


Diagnostic Potential of Exosomal and Non-Exosomal Biomarkers in Lung Cancer: A Comparative Analysis Using a Rat Model of Lung Carcinogenesis

 ,
, 
Abstract
1. Introduction
2. Results
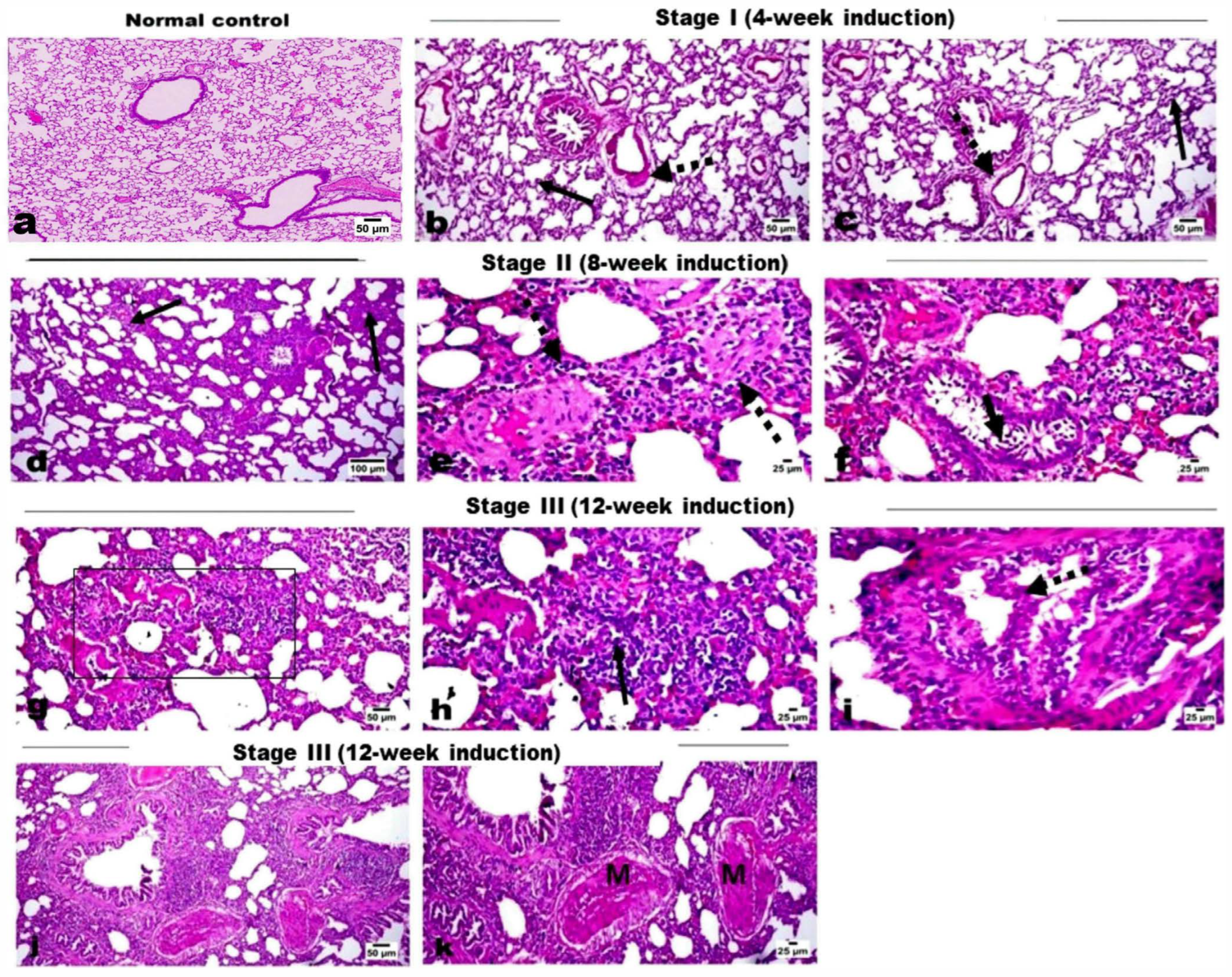
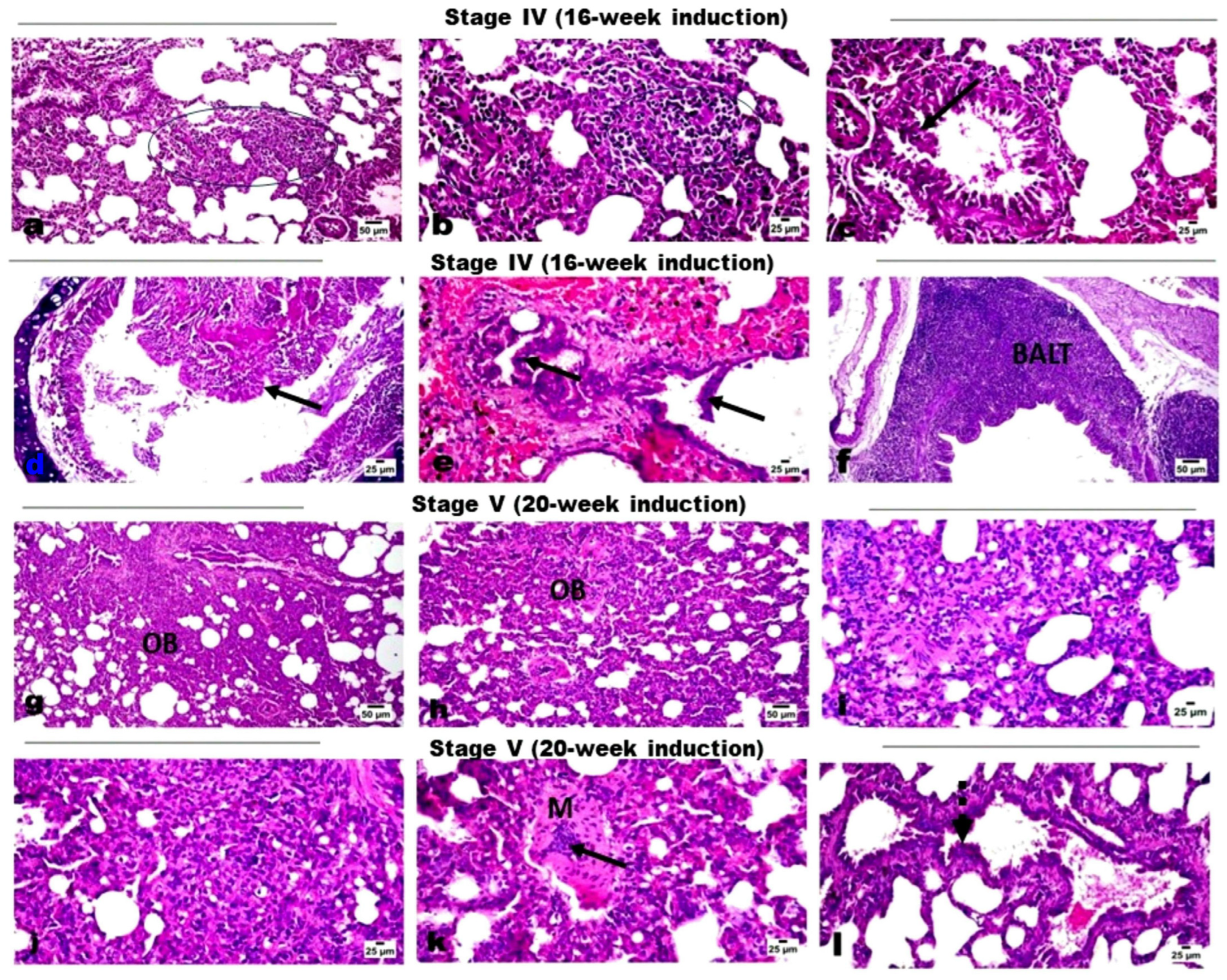
2.1. Histopathological Evaluation
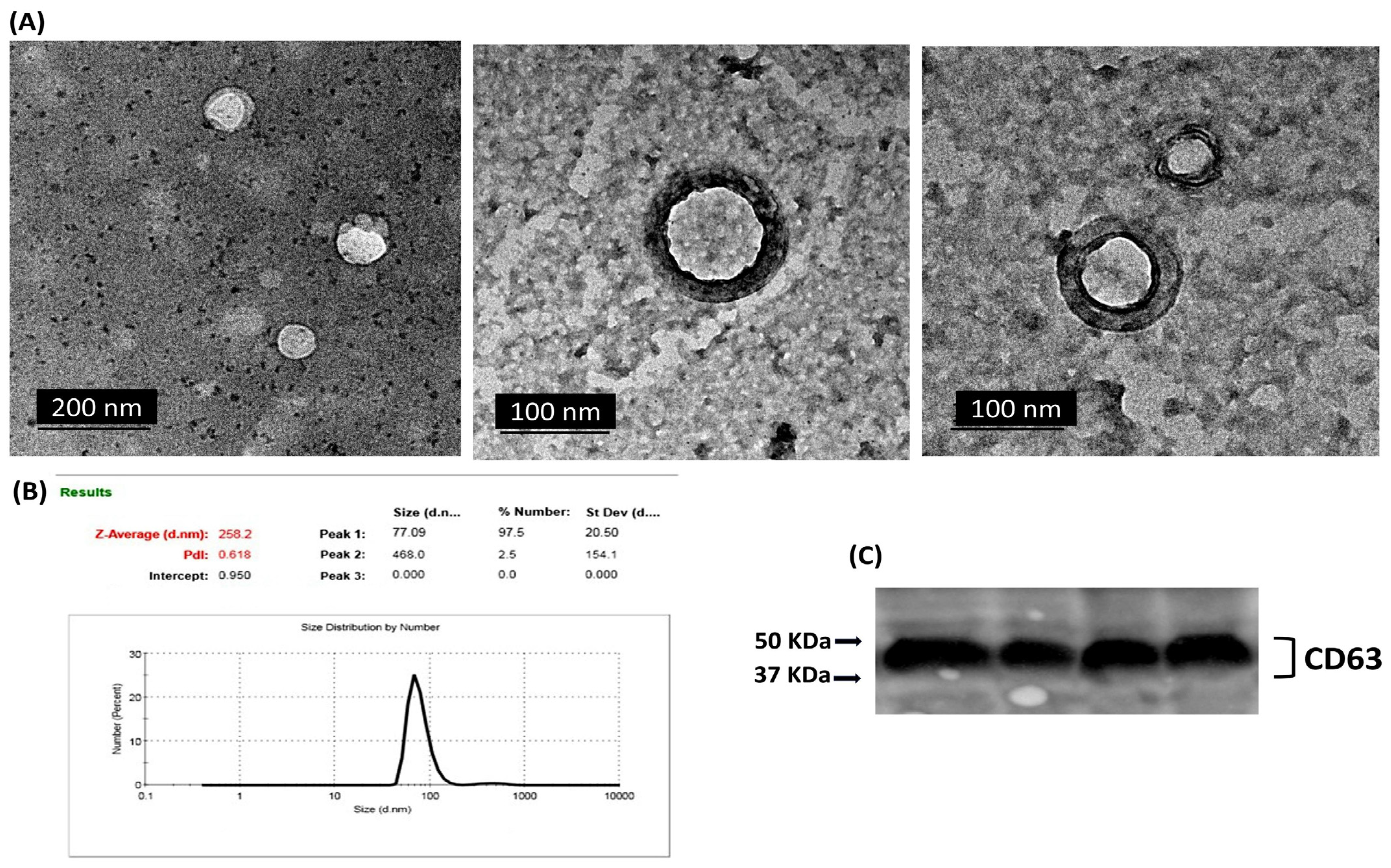
2.2. Characterization of Isolated Exosomes
2.3. Expression Patterns of miRNAs in Exosomal and Non-Exosomal Fractions
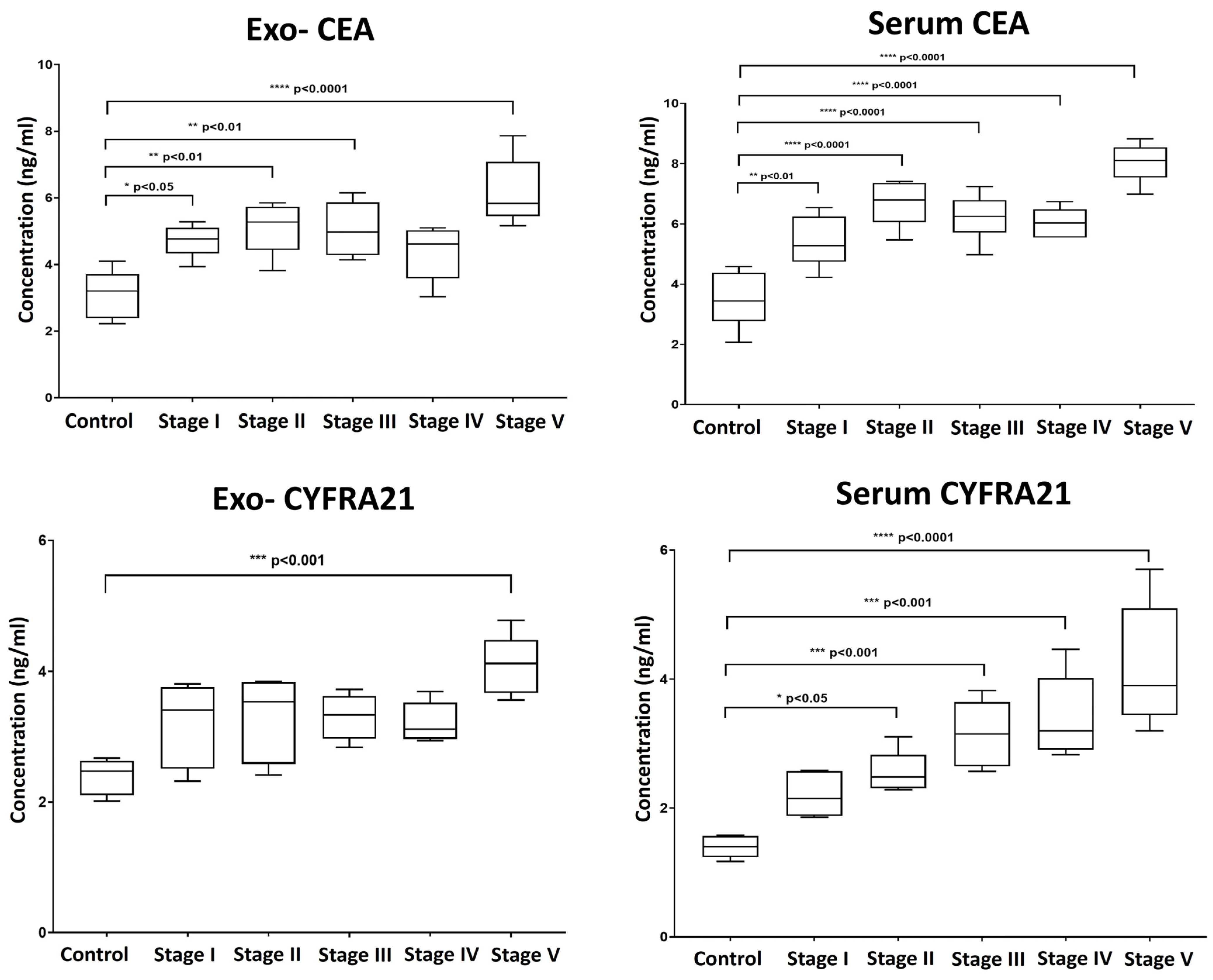
2.4. Levels of CEA and CYPF21 in Serum and Exosomes
2.5. Diagnostic Performance of Exosomal and Non-Exosomal miRNAs and Protein Tumor Markers
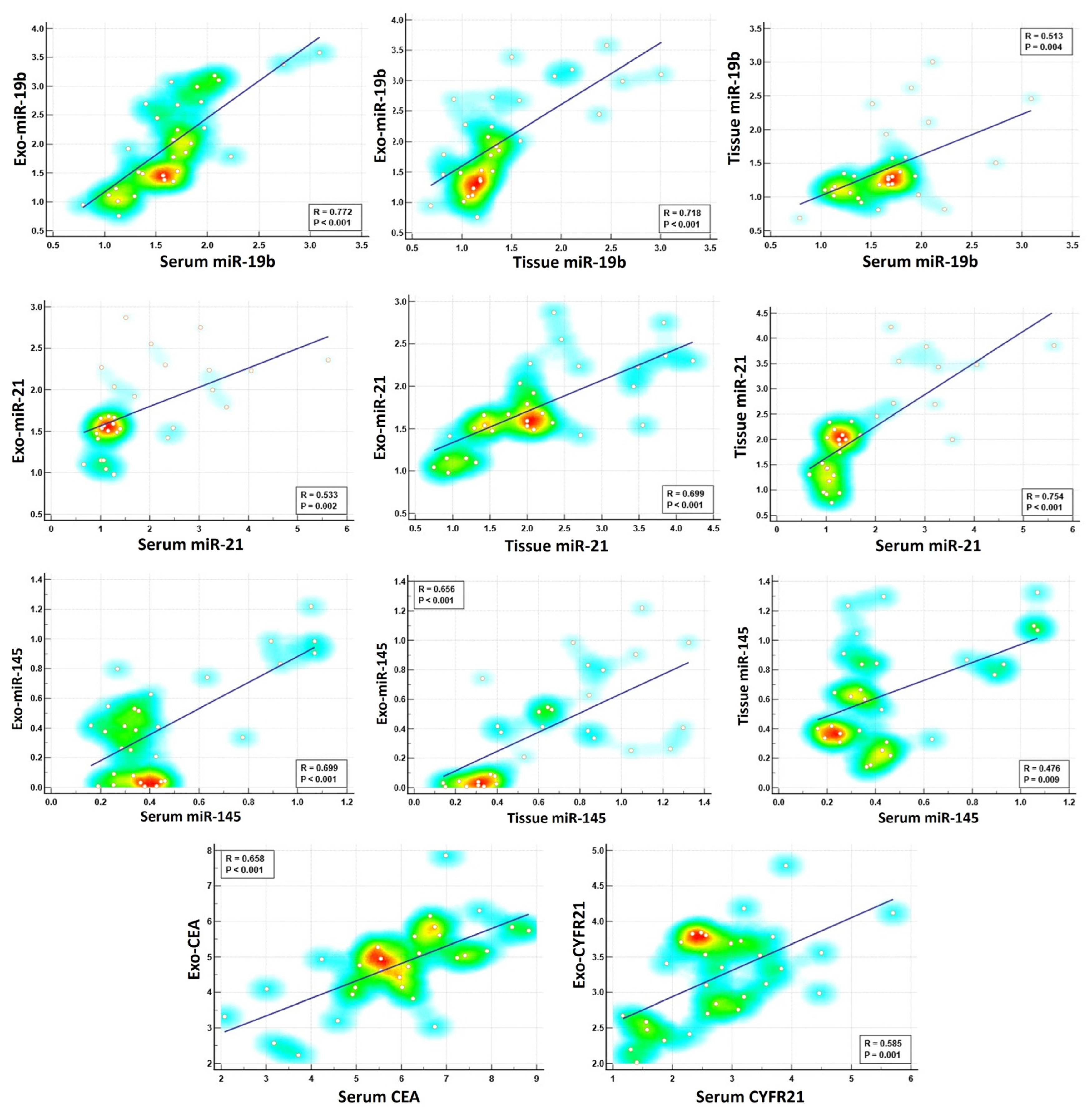
2.6. Correlation Between Exosomal and Non-Exosomal Biomarkers
3. Discussion
4. Materials and Methods
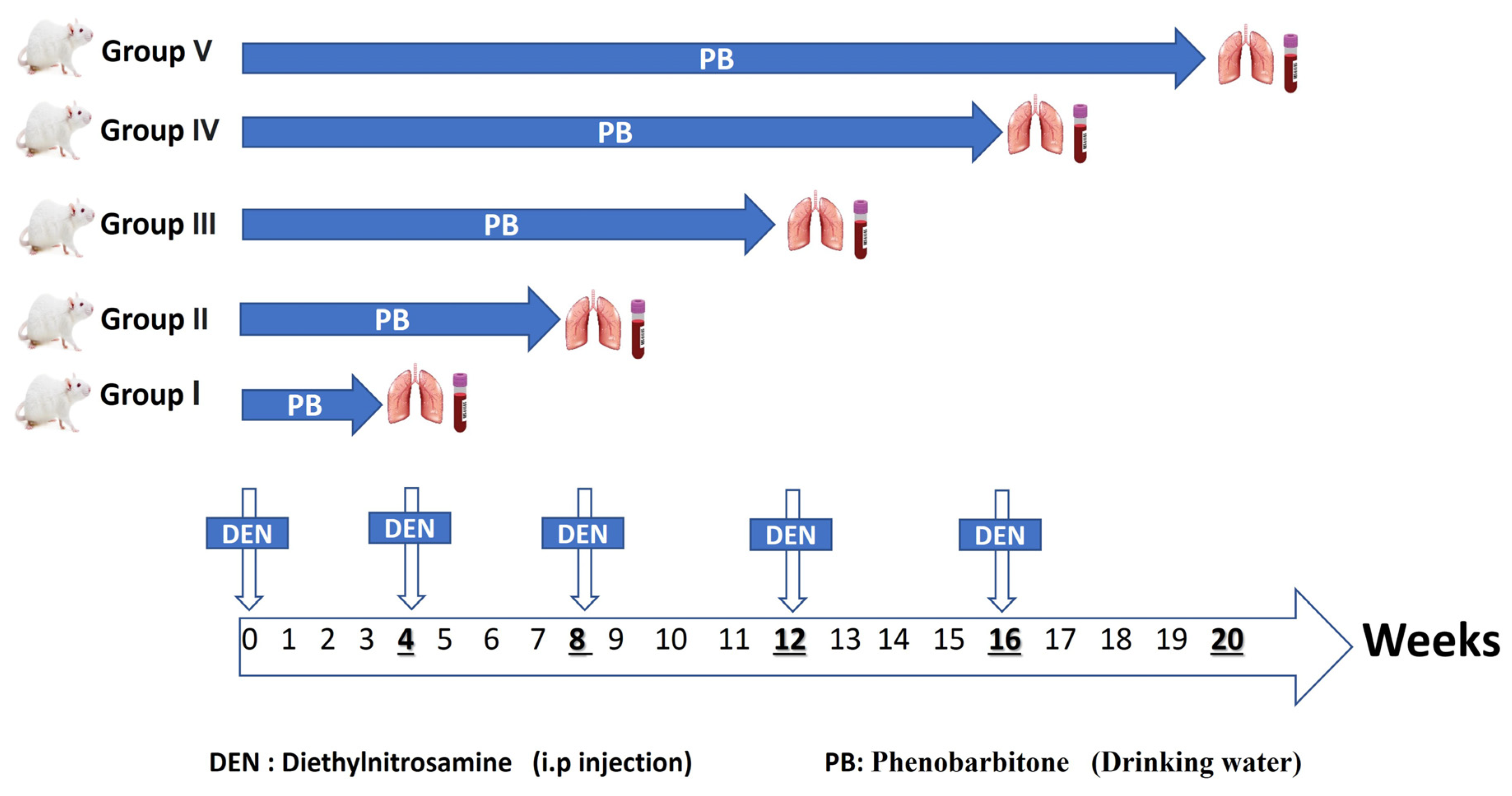
4.1. Experimental Animals
4.2. Induction of Lung Carcinogenesis
4.3. Histopathological Examination
4.4. Exosome Isolation and Characterization
4.4.1. Exosome Isolation
4.4.2. Transmission Electron Microscopy (TEM) and Dynamic Light Scattering (DLS)
4.4.3. Total Protein Content
4.4.4. Western Blot
4.5. Selection of Candidate miRNAs
4.6. RNA Extraction from Isolated Exosomes, Serum, and Tissue Samples and Expression Analysis
4.7. Evaluating Levels of Serum and Exosomal Carcinoembryonic Antigen (CEA) and Cytokeratin Fragment Antigen-21 (CYFRA 21-1)
4.8. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| Ago | Argonaute |
| AHSG | Alpha-2-HS-Glycoprotein |
| aGvHD | Acute Graft-versus-Host Disease |
| CEA | Carcinoembryonic Antigen |
| CYFRA 21-1 | Cytokeratin Fragment Antigen-21 |
| DEN | N-Diethylnitrosamine |
| DLS | Dynamic Light Scattering |
| ELISA | Enzyme-linked Immunosorbent Assay |
| EV | Extracellular Vesicle |
| HDL | High-density lipoproteins |
| LRG1 | Leucine-Rich a2-Glycoprotein |
| NSCLC | Non-Small Cell Lung Cancer |
| PB | Phenobarbitone |
| PTEN | Phosphatase and Tensin Homolog |
| TEM | Transmission Electron Microscopy |
References
- Kratzer, T.B.; Bandi, P.; Freedman, N.D.; Smith, R.A.; Travis, W.D.; Jemal, A.; Siegel, R.L. Lung Cancer Statistics. Cancer 2023, 2024, 130. [Google Scholar]
- Uprety, D.; Parikh, K.; Sawkar, A.; Dimou, A.; Leventakos, K. Changing Paradigm in Advanced and Metastatic Non-Small Cell Lung Cancer. J. Thorac. Dis. 2020, 12, 6992–7001. [Google Scholar] [CrossRef] [PubMed]
- Casagrande, G.M.S.; de Oliveira Silva, M.; Reis, R.M.; Leal, L.F. Liquid Biopsy for Lung Cancer: Up-to-Date and Perspectives for Screening Programs. Int. J. Mol. Sci. 2023, 24, 2505. [Google Scholar] [CrossRef] [PubMed]
