. [Google Scholar] [CrossRef]
- Wang, B.; Lu, Y.; Feng, E. Hsa_circ_0001610 Knockdown Modulates MiR-646-STAT3 Axis to Suppress Endometrial Carcinoma Progression. J. Gene Med. 2021, 23, e3337. [Google Scholar] [CrossRef]
- Liu, Y.; Chen, S.; Zong, Z.H.; Guan, X.; Zhao, Y. CircRNA WHSC1 Targets the MiR-646/NPM1 Pathway to Promote the Development of Endometrial Cancer. J. Cell. Mol. Med. 2020, 24, 6898–6907. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.X.; Wang, C.; Mao, L.W.; Wang, Y.L.; Xia, L.Q.; Zhao, W.; Shen, J.; Chen, J. Long Noncoding RNA HOTAIR Mediates the Estrogen-Induced Metastasis of Endometrial Cancer Cells via the MiR-646/NPM1 Axis. Am. J. Physiol. Cell. Physiol. 2018, 314, C690–C701. [Google Scholar] [CrossRef]
- Sun, X.; Dongol, S.; Qiu, C.; Xu, Y.; Sun, C.; Zhang, Z.; Yang, X.; Zhang, Q.; Kong, B. MiR-652 Promotes Tumor Proliferation and Metastasis by Targeting RORA in Endometrial Cancer. Mol. Cancer Res. 2018, 16, 1927–1939. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.-F.; Wen, L.-N. LncRNA SNHG14 Promotes Proliferation of Endometrial Cancer through Regulating MicroRNA-655-3p. Eur. Rev. Med. Pharmacol. Sci. 2020, 24, 10410–10418. [Google Scholar] [CrossRef] [PubMed]
- Lai, T.; Qiu, H.; Si, L.; Zhen, Y.; Chu, D.; Guo, R. Long Noncoding RNA BMPR1B-AS1 Facilitates Endometrial Cancer Cell Proliferation and Metastasis by Sponging MiR-7-2-3p to Modulate the DCLK1/Akt/NF-ΚB Pathway. Cell. Cycle 2022, 21, 1599–1618. [Google Scholar] [CrossRef]
- Zhang, J.; Zhang, Y.; Zhang, J. N6-Methyladenosine Modified Circ-NAB1 Modulates Cell Cycle and Epithelialmesenchymal Transition via CDKN3 in Endometrial Cancer. Cell. Mol. Biol. 2024, 70, 161–169. [Google Scholar] [CrossRef]
- Ke, J.; Shen, Z.; Hu, W.; Li, M.; Shi, Y.; Xie, Z.; Wu, D. LncRNA DCST1-AS1 Was Upregulated in Endometrial Carcinoma and May Sponge MiR-92a-3p to Upregulate Notch1. Cancer Manag. Res. 2020, 12, 1221–1227. [Google Scholar] [CrossRef]
- Chen, S.; Chen, X.; Sun, K.-X.; Xiu, Y.-L.; Liu, B.-L.; Feng, M.-X.; Sang, X.-B.; Zhao, Y. MicroRNA-93 Promotes Epithelial–Mesenchymal Transition of Endometrial Carcinoma Cells. PLoS ONE 2016, 11, e0165776. [Google Scholar] [CrossRef]
- Zhang, K.; Cai, Y.; Zhou, Q.; Sun, H.; Wei, J. Long Non-Coding RNA SNHG14 Impedes Viability, Migration and Invasion of Endometrial Carcinoma Cells Through Modulating MiR-93-5p/ZBTB7A Axis. Cancer Manag. Res. 2020, 12, 9515–9525. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Z.; Xu, Y.-P.; Wang, L.-J.; Kong, Y. MiR-940 Potentially Promotes Proliferation and Metastasis of Endometrial Carcinoma through Regulation of MRVI1. Biosci. Rep. 2019, 39, BSR20190077. [Google Scholar] [CrossRef]
- He, Z.; Xu, H.; Meng, Y.; Kuang, Y. MiR-944 Acts as a Prognostic Marker and Promotes the Tumor Progression in Endometrial Cancer. Biomed. Pharmacother. 2017, 88, 902–910. [Google Scholar] [CrossRef]
- Li, Y.; Zhang, Z.; Zhang, X.; Lin, Y.; Luo, T.; Xiao, Z.; Zhou, Q. A Dual PI3K/AKT/MTOR Signaling Inhibitor MiR-99a Suppresses Endometrial Carcinoma. Am. J. Transl. Res. 2016, 8, 719–731. [Google Scholar]
- O’Brien, J.; Hayder, H.; Zayed, Y.; Peng, C. Overview of MicroRNA Biogenesis, Mechanisms of Actions, and Circulation. Front. Endocrinol. 2018, 9, 402. [Google Scholar] [CrossRef]
- Piergentili, R.; Marinelli, E.; Cucinella, G.; Lopez, A.; Napoletano, G.; Gullo, G.; Zaami, S. MiR-125 in Breast Cancer Etiopathogenesis: An Emerging Role as a Biomarker in Differential Diagnosis, Regenerative Medicine, and the Challenges of Personalized Medicine. Noncoding RNA 2024, 10, 16. [Google Scholar] [CrossRef] [PubMed]
- Katsushima, K.; Natsume, A.; Ohka, F.; Shinjo, K.; Hatanaka, A.; Ichimura, N.; Sato, S.; Takahashi, S.; Kimura, H.; Totoki, Y.; et al. Targeting the Notch-Regulated Non-Coding RNA TUG1 for Glioma Treatment. Nat. Commun. 2016, 7, 13616. [Google Scholar] [CrossRef]
- Wu, R.; Liu, W.; Yang, Q.; Zhang, J.; Hou, P.; Xiong, J.; Wu, L.; Li, E. LncTUG1 Promotes Hepatocellular Carcinoma Immune Evasion via Upregulating PD-L1 Expression. Sci. Rep. 2023, 13, 16998. [Google Scholar] [CrossRef] [PubMed]
- Beadle, G.W.; Tatum, E.L. Genetic Control of Biochemical Reactions in Neurospora. Proc. Natl. Acad. Sci. USA 1941, 27, 499–506. [Google Scholar] [CrossRef] [PubMed]
