
Exploration of the Strategy for Improving the Expression of Heterologous Sweet Protein Monellin in Aspergillus niger


Abstract
1. Introduction
2. Materials and Methods
2.1. Strains and Media
2.2. Construction of Recombinant Plasmids
2.3. Genetic Regulation of A. niger Based on the CRISPR/Cas9 System
2.4. RNA Extraction and Quantitative Real-Time PCR (qRT-PCR) Analysis
2.5. Characterization of Monellin Expression
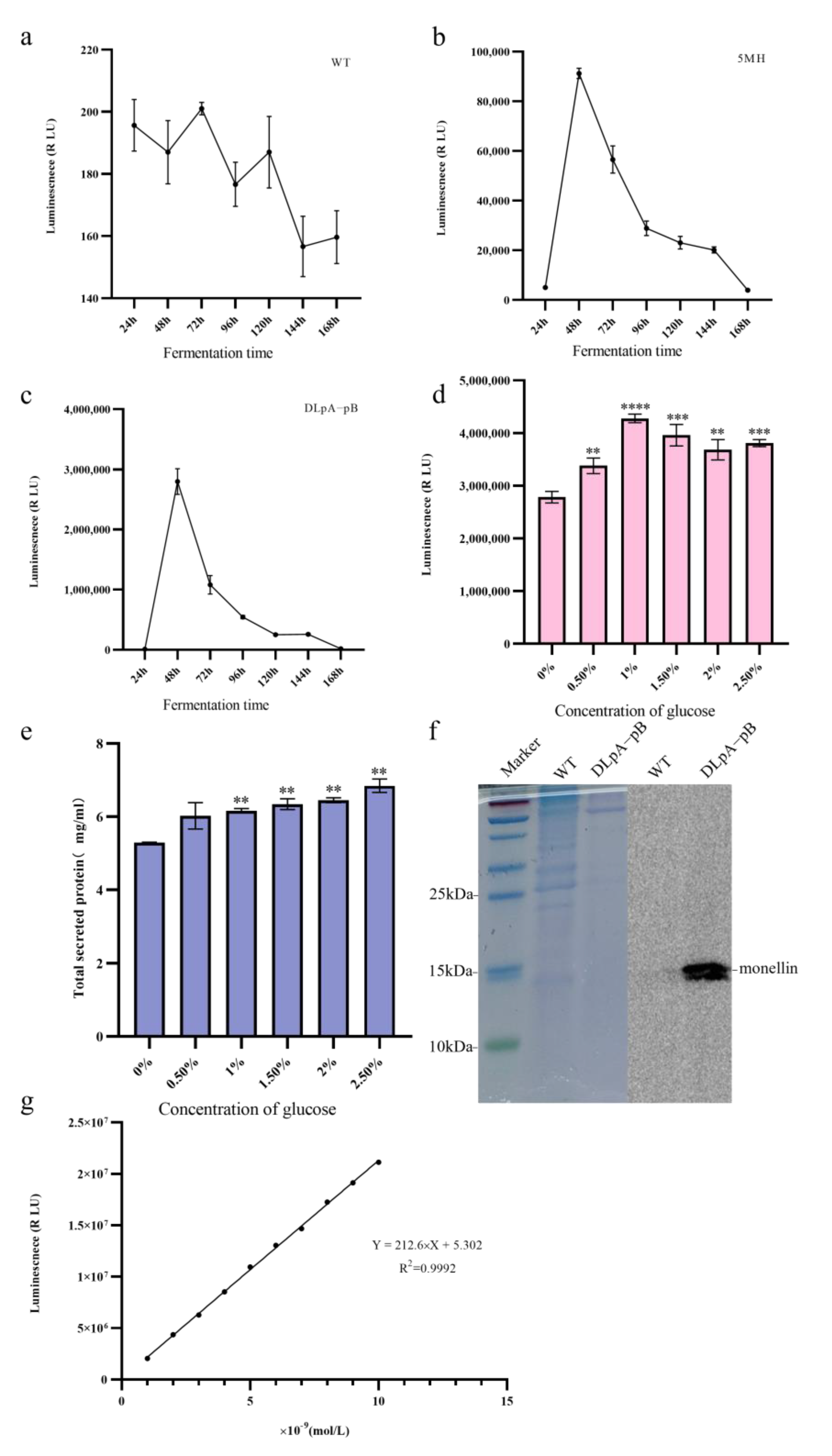
2.6. Quantification and Blotting of Monellin
3. Results
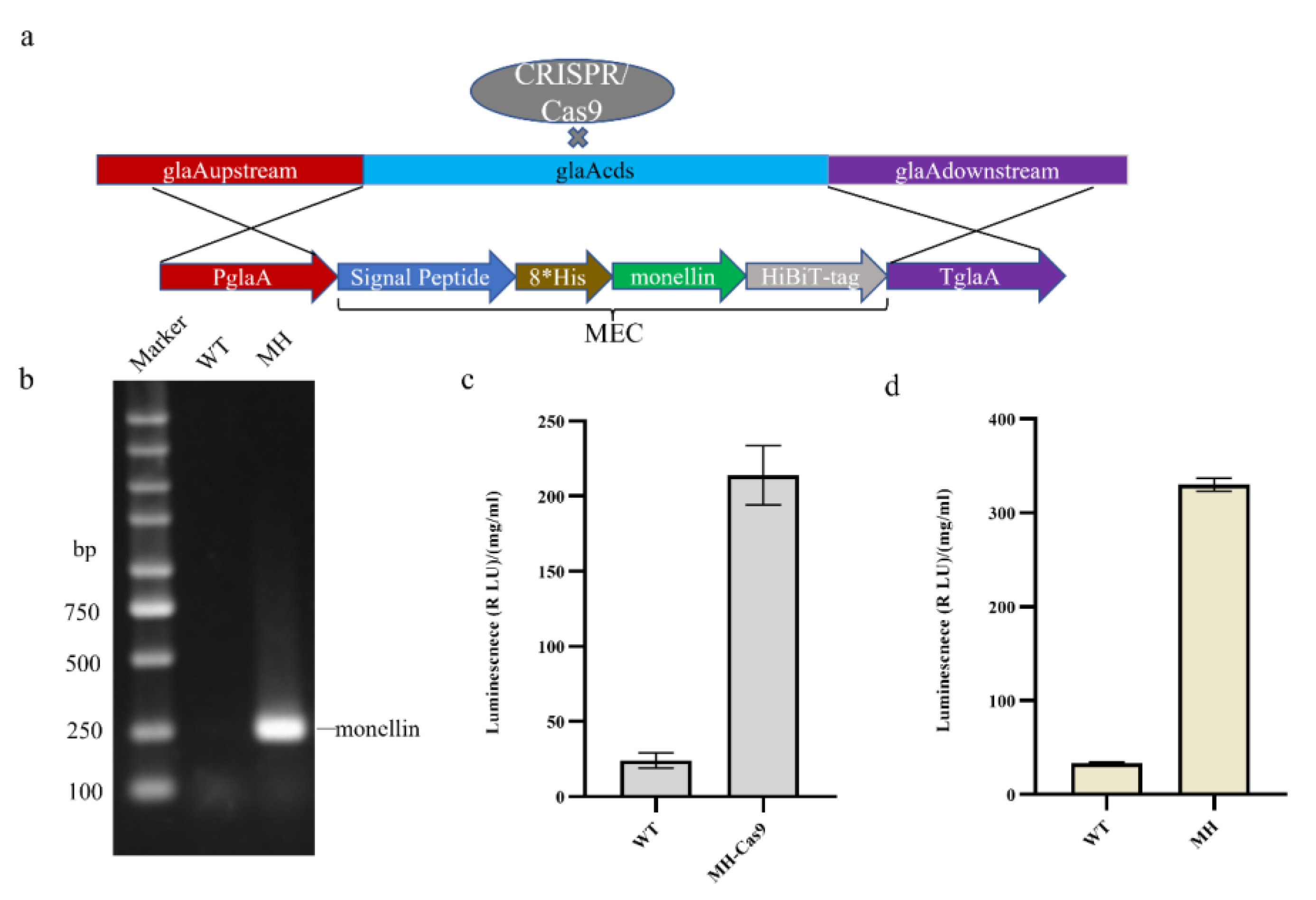
3.1. Establishment of the Monellin Expression Model
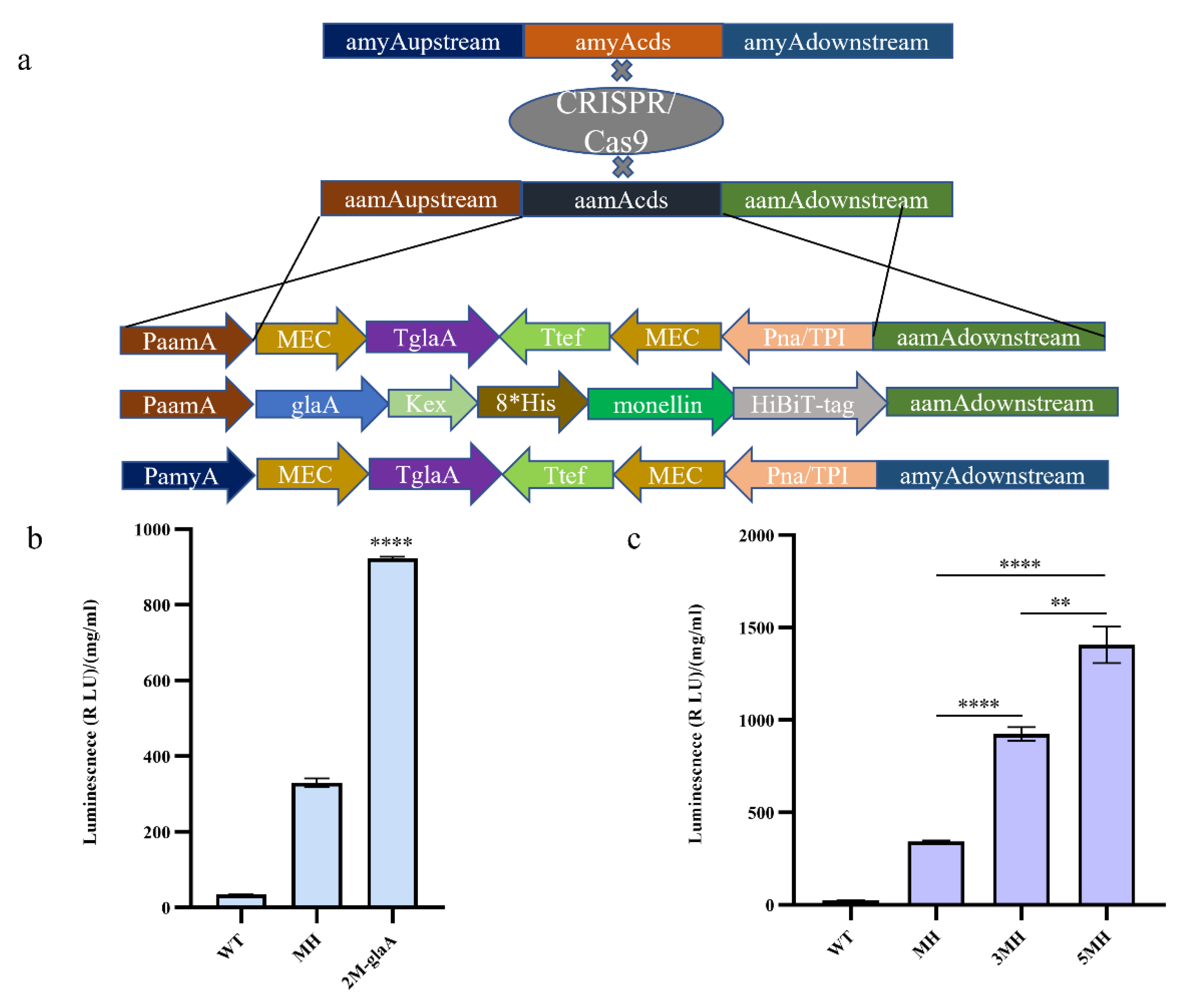
3.2. Increasing the Copy Number of Monellin to Enhance Its Expression
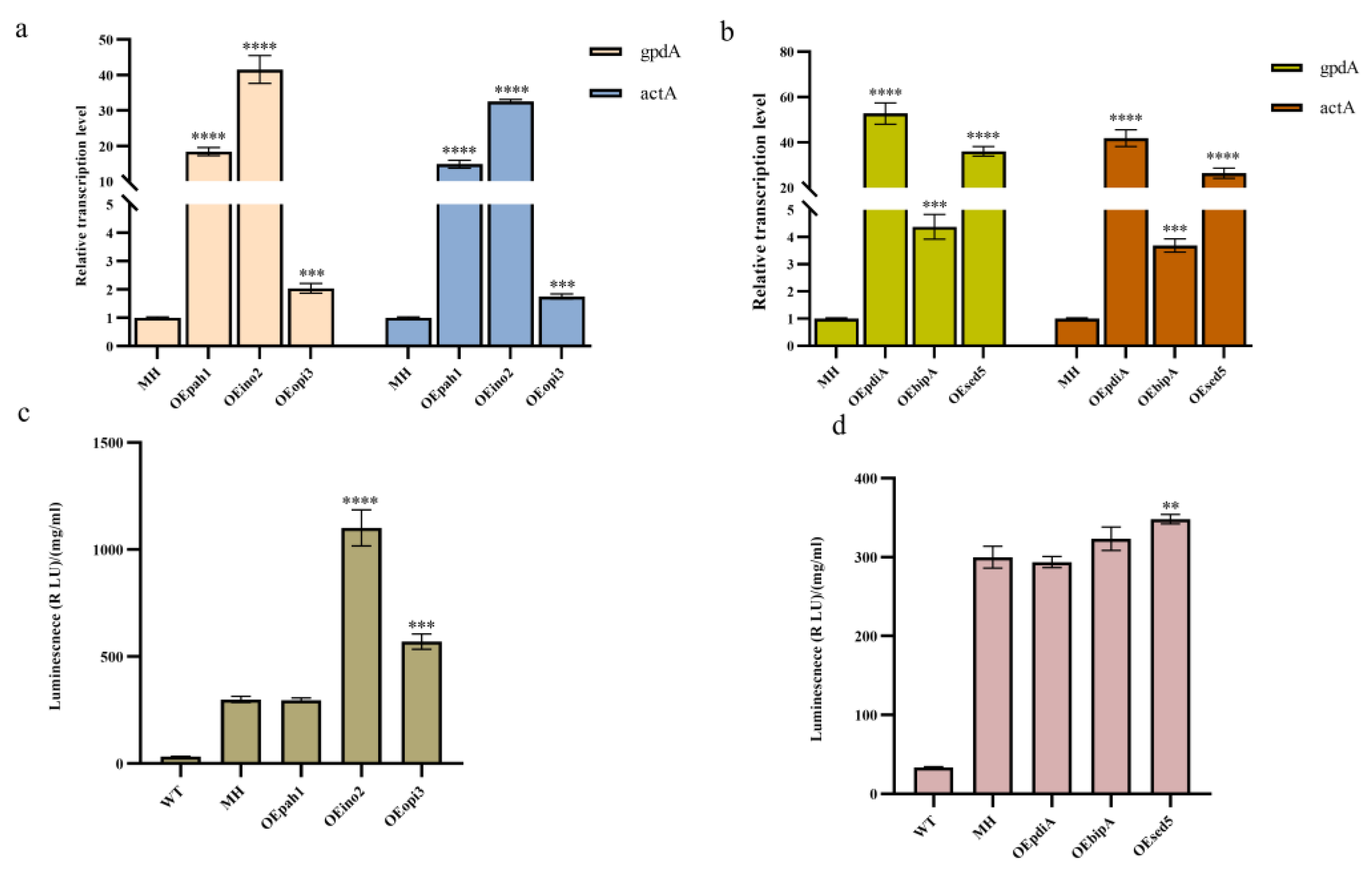
3.3. Effects of Overexpression of Key Genes in the Phospholipid Synthesis Pathway and the UPR Pathway on Monellin Expression
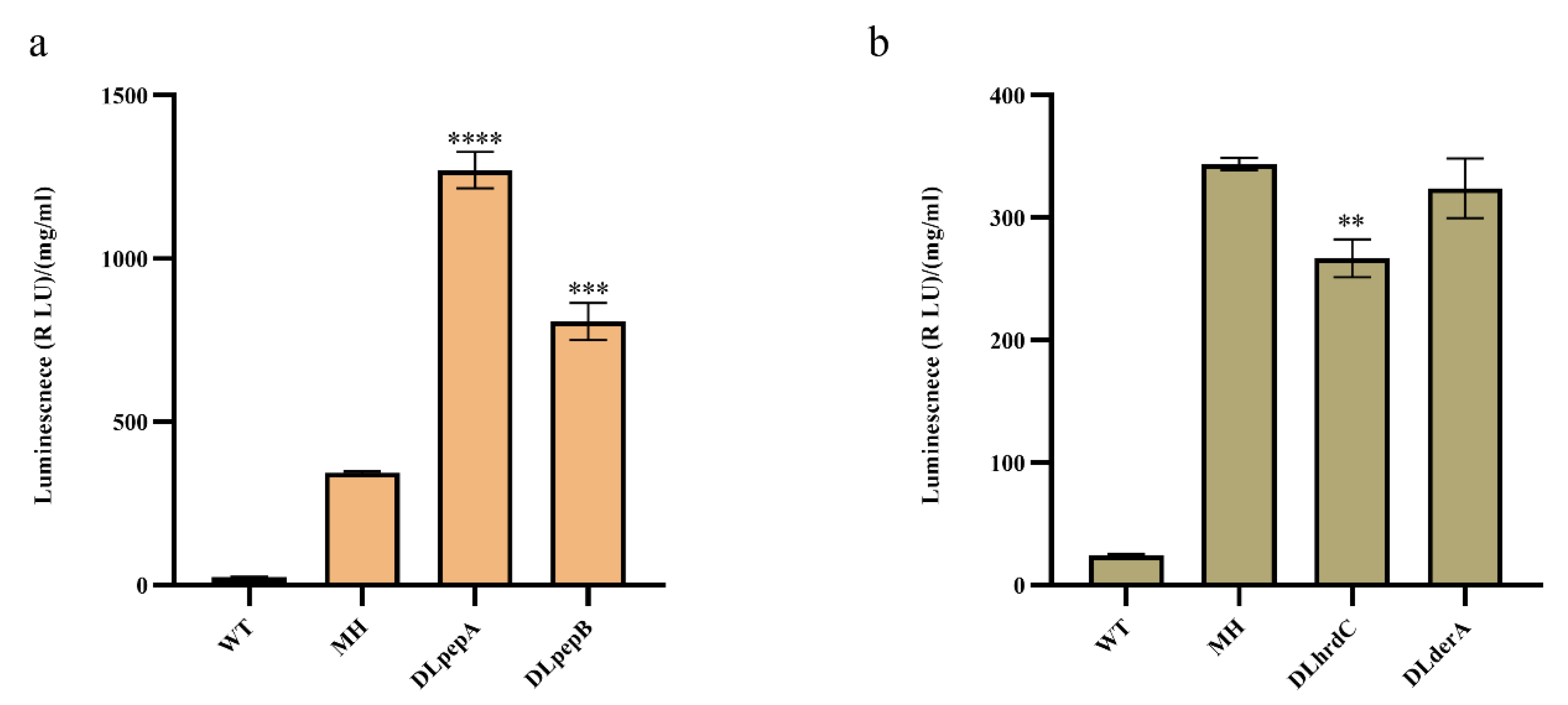
3.4. Effects of Knocking out Key Genes in the Extracellular Protease and ERAD Pathways on Monellin Expression
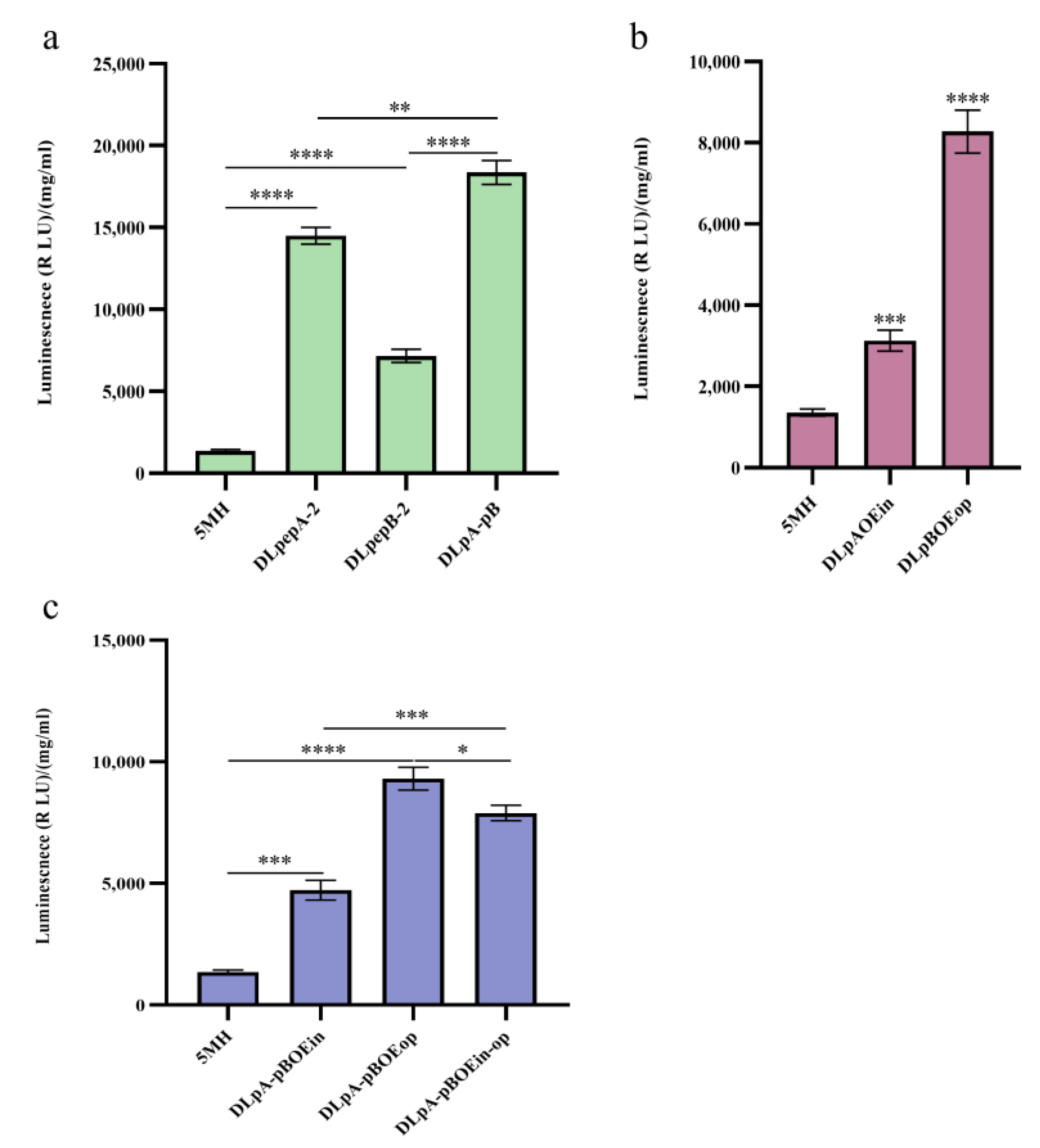
3.5. Effects of a Combination of Knockout Protease Genes and Overexpression of Key Genes for Phospholipid Synthesis on Monellin Expression
3.6. Optimization of Fermentation Medium and Monellin–HiBiT-Tag Blotting
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Krijgsheld, P.; Bleichrodt, R.; Veluw, G.; Wang, F.; Müller, W.; Dijksterhuis, J.; Wsten, H.A.B. Development in Aspergillus. Stud. Mycol. 2013, 74, 1–29. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Lu, J.; Zhang, G.; Liu, S.; Zhou, J.; Du, G.; Chen, J.J. Recent advances in the development of Aspergillus for protein production. Bioresour. Technol. 2022, 348, 126768. [Google Scholar] [CrossRef]
- Abarca, M.L.; Accensi, F.; Cano, J.; Cabañes, F. Taxonomy and significance of black aspergilli.” Antonie Van Leeuwenhoek 86. Antonie Leeuwenhoek Int. J. Gen. Mol. Microbiol. 2004, 86, 33–49. [Google Scholar] [CrossRef]
- Li, C.; Zhou, J.; Du, G.; Chen, J.; Takahashi, S.; Liu, S. Developing Aspergillus niger as a cell factory for food enzyme production. Biotechnol. Adv. 2020, 44, 107630. [Google Scholar] [CrossRef] [PubMed]
- Sakekar, A.A.; Gaikwad, S.R.; Punekar, N.S. Protein expression and secretion by filamentous fungi. J. Biosci. 2021, 46, 5. [Google Scholar] [CrossRef] [PubMed]