- Winkle, M.; El-Daly, S.M.; Fabbri, M.; Calin, G.A. Noncoding RNA Therapeutics—Challenges and Potential Solutions. Nat. Rev. Drug Discov. 2021, 20, 629–651. [Google Scholar] [CrossRef] [PubMed]
- Valihrach, L.; Androvic, P.; Kubista, M. Circulating MiRNA Analysis for Cancer Diagnostics and Therapy. Mol. Asp. Med. 2020, 72, 100825. [Google Scholar] [CrossRef]
- El-Daly, S.M.; Gouhar, S.A.; Abd Elmageed, Z.Y. Circulating MicroRNAs as Reliable Tumor Biomarkers: Opportunities and Challenges Facing Clinical Application. J. Pharmacol. Exp. Ther. 2023, 384, 35–51. [Google Scholar] [CrossRef]
- Drula, R.; Ott, L.F.; Berindan-Neagoe, I.; Pantel, K.; Calin, G.A. Micrornas from Liquid Biopsy Derived Extracellular Vesicles: Recent Advances in Detection and Characterization Methods. Cancers 2020, 12, 2009. [Google Scholar] [CrossRef]
- Witwer, K.W.; Buzás, E.I.; Bemis, L.T.; Bora, A.; Lässer, C.; Lötvall, J.; Nolte-‘t Hoen, E.N.; Piper, M.G.; Sivaraman, S.; Skog, J.; et al. Standardization of Sample Collection, Isolation and Analysis Methods in Extracellular Vesicle Research. J. Extracell. Vesicles 2013, 2, 20360. [Google Scholar] [CrossRef]
- Zhang, Y.; Liu, Y.; Liu, H.; Tang, W.H. Exosomes: Biogenesis, Biologic Function and Clinical Potential. Cell Biosci. 2019, 9, 19. [Google Scholar] [CrossRef]
- Zhou, B.; Xu, K.; Zheng, X.; Chen, T.; Wang, J.; Song, Y.; Shao, Y.; Zheng, S. Application of Exosomes as Liquid Biopsy in Clinical Diagnosis. Signal Transduct. Target. Ther. 2020, 5, 144. [Google Scholar] [CrossRef]
- Yu, D.; Li, Y.; Wang, M.; Gu, J.; Xu, W.; Cai, H.; Fang, X.; Zhang, X. Exosomes as a New Frontier of Cancer Liquid Biopsy. Mol. Cancer 2022, 21, 56. [Google Scholar] [CrossRef] [PubMed]
- Kamal, N.N.S.B.N.M.; Shahidan, W.N.S. Non-Exosomal and Exosomal Circulatory MicroRNAs: Which Are More Valid as Biomarkers? Front. Pharmacol. 2020, 10, 1500. [Google Scholar]
- Sanz-Rubio, D.; Martin-Burriel, I.; Gil, A.; Cubero, P.; Forner, M.; Khalyfa, A.; Marin, J.M. Stability of Circulating Exosomal MiRNAs in Healthy Subjects Article. Sci. Rep. 2018, 8, 10306. [Google Scholar] [CrossRef] [PubMed]
- Endzelinš, E.; Berger, A.; Melne, V.; Bajo-Santos, C.; Sobolevska, K.; Abols, A.; Rodriguez, M.; Šantare, D.; Rudnickiha, A.; Lietuvietis, V.; et al. Detection of Circulating MiRNAs: Comparative Analysis of Extracellular Vesicle-Incorporated MiRNAs and Cell-Free MiRNAs in Whole Plasma of Prostate Cancer Patients. BMC Cancer 2017, 17, 730. [Google Scholar] [CrossRef] [PubMed]