- Williams, R.J. Biochemical Institute Studies IV—Individual Metabolic Patterns and Human Disease: An Exploratory Study Utilizing Predominantly Paper Chromatographic Methods; U. Texas Publication No. 5109; University of Texas: Austin, TX, USA, 1951; 205p. [Google Scholar]
- Wishart, D.S.; Tzur, D.; Knox, C.; Eisner, R.; Guo, A.C.; Young, N.; Cheng, D.; Jewell, K.; Arndt, D.; Sawhney, S.; et al. HMDB: The Human Metabolome Database. Nucleic Acids Res. 2007, 35, D521–D526. [Google Scholar] [CrossRef]
- Pandey, S. Metabolomics for the identification of biomarkers in endometriosis. Arch. Gynecol. Obstet. 2024, 310, 2823–2827. [Google Scholar] [CrossRef] [PubMed]
- Raffone, A.; Troisi, J.; Boccia, D.; Travaglino, A.; Capuano, G.; Insabato, L.; Mollo, A.; Guida, M.; Zullo, F. Metabolomics in endometrial cancer diagnosis: A systematic review. Acta Obstet. Gynecol. Scand. 2020, 99, 1135–1146. [Google Scholar] [CrossRef]
- Albertí-Valls, M.; Megino-Luque, C.; Macià, A.; Gatius, S.; Matias-Guiu, X.; Eritja, N. Metabolomic-Based Approaches for Endometrial Cancer Diagnosis and Prognosis: A Review. Cancers 2023, 16, 185. [Google Scholar] [CrossRef]
- Li, Z.; Munim, M.B.; Sharygin, D.A.; Bevis, B.J.; Vander Heiden, M.G. Understanding the Warburg Effect in Cancer. Cold Spring Harb. Perspect. Med. 2024, a041532. [Google Scholar] [CrossRef]
- Hishinuma, E.; Shimada, M.; Matsukawa, N.; Shima, Y.; Li, B.; Motoike, I.N.; Shibuya, Y.; Hagihara, T.; Shigeta, S.; Tokunaga, H.; et al. Identification of predictive biomarkers for endometrial cancer diagnosis and treatment response monitoring using plasma metabolome profiling. Cancer Metab. 2023, 11, 16. [Google Scholar] [CrossRef]
- Chang, L.; Xia, J. MicroRNA Regulatory Network Analysis Using miRNet 2.0. Methods Mol. Biol. 2023, 2594, 185–204. [Google Scholar] [CrossRef]
- Androutsopoulos, G.; Styliara, I.; Zarogianni, E.; Lazurko, N.; Valasoulis, G.; Michail, G.; Adonakis, G. The ErbB Signaling Network and Its Potential Role in Endometrial Cancer. Epigenomes 2023, 7, 24. [Google Scholar] [CrossRef] [PubMed]
- Fatima, I.; Barman, S.; Rai, R.; Thiel, K.W.W.; Chandra, V. Targeting Wnt Signaling in Endometrial Cancer. Cancers 2021, 13, 2351. [Google Scholar] [CrossRef]
- Gao, S.; Wang, J.; Wang, T.; Wang, J. Is Insulin Resistance a High-Risk Factor for Postmenopausal Endometrial Cancer: Insights from the Triglyceride Glucose (TyG) Index and the Metabolic Score for Insulin Resistance (METS-IR). Int. J. Womens. Health 2024, 16, 2355–2363. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Li, Q.; Niu, X.; Wang, G.; Zheng, S.; Fu, G.; Wang, Z. MiR-143 Inhibits Bladder Cancer Cell Proliferation and Enhances Their Sensitivity to Gemcitabine by Repressing IGF-1R Signaling. Oncol. Lett. 2017, 13, 435–440. [Google Scholar] [CrossRef]
- Li, D.; Zhong, S.; Zhu, Z.; Jiang, X.; Zhang, J.; Gu, J.; Chen, F. LncRNA MAFG-AS1 Promotes the Progression of Bladder Cancer by Targeting the MiR-143-3p/COX-2 Axis. Pathobiology 2020, 87, 345–355. [Google Scholar] [CrossRef]
- Xiang, W.; Lyu, L.; Huang, T.; Zheng, F.; Yuan, J.; Zhang, C.; Jiang, G. The Long Non-coding RNA SNHG1 Promotes Bladder Cancer Progression by Interacting with MiR-143-3p and EZH2. J. Cell. Mol. Med. 2020, 24, 11858–11873. [Google Scholar] [CrossRef]
- Luo, J.; Chen, J.; Li, H.; Yang, Y.; Yun, H.; Yang, S.; Mao, X. LncRNA UCA1 Promotes the Invasion and EMT of Bladder Cancer Cells by Regulating the MiR-143/HMGB1 Pathway. Oncol. Lett. 2017, 14, 5556–5562. [Google Scholar] [CrossRef]
- Bell, D.W.; Ellenson, L.H. Molecular Genetics of Endometrial Carcinoma. Annu. Rev. Pathol. Mech. Dis. 2019, 14, 339–367. [Google Scholar] [CrossRef]
- Żyła, M.M.; Kostrzewa, M.; Litwińska, E.; Szpakowski, A.; Radosław Wilczyński, J.; Stetkiewicz, T. The Role of Angiogenic Factors in Endometrial Cancer. Menopausal Rev. 2014, 2, 122–126. [Google Scholar] [CrossRef]
- Hong, D.S.; Kang, Y.-K.; Borad, M.; Sachdev, J.; Ejadi, S.; Lim, H.Y.; Brenner, A.J.; Park, K.; Lee, J.-L.; Kim, T.-Y.; et al. Phase 1 Study of MRX34, a Liposomal MiR-34a Mimic, in Patients with Advanced Solid Tumours. Br. J. Cancer 2020, 122, 1630–1637. [Google Scholar] [CrossRef]
