
Temporal Patterns and Inter-Correlations among Physical and Antioxidant Attributes and Enzyme Activities of Apricot Fruit Inoculated with Monilinia laxa under Salicylic Acid and Methyl Jasmonate Treatments under Shelf-Life Conditions


 ,
, 
Abstract
1. Introduction
2. Materials and Methods
2.1. Isolation of M. laxa
2.2. Plant Materials and Experimental Setup for SA and MeJA Treatments
2.3. Brown Rot Measures: Brown Rot Incidence and Lesion Diameter
2.4. Physical Attributes: Fruit Firmness and Lignin Content
2.5. Antioxidant Properties: Total Soluble Phenol Content and Total Antioxidant Capacity
2.6. Activity of Defense-Related Enzymes
2.7. Statistical Analysis
2.7.1. ANOVA
2.7.2. Correlation and Regression Analysis among Measurements
2.7.3. Principal Component Analysis
3. Results
3.1. Experiment 1: Brown Rot Incidence and Lesion Diameter in Three Concentrations of MeJA and SA Treatments
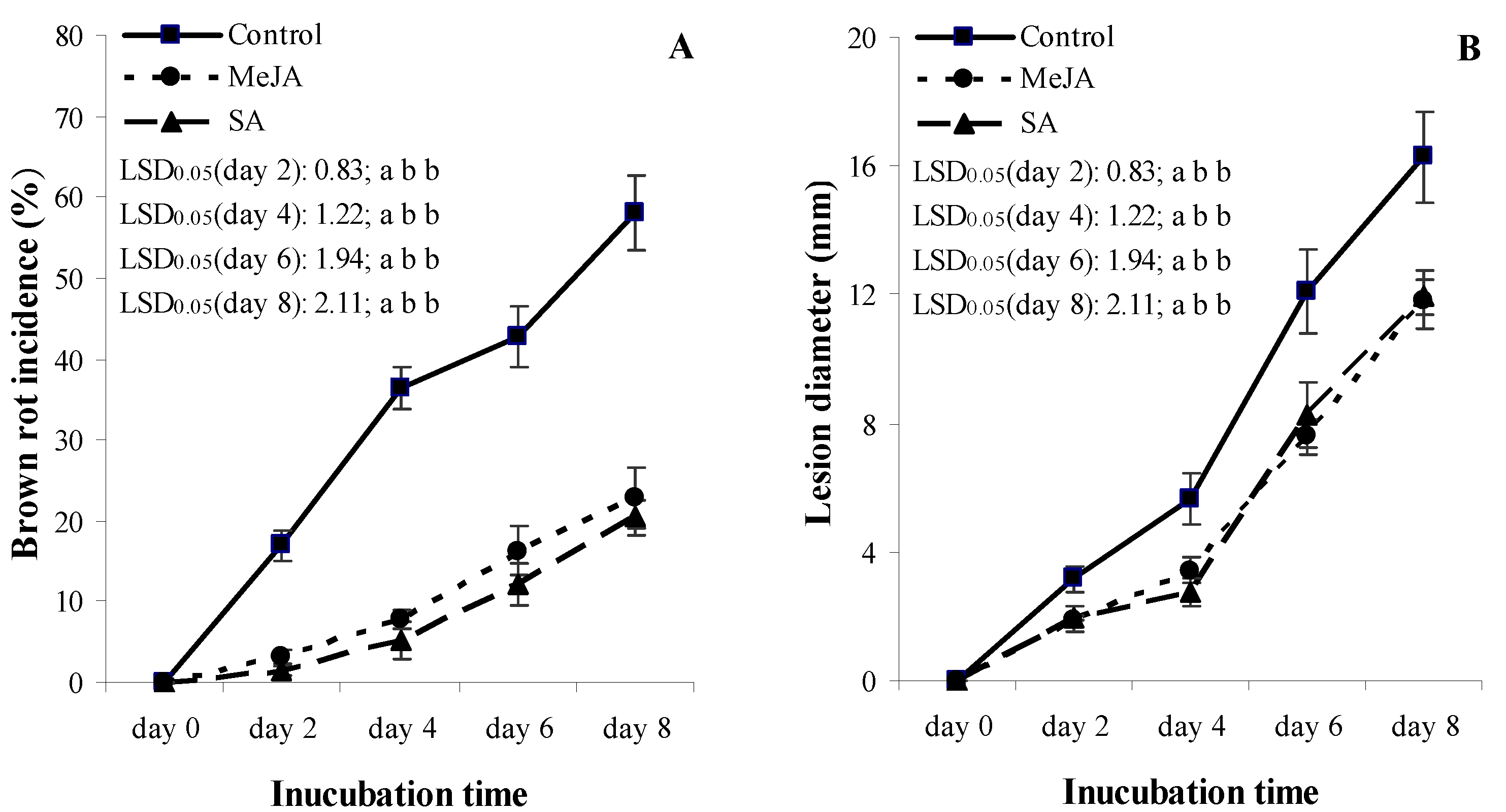
3.2. Experiment 2: Temporal Patterns of Brown Rot Measures
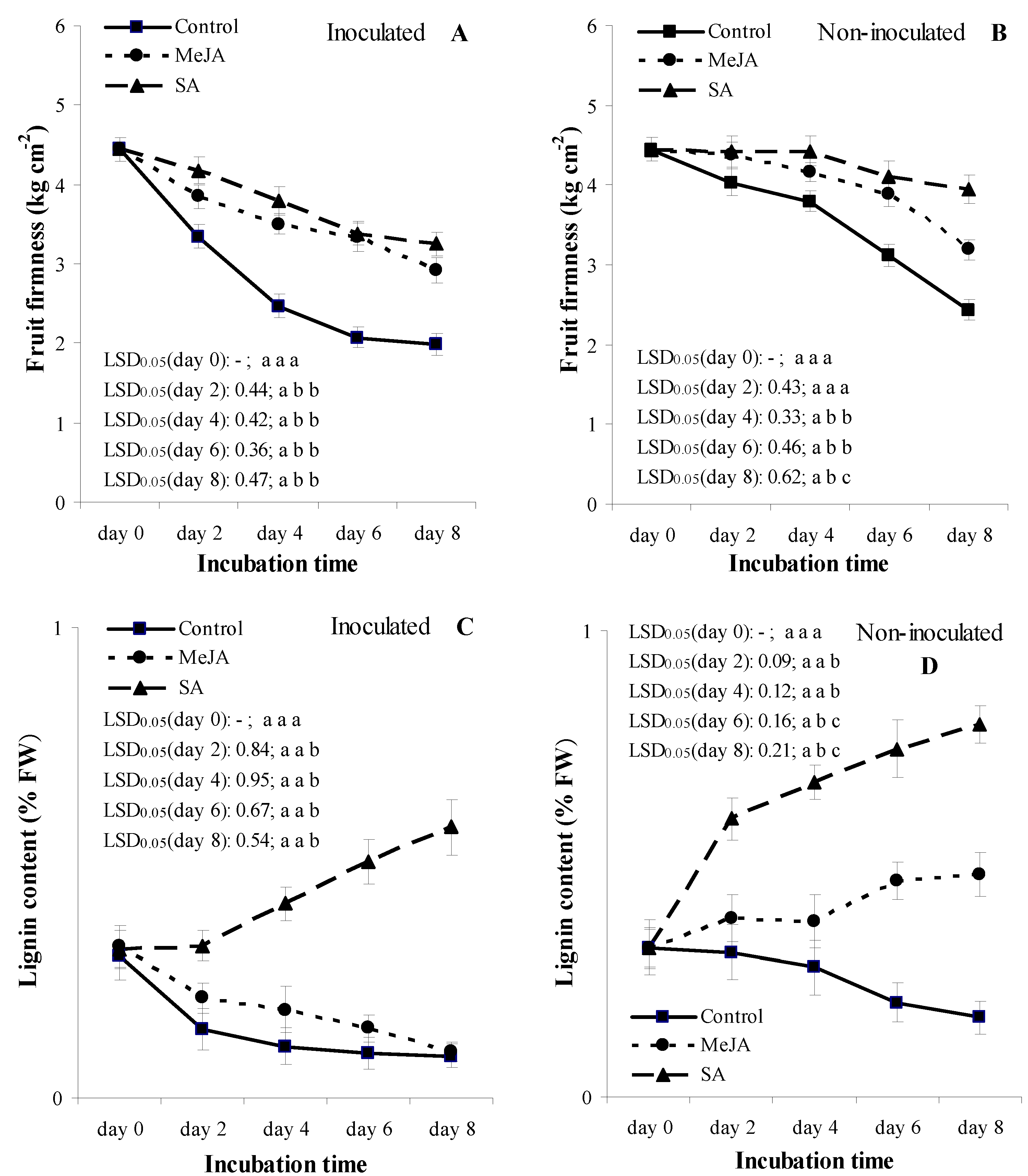
3.3. Experiment 2: Temporal Patterns of Fruit Firmness and Lignin Content
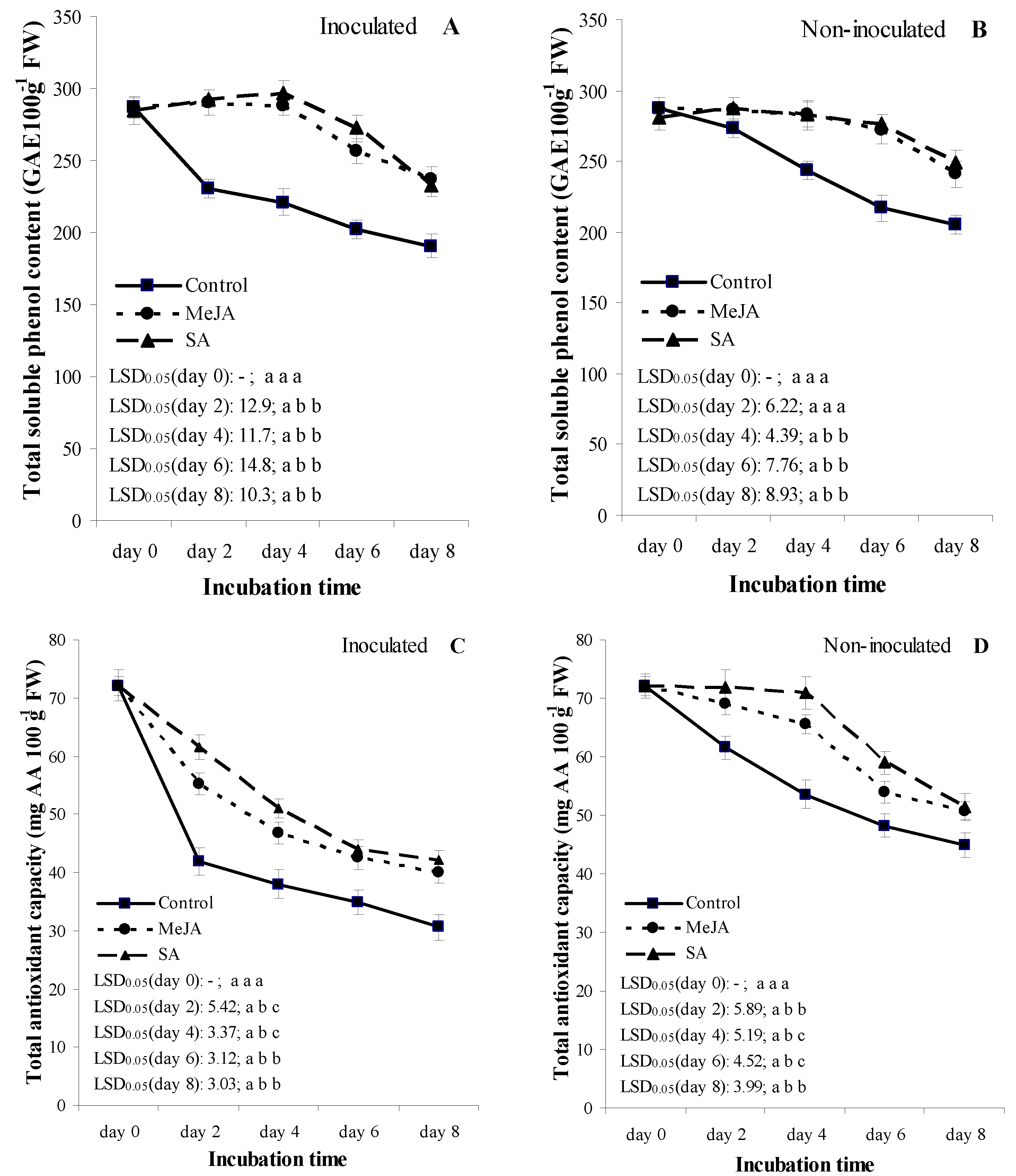
3.4. Experiment 2: Temporal Patterns of Total Soluble Phenol Content and Total Antioxidant Capacity
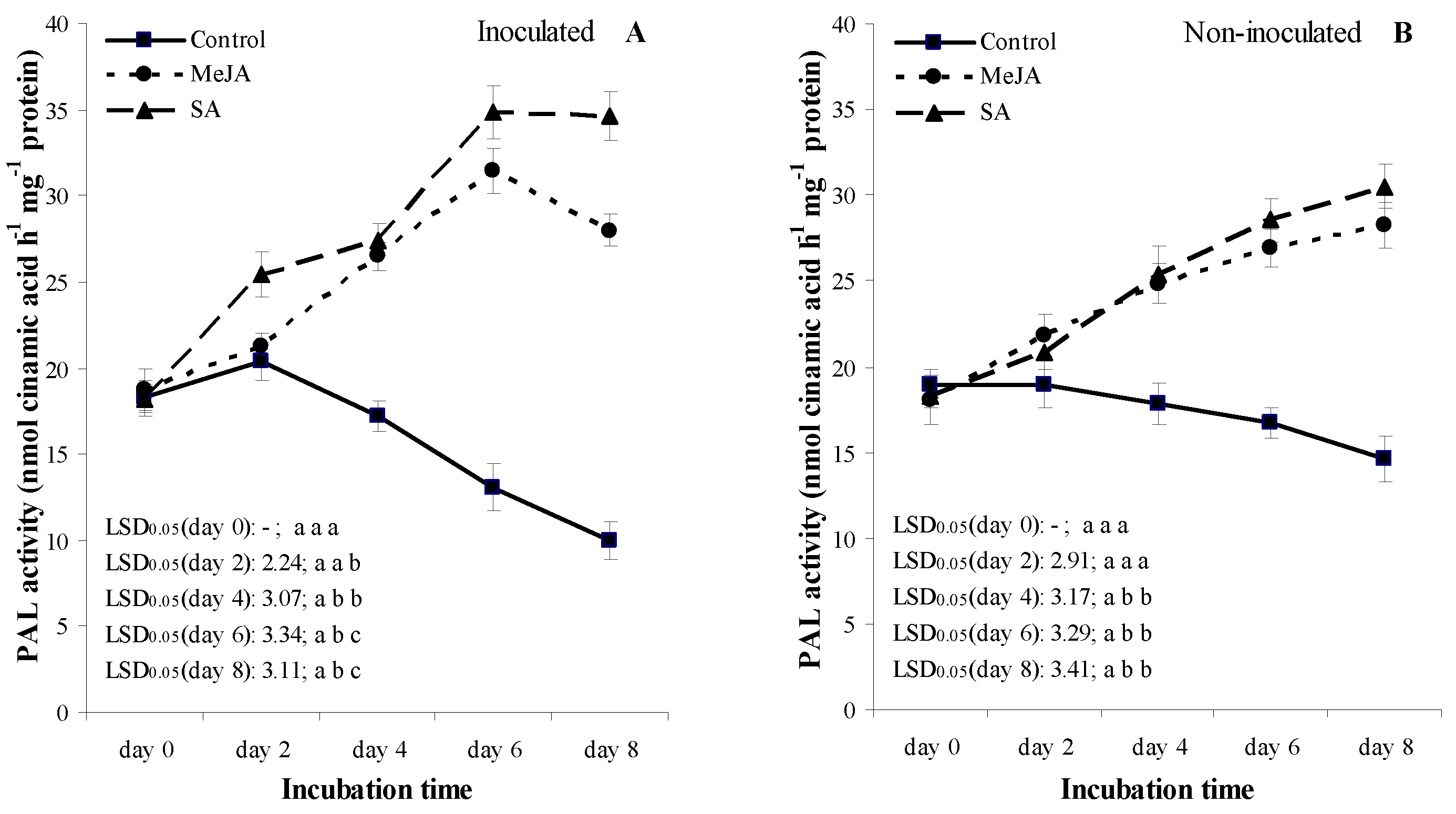