- Tian, L.; Shan, W.; Zhang, Y.; Lv, X.; Li, X.; Wei, C. Up-Regulation of MiR-21 Expression Predicate Advanced Clinicopathological Features and Poor Prognosis in Patients with Non-Small Cell Lung Cancer. Pathol. Oncol. Res. 2016, 22, 161–167. [Google Scholar] [CrossRef]
- Xue, X.; Liu, Y.; Wang, Y.; Meng, M.; Wang, K.; Zang, X.; Zhao, S.; Sun, X.; Cui, L.; Pan, L.; et al. MiR-21 and MiR-155 Promote Non-Small Cell Lung Cancer Progression by Downregulating SOCS1, SOCS6, and PTEN. Oncotarget 2016, 7, 84508–84519. [Google Scholar] [CrossRef]
- Baumgartner, U.; Berger, F.; Hashemi Gheinani, A.; Burgener, S.S.; Monastyrskaya, K.; Vassella, E. MiR-19b Enhances Proliferation and Apoptosis Resistance via the EGFR Signaling Pathway by Targeting PP2A and BIM in Non-Small Cell Lung Cancer. Mol. Cancer 2018, 17, 44. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Zeng, H.; Guo, Y.; Liu, P.; Pan, H.; Deng, A.; Hu, J. MiRNA-145 Inhibits Non-Small Cell Lung Cancer Cell Proliferation by Targeting c-Myc. J. Exp. Clin. Cancer Res. 2010, 29, 151. [Google Scholar] [CrossRef]
- Ling, D.J.; Chen, Z.S.; Zhang, Y.D.; Liao, Q.D.; Feng, J.X.; Zhang, X.Y.; Shi, T.S. MicroRNA-145 Inhibits Lung Cancer Cell Metastasis. Mol. Med. Rep. 2015, 11, 3108–3114. [Google Scholar] [CrossRef]
- Zhang, H.; Mao, F.; Shen, T.; Luo, Q.; Ding, Z.; Qian, L.; Huang, J. Plasma MiR-145, MiR-20a, MiR-21 and MiR-223 as Novel Biomarkers for Screening Early-Stage Non-Small Cell Lung Cancer. Oncol. Lett. 2017, 13, 669–676. [Google Scholar] [CrossRef]
- Zhang, Z.H.; Han, Y.W.; Liang, H.; Wang, L.M. Prognostic Value of Serum CYFRA21-1 and CEA for Non-Small-Cell Lung Cancer. Cancer Med. 2015, 4, 1633–1638. [Google Scholar] [CrossRef] [PubMed]
- Anand, S.; Samuel, M.; Kumar, S.; Mathivanan, S. Ticket to a Bubble Ride: Cargo Sorting into Exosomes and Extracellular Vesicles. Biochim. Biophys. Acta Proteins Proteom. 2019, 1867, 140203. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.; Cao, Y.; He, Z.; He, J.; Hu, C.; Duan, H.; Jiang, J. Serum Levels of MiR-19b and MiR-146a as Prognostic Biomarkers for Non-Small Cell Lung Cancer. Tohoku J. Exp. Med. 2014, 232, 85–95. [Google Scholar] [CrossRef]
- Fan, Y.; Yin, S.; Hao, Y.; Yang, J.; Zhang, H.; Sun, C.; Ma, M.; Chang, Q.; Xi, J.J. MiR-19b Promotes Tumor Growth and Metastasis via Targeting TP53. RNA 2014, 20, 765–772. [Google Scholar] [CrossRef]
- Lin, Q.; Chen, T.; Lin, Q.; Lin, G.; Lin, J.; Chen, G.; Guo, L. Serum MiR-19a Expression Correlates with Worse Prognosis of Patients with Non-Small Cell Lung Cancer. J. Surg. Oncol. 2013, 107, 767–771. [Google Scholar] [CrossRef]
- Wang, L.; Wu, J.; Ye, N.; Li, F.; Zhan, H.; Chen, S.; Xu, J. Plasma-Derived Exosome MiR-19b Acts as a Diagnostic Marker for Pancreatic Cancer. Front. Oncol. 2021, 11, 11–2021. [Google Scholar] [CrossRef]