- Beg, M.S.; Brenner, A.J.; Sachdev, J.; Borad, M.; Kang, Y.-K.; Stoudemire, J.; Smith, S.; Bader, A.G.; Kim, S.; Hong, D.S. Phase I Study of MRX34, a Liposomal MiR-34a Mimic, Administered Twice Weekly in Patients with Advanced Solid Tumors. Invest. New Drugs 2017, 35, 180–188. [Google Scholar] [CrossRef] [PubMed]
- Peltier, H.J.; Kelnar, K.; Bader, A.G. Effects of MRX34, a Liposomal MiR-34 Mimic, on Target Gene Expression in Human White Blood Cells (HWBCs): QRT-PCR Results from a First-in-Human Trial of MicroRNA Cancer Therapy. J. Clin. Oncol. 2016, 34, e14090. [Google Scholar] [CrossRef]
- Mechahougui, H.; Gutmans, J.; Colarusso, G.; Gouasmi, R.; Friedlaender, A. Advances in Personalized Oncology. Cancers 2024, 16, 2862. [Google Scholar] [CrossRef] [PubMed]
- Shao, J.; Ma, J.; Zhang, Q.; Li, W.; Wang, C. Predicting Gene Mutation Status via Artificial Intelligence Technologies Based on Multimodal Integration (MMI) to Advance Precision Oncology. Semin. Cancer Biol. 2023, 91, 1–15. [Google Scholar] [CrossRef]
- The ESMO Personalised Medicine Task Force. What Is Personalised Medicine? Let Us Help You Understand. Available online: https://www.esmo.org/content/download/20122/337223/1/ESMO-Patient-Guide-Personalised-Cancer-Medicine.pdf (accessed on 10 February 2025).
- Salari, P.; Larijani, B. Ethical Issues Surrounding Personalized Medicine: A Literature Review. Acta Med. Iran. 2017, 55, 209–217. Fiore, R.N.; Goodman, K.W. Precision Medicine Ethics: Selected Issues and Developments in next-Generation Sequencing, Clinical Oncology, and Ethics. Curr. Opin. Oncol. 2016, 28, 83–87. [Google Scholar] [CrossRef]
- Krzyszczyk, P.; Acevedo, A.; Davidoff, E.J.; Timmins, L.M.; Marrero-Berrios, I.; Patel, M.; White, C.; Lowe, C.; Sherba, J.J.; Hartmanshenn, C.; et al. The Growing Role of Precision and Personalized Medicine for Cancer Treatment. Technology 2018, 6, 79–100. [Google Scholar] [CrossRef]
- Gazola, A.A.; Lautert-Dutra, W.; Archangelo, L.F.; Reis, R.B.D.; Squire, J.A. Precision Oncology Platforms: Practical Strategies for Genomic Database Utilization in Cancer Treatment. Mol. Cytogenet. 2024, 17, 28. [Google Scholar] [CrossRef]
- Zeng, J.; Shufean, M.A. Molecular-Based Precision Oncology Clinical Decision Making Augmented by Artificial Intelligence. Emerg. Top Life Sci. 2021, 5, 757–764. [Google Scholar] [CrossRef]
- European Commission. Europe’s Beating Cancer Plan Communication from the Commission to the European Parliament and the Council. Issued in February 2022. Available online: https://health.ec.europa.eu/system/files/2022-02/eu_cancer-plan_en_0.pdf (accessed on 20 August 2023).
- European Commission. Directorate General for Research and Innovation. Conquering Cancer: Mission Possible; Publications Office: Luxembourg, 2020. Available online: https://op.europa.eu/en/publication-detail/-/publication/b389aad3-fd56-11ea-b44f-01aa75ed71a1/https://op.europa.eu/en/publication-detail/-/publication/b389aad3-fd56-11ea-b44f-01aa75ed71a1 (accessed on 29 August 2023).
- Hickman, J.A.; Tannock, I.F.; Meheus, L.; Hutchinson, L. The European Union and personalised cancer medicine. Eur. J. Cancer 2021, 150, 95–98. [Google Scholar] [CrossRef] [PubMed]
- Gambardella, V.; Tarazona, N.; Cejalvo, J.M.; Lombardi, P.; Huerta, M.; Roselló, S.; Fleitas, T.; Roda, D.; Cervantes, A. Personalized Medicine: Recent Progress in Cancer Therapy. Cancers 2020, 12, 1009. [Google Scholar] [CrossRef] [PubMed]
- Hao, W.; Rajendran, B.K.; Cui, T.; Sun, J.; Zhao, Y.; Palaniyandi, T.; Selvam, M. Advances in Predicting Breast Cancer Driver Mutations: Tools for Precision Oncology (Review). Int. J. Mol. Med. 2025, 55, 6. [Google Scholar] [CrossRef] [PubMed]
- Piergentili, R.; Basile, G.; Nocella, C.; Carnevale, R.; Marinelli, E.; Patrone, R.; Zaami, S. Using ncRNAs as Tools in Cancer Diagnosis and Treatment—The Way towards Personalized Medicine to Improve Patients’ Health. IJMS 2022, 23, 9353. [Google Scholar] [CrossRef]
- PCM4EU—Personalised cancer medicine for all EU citizens. Project set to run from 1st January 2023 to 30th June 2025. Available online: https://health.ec.europa.eu/document/download/67f62913-43d0-4092-bfed-2347caadcfe1_en?filename=fs_cancer_pcm4eu.pdf (accessed on 10 February 2025).