- Ward, O.P. Production of recombinant proteins by filamentous fungi. (Special Issue: Production of recombinant proteins.). Biotechnol. Adv. Int. Rev. J. 2012, 30, 1119–1139. [Google Scholar] [CrossRef]
- Fleissner, A.; Dersch, P. Expression and export: Recombinant protein production systems for Aspergillus. Appl. Microbiol. Biotechnol. 2010, 87, 1255–1270. [Google Scholar] [CrossRef]
- Peberdy, J.F. Protein secretion in filamentous fungi—Trying to understand a highly productive black box. Trends Biotechnol. 1994, 12, 50–57. [Google Scholar] [CrossRef] [PubMed]
- Kluge, J.; Terfehr, D.; Kuck, U. Inducible promoters and functional genomic approaches for the genetic engineering of filamentous fungi. Appl. Microbiol. Biotechnol. 2018, 102, 6357–6372. [Google Scholar] [CrossRef]
- Liu, F.L.; Wang, B.; Ye, Y.R.; Pan, L. High level expression and characterization of tannase tan? Using Aspergillus niger SH-2 with low-background endogenous secretory proteins as the host. Protein Expr. Purif. 2018, 144, 71–75. [Google Scholar] [CrossRef]
- Prathumpai, W.; Flitter, S.J.; McIntyre, M.; Nielsen, J. Lipase production by recombinant strains of Aspergillus niger expressing a lipase-encoding gene from Thermomyces lanuginosus. Appl. Microbiol. Biotechnol. 2004, 65, 714–719. [Google Scholar] [CrossRef] [PubMed]
- vanKuyk, P.A.; Benen, J.A.E.; Wosten, H.A.B.; Visser, J.; de Vries, R.P. A broader role for AmyR in Aspergillus niger: Regulation of the utilisation of d-glucose or d-galactose containing oligo- and polysaccharides. Appl. Microbiol. Biotechnol. 2012, 93, 285–293. [Google Scholar] [CrossRef]
- Dong, H.Z.; Zheng, J.W.; Yu, D.; Wang, B.; Pan, L. Efficient genome editing in Aspergillus niger with an improved recyclable CRISPR-HDR toolbox and its application in introducing multiple copies of heterologous genes. J. Microbiol. Methods 2019, 163, 105655. [Google Scholar] [CrossRef]
- Dong, L.B.; Yu, D.; Lin, X.T.; Wang, B.; Pan, L. Improving expression of thermostable trehalase from Myceliophthora sepedonium in Aspergillus niger mediated by the CRISPR/Cas9 tool and its purification, characterization. Protein Expr. Purif. 2020, 165, 105482. [Google Scholar] [CrossRef] [PubMed]
- Faus, I.; del Moral, C.; Adroer, N.; del Rio, J.L.; Patino, C.; Sisniega, H.; Casas, C.; Blade, J.; Rubio, V. Secretion of the sweet-tasting protein thaumatin by recombinant strains of Aspergillus niger var. awamori. Appl. Microbiol. Biotechnol. 1998, 49, 393–398. [Google Scholar] [CrossRef] [PubMed]