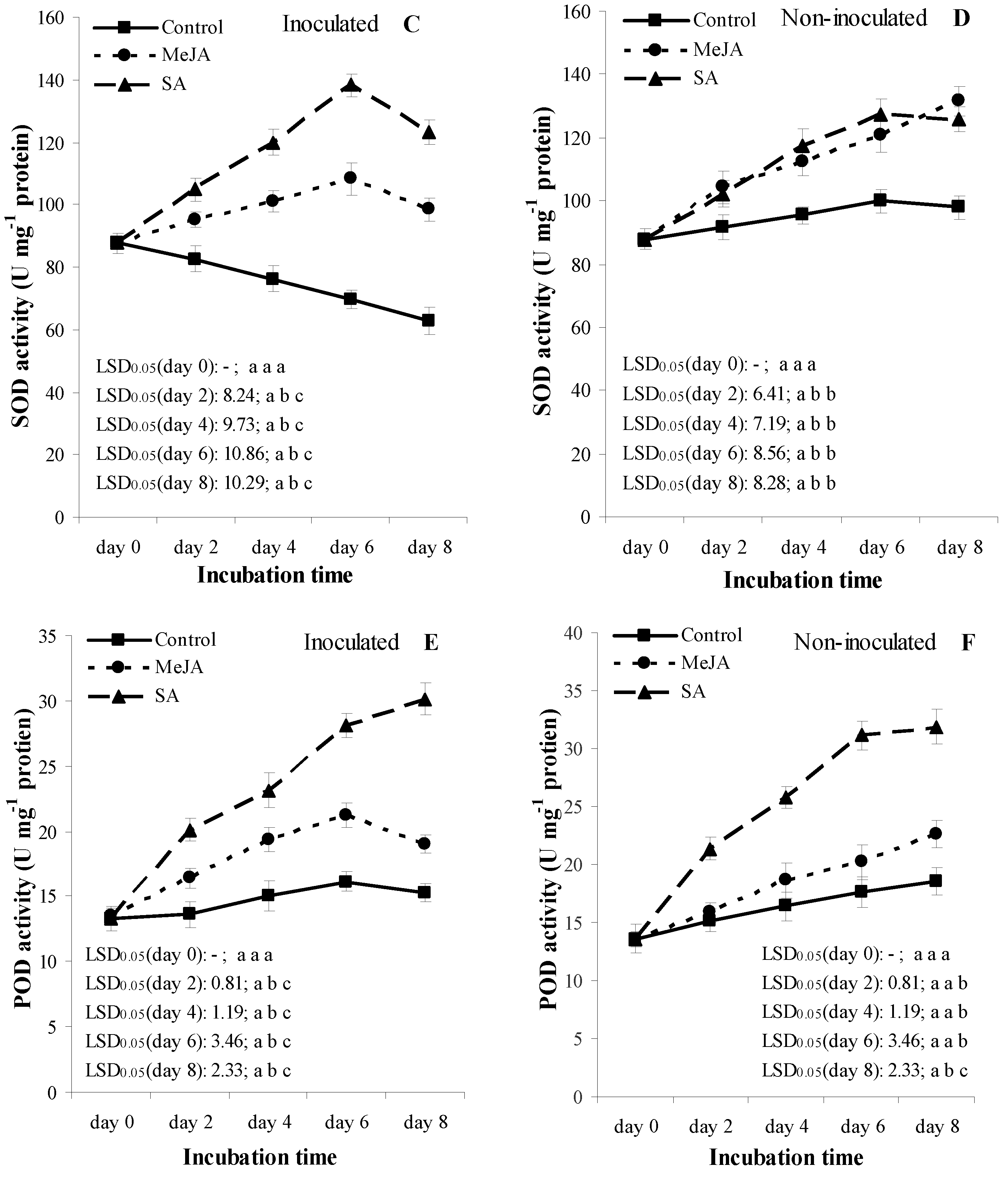
3.5. Experiment 2: Temporal Patterns of the Activity of Defense-Related Enzymes
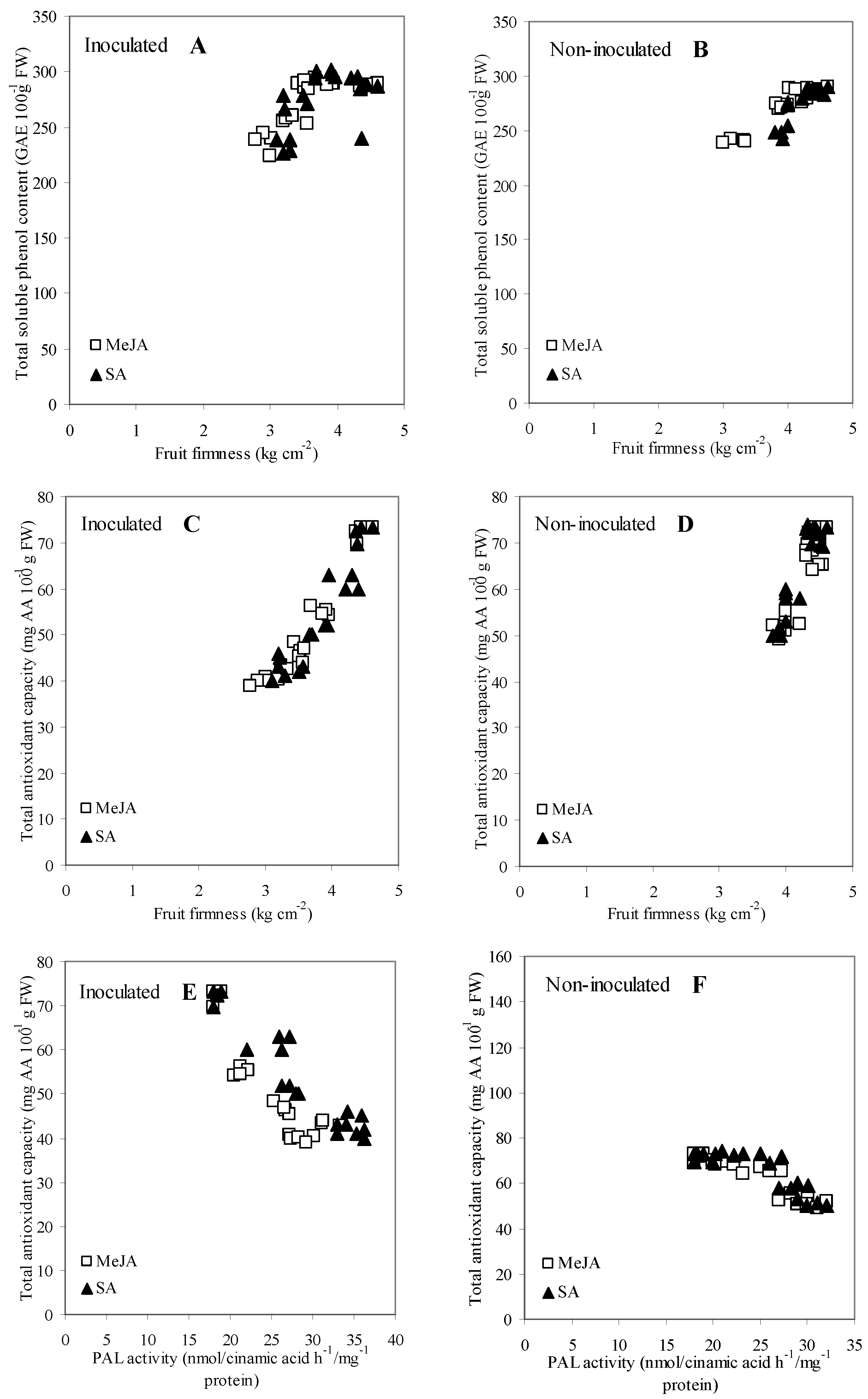
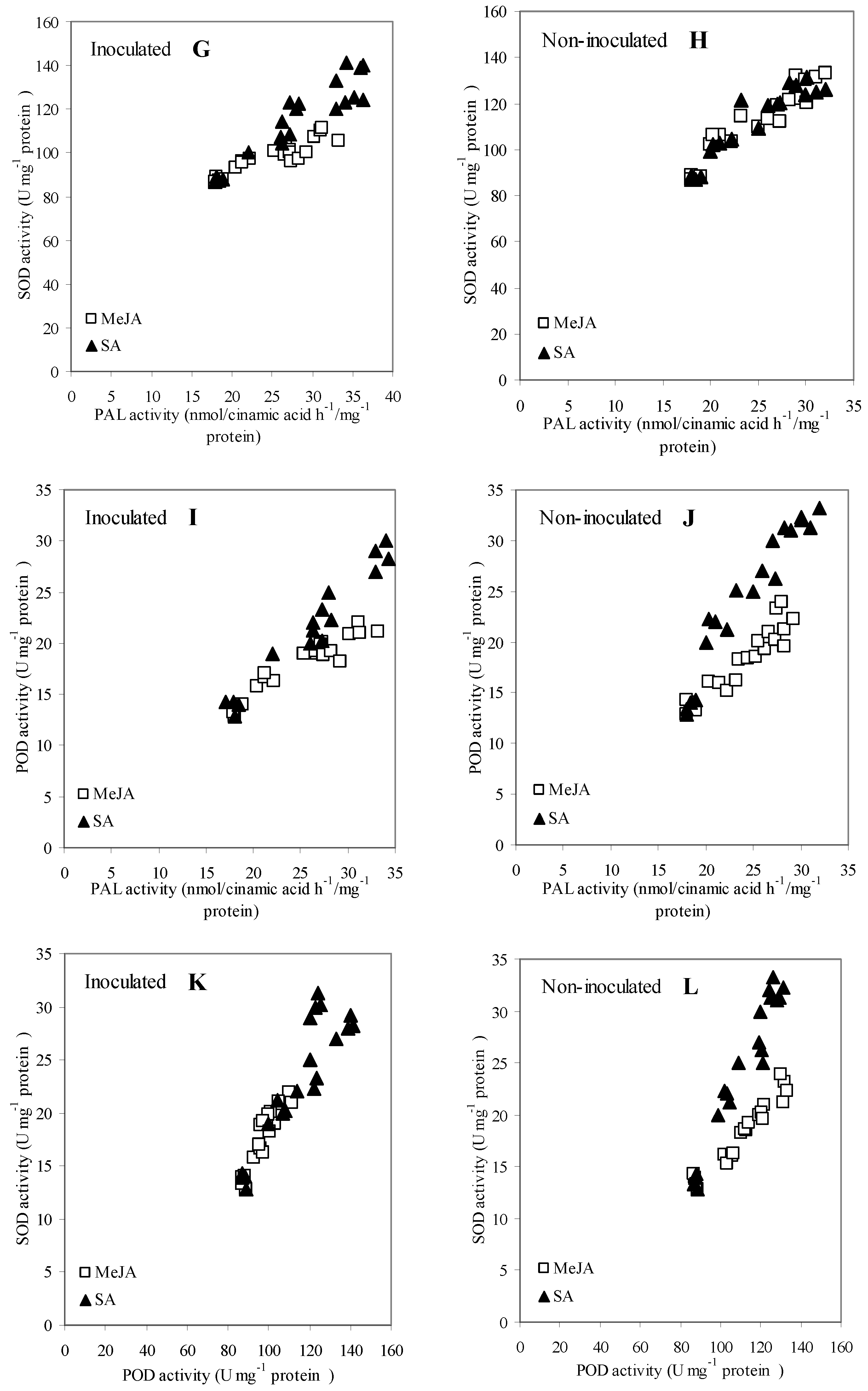
3.6. Correlation and Regression Analysis among Measurements
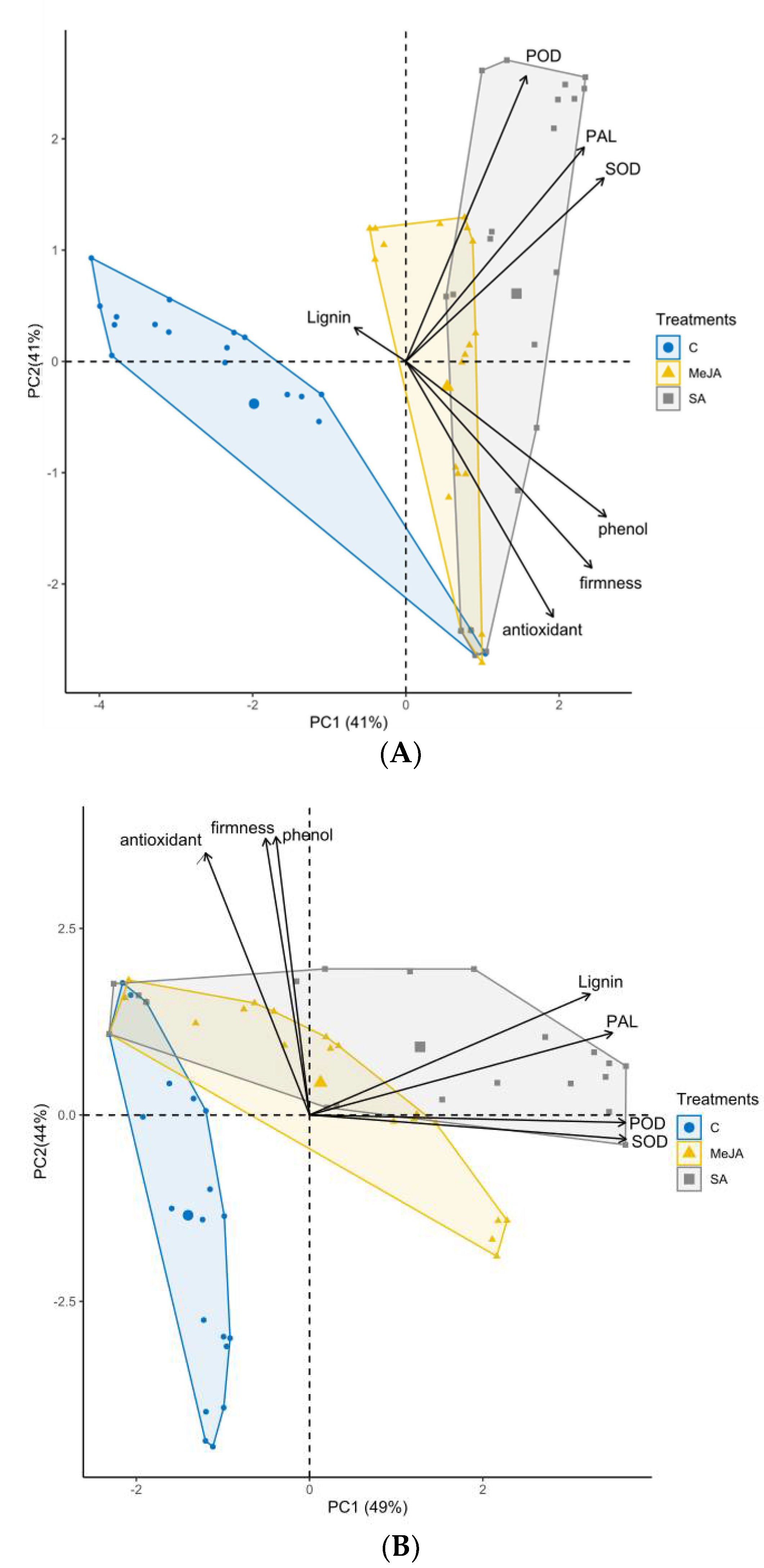
3.7. Principal Component Analysis among Measurements
4. Discussion
5. Conclusions
- i.
- Direct reducing effect of disease incidence (BRI) and lesion diameter (LD) on infected fruits;
- ii.
- Indirect reducing effect by increasing physical and antioxidant attributes (FF, LC, TAC and TSPC) of infected fruit, which resulted in an increase in fungal antagonistic compounds such as phenols and antioxidants in the infected fruits;
- iii.
- Indirect reducing effect by inducing defense mechanisms in the infected fruit cells, which resulted in an increase in production and activity of defense-related enzymes such as PAL, POD and SOD; thus, the lignification of the cell wall was increased, which made the cell wall more mechanically rigid against fungal attack.
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Wormald, H. The Brown Rot Disease of Fruit Trees; Technical Bulletin, no. 3; Ministry of Agriculture, Fisheries and Food: London, UK, 1954; pp. 1–337.