- European Cancer Imaging Initiative. Launched on 23 January 2023. Available online: https://digital-strategy.ec.europa.eu/en/policies/cancer-imaging (accessed on 10 February 2025).
- European Commission. Smart Specialisation Platform. European Partnership on Personalised Medicine. TOPIC ID: HORIZON-HLTH-2023-CARE-08-01. Last Updated on 9 August 2023. Available online: https://ec.europa.eu/info/funding-tenders/opportunities/portal/screen/opportunities/topic-details/horizon-hlth-2023-care-08-01 (accessed on 30 August 2023).
- Marchant, G.E.; Lindor, R.A. Personalized Medicine and Genetic Malpractice. Genet. Med. 2013, 15, 921–922. [Google Scholar] [CrossRef]
- Brothers, K.B.; Rothstein, M.A. Ethical, Legal and Social Implications of Incorporating Personalized Medicine into Healthcare. Per. Med. 2015, 12, 43–51. [Google Scholar] [CrossRef]
- Ledford, H. US Personalized-Medicine Industry Takes Hit from Supreme Court. Nature 2016, 536, 382. [Google Scholar] [CrossRef]
- Donohue, K.E.; Gooch, C.; Katz, A.; Wakelee, J.; Slavotinek, A.; Korf, B.R. Pitfalls and Challenges in Genetic Test Interpretation: An Exploration of Genetic Professionals Experience with Interpretation of Results. Clin. Genet. 2021, 99, 638–649. [Google Scholar] [CrossRef]
- Berger, S.M.; Appelbaum, P.S.; Siegel, K.; Wynn, J.; Saami, A.M.; Brokamp, E.; O’Connor, B.C.; Hamid, R.; Martin, D.M.; Chung, W.K. Challenges of Variant Reinterpretation: Opinions of Stakeholders and Need for Guidelines. Genet. Med. 2022, 24, 1878–1887. [Google Scholar] [CrossRef]
- Galante, N.; Bedeschi, M.F.; Beltrami, B.; Bailo, P.; Silva Palomino, L.A.; Piccinini, A. Reviewing Hereditary Connective Tissue Disorders: Proposals of Harmonic Medicolegal Assessments. Int. J. Legal Med. 2024, 138, 2507–2522. [Google Scholar] [CrossRef]
- Milunsky, A. Obstetrics, Genetics, and Litigation. Acta Obstet. Gynecol. Scand. 2021, 100, 1097–1105. [Google Scholar] [CrossRef] [PubMed]
- Statello, L.; Guo, C.-J.; Chen, L.-L.; Huarte, M. Gene Regulation by Long Non-Coding RNAs and Its Biological Functions. Nat. Rev. Mol. Cell. Biol. 2021, 22, 96–118. [Google Scholar] [CrossRef]
- Tsai, M.-C.; Manor, O.; Wan, Y.; Mosammaparast, N.; Wang, J.K.; Lan, F.; Shi, Y.; Segal, E.; Chang, H.Y. Long Noncoding RNA as Modular Scaffold of Histone Modification Complexes. Science 2010, 329, 689–693. [Google Scholar] [CrossRef]
- Gulìa, C.; Signore, F.; Gaffi, M.; Gigli, S.; Votino, R.; Nucciotti, R.; Bertacca, L.; Zaami, S.; Baffa, A.; Santini, E.; et al. Y RNA: An Overview of Their Role as Potential Biomarkers and Molecular Targets in Human Cancers. Cancers 2020, 12, 1238. [Google Scholar] [CrossRef] [PubMed]