- Feng, Y.H.; Tsao, C.J. Emerging Role of MicroRNA-21 in Cancer (Review). Biomed. Rep. 2016, 5, 395–402. [Google Scholar] [CrossRef]
- El-Daly, S.M.; Morsy, S.M.; Medhat, D.; El-Bana, M.A.; Latif, Y.A.; Omara, E.A.; Awadallah, J.R.; Gamal-Eldeen, A.M. The Diagnostic Efficacy of Circulating MiRNAs in Monitoring the Early Development of Colitis-Induced Colorectal Cancer. J. Cell Biochem. 2019, 120, 16668–16680. [Google Scholar] [CrossRef]
- Bica-Pop, C.; Cojocneanu-Petric, R.; Magdo, L.; Raduly, L.; Gulei, D.; Berindan-Neagoe, I. Overview upon MiR-21 in Lung Cancer: Focus on NSCLC. Cell. Mol. Life Sci. 2018, 75, 3539–3551. [Google Scholar] [CrossRef]
- Bautista-Sánchez, D.; Arriaga-Canon, C.; Pedroza-Torres, A.; De La Rosa-Velázquez, I.A.; González-Barrios, R.; Contreras-Espinosa, L.; Montiel-Manríquez, R.; Castro-Hernández, C.; Fragoso-Ontiveros, V.; Álvarez-Gómez, R.M.; et al. The Promising Role of MiR-21 as a Cancer Biomarker and Its Importance in RNA-Based Therapeutics. Mol. Ther. Nucleic Acids 2020, 20, 409–420. [Google Scholar] [CrossRef]
- Soliman, S.E.S.; Abdelaleem, A.H.; Alhanafy, A.M.; Ibrahem, R.A.L.; Elhaded, A.S.A.; Assar, M.F.A. Circulating MiR-21-5p and MiR-126-3p: Diagnostic, Prognostic Value, and Multivariate Analysis in Non-Small-Cell Lung Cancer. Mol. Biol. Rep. 2021, 48, 2543–2552. [Google Scholar] [CrossRef]
- Gao, W.; Lu, X.; Liu, L.; Xu, J.; Feng, D.; Shu, Y. MiRNA-21: A Biomarker Predictive for Platinum-Based Adjuvant Chemotherapy Response in Patients with Non-Small Cell Lung Cancer. Cancer Biol. Ther. 2012, 13, 330–340. [Google Scholar] [CrossRef]
- Catelan, S.; Olioso, D.; Santangelo, A.; Scapoli, C.; Tamanini, A.; Pinna, G.; Sala, F.; Lippi, G.; Nicolato, A.; Cabrini, G.; et al. MiRNAs in Serum Exosomes for Differential Diagnosis of Brain Metastases. Cancers 2022, 14, 3493. [Google Scholar] [CrossRef]
- Shi, J. Considering Exosomal MiR-21 as a Biomarker for Cancer. J. Clin. Med. 2016, 5, 42. [Google Scholar] [CrossRef]
- Hang, W.; Feng, Y.; Sang, Z.; Yang, Y.; Zhu, Y.; Huang, Q.; Xi, X. Downregulation of MiR-145-5p in Cancer Cells and Their Derived Exosomes May Contribute to the Development of Ovarian Cancer by Targeting CT. Int. J. Mol. Med. 2019, 43, 256–266. [Google Scholar] [CrossRef]
- Zhao, C.; Xu, Y.; Zhang, Y.; Tan, W.; Xue, J.; Yang, Z.; Zhang, Y.; Lu, Y.; Hu, X. Downregulation of MiR-145 Contributes to Lung Adenocarcinoma Cell Growth to Form Brain Metastases. Oncol. Rep. 2013, 30, 2027–2034. [Google Scholar] [CrossRef]