- Byrde, R.J.W.; Willetts, H.J. The Brown Rot Fungi of Fruit: Their Biology and Control; Pergamon Press: Oxford, UK, 1977; pp. 1–171. [Google Scholar]
- Rungjindamai, N.; Jeffries, P.; Xu, X.M. Epidemiology and management of brown rot on stone fruit caused by Monilinia laxa. Eur. J. Plant Pathol. 2014, 140, 1–17. [Google Scholar] [CrossRef]
- Holb, I. Brown rot: Causes, detection and control of Monilinia spp. affecting tree fruit. In Integrated Management of Diseases and Insect Pests of Tree Fruit; Fountain, X.M., Ed.; Burleigh Dodds Science Publishing: Cambridge, UK, 2019; pp. 103–150. [Google Scholar]
- Balsells-Llauradó, M.; Silva, C.J.; Usall, J.; Vall-Llaura, N.; Serrano-Prieto, S.; Teixidó, N.; Torres, R. Depicting the battle between nectarine and Monilinia laxa: The fruit developmental stage dictates the effectiveness of the host defenses and the pathogen’s infection strategies. Hortic. Res. 2020, 7, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Batra, L.R. World Species of Monilinia: Their Ecology, Biosystematics and Control; Cramer: Berlin, Germany, 1991; pp. 1–246. [Google Scholar]
- Holb, I.J. The brown rot fungi of fruit crops (Monilinia spp.). II. Important features of their epidemiology. Int. J. Hortic. Sci. 2004, 10, 17–35. [Google Scholar] [CrossRef]
- Everhart, S.E.; Askew, A.; Seymour, L.; Holb, I.J.; Scherm, H. Characterization of three-dimensional spatial aggregation and association patterns of brown rot symptoms within intensively mapped sour cherry trees. Ann. Bot. 2011, 108, 1195–1202. [Google Scholar] [CrossRef][Green Version]
- Holb, I.J.; Kunz, S. Integrated control of brown rot blossom blight by combining approved chemical control options with Aureobasidium pullulans in organic cherry production. Crop Prot. 2013, 54, 114–120. [Google Scholar] [CrossRef]
- Holb, I.J.; Szőke, S.; Abonyi, F. Temporal development and relationship amongst brown rot blossom blight, fruit blight and fruit rot in integrated and organic sour cherry orchards. Plant Pathol. 2013, 62, 799–808. [Google Scholar] [CrossRef]
- Larena, I.; Villarino, M.; Melgarejo, P.; Cal, A.D. Epidemiological studies of brown rot in Spanish cherry orchards in the Jerte Valley. J. Fungi 2021, 7, 203. [Google Scholar] [CrossRef] [PubMed]
- Villarino, M.; Melgarejo, P.; Usall, J.; Segarra, J.; Lamarca, N.; de Cal, A. Secondary inoculum dynamics of Monilinia spp. and relationship to the incidence of postharvest brown rot in peaches and the weather conditions during the growing season. Eur. J. Plant Pathol. 2012, 133, 585–598. [Google Scholar] [CrossRef]
- Garcia-Benitez, C.; Melgarejo, P.; de Cal, A. Fruit maturity and post-harvest environmental conditions influence the pre-penetration stages of Monilinia infections in peaches. Int. J. Food Microbiol. 2017, 241, 117–122. [Google Scholar] [CrossRef]
- Garcia-Benitez, C.; Casals, C.; Usall, J.; Sánchez-Ramos, I.; Melgarejo, P.; de Cal, A. Impact of postharvest handling on preharvest latent infections caused by Monilinia spp. in nectarines. J. Fungi 2020, 6, 266. [Google Scholar] [CrossRef]
- Smilanick, J.L.; Margosan, D.A.; Henson, D.J. Evaluation of heated solutions of sulfur dioxide, ethanol, and hydrogen peroxide to control postharvest green mold of lemons. Plant Dis. 1995, 79, 742–747. [Google Scholar] [CrossRef]
- Tian, S.; Torres, R.; Ballester, A.R.; Li, B.; Vilanova, L.; González-Candelas, L. Molecular aspects in pathogen-fruit interactions: Virulence and resistance. Postharvest Biol. Technol. 2016, 122, 11–21. [Google Scholar] [CrossRef]
- Bernat, M.; Casals, C.; Teixidó, N.; Torres, R.; Carballo, B.C.; Usall, J. Efficacy of environmental friendly disinfectants against the major postharvest pathogens of stone fruits on plastic and wood surfaces. Food Sci. Technol. Int. 2019, 25, 109–119. [Google Scholar] [CrossRef] [PubMed]
- Droby, S.; Wisniewski, M.; Teixidó, N.; Spadaro, D.; Jijakli, M.H. Biocontrol of postharvest diseases with antagonistic microorganisms. In Postharvest Pathology of Fresh Horticultural Produce; Paulo, L., Smilanick, J.L., Eds.; CRC Press: Boca Raton, FL, USA, 2019; pp. 463–498. [Google Scholar]
- Torres, R.; Vilanova, L.; Usall, J.; Teixidó, N. Insights into Fruit Defense Mechanisms Against the Main Postharvest Pathogens of Apples and Oranges. In Postharvest Pathology. Plant Pathology in the 21st Century; Spadaro, D., Droby, S., Gullino, M.L., Eds.; Springer: Cham, Switzerland, 2021; Volume 11, pp. 21–40. [Google Scholar]
- Kessmann, H.; Staub, T.; Hofmann, C.; Maetzke, T.; Herzog, J. Induction of systemic acquired disease resistance in plants by chemicals. Annu. Rev. Phytopathol. 1994, 32, 439–459. [Google Scholar] [CrossRef] [PubMed]
- Lawton, K.; Friedrich, L.; Hunt, M.; Weymann, K.; Kessmann, H.; Staub, T.; Ryals, J. Benzothiadiazole induces disease resistance in Arabidopsis by activation of the systemic acquired resistance signal transduction pathway. Plant J. 1996, 10, 71–82. [Google Scholar] [CrossRef] [PubMed]
- Schroder, M.; Hahlbrock, K.; Kombrink, E. Temporal and spatial patterns of β 1,3-glucanase and chitinase induction in potato leaves infected by Phytophthora infestans. Plant J. 1992, 2, 161–172. [Google Scholar] [CrossRef]
- Droby, S.; Vinokur, V.; Weiss, B.; Cohen, L.; Daus, A.; Goldschmidt, E.E.; Porat, R. Induction of resistance to Penicillium digitatum in grapefruit by the yeast biocontrol agent Candida oleophila. Phytopathology 2002, 92, 393–399. [Google Scholar] [CrossRef]
- Qin, G.Z.; Tian, S.P.; Xu, Y.; Wan, Y.K. Enhancement of biocontrol efficacy of antagonistic yeasts by salicylic acid in sweet cherry fruit. Physiol. Mol. Plant Pathol. 2003, 62, 147–154. [Google Scholar] [CrossRef]
- Durrant, W.E.; Dong, X. Systemic acquired resistance. Annu. Rev. Phytopathol. 2004, 42, 185–209. [Google Scholar] [CrossRef]
- Luna, E.; Bruce, T.J.; Roberts, M.R.; Flors, V.; Ton, J. Next-generation systemic acquired resistance. Plant Physiol. 2012, 158, 844–853. [Google Scholar] [CrossRef]
- Blanco, P.R.; Medina, E.N.; Lopez, R.J.A.; Gonzalez, R.J.A.; Villalba, J.M.; Moyano, E.; Bokshi, A.I.; Morris, S.C.; Deverall, B.J. Effects of benzothiadiazole and acetylsalicylic acid on β-1,3-glucanase activity and disease resistance in potato. Plant Pathol. 2003, 52, 22–27. [Google Scholar]
- Huang, Y.; Deverall, B.J.; Tang, W.H.; Wang, W.; Wu, F.W. Foliar application of acibenzolar-S-methyl and protection of postharvest rock melons and Hami melons from disease. Eur. J. Plant Pathol. 2000, 106, 651–656. [Google Scholar] [CrossRef]
- Liu, X.; Jiang, W.B.; Bi, Y.; Luo, Y.B. Postharvest BTH treatment induces resistance of peach (Prunus persica L. cv. Jiubao) fruit to infection by Penicillium expansum and enhances activity of fruit defense mechanisms. Postharvest Biol. Technol. 2005, 35, 263–269. [Google Scholar] [CrossRef]
- Terry, L.A.; Joyce, D.C. Suppression of grey mould on strawberry fruit with the chemical plant activator acibenzolar. Pest Manag. Sci. 2000, 56, 989–992. [Google Scholar] [CrossRef]
- Klessig, D.F.; Choi, H.W.; Dempsey, D.M.A. Systemic acquired resistance and salicylic acid: Past, present, and future. Mol. Plant Microb. Interact. 2018, 31, 871–888. [Google Scholar] [CrossRef]