- Cao, J. The Functional Role of Long Non-Coding RNAs and Epigenetics. Biol. Proced. Online 2014, 16, 42. [Google Scholar] [CrossRef]
- Qian, Y.; Shi, L.; Luo, Z. Long Non-Coding RNAs in Cancer: Implications for Diagnosis, Prognosis, and Therapy. Front. Med. 2020, 7, 612393. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| miRNA | lncRNA | mRNA Target | Reference | Figure |
|---|---|---|---|---|
| miR-101 | undefined | COX2 | [30] | 1 |
| miR-101 | undefined | EZH2 | [31] | 1 |
| miR-101 | undefined | FOS | [31] | 1 |
| miR-101 | undefined | MCL-1 | [31] | 1 |
| miR-101-3p | undefined | EZH2 | [32] | 1 |
| miR-103 | GAS5 | PTEN | [33] | 1 |
| miR-103 | undefined | ZO-1 | [34] | 1 |
| miR-105 | circ_0109046 | SOX9 | [35] | 1 |
| miR-106a | undefined | BCL2L11 | [36] | no |
| miR-106b | undefined | p21 | [37] | no |
| miR-107-5p | undefined | ERα | [38] | no |
| miR-10b | undefined | HOXB3 | [39] | no |
| miR-1227 | circ_TNFRSF21 | MAPK13 | [40] | no |
| miR-124-3p | H19 | ITGB3 | [41] | 1 |
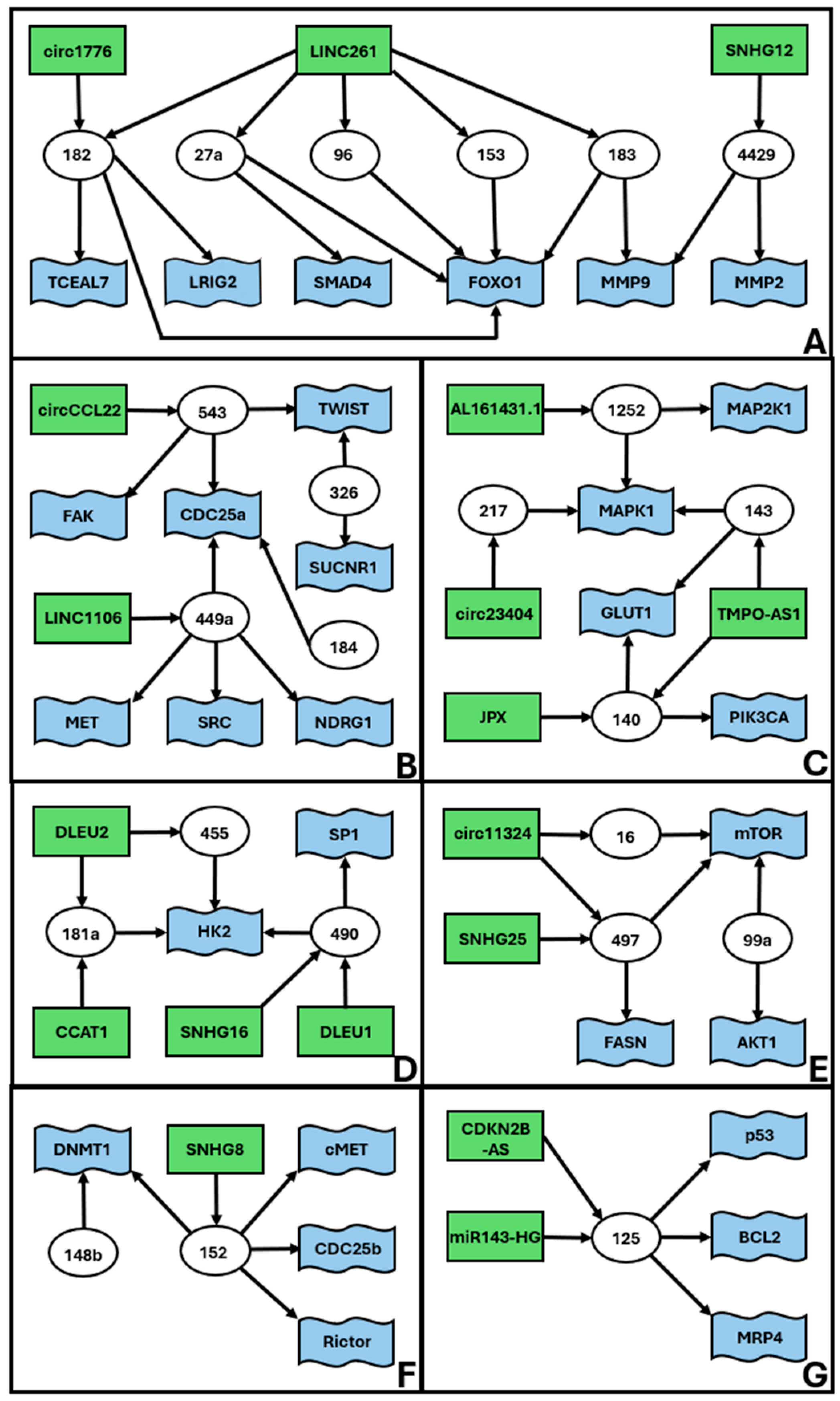
| miR-1252-5p | AL161431.1 | MAP2K1 | [42] | 2C |
| miR-1252-5p | AL161431.1 | MAPK1 | [42] | 2C |
| miR-125a | miR143HG | p53 | [43] | 2G |
| miR-125a-5p | CDKN2B-AS | BCL2 | [44] | 2G |
| miR-125a-5p | CDKN2B-AS | MRP4 | [44] | 2G |
| miR-126 | ATB | undefined | [45] | 1 |
| miR-126-5p | circ_0002577 | MACC1 | [46] | 1 |
| miR-1271 | undefined | CDK1 | [47] | 1 |
| miR-1271 | undefined | LDHA | [48] | 1 |
| miR-1271-5p | circ_0000043 | CTNND1 | [49] | 1 |
| miR-1271-5p | circ_0039569 | PHF6 | [50] | 1 |
| miR-129-2-3p | XIST | CCP110 | [51] | no |
| miR-129-5p | PCGEM1 | STAT3 | [52] | 1 |
| miR-130b | undefined | ZEB1 | [53] | no |
| miR-132-3p | undefined | MTFR2 | [54] | no |
| miR-134 | undefined | POGLUT1 | [55] | no |
| miR-136 | circ_0000043 | NOTCH3 | [56] | 1 |
| miR-136 | circ_0109046 | HMGA2 | [57] | 1 |
| miR-136-5p | DSCAM-AS1 | undefined | [58] | 1 |
| miR-137 | undefined | EZH2 | [59] | 1 |
| miR-137 | undefined | LSD1 | [59] | 1 |
| miR-139-5p | circ_0001610 | CCNB1 | [60] | 1 |
| miR-139-5p | undefined | HOXA10 | [61] | 1 |
| miR-140 | TMPO-AS1 | GLUT1 | [62] | 2C |
| miR-140-3p | JPX | PIK3CA | [63] | 2C |
| miR-141-3p | MIR22HG | DAPK1 | [64] | no |
| miR-143 | TMPO-AS1 | GLUT1 | [62] | 2C |
| miR-143 | undefined | MAPK1 | [65] | 2C |
| miR-144-3p | NEAT1 | EZH2 | [66] | 1 |
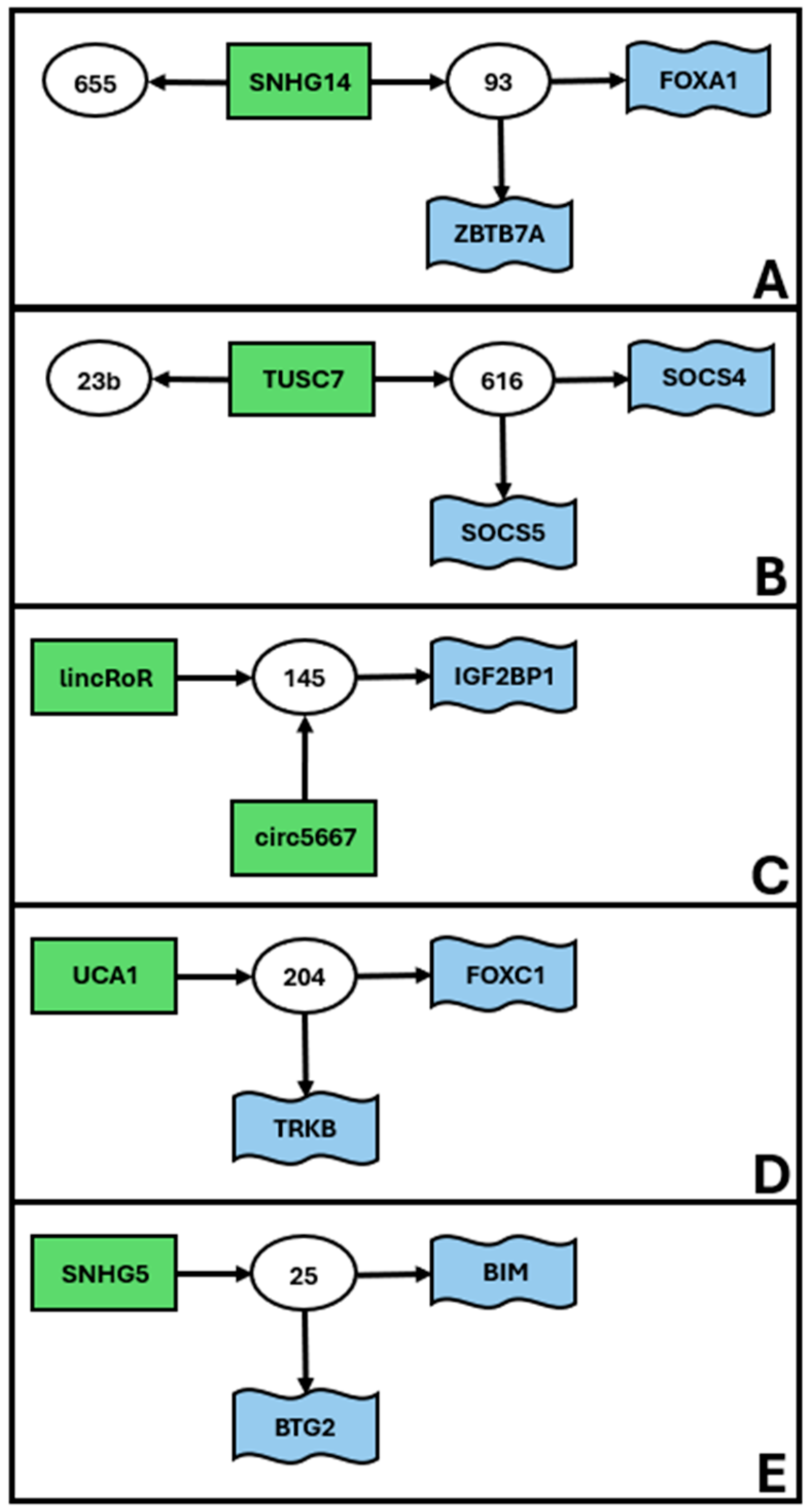
| miR-145 | linc-RoR | undefined | [67] | 3C |
| miR-145-5p | circ_0005667 | IGF2BP1 | [68] | 3C |
| miR-148b | undefined | DNMT1 | [69] | 2F |
| miR-149-5p | circ_0061140 | STAT3 | [70] | 1 |
| miR-152 | SNHG8 | c-MET | [71] | 2F |
| miR-152 | undefined | CDC25B | [72] | 2F |