- Moralejo, F.J.; Cardoza, R.E.; Gutierrez, S.; Martin, J.F. Thaumatin production in Aspergillus awamori by use of expression cassettes with strong fungal promoters and high gene dosage. Appl. Environ. Microbiol. 1999, 65, 1168–1174. [Google Scholar] [CrossRef] [PubMed]
- Lombrana, M.; Moralejo, F.J.; Pinto, R.; Martin, J.F. Modulation of Aspergillus awamori thaumatin secretion by modification of bipA gene expression. Appl. Environ. Microbiol. 2004, 70, 5145–5152. [Google Scholar] [CrossRef] [PubMed]
- Moralejo, F.J.; Watson, A.J.; Jeenes, D.J.; Archer, D.B.; Martin, J.F. A defined level of protein disulfide isomerase expression is required for optimal secretion of thaumatin by Aspergillus awamori. Mol. Genet. Genom. 2001, 266, 246–253. [Google Scholar] [CrossRef]
- Valkonen, M.; Ward, M.; Wang, H.M.; Penttila, M.; Saloheimo, M. Improvement of foreign-protein production in Aspergillus niger var. awamori by constitutive induction of the unfolded-protein response. Appl. Environ. Microbiol. 2003, 69, 6979–6986. [Google Scholar] [CrossRef]
- Carvalho, N.; Arentshorst, M.; Kooistra, R.; Stam, H.; Sagt, C.M.; van den Hondel, C.; Ram, A.F.J. Effects of a defective ERAD pathway on growth and heterologous protein production in Aspergillus niger. Appl. Microbiol. Biotechnol. 2011, 89, 357–373. [Google Scholar] [CrossRef]
- Conesa, A.; Jeenes, D.; Archer, D.B.; van den Hondel, C.; Punt, P.J. Calnexin overexpression increases manganese peroxidase production in Aspergillus niger. Appl. Environ. Microbiol. 2002, 68, 846–851. [Google Scholar] [CrossRef] [PubMed]
- Moralejo, F.J.; Cardoza, R.E.; Gutierrez, S.; Lombrana, M.; Fierro, F.; Martin, J.F. Silencing of the aspergillopepsin B (pepB) gene of Aspergillus awamori by antisense RNA expression or protease removal by gene disruption results in a large increase in thaumatin production. Appl. Environ. Microbiol. 2002, 68, 3550–3559. [Google Scholar] [CrossRef]
- Moralejo, F.J.; Cardoza, R.E.; Gutierrez, S.; Sisniega, H.; Faus, I.; Martin, J.F. Overexpression and lack of degradation of thaumatin in an aspergillopepsin A-defective mutant of Aspergillus awamori containing an insertion in the pepA gene. Appl. Microbiol. Biotechnol. 2000, 54, 772–777. [Google Scholar] [CrossRef]