- Gaffney, T.; Friedrich, L.; Vernooij, B.; Negrotto, D.; Nye, G.; Uknes, S.; Ryals, J. Requirement of salicylic acid for the induction of systemic acquired resistance. Science 1993, 261, 754–756. [Google Scholar] [CrossRef]
- Thaler, J.S.; Fidantsef, A.L.; Duney, S.S.; Bostock, R.M. Trade-offs in plant defense against pathogens and herbivores: A field demonstration of chemical elicitors of induced resistance. J. Chem. Ecol. 1999, 25, 1597–1609. [Google Scholar] [CrossRef]
- Yao, H.J.; Tian, S.P. Effects of pre- and post-harvest application of salicylic acid or methyl jasmonate on inducing disease resistance of sweet cherry fruit in storage. Postharvest Biol. Technol. 2005, 35, 253–262. [Google Scholar] [CrossRef]
- Yao, H.J.; Tian, S.P. Effects of a biocontrol agent and methyl jasmonate on postharvest disease of peach fruit and the possible mechanism involved. J. Appl. Microbiol. 2005, 98, 941–950. [Google Scholar] [CrossRef]
- Vlot, A.C.; Dempsey, D.M.A.; Klessig, D.F. Salicylic acid; a multifaceted hormone to combat disease. Annu. Rev. Phytopathol. 2009, 47, 177–206. [Google Scholar] [CrossRef]
- Prusky, D.; Alkan, N.; Mengiste, T.; Fluhr, R. Quiescent and necrotrophic lifestyle choice during postharvest disease development. Annu. Rev. Phytopathol. 2013, 51, 155–176. [Google Scholar] [CrossRef] [PubMed]
- Pieterse, C.M.; van der Does, D.; Zamioudis, C.; Leon-Reyes, A.; van Wees, S.C. Hormonal modulation of plant immunity. Annu. Rev. Cell Dev. Biol. 2012, 28, 489–521. [Google Scholar] [CrossRef] [PubMed]
- Kamle, M.; Borah, R.; Bora, H.; Jaiswal, A.K.; Singh, R.K.; Kumar, P. Systemic Acquired Resistance (SAR) and Induced Systemic Resistance (ISR): Role and mechanism of action against phytopathogens. In Fungal Biotechnology and Bioengineering; El-Latif, A., Ram, H., Upadhyay, S., Dutt, G., Chakravarthula, S., Vijai, M., Gupta, K., Eds.; Springer: Cham, Switzerland, 2020; pp. 457–470. [Google Scholar]
- Fresno, D.H.; Munné-Bosch, S. Differential tissue-specific jasmonic acid, salicylic acid, and abscisic acid dynamics in sweet cherry development and their implications in fruit-microbe interactions. Front. Plant Sci. 2021, 12, 640. [Google Scholar] [CrossRef]
- Ryals, J.A.; Neuenschwander, U.H.; Willits, M.G.; Molina, A.; Steiner, H.Y.; Hunt, M.D. Systemic acquired resistance. Plant Cell 1996, 8, 1809–1819. [Google Scholar] [CrossRef] [PubMed]
- Raskin, I. Role of salicylic acid in plants. Annu. Rev. Plant Biol. 1992, 43, 439–463. [Google Scholar] [CrossRef]
- Asghari, M.; Aghdam, M.S. Impact of salicylic acid on post-harvest physiology of horticultural crops. Trends Food Sci. Technol. 2010, 21, 502–509. [Google Scholar] [CrossRef]
- Tripathi, D.; Raikhy, G.; Kumar, D. Chemical elicitors of systemic acquired resistance—Salicylic acid and its functional analogs. Curr. Plant Biol. 2019, 17, 48–59. [Google Scholar] [CrossRef]
- Zainuri, D.C.; Joyce, H.; Wearing, L.; Terry, L. Effects of phosphonate and salicylic acid treatments on anthracnose disease development and ripening of ‘Kensington Pride’ mango fruit. Aust. J. Expt. Agric. 2001, 41, 805–881. [Google Scholar] [CrossRef]
- Zeng, K.; Cao, J.; Jiang, W. Enhancing disease resistance in harvested mango (Mangifera indica L. cv. ‘Matisu’) fruit by salicylic acid. J. Sci. Food Agric. 2006, 86, 694–698. [Google Scholar] [CrossRef]
- Xu, X.; Tian, S.P. Salicylic acid alleviated pathogen-induced oxidative stress in harvested sweet cherry fruit. Postharvest Biol. Technol. 2008, 49, 379–385. [Google Scholar] [CrossRef]
- Chan, Z.L.; Qin, G.Z.; Xu, X.B.; Li, B.Q.; Tian, S.P. Proteome approach to characterize proteins induced by antagonist yeast and salicylic acid in peach fruit. J. Proteome Res. 2007, 6, 1677–1688. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Cao, S.; Cai, Y.; Zheng, Y. Combination of salicylic acid and ultrasound to control postharvest blue mold caused by Penicillium expansum in peach fruit. Innov. Food Sci. Emerg. Technol. 2011, 12, 310–314. [Google Scholar] [CrossRef]
- Zhang, H.; Ma, L.; Wang, L.; Jiang, S.; Dong, Y.; Zheng, X. Biocontrol of gray mold decay in peach fruit by integration of antagonistic yeast with salicylic acid and their effects on postharvest quality parameters. Biol. Control. 2008, 47, 60–65. [Google Scholar] [CrossRef]
- Ezzat, A.; Szabó, Z.; Nyéki, J.; Holb, I.J. Preliminary results on salicylic acid treatment on brown rot caused by Monilinia laxa on Jumbo Cot fruit, Prunus armeniaca L. Int. J. Hortic. Sci. 2013, 19, 37–39. [Google Scholar] [CrossRef]
- Gačnik, S.; Munda, A.; Petkovšek, M.M. Effect of salicylic and methyl-salicylic acid on mycelial growth of different fungi and on infection of apple fruits with Monilinia laxa; Zbornik predavanj in referatov. In Proceedings of the 14th Slovensko Posvetovanje o Varstvu Rastlin z Mednarodno Udeležbo, Maribor, Slovenija, 5–6 March 2019; pp. 513–518. [Google Scholar]
- Lyousfi, N.; Lahlali, R.; Letrib, C.; Belabess, Z.; Ouaabou, R.; Ennahli, S.; Barka, E.A. Improving the biocontrol potential of bacterial antagonists with salicylic acid against brown rot disease and impact on nectarine fruits quality. Agronomy. 2021, 11, 209. [Google Scholar] [CrossRef]
- Reyes-Díaz, M.; Lobos, T.; Cardemil, L.; Nunes-Nesi, A.; Retamales, J.; Jaakola, L.; Ribera-Fonseca, A. Methyl jasmonate: An alternative for improving the quality and health properties of fresh fruits. Molecules 2016, 21, 567. [Google Scholar] [CrossRef]
- Tamari, G.; Borochov, A.; Atzorn, R.; Weiss, D. Methyl jasmonate induces pigmentation and flavonoid gene expression in petunia corollas: A possible role in wound response. Physiol. Plant. 1995, 94, 45–50. [Google Scholar] [CrossRef]
- Van Loon, L.C. Pathogenesis-related proteins. Plant Mol. Biol. 1995, 4, 111–116. [Google Scholar] [CrossRef]
- Bostock, R.M. Signal conflicts and synergies in induced resistance to multiple attackers. Physiol. Mol. Plant Pathol. 1999, 55, 99–109. [Google Scholar] [CrossRef]
- Campos, M.L.; Kang, J.H.; Howe, G.A. Jasmonate-triggered plant immunity. J. Chem. Ecol. 2014, 40, 657–675. [Google Scholar] [CrossRef] [PubMed]
- Cao, S.F.; Zheng, Y.H.; Yang, Z.F.; Tang, S.S.; Jin, P.; Wang, K.T.; Wang, X.M. Effect of methyl jasmonate on the inhibition of Colletotrichum acutatum infection in loquat fruit and the possible mechanisms. Postharvest Biol. Technol. 2008, 49, 301–307. [Google Scholar] [CrossRef]
- Jin, P.; Zheng, Y.; Tang, S.; Rui, H.; Wang, C. Enhancing disease resistance in peach fruit with methyl jasmonate. J. Sci. Food Agric. 2009, 89, 802–808. [Google Scholar] [CrossRef]
- Wang, L.; Jin, P.; Wang, J.; Jiang, L.; Shan, T.; Zheng, Y. Methyl jasmonate primed defense responses against Penicillium expansum in sweet cherry fruit. Plant Mol. Biol. Rep. 2015, 33, 1464–1471. [Google Scholar] [CrossRef]
- Tsao, R.; Zhou, T. Interaction of monoterpenoids, methyl jasmonate, and Ca2+ in controlling postharvest brown rot of sweet cherry. HortScience 2000, 35, 1304–1307. [Google Scholar] [CrossRef]