| miR-152 | undefined | DNMT1 | [73] | 2F |
| miR-152 | undefined | Rictor | [73] | 2F |
| miR-153 | LINC00261 | FOXO1 | [74] | 2A |
| miR-155 | undefined | AGTR1 | [75] | no |
| miR-15a-5p | undefined | WNT3A | [76] | no |
| miR-16-5p | circ_0011324 | mTOR | [77] | 2E |
| miR-181a | DLEU2 | HK2 | [78] | 2D |
| miR-181a-5p | CCAT1 | undefined | [79] | 2D |
| miR-181c | undefined | PTEN | [80] | 1 |
| miR-182 | circ_0001776 | LRIG2 | [81] | 2A |
| miR-182 | LINC00261 | FOXO1 | [74] | 2A |
| miR-182 | undefined | TCEAL7 | [82] | 2A |
| miR-183 | LINC00261 | FOXO1 | [74] | 2A |
| miR-183 | undefined | MMP9 | [83] | 2A |
| miR-184 | undefined | CDC25A | [84] | 2B |
| miR-191 | undefined | TET1 | [85] | no |
| miR-192-5p | undefined | ALX1 | [86] | no |
| miR-195 | undefined | GPER | [87] | 1 |
| miR-195 | undefined | SOX4 | [88] | 1 |
| miR-195-5p | circ_0003390 | VEGFA | [89] | 1 |
| miR-195-5p | PVT1 | FGF2 | [90] | 1 |
| miR-195-5p | PVT1 | FGFR1 | [90] | 1 |
| miR-196a-5p | LOC134466 | TAC1 | [91] | no |
| miR-197 | circ_0002577 | CTNND1 | [92] | 1 |
| miR-197 | circ_0039569 | HMGA1 | [93] | 1 |
| miR-200a | undefined | FOXA2 | [94] | no |
| miR-200b | undefined | TIMP2 | [95] | no |
| miR-200c | MALAT1 | undefined | [96] | 1 |
| miR-200c | PTENP1 | PTEN | [97] | 1 |
| miR-200c | undefined | BMI-1 | [98] | 1 |
| miR-200c | undefined | BRD7 | [99] | 1 |
| miR-200c-3p | RP11-379k17.4 | NOXA | [100] | 1 |
| miR-204 | undefined | FOXC1 | [101] | 3D |
| miR-204-5p | UCA1 | undefined | [102] | 3D |
| miR-204-5p | undefined | TrkB | [103] | 3D |
| miR-205 | undefined | ESRRG | [104] | 1 |
| miR-205 | undefined | PTEN | [105] | 1 |
| miR-205-5p | LA16c-313D11.11 | PTEN | [106] | 1 |
| miR-205-5p | RP11-395G23.3 | PTEN | [107] | 1 |
| miR-206 | undefined | HDAC6 | [108] | no |
| miR-210-3p | undefined | RUNX1T1 | [109] | no |
| miR-214 | DANCR | undefined | [110] | no |
| miR-21-5p | undefined | SOX17 | [111] | no |
| miR-216a | MEG3 | PD-L1 | [112] | 1 |
| miR-217 | circ_0023404 | MAPK1 | [113] | 2C |
| miR-218 | undefined | ADD2 | [114] | no |
| miR-222-3p | GAS5 | p27 | [115] | 1 |
| miR-223 | undefined | IGF-1R | [116] | 1 |
| miR-23b | TUSC7 | undefined | [117] | 3B |
| miR-23c | LINC01410 | CHD7 | [118] | no |
| miR-25 | undefined | BIM | [37] | 3E |
| miR-25-3p | SNHG5 | BTG2 | [119] | 3E |
| miR-26a/b-5p | NEAT1 | STAT3 | [120] | 1 |
| miR-27a | LINC00261 | FOXO1 | [74] | 2A |
| miR-27a-5p | undefined | SMAD4 | [121] | 2A |
| miR-27b | RBAT1 | undefined | [122] | no |
| miR-27b-3p | undefined | MARCH7 | [123] | no |
| miR-28-5p | LOXL1-AS1 | RAP1B | [124] | no |
| miR-298 | circ_0005797 | CTNND1 | [125] | 1 |