- Gramazio, S.; Trauth, J.; Bezold, F.; Essen, L.O.; Taxis, C.; Spadaccini, R. Light-induced fermenter production of derivatives of the sweet protein monellin is maximized in prestationary Saccharomyces cerevisiae cultures. Biotechnol. J. 2022, 17, 10. [Google Scholar] [CrossRef]
- Morris, J.A.; Martenson, R.; Deibler, G.; Cagan, R.H. Characterization of monellin, a protein that tastes sweet. J. Biol. Chem. 1973, 248, 534–539. [Google Scholar] [CrossRef]
- Picone, D.; Temussi, P.A. Dissimilar sweet proteins from plants: Oddities or normal components? Plant Sci. 2012, 195, 135–142. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.H.; Kang, C.H.; Kim, R.; Cho, J.M.; Lee, Y.B.; Lee, T.K. Redesigning a sweet protein: Increased stability and renaturability. Protein Eng. 1989, 2, 571–575. [Google Scholar] [CrossRef] [PubMed]
- Schwinn, M.K.; Machleidt, T.; Zimmerman, K.; Eggers, C.T.; Dixon, A.S.; Hurst, R.; Hall, M.P.; Encell, L.P.; Binkowski, B.F.; Wood, K.V. CRISPR-Mediated Tagging of Endogenous Proteins with a Luminescent Peptide. Acs Chem. Biol. 2018, 13, 467–474. [Google Scholar] [CrossRef]
- Yu, L.Y.; Li, L.X.; Yao, L.L.; Zheng, J.W.; Wang, B.; Pan, L. A Special Phenotype of Aconidial Aspergillus niger SH2 and Its Mechanism of Formation via CRISPRi. J. Fungi 2022, 8, 679. [Google Scholar] [CrossRef]
- Yin, C.; Wang, B.; He, P.; Lin, Y.; Pan, L. Genomic analysis of the aconidial and high-performance protein producer, industrially relevant Aspergillus niger SH2 strain. Gene 2014, 541, 107–114. [Google Scholar] [CrossRef]
- Li, L.X.; Yu, L.Y.; Wang, B.; Pan, L. Impact of overexpressing NADH kinase on glucoamylase production in Aspergillus niger. J. Ind. Microbiol. Biotechnol. 2022, 49. [Google Scholar] [CrossRef]
- Huang, L.G.; Li, X.J.; Dong, L.B.; Wang, B.; Pan, L. Profiling of chromatin accessibility identifies transcription factor binding sites across the genome of Aspergillus species. BMC Biol. 2021, 19. [Google Scholar] [CrossRef]
- Chumnanpuen, P.; Nookaew, I.; Nielsen, J. Integrated analysis, transcriptome-lipidome, reveals the effects of INO-level (INO2 and INO4) on lipid metabolism in yeast. BMC Syst. Biol. 2013, 7. [Google Scholar] [CrossRef]
- Dong, H.Z.; Yu, D.; Wang, B.; Pan, L. Identification and Characterization of a Novel Basic Helix-Loop-Helix Transcription Factor of Phospholipid Synthesis Regulation in Aspergillus niger. Front. Microbiol. 2020, 10, 2985. [Google Scholar] [CrossRef]