- Ezzat, A.; Hegedűs, A.; Szabó, S.; Ammar, A.; Szabó, Z.; Nyéki, J.; Molnár, B.; Holb, I.J. Temporal changes and correlations between quality loss parameters, antioxidant properties and enzyme activities in apricot fruit treated with methyl jasmonate and salicylic acid during cold storage and shelf-life. Appl. Sci. 2020, 10, 8071. [Google Scholar] [CrossRef]
- Maisuthisakul, P.; Pasuk, S.; Ritthiruangdejc, P. Relationship between antioxidant properties and chemical composition ofsome Thai plants. J. Food Compos. Anal. 2008, 21, 229–240. [Google Scholar] [CrossRef]
- Du, G.; Li, M.; Ma, F.; Lian, D. Antioxidant capacity and the relationship with polyphenol and vitamin C in Actinidia fruits. Food Chem. 2009, 113, 557–562. [Google Scholar] [CrossRef]
- Sulaiman, S.F.; Yusoff, N.A.; Eldeen, I.M.; Seow, E.M.; Sajak, A.A.B.; Supriatno Ooi, K.L. Correlation between total phenolic and mineral contents with antioxidant activity of eight Malaysian bananas (Musa sp.). J. Food Compos. Anal. 2011, 24, 1–10. [Google Scholar] [CrossRef]
- Ulewicz-Magulska, B.; Wesolowski, M. Total phenolic contents and antioxidant potential of herbs used for medical and culinary purposes. Plant Foods Hum. Nutr. 2019, 74, 61–67. [Google Scholar] [CrossRef]
- Ezzat, A.; El-Sherif, A.R.; Elgear, D.; Szabo, S.; Holb, I.J. A comparison of fruit and leaf parameters of apple in three orchard training systems. Zemdirb. Agric. 2020, 107, 373–382. [Google Scholar] [CrossRef]
- De Cal, A.; Melgarejo, P. Effects of long-wave UV light on Monilinia growth and identification of species. Plant Dis. 1999, 83, 62–65. [Google Scholar] [CrossRef]
- Gell, I.; Larena, I.; Melgarejo, P. Genetic diversity in Monilinia laxa populations in peach orchards in Spain. J. Phytopathol. 2007, 155, 549–556. [Google Scholar] [CrossRef]
- Fazekas, M.; Madar, A.; Sipiczki, M.; Miklós, I.; Holb, I.J. Genetic diversity in Monilinia laxa populations in stone fruit species in Hungary. World J. Microbiol. Biotechnol. 2014, 30, 1879–1892. [Google Scholar] [CrossRef] [PubMed]
- Femenia, A.; Garcia-Conesab, M.; Simal, S.; Rossello, C. Characterisation of the cell walls of loquat (Eriobotrya japonica L.) fruit tissues. Carbohydr. Polym. 1998, 35, 169–171. [Google Scholar] [CrossRef]
- Singleton, V.L.; Rossi, J.A. Colorimetry of total phenolics with phosphomolybdic phosphotungstic acid “reagents”. Am. J. Enol. Vitic. 1965, 16, 144–158. [Google Scholar]
- Benzie, I.F.F.; Strain, J.J. The ferric reducing ability of plasma (FRAP) as a measure of “antioxidant power”: The FRAP assay. Anal. Biochem. 1996, 239, 70–76. [Google Scholar] [CrossRef] [PubMed]
- Assis, J.S.; Maldonado, R.; Mnoz, T.; Escribano, M.I.; Merodio, C. Effect of high carbon dioxid concentration on PAL activity and phenol content in ripening cherimoya fruit. Postharvest Biol. Technol. 2001, 23, 33–39. [Google Scholar] [CrossRef]
- Rao, M.V.; Paliyath, G.; Ormord, D.P. Ultraviolet-B- and ozon induced biochemical changes in antioxidant enzymes of Arabidopsis thaliana. Plant Physiol. 1996, 110, 125–136. [Google Scholar] [CrossRef]
- Kochba, J.; Lavee, S.; Spiege, R.P. Difference in peroxidase activity and isoenzymes in embryogenic and non-embryogenic ‘Shamouti’ orange ovular callus lines. Plant Cell Physiol. 1977, 18, 463–467. [Google Scholar] [CrossRef]
- Basto, M.; Pereira, J.M. An SPSSR-menu for ordinal factor analysis. J. Stat. Softw. 2012, 46, 1–29. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020; Available online: https://www.R-project.org/ (accessed on 10 May 2020).
- Revelle, W. Psych: Procedures for Personality and Psychological Research; Northwestern University: Evanston, IL, USA, 2015; Available online: http://CRAN.R-project.org/package=psych (accessed on 12 June 2015).
- Lê, S.; Josse, J.; Husson, F. FactoMineR: An R package for multivariate analysis. J. Stat. Softw. 2008, 25, 30294. Available online: https://www.jstatsoft.org/article/view/v025i01 (accessed on 25 October 2008). [CrossRef]
- Kassambara, A.; Mundt, A. Factoextra: Extract and Visualize the Results of Multivariate Data Analyses. R Package Version 1.0.6.2019. Available online: https://CRAN.R-project.org/package=factoextra (accessed on 1 June 2019).
- Leslie, C.A.; Romani, R.J. Salicylic acid: A new inhibitor of ethylene biosynthesis. Plant Cell Rep. 1986, 5, 144–146. [Google Scholar] [CrossRef] [PubMed]
- Ezzat, A.; Ammar, A.; Szabó, Z.; Nyéki, J.; Holb, I.J. Postharvest treatments with methyl jasmonate and salicylic acid for maintaining physico-chemical characteristics and sensory quality properties of apricot fruit during cold storage and shelf-life. Pol. J. Food Nutr. Sci. 2017, 67, 159–166. [Google Scholar] [CrossRef]
- Baswal, A.K.; Dhaliwal, H.S.; Singh, Z.; Mahajan, B.V.C.; Gill, K.S. Postharvest application of methyl jasmonate, 1-methylcyclopropene and salicylic acid extends the cold storage life and maintain the quality of ‘Kinnow’ mandarin (Citrus nobilis L. X C. deliciosa L.) fruit. Postharvest Biol. Technol. 2020, 161, 111064. [Google Scholar] [CrossRef]
- Babalar, M.; Asghari, M.; Talaei, A.; Khosroshahi, A. Effect of pre- and postharvest salicylic acid treatment on ethylene production, fungal decay and overall quality of Selva strawberry fruit. Food Chem. 2007, 105, 449–453. [Google Scholar] [CrossRef]
- Whetten, R.; Sederoff, R. Lignin biosynthesis. Plant Cell 1995, 7, 1001–1013. [Google Scholar] [CrossRef]
- Muro-Villanueva, F.; Mao, X.; Chapple, C. Linking phenylpropanoid metabolism, lignin deposition, and plant growth inhibition. Curr. Opin. Biotechnol. 2019, 56, 202–208. [Google Scholar] [CrossRef] [PubMed]
- Uritani, I.; Oba, K. The tissue slice system as a model for studies of host-parasite relationships. In Biochemistry of Wounded Plant Tissues; Kahl, G., Ed.; Walter de Gruyter: New York, NY, USA, 1978; pp. 287–308. [Google Scholar]
- Vance, C.P.; Kirk, T.K. Sherwood lignification as a mechanism of disease resistance. Annu. Rev. Phytopathol. 1980, 18, 259–288. [Google Scholar] [CrossRef]
- Lu, Y.C.; Lu, Y.; Fan, X. Structure and characteristics of lignin. In Lignin; Sharma, S., Kumar, A., Eds.; Springer Series on Polymer and Composite Materials; Springer: Cham, Switzerland, 2020; pp. 17–75. [Google Scholar]
- Govender, N.T.; Mahmood, M.; Seman, I.A.; Wong, M.Y. The phenylpropanoid pathway and lignin in defense against ganoderma boninense colonized root tissues in oil palm (Elaeis guineensis Jacq.). Front. Plant Sci. 2017, 8, 1395. [Google Scholar] [CrossRef]
- Su, X.G.; Zheng, Y.H.; Feng, L.; Zhang, L.; Wang, F. Effects of exogenous MeJA on postharvest senescence and decay of vegetable soybean pods. J. Plant Physiol. Mol. Biol. 2003, 29, 52–58. [Google Scholar]
- Solecka, D.; Kacperska, A. Phenylpropanoid deficiency affects the course of plant acclimation to cold. Plant Physiol. 2003, 119, 253–262. [Google Scholar] [CrossRef]
- Wang, K.; Jiang, S.; Pu, T.; Fan, L.; Su, F.; Ye, M. Antifungal activity of phenolic monoterpenes and structure-related compounds against plant pathogenic fungi. Nat. Prod. Res. 2019, 33, 1423–1430. [Google Scholar] [CrossRef] [PubMed]