- Yin, Y.; Yan, Z.P.; Lu, N.N.; Xu, Q.; He, J.; Qian, X.; Yu, J.; Guan, X.; Jiang, B.H.; Liu, L.Z. Downregulation of MiR-145 Associated with Cancer Progression and VEGF Transcriptional Activation by Targeting N-RAS and IRS1. Biochim. Biophys. Acta Gene Regul. Mech. 2013, 1829, 239–247. [Google Scholar] [CrossRef]
- El-Toukhy, S.E.; El-Daly, S.M.; Kamel, M.M.; Nabih, H.K. The Diagnostic Significance of Circulating MiRNAs and Metabolite Profiling in Early Prediction of Breast Cancer in Egyptian Women. J. Cancer Res. Clin. Oncol. 2022, 149, 5437–5451. [Google Scholar] [CrossRef]
- Ye, Z.; Shen, N.; Weng, Y.; Li, K.; Hu, L.; Liao, H.; An, J.; Liu, L.; Lao, S.; Cai, S. Low MiR-145 Silenced by DNA Methylation Promotes NSCLC Cell Proliferation, Migration and Invasion by Targeting Mucin 1. Cancer Biol. Ther. 2015, 16, 1071–1079. [Google Scholar] [CrossRef]
- Sachdeva, M.; Mo, Y.Y. MicroRNA-145 Suppresses Cell Invasion and Metastasis by Directly Targeting Mucin 1. Cancer Res. 2010, 70, 378–387. [Google Scholar] [CrossRef]
- Tao, S.; Ju, X.; Zhou, H.; Zeng, Q. Circulating MicroRNA-145 as a Diagnostic Biomarker for Non-Small-Cell Lung Cancer: A Systemic Review and Meta-Analysis. Int. J. Biol. Markers 2020, 35, 51–60. [Google Scholar] [CrossRef]
- Geater, S.L.; Chaniad, P.; Trakunram, K.; Keeratichananont, W.; Buya, S.; Thongsuksai, P.; Raungrut, P. Diagnostic and Prognostic Value of Serum MiR-145 and Vascular Endothelial Growth Factor in Non-Small Cell Lung Cancer. Oncol. Lett. 2022, 23, 1–11. [Google Scholar] [CrossRef]
- Groot, M.; Lee, H. Sorting Mechanisms for MicroRNAs into Extracellular Vesicles and Their Associated Diseases. Cells 2020, 9, 1044. [Google Scholar] [CrossRef]
- Drula, R.; Pardini, B.; Fu, X.; De los Santos, M.C.; Jurj, A.; Pang, L.; El-Daly, S.M.; Fabris, L.; Knutsen, E.; Dragomir, M.P.; et al. 17β-Estradiol Promotes Extracellular Vesicle Release and Selective MiRNA Loading in ERα-Positive Breast Cancer. Proc. Natl. Acad. Sci. USA 2023, 120, e2122053120. [Google Scholar] [CrossRef]
- Koppers-Lalic, D.; Hackenberg, M.; Bijnsdorp, I.V.; van Eijndhoven, M.A.J.; Sadek, P.; Sie, D.; Zini, N.; Middeldorp, J.M.; Ylstra, B.; de Menezes, R.X.; et al. Nontemplated Nucleotide Additions Distinguish the Small RNA Composition in Cells from Exosomes. Cell Rep. 2014, 8, 1649–1658. [Google Scholar] [CrossRef]
- Crossland, R.E.; Norden, J.; Juric, M.K.; Pearce, K.F.; Lendrem, C.; Bibby, L.A.; Collin, M.; Greinix, H.T.; Dickinson, A.M. Serum and Extracellular Vesicle MicroRNAs MiR-423, MiR-199, and MiR-93* as Biomarkers for Acute Graft-versus-Host Disease. Front. Immunol. 2017, 8, 446–1446. [Google Scholar] [CrossRef]