- Dong, H.Z.; Wang, B.; Pan, L. Study on the interaction mechanism of phospholipid imbalance and endoplasmic reticulum protein secretion imbalance in Aspergillus niger. Biochim. Biophys. Acta-Biomembr. 2021, 1863, 183530. [Google Scholar] [CrossRef] [PubMed]
- Carman, G.M.; Han, G.S. Regulation of Phospholipid Synthesis in the Yeast Saccharomyces cerevisiae. Annu. Rev. Biochem. 2011, 80, 859–883. [Google Scholar] [CrossRef]
- Van Zyl, J.H.D.; Den Haan, R.; Van Zyl, W.H. Over-expression of native Saccharomyces cerevisiae exocytic SNARE genes increased heterologous cellulase secretion. Appl. Microbiol. Biotechnol. 2014, 98, 5567–5578. [Google Scholar] [CrossRef]
- Chaney, J.L.; Clark, P.L. Roles for Synonymous Codon Usage in Protein Biogenesis. Annu. Rev. Biophys. 2015, 44, 143–166. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Yu, J.W.; Li, Z.X.; Yin, W.B. Rational design for fungal laccase production in the model host Aspergillus nidulans. Sci. China-Life Sci. 2019, 62, 84–94. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.G.; Dong, L.B.; Wang, B.; Pan, L. The transcription factor PrtT and its target protease profiles in Aspergillus niger are negatively regulated by carbon sources. Biotechnol. Lett. 2020, 42, 613–624. [Google Scholar] [CrossRef]
- Kamaruddin, N.; Storms, R.; Mahadi, N.M.; Illias, R.M.; Abu Bakar, F.D.; Murad, A.M.A. Reduction of Extracellular Proteases Increased Activity and Stability of Heterologous Protein in Aspergillus niger. Arab. J. Sci. Eng. 2018, 43, 3327–3338. [Google Scholar] [CrossRef]
- Sharon, H.; Hagag, S.; Osherov, N. Transcription Factor PrtT Controls Expression of Multiple Secreted Proteases in the Human Pathogenic Mold Aspergillus fumigatus. Infect. Immun. 2009, 77, 4051–4060. [Google Scholar] [CrossRef]
- Shemesh, E.; Hanf, B.; Hagag, S.; Attias, S.; Shadkchan, Y.; Fichtman, B.; Harel, A.; Krüger, T.; Brakhage, A.A.; Kniemeyer, O.J.O. Phenotypic and Proteomic Analysis of the Aspergillus fumigatus ΔPrtT, ΔXprG and ΔXprG/ΔPrtT Protease-Deficient Mutants. Front. Microbiol. 2017, 8, 2490. [Google Scholar] [CrossRef] [PubMed]
- Mattern, I.E.; van Noort, J.M.; van den Berg, P.; Archer, D.B.; Roberts, I.N.; van den Hondel, C.A. Isolation and characterization of mutants of Aspergillus niger deficient in extracellular proteases. Mol. Gen. Genet. MGG 1992, 234, 332–336. [Google Scholar] [CrossRef]
- Heimel, K. Unfolded protein response in filamentous fungi-implications in biotechnology. Appl. Microbiol. Biotechnol. 2015, 99, 121–132. [Google Scholar] [CrossRef]