| miR-299 | TUG1 | VEGFA | [126] | 1 |
| miR-29a-3p | circ_FOXO3 | HDAC4 | [127] | no |
| miR-29b | undefined | PTEN | [128] | 1 |
| miR-29b | undefined | VEGFA | [129] | 1 |
| miR-302a-3p | LINC01016 | NFYA | [130] | * |
| miR-302a-5p | undefined | HMGA2 | [131] | 1 |
| miR-30c | undefined | MTA1 | [132] | no |
| miR-3130-3p | LINC01016 | NFYA | [130] | * |
| miR-320a | undefined | IGF-1R | [133] | 1 |
| miR-320a | undefined | eIF4E | [134] | 1 |
| miR-324-5p | circ_0067835 | HMGA1 | [135] | 1 |
| miR-326 | undefined | SUCNR1/GPR91 | [136] | 2B |
| miR-326 | undefined | TWIST1 | [137] | 2B |
| miR-329-3p | undefined | E2F1 | [138] | no |
| miR-335 | undefined | RBM10 | [139] | no |
| miR-340-5p | undefined | eIF4E | [134] | 1 |
| miR-34a | OC1 | PD-L1 | [140] | 1 |
| miR-34a | undefined | L1CAM | [141] | 1 |
| miR-34a | undefined | NOTCH1 | [142] | 1 |
| miR-34a-5p | TUG1 | VEGFA | [126] | 1 |
| miR-34c | undefined | E2F3 | [143] | no |
| miR-34c | undefined | IL-6R | [144] | no |
| miR-361 | GATA3-AS1 | ARRB2 | [145] | 1 |
| miR-361 | NEAT1 | KPNA4 | [146] | 1 |
| miR-361 | NEAT1 | MEF2D | [146] | 1 |
| miR-361 | NEAT1 | PDE4B | [146] | 1 |
| miR-361 | NEAT1 | ROCK1 | [146] | 1 |
| miR-361 | NEAT1 | STAT3 | [146] | 1 |
| miR-361 | NEAT1 | VEGFA | [146] | 1 |
| miR-361 | NEAT1 | WNT7A | [146] | 1 |
| miR-361-3p | circ_0075960 | SH2B1 | [147] | 1 |
| miR-363 | NR2F1-AS1 | SOX4 | [148] | 1 |
| miR-367-3p | undefined | HMGA2 | [131] | 1 |
| miR-372 | undefined | RhoC | [149] | no |
| miR-377-3p | circ_ZNF608 | COX2 | [150] | 1 |
| miR-377-3p | VPS9D1-AS1 | SGK1 | [151] | 1 |
| miR-378a-3p | BLACAT2 | YY1 | [152] | no |
| miR-381 | undefined | IGF-1R | [153] | 1 |
| miR-409 | undefined | SMAD2 | [154] | no |
| miR-424 | undefined | CPEB2 | [155] | 1 |
| miR-424 | undefined | E2F7 | [156] | 1 |
| miR-424 | undefined | GPER | [157] | 1 |
| miR-424 | undefined | IGF-1R | [158] | 1 |
| miR-4429 | SNHG12 | MMP2 | [159] | 2A |
| miR-4429 | SNHG12 | MMP9 | [159] | 2A |
| miR-449a | LINC01106 | MET | [160] | 2B |
| miR-449a | undefined | CDC25A | [161] | 2B |
| miR-449a | undefined | NDRG1 | [162] | 2B |
| miR-449a | undefined | SRC | [163] | 2B |
| miR-455 | DLEU2 | HK2 | [78] | 2D |
| miR-485-5p | LINC01224 | AKT3 | [164] | no |
| miR-486-5p | undefined | MARK1 | [165] | no |
| miR-490 | DLEU1 | SP1 | [166] | 2D |
| miR-490-3p | SNHG16 | HK2 | [167] | 2D |
| miR-494-3p | undefined | PTEN | [168] | 1 |
| miR-495 | undefined | PIK3R1 | [169] | no |
| miR-497-5p | circ_0011324 | mTOR | [77] | 2E |
| miR-497-5p | SNHG25 | FASN | [170] | 2E |
| miR-498-5p | HOXB-AS3 | ADAM9 | [171] | no |
| miR-505 | undefined | TGFA | [172] | no |