- Jakobsen, K.R.; Paulsen, B.S.; Bæk, R.; Varming, K.; Sorensen, B.S.; Jørgensen, M.M. Exosomal Proteins as Potential Diagnostic Markers in Advanced Non-Small Cell Lung Carcinoma. J. Extracell. Vesicles 2015, 4, 26659. [Google Scholar] [CrossRef]
- Okamura, K.; Takayama, K.; Izumi, M.; Harada, T.; Furuyama, K.; Nakanishi, Y. Diagnostic Value of CEA and CYFRA 21-1 Tumor Markers in Primary Lung Cancer. Lung Cancer 2013, 80, 45–49. [Google Scholar] [CrossRef]
- Zhang, C.; Yang, X.; Jiang, T.; Yan, C.; Xu, X.; Chen, Z. Tissue-Derived Extracellular Vesicles: Isolation, Purification, and Multiple Roles in Normal and Tumor Tissues. Life Sci. 2023, 321, 121624. [Google Scholar] [CrossRef]
- De Seranno, S.; Meuwissen, R. Progress and Applications of Mouse Models for Human Lung Cancer. Eur. Respir. J. 2010, 35, 426–443. [Google Scholar] [CrossRef]
- Bai, J.; Wei, X.; Zhang, X.; Wu, C.; Wang, Z.; Chen, M.; Wang, J. Microfluidic Strategies for the Isolation and Profiling of Exosomes. TrAC-Trends Anal. Chem. 2023, 158, 116834. [Google Scholar] [CrossRef]
- Qian, C.; Xiao, Y.; Wang, J.; Li, Y.; Li, S.; Wei, B.; Du, W.; Feng, X.; Chen, P.; Liu, B.F. Rapid Exosomes Concentration and in Situ Detection of Exosomal MicroRNA on Agarose-Based Microfluidic Chip. Sens. Actuators B Chem. 2021, 333, 129559. [Google Scholar] [CrossRef]
- Tong, Z.; Yang, D.; Shen, C.; Li, C.; Xu, X.; Li, Q.; Wu, Z.; Ma, H.; Chen, F.; Mao, H. Rapid Automated Extracellular Vesicle Isolation and MiRNA Preparation on a Cost-Effective Digital Microfluidic Platform. Anal. Chim. Acta 2024, 1296, 342337. [Google Scholar] [CrossRef] [PubMed]
- Sivalingam, K.; Amirthalingam, V.; Ganasan, K.; Huang, C.Y.; Viswanadha, V.P. Neferine Suppresses Diethylnitrosamine-Induced Lung Carcinogenesis in Wistar Rats. Food Chem. Toxicol. 2019, 123, 385–398. [Google Scholar] [CrossRef] [PubMed]
- Giri, R.K.; Baral, R.N.; Das, B.R. Induction of Lung Carcinogenesis in AKR-Mice by N-Nitrosodiethylamine/Phenobarbitone, Associated with High Expression of c-Mye and c-Jun Oncoproteins. Cancer Lett. 1997, 112, 57–63. [Google Scholar] [CrossRef] [PubMed]
- Man, S.; Li, J.; Qiu, P.; Liu, J.; Liu, Z.; Ma, L.; Gao, W. Inhibition of Lung Cancer in Diethylnitrosamine-Induced Mice by Rhizoma Paridis Saponins. Mol. Carcinog. 2017, 56, 1405–1413. [Google Scholar] [CrossRef]
- van der Pol, E.; Coumans, F.A.W.; Grootemaat, A.E.; Gardiner, C.; Sargent, I.L.; Harrison, P.; Sturk, A.; van Leeuwen, T.G.; Nieuwland, R. Particle Size Distribution of Exosomes and Microvesicles Determined by Transmission Electron Microscopy, Flow Cytometry, Nanoparticle Tracking Analysis, and Resistive Pulse Sensing. J. Thromb. Haemost. 2014, 12, 1182–1192. [Google Scholar] [CrossRef]