- Lubeck, M.; Lubeck, P.S. Fungal Cell Factories for Efficient and Sustainable Production of Proteins and Peptides. Microorganisms 2022, 10, 753. [Google Scholar] [CrossRef] [PubMed]
- Zhu, S.-Y.; Xu, Y.; Yu, X.-W. Improved Homologous Expression of the Acidic Lipase from Aspergillus niger. J. Microbiol. Biotechnol. 2020, 30, 196–205. [Google Scholar] [PubMed]
- Rojas-Sánchez, U.; López-Calleja, A.C.; Millán-Chiu, B.E.; Fernández, F.; Loske, A.M.; Gómez-Lim, M.A. Enhancing the yield of human erythropoietin in Aspergillus niger by introns and CRISPR-Cas9. Protein Expr. Purif. 2020, 168, 105570. [Google Scholar] [CrossRef]
- Travis, S.M.; Kokona, B.; Fairman, R.; Hughson, F.M. Roles of singleton tryptophan motifs in COPI coat stability and vesicle tethering. Proc. Natl. Acad. Sci. USA 2019, 116, 24031–24040. [Google Scholar] [CrossRef]
- Thak, E.J.; Yoo, S.J.; Moon, H.Y.; Kang, H.A. Yeast synthetic biology for designed cell factories producing secretory recombinant proteins. Fems Yeast Res. 2020, 20. [Google Scholar] [CrossRef]
- Tegelaar, M.; Aerts, D.; Teertstra, W.R.; Wösten, H.A. Spatial induction of genes encoding secreted proteins in micro-colonies of Aspergillus niger. Sci. Rep. 2020, 10, 1–10. [Google Scholar] [CrossRef]
- Wucherpfennig, T.; Lakowitz, A.; Krull, R. Comprehension of viscous morphology—Evaluation of fractal and conventional parameters for rheological characterization of Aspergillus niger culture broth. J. Biotechnol. 2013, 163, 124–132. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| A. niger Strain | Parental Strain | Description | Reference |
|---|---|---|---|
| WT | SH-2 | ∆kusA, ∆pyrG | [30] |
| MH | WT | one copy of monellin, ∆glaA | This study |
| DLpepA | MH | ∆pepA | This study |
| DLpepB | MH | ∆pepB | This study |
| DLderA | MH | ∆derA | This study |
| DLhrdC | MH | ∆hrdC | This study |
| OEpah1 | MH | oepah1 | This study |
| OEino2 | MH | oeino2 | This study |
| OEopi3 | MH | oeopi3 | This study |
| OEpdiA | MH | oepdiA | This study |
| OEbipA | MH | oebipA | This study |
| OEsed5 | MH | oesed5 | This study |
| 3MH | MH | Three copies of monellin, ∆amyA | This study |
| 2M-glaA | MH | Fusion of glaA and monellin, ∆amyA | This study |
| 5MH | 3MH | Five copies of monellin, ∆aamA | This study |
| DLpepA-2 | 5MH | ∆pepA | This study |