| miR-520h | circ_0001860 | SMAD7 | [173] | no |
| miR-522 | undefined | MAOB | [174] | no |
| miR-543 | circ_CCL22 | CDC25A | [175] | 2B |
| miR-543 | undefined | FAK | [176] | 2B |
| miR-543 | undefined | TWIST1 | [177] | 2B |
| miR-545-3p | AFAP-AS1 | VEGFA | [177] | 1 |
| miR-545-3p | circ_IFT80 | FAM98A | [178] | 1 |
| miR-589-5p | undefined | TRIP6 | [179] | no |
| miR-6076 | CHL1-AS1 | CHL1 | [180] | no |
| miR-612 | H19 | HOXA10 | [181] | 1 |
| miR-612 | PVT1 | CENP-H | [182] | 1 |
| miR-616 | TUSC7 | SOCS4 | [183] | 3B |
| miR-616 | TUSC7 | SOCS5 | [184] | 3B |
| miR-625 | circ_0007534 | ZEB2 | [184] | 1 |
| miR-625-5p | circ_0002577 | IGF-1R | [185] | 1 |
| miR-626 | circ_0000437 | CDKN1B | [186] | no |
| miR-636 | MONC | GLCE | [187] | no |
| miR-638 | undefined | MEF2C | [188] | no |
| miR-646 | circ_0001610 | STAT3 | [189] | 1 |
| miR-646 | circ_WHSC1 | NPM1 | [190] | 1 |
| miR-646 | HOTAIR | NPM1 | [191] | 1 |
| miR-652 | undefined | RORA | [192] | no |
| miR-655-3p | SNHG14 | undefined | [193] | 3A |
| miR-7-2-3p | BMPR1B-AS1 | DCLK1 | [194] | no |
| miR-876-3p | circ_NAB1 | CDKN3 | [195] | no |
| miR-92a-3p | DCST1-AS1 | NOTCH1 | [196] | 1 |
| miR-93 | undefined | FOXA1 | [197] | 3A |
| miR-93-5p | SNHG14 | ZBTB7A | [198] | 3A |
| miR-940 | undefined | MRVI1 | [199] | no |
| miR-944 | undefined | CADM2 | [200] | no |
| miR-96 | LINC00261 | FOXO1 | [74] | 2A |
| miR-99a | undefined | AKT1 | [201] | 2E |
| miR-99a | undefined | mTOR | [201] | 2E |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Piergentili, R.; Sechi, S.; De Paola, L.; Zaami, S.; Marinelli, E. Building a Hand-Curated ceRNET for Endometrial Cancer, Striving for Clinical as Well as Medicolegal Soundness: A Systematic Review. Non-Coding RNA 2025, 11, 34. https://doi.org/10.3390/ncrna11030034
Piergentili R, Sechi S, De Paola L, Zaami S, Marinelli E. Building a Hand-Curated ceRNET for Endometrial Cancer, Striving for Clinical as Well as Medicolegal Soundness: A Systematic Review. Non-Coding RNA. 2025; 11(3):34. https://doi.org/10.3390/ncrna11030034
Chicago/Turabian StylePiergentili, Roberto, Stefano Sechi, Lina De Paola, Simona Zaami, and Enrico Marinelli. 2025. "Building a Hand-Curated ceRNET for Endometrial Cancer, Striving for Clinical as Well as Medicolegal Soundness: A Systematic Review" Non-Coding RNA 11, no. 3: 34. https://doi.org/10.3390/ncrna11030034
APA StylePiergentili, R., Sechi, S., De Paola, L., Zaami, S., & Marinelli, E. (2025). Building a Hand-Curated ceRNET for Endometrial Cancer, Striving for Clinical as Well as Medicolegal Soundness: A Systematic Review. Non-Coding RNA, 11(3), 34. https://doi.org/10.3390/ncrna11030034