- Lyu, T.S.; Ahn, Y.; Im, Y.J.; Kim, S.S.; Lee, K.H.; Kim, J.; Choi, Y.; Lee, D.; Kang, E.S.; Jin, G.; et al. The Characterization of Exosomes from Fibrosarcoma Cell and the Useful Usage of Dynamic Light Scattering (DLS) for Their Evaluation. PLoS ONE 2021, 16, e0231994. [Google Scholar] [CrossRef]
- Karttunen, J.; Heiskanen, M.; Navarro-Ferrandis, V.; Das Gupta, S.; Lipponen, A.; Puhakka, N.; Rilla, K.; Koistinen, A.; Pitkänen, A. Precipitation-Based Extracellular Vesicle Isolation from Rat Plasma Co-Precipitate Vesicle-Free MicroRNAs. J. Extracell. Vesicles 2019, 8, 1555410. [Google Scholar] [CrossRef]
- Zaporozhchenko, I.A.; Morozkin, E.S.; Skvortsova, T.E.; Ponomaryova, A.A.; Rykova, E.Y.; Cherdyntseva, N.V.; Polovnikov, E.S.; Pashkovskaya, O.A.; Pokushalov, E.A.; Vlassov, V.V.; et al. Plasma MiR-19b and MiR-183 as Potential Biomarkers of Lung Cancer. PLoS ONE 2016, 11, e0165261. [Google Scholar] [CrossRef]
- Aushev, V.N.; Zborovskaya, I.B.; Laktionov, K.K.; Girard, N.; Cros, M.P.; Herceg, Z.; Krutovskikh, V. Comparisons of MicroRNA Patterns in Plasma before and after Tumor Removal Reveal New Biomarkers of Lung Squamous Cell Carcinoma. PLoS ONE 2013, 8, e78649. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez, M.; Silva, J.; López-Alfonso, A.; López-Muñiz, M.B.; Peña, C.; Domínguez, G.; García, J.M.; López-Gónzalez, A.; Méndez, M.; Provencio, M.; et al. Different Exosome Cargo from Plasma/Bronchoalveolar Lavage in Non-Small-Cell Lung Cancer. Genes Chromosomes Cancer 2014, 53, 713–724. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | AUC | Cut-Off Value | Sensitivity % | Specificity % | p Value | 95% CI |
|---|---|---|---|---|---|---|
| Exo-miR-19b | 0.920 | 1.294 | 80.0% | 100.0% | 0.028 | 0.738–1.000 |
| Serum miR-19b | 1.000 | 1.215 | 100.0% | 100.0% | 0.009 | 1.000–1.000 |
| Tissue miR-19b | 0.880 | 1.170 | 80.0% | 100.0% | 0.047 | 0.640–1.000 |
| Exo-miR-21 | 1.000 | 1.279 | 100.0% | 100.0% | 0.009 | 1.000–1.000 |
| Serum miR-21 | 0.625 | 1.018 | 66.7% | 66.7% | 0.471 | 0.288–0.962 |
| Tissue miR-21 | 0.880 | 1.235 | 80.0% | 80.0% | 0.047 | 0.662–1.000 |
| Exo-miR-145 | 1.000 | 0.816 | 100.0% | 100.0% | 0.009 | 1.000–1.000 |
| Serum miR-145 | 1.000 | 0.663 | 100.0% | 100.0% | 0.004 | 1.000–1.000 |
| Tissue miR-145 | 0.520 | 1.058 | 60.0% | 60.0% | 0.917 | 0.135–0.905 |
| Exo-CEA | 0.960 | 4.413 | 80.0% | 100.0% | 0.016 | 0.843–1.000 |
| Serum CEA | 0.944 | 4.750 | 83.3% | 100.0% | 0.010 | 0.814–1.000 |