| DLpepB-2 | 5MH | ∆pepB | This study |
| DLpA-pB | 5MH | ∆pepA, ∆pepB | This study |
| DLpAOEin | 5MH | ∆pepA, oe ino2 | This study |
| DLpBOEop | 5MH | ∆pepB, oe opi3 | This study |
| DLpA-pBOEin | 5MH | ∆pepA, ∆pepB, oe ino2 | This study |
| DLpA-pBOEop | 5MH | ∆pepA, ∆pepB, oe opi3 | This study |
| DLpA-pBOEin-op | 5MH | ∆pepA, ∆pepB, oe ino2, oe opi3 | This study |
| A. niger Strain | Gene Expression Cassette | CRISPR/Cas9 Plasmids | Reference |
|---|---|---|---|
| pFC330, | [31] | ||
| pFC332-Dual loci | [13] | ||
| MH | pMD18T-PglaA-M-TglaA | pFC330-1(PAfU6-glaAsgRNA-TU6) | This study |
| DLpepA | pMD18T-UpepA-DpepA | pFC330-2(PAfU6-pepAsgRNA-TU6) | This study |
| DLpepB | pMD18T-UpepB-DpepB | pFC330-3(PAfU6-pepBsgRNA-TU6) | This study |
| DLderA | pMD18T-UderA-DderA | pFC330-4(PAfU6-derAsgRNA-TU6) | This study |
| DLhrdC | pMD18T-UhrdC-DhrdC | pFC330-5(PAfU6-hrdCsgRNA-TU6) | This study |
| OEpah1 | pMD18T-PaamA-pha1-TaamA | pFC330-6(PAfU6-aamAsgRNA-TU6) | This study |
| OEino2 | pMD18T-PaamA-ino2-TaamA | pFC330-6(PAfU6-aamAsgRNA-TU6) | This study |
| OEopi3 | pMD18T-PaamA-opi3-TaamA | pFC330-6(PAfU6-aamAsgRNA-TU6) | This study |
| OEpdiA | pMD18T-PaamA-pdiA-TaamA | pFC330-6(PAfU6-aamAsgRNA-TU6) | This study |
| OEbipA | pMD18T-PaamA-bipA-TaamA | pFC330-6(PAfU6-aamAsgRNA-TU6) | This study |
| OEsed5 | pMD18T-PaamA-sed5-TaamA | pFC330-6(PAfU6-aamAsgRNA-TU6) | This study |
| 3MH | pMD18T-Pna/TPI-M-TglaA-Ttef-M-Pna/TPI-DamyA | pFC330-7(PAfU6-amyAsgRNA-TU6) | This Study |
| 2M-glaA | pMD18T-PaamA-glaA-M-TaamA | pFC330-6(PAfU6-aamAsgRNA-TU6) | This study |
| 5MH | pMD18T-PaamA-M-TglaA-Ttef-M-Pna/TPI-TaamA | pFC330-6PAfU6-aamAsgRNA-TU6 | This study |
| DLpepA-2 | pMD18T-UpepA-DpepA | pFC330-2(PAfU6-pepAsgRNA-TU6) | This study |
| DLpepB-2 | pMD18T-UpepB-DpepB | pFC330-3(PAfU6-pepBsgRNA-TU6) | This study |
| DLpA-pB | pMD18T-UpepA-DpepA pMD18T-UpepB-DpepB | pFC330-8(PAoU6-pepBsgRNA-TU6-PAfU6-pepAsgRNA-TU6) | This study |
| DLpAOEin | pMD18T-UpepA-PaamA-ino2-TpepA | pFC330-2(PAfU6-pepAsgRNA-TU6) | This study |
| DLpBOEop | pMD18T-UpepB-PaamA-opi3-TpepB | pFC330-3(PAoU6-pepBsgRNA-TU6) | This study |
| DLpA-pBOEin | pMD18T-UpepA-PaamA-ino2-TpepA pMD18T-UpepB-DpepB | pFC330-8(PAoU6-pepBsgRNA-TU6-PAfU6-pepAsgRNA-TU6) | This study |
| DLpA-pBOEop | pMD18T-UpepA-DpepA pMD18T-UpepB-PaamA-opi3-TpepB | pFC330-8(PAoU6-pepBsgRNA-TU6-PAfU6-pepAsgRNA-TU6) | This study |