| Exo-CYFRA21 | 0.880 | 2.688 | 80.0% | 100.0% | 0.047 | 0.640–1.000 |
| Serum CYFRA21 | 1.000 | 1.719 | 100.0% | 100.0% | 0.009 | 1.000–1.000 |
| r (Correlation Coefficient) | p Value | |
|---|---|---|
| Exo-miR-19b with serum miR-19b | 0.772 ** | p < 0.001 |
| Exo-miR-19b with tissue miR-19b | 0.718 ** | p < 0.001 |
| Exo-miR-21 with serum miR-21 | 0.533 ** | p < 0.002 |
| Exo-miR-21 with tissue miR-21 | 0.699 ** | p < 0.001 |
| Exo-miR-145 with serum miR-145 | 0.699 ** | p < 0.001 |
| Exo-miR-145 with tissue miR-145 | 0.656 ** | p < 0.001 |
| Exo-CEA with serum CEA | 0.658 ** | P < 0.001 |
| Exo-CYFRA21 with serum CYFRA21 | 0.585 ** | p = 0.001 |
| Exo-CEA with Exo-CYFRA21 | 0.568 ** | p = 0.001 |
| Serum CEA with serum CYFRA21 | 0.745 ** | p < 0.001 |
| Exo-miR-19b with Exo-miR-21 | 0.676 ** | p < 0.001 |
| Exo-miR-19b with Exo-miR-145 | −0.790 ** | p < 0.001 |
| Exo-miR-21 with Exo-miR-145 | −0.622 ** | p < 0.001 |
| Exo-miR-19b with Exo-CEA | 0.486 ** | p = 0.007 |
| Exo-miR-19b with Exo-CYFRA21 | 0.527 ** | p = 0.003 |
| Exo-miR-21 with Exo-CEA | 0.361 | p = 0.05 |
| Exo-miR-21with Exo-CYFRA21 | 0.396 * | p = 0.03 |
| Exo miR-145 with Exo-CEA | −0.565 ** | p = 0.001 |
| Exo miR-145 with Exo-CYFRA21 | −0.668 ** | p < 0.001 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
El-Daly, S.M.; Abdelrahman, S.S.; Abd El-Jawad, A.M.; Abdel-Monem, M.A.; El-Saeed, G.S.M. Diagnostic Potential of Exosomal and Non-Exosomal Biomarkers in Lung Cancer: A Comparative Analysis Using a Rat Model of Lung Carcinogenesis. Non-Coding RNA 2025, 11, 47. https://doi.org/10.3390/ncrna11030047
El-Daly SM, Abdelrahman SS, Abd El-Jawad AM, Abdel-Monem MA, El-Saeed GSM. Diagnostic Potential of Exosomal and Non-Exosomal Biomarkers in Lung Cancer: A Comparative Analysis Using a Rat Model of Lung Carcinogenesis. Non-Coding RNA. 2025; 11(3):47. https://doi.org/10.3390/ncrna11030047
Chicago/Turabian StyleEl-Daly, Sherien M., Sahar S. Abdelrahman, Amira Mohamed Abd El-Jawad, Mahmoud A. Abdel-Monem, and Gamila S. M. El-Saeed. 2025. "Diagnostic Potential of Exosomal and Non-Exosomal Biomarkers in Lung Cancer: A Comparative Analysis Using a Rat Model of Lung Carcinogenesis" Non-Coding RNA 11, no. 3: 47. https://doi.org/10.3390/ncrna11030047
APA StyleEl-Daly, S. M., Abdelrahman, S. S., Abd El-Jawad, A. M., Abdel-Monem, M. A., & El-Saeed, G. S. M. (2025). Diagnostic Potential of Exosomal and Non-Exosomal Biomarkers in Lung Cancer: A Comparative Analysis Using a Rat Model of Lung Carcinogenesis. Non-Coding RNA, 11(3), 47. https://doi.org/10.3390/ncrna11030047