| DLpA-pBOEin-op | pMD18T-UpepA-PaamA-ino2-TpepA pMD18T-UpepB-PaamA-opi3-TpepB | pFC330-8(PAoU6-pepBsgRNA-TU6-PAfU6-pepAsgRNA-TU6) | This study |
| A. niger Strain | Parental Strain | Description | Compared with MH |
|---|---|---|---|
| MH | WT | one copy of monellin, ∆glaA | 1 |
| DLpepA | MH | ∆pepA | 3.7 times increase |
| DLpepB | MH | ∆pepB | 2.35 times increase |
| DLderA | MH | ∆derA | No impact |
| DLhrdC | MH | ∆hrdC | Decreased by 22.3% |
| OEpah1 | MH | oe pah1 | No impact |
| OEino2 | MH | oe ino2 | 3.67 times increase |
| OEopi3 | MH | oe opi3 | 1.90 times increase |
| OEpdiA | MH | oe pdiA | No impact |
| OEbipA | MH | oe bipA | No impact |
| OEsed5 | MH | oe sed5 | Increased by 22.3% |
| 3MH | MH | Three copies of monellin, ∆amyA | 2.70 times increase |
| 2M-glaA | MH | Fusion of glaA and monellin, ∆amyA | 2.80 times increase |
| 5MH | 3MH | Five copies of monellin, ∆aamA | 4.10 times increase |
| DLpepA-2 | 5MH | ∆pepA | 43.87 times increase |
| DLpepB-2 | 5MH | ∆pepB | 21.65 times increase |
| DLpA-pB | 5MH | ∆pepA, ∆pepB | 55.51 times increase |
| DLpAOEin | 5MH | ∆pepA, oe ino2 | 9.43 times increase |
| DLpBOEop | 5MH | ∆pepB, oe opi3 | 25.01 times increase |
| DLpA-pBOEin | 5MH | ∆pepA, ∆pepB, oe ino2 | 14.27 times increase |
| DLpA-pBOEop | 5MH | ∆pepA, ∆pepB, oe opi3 | 28.17 times increase |
| DLpA-pBOEin-op | 5MH | ∆pepA, ∆pepB, oe ino2, oe opi3 | 23.78 times increase |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, K.; Zheng, J.; Yu, L.; Wang, B.; Pan, L. Exploration of the Strategy for Improving the Expression of Heterologous Sweet Protein Monellin in Aspergillus niger. J. Fungi 2023, 9, 528. https://doi.org/10.3390/jof9050528
Li K, Zheng J, Yu L, Wang B, Pan L. Exploration of the Strategy for Improving the Expression of Heterologous Sweet Protein Monellin in Aspergillus niger. Journal of Fungi. 2023; 9(5):528. https://doi.org/10.3390/jof9050528
Chicago/Turabian StyleLi, Ke, Junwei Zheng, Leyi Yu, Bin Wang, and Li Pan. 2023. "Exploration of the Strategy for Improving the Expression of Heterologous Sweet Protein Monellin in Aspergillus niger" Journal of Fungi 9, no. 5: 528. https://doi.org/10.3390/jof9050528
APA StyleLi, K., Zheng, J., Yu, L., Wang, B., & Pan, L. (2023). Exploration of the Strategy for Improving the Expression of Heterologous Sweet Protein Monellin in Aspergillus niger. Journal of Fungi, 9(5), 528. https://doi.org/10.3390/jof9050528