- Tsai, C.J.; Harding, S.A.; Tschaplinshi, T.J.; Lindroth, R.L.; Yuan, Y. Genome-wide analysis of the structural genes regulating defense phenylpropanoid metabolism in Populus. New Phytol. 2006, 172, 47–56. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.; Shahzad, B.; Rehman, A.; Bhardwaj, R.; Landi, M.; Zheng, B. Response of phenylpropanoid pathway and the role of polyphenols in plants under abiotic stress. Molecules 2019, 24, 2452. [Google Scholar] [CrossRef] [PubMed]
- Reimers, P.J.; Leach, I.E. Race-specific resistance to Xanthomonas oryzae pv. oryzae conferred by bacterial blight resistance gene Xa-10 in rice Oryza sativa involves accumulation of a lignin-like substance in host tissues. Physiol. Mol. Plant Pathol. 1999, 38, 39–55. [Google Scholar]
- De Ascensao, A.F.R.D.C.; Dubrey, I.A. Soluble and wall-bound phenolic polymers in Musa acuminata roots exposed to elicitors from Fusarium oxysporum f.sp. cubens. Phytochemistry 2003, 63, 679–686. [Google Scholar] [CrossRef]
- Jiang, S.; Han, S.; He, D.; Cao, G.; Fang, K.; Xiao, X.; Wan, X. The accumulation of phenolic compounds and increased activities of related enzymes contribute to early defense against walnut blight. Physiol. Mol. Plant Pathol. 2019, 108, 101433. [Google Scholar] [CrossRef]
- Mendoza, D.; Cuaspud, O.; Arias, J.P.; Ruiz, O.; Arias, M. Effect of salicylic acid and methyl jasmonate in the production of phenolic compounds in plant cell suspension cultures of Thevetia peruviana. Biotechnol. Rep. 2018, 19, e00273. [Google Scholar] [CrossRef]
- Cappellari, L.D.R.; Santoro, M.V.; Schmidt, A.; Gershenzon, J.; Banchio, E. Improving phenolic total content and monoterpene in Mentha x piperita by using salicylic acid or methyl jasmonate combined with Rhizobacteria inoculation. Int. J. Mol. Sci. 2020, 21, 50. [Google Scholar] [CrossRef]
- Zhu, Z.; Tian, S.P. Resistant responses of tomato fruit treated with exogenous methyl jasmonate to Botrytis cinerea infection. Sci. Hortic. 2012, 142, 38–43. [Google Scholar] [CrossRef]
- Zhang, L.; Zhang, F.; Melotto, M.; Yao, J.; He, S.Y. Jasmonate signaling and manipulation by pathogens and insects. J. Exp. Bot. 2017, 68, 1371–1385. [Google Scholar] [CrossRef]
- Poveda, J. Use of plant-defense hormones against pathogen-diseases of postharvest fresh produce. Physiol. Mol. Plant Pathol. 2020, 111, 101521. [Google Scholar] [CrossRef]
- Milosevic, N.; Slusarenko, A.J. Active oxygen metabolism and lignification in the hypersensitive response in bean. Physiol. Mol. Plant. Pathol. 1996, 49, 143–157. [Google Scholar] [CrossRef]
- Yadav, V.; Wang, Z.; Wei, C.; Amo, A.; Ahmed, B.; Yang, X.; Zhang, X. Phenylpropanoid pathway engineering: An emerging approach towards plant defense. Pathogens 2020, 9, 312. [Google Scholar] [CrossRef] [PubMed]
- Saltveit, M.E. Chilling injury is reduced in cucumber and rice seedlings and in tomato pericarp discs by heat-shocks applied after chilling. Postharvest Biol. Technol. 2001, 21, 169–177. [Google Scholar] [CrossRef]
- Inze, D.; Montagu, M.V. Oxidative stress in plants. Curr. Opin. Biotechnol. 1995, 6, 153–158. [Google Scholar] [CrossRef]
- Berwal, M.; Ram, C. Superoxide dismutase: A stable biochemical marker for abiotic stress tolerance in higher plants. Abiotic Biotic Stress Plants 2018, 1, 1–10. [Google Scholar]
- Chittoor, J.M.; Leach, J.E.; White, F.F. Induction of peroxidase during defense against pathogens. In Pathogenesis: Related Proteins in Plants; Datta, S.K., Muthukrishnan, S., Eds.; CRC Press: Boca Raton, FL, USA, 1999; pp. 1–291. [Google Scholar]
- Pandey, V.P.; Awasthi, M.; Singh, S.; Tiwari, S.; Dwivedi, U.N. A comprehensive review on function and application of plant peroxidases. Anal. Biochem. 2017, 6, 1009–2161. [Google Scholar] [CrossRef]
- Vera-Estrella, R.; Blumwald, E.; Higgins, V.G. Effect of specific elicitors of Cladosporium fulvum on tomato suspension cells. Plant Physiol. 1992, 99, 1208–1215. [Google Scholar] [CrossRef]
- Ali, M.; Cheng, Z.; Ahmad, H.; Hayat, S. Reactive oxygen species (ROS) as defenses against a broad range of plant fungal infections and case study on ROS employed by crops against Verticillium dahliae wilts. J. Plant Interact. 2018, 13, 353–363. [Google Scholar] [CrossRef]
- Francoz, E.; Ranocha, P.; Le Ru, A.; Martinez, Y.; Fourquaux, I.; Jauneau, A.; Burlat, V. Pectin demethylesterification generates platforms that anchor peroxidases to remodel plant cell wall domains. Dev. Cell 2019, 48, 261–276. [Google Scholar] [CrossRef] [PubMed]
- Huckelhoven, R.; Fodor, J.; Preis, C.; Kogel, K.H. Hypersensitive cell death and papilla formation in barley attacked by the powdery mildew fungus are associated with hydrogen peroxide but not with salicylic acid accumulation. Plant Physiol. 1999, 119, 1251–1260. [Google Scholar] [CrossRef] [PubMed]
- Kristensen, B.K.; Bloch, H.; Rasmussen, S.K. Barley coleoptile peroxidases. Purification, molecular cloning, and induction by pathogens. Plant Physiol. 1999, 120, 501–512. [Google Scholar] [CrossRef] [PubMed]
- Cvikrová, M.; Malá, J.; Hrubcová, M. Eder; J. Soluble and cell wall-bound phenolics and lignin in Ascocalyx abietina infected Norway spruces. Plant Sci. 2006, 170, 563–570. [Google Scholar] [CrossRef]
- Moosa, A.; Sahi, S.T.; Khan, S.A.; Malik, A.U. Salicylic acid and jasmonic acid can suppress green and blue moulds of citrus fruit and induce the activity of polyphenol oxidase and peroxidase. Folia Hortic. 2019, 31, 195–204. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatments | Brown Rot Incidence (%) | Lesion Diameter (mm) |
|---|---|---|
| Water-treated control | 58.42 ± 3.45 a 1 | 15.49 ± 1.01 a |
| MeJA | ||
| 0.1 mmol L−1 | 36.48 ± 1.96 b | 15.77 ± 0.45 a |
| 0.4 mmol L−1 | 22.84 ± 0.08 c | 11.09 ± 0.69 c |
| 0.7 mmol L−1 | 23.50 ± 0.72 c | 12.81 ± 0.23 b |
| SA | ||
| 0.5 mmol L−1 | 32.35 ± 2.09 b | 12.99 ± 0.59 b |
| 2 mmol L−1 | 21.50 ± 2.12 c | 10.31 ± 0.47 c |
| 5 mmol L−1 | 22.50 ± 0.70 c | 10.44 ± 0.61 c |
| LSD0.05 | 3.86 | 1.02 |
| Methyl Jasmonate (MeJA) | ||||||
|---|---|---|---|---|---|---|
| FF | LC | TSPC | TAC | PAL | SOD | |
| LC | 0.885 | |||||
| 0.011 | ||||||
| TSPC | 0.800 | 0.668 | ||||
| 0.046 | 0.134 | |||||
| TAC | 0.956 | 0.900 | 0.668 | |||
| <0.001 | 0.005 | 0.134 | ||||
| PAL | −0.838 | −0.736 | −0.663 | −0.899 | ||
| 0.026 | 0.079 | 0.139 | 0.007 | |||
| SOD | −0.698 | −0.681 | −0.453 | −0.817 | 0.909 | |
| 0.104 | 0.121 | 0.361 | 0.041 | 0.003 | ||
| POD | −0.827 | −0.727 | −0.553 | −0.911 | 0.952 | 0.936 |
| 0.028 | 0.085 | 0.294 | 0.003 | <0.001 | 0.001 | |
| Salicylic Acid (SA) | ||||||
| FF | LC | TSPC | TAC | PAL | SOD | |
| LC | 0.783 | |||||
| 0.061 | ||||||
| TSPC | 0.882 | −0.573 | ||||
| 0.006 | 0.261 | |||||
| TAC | 0.941 | −0.867 | 0.439 | |||
| <0.001 | 0.012 | 0.384 | ||||
| PAL | −0.943 | 0.859 | −0.412 | −0.951 | ||
| <0.001 | 0.016 | 0.408 | <0.001 | |||
| SOD | −0.885 | 0.765 | −0.189 | −0.913 | 0.936 | |
| 0.006 | 0.071 | 0.727 | 0.003 | <0.001 | ||
| POD | −0.949 | 0.889 | −0.455 | −0.974 | 0.979 | 0.903 |
| <0.001 | 0.006 | 0.358 | <0.001 | <0.001 | 0.004 | |
| Methyl Jasmonate (MeJA) | ||||||
|---|---|---|---|---|---|---|
| FF | LC | TSPC | TAC | PAL | SOD | |
| LC | 0.598 | |||||
| 0.232 | ||||||
| TSPC | 0.943 | −0.612 | ||||
| <0.001 | 0.204 | |||||
| TAC | 0.896 | −0.714 | 0.861 | |||
| 0.002 | 0.092 | 0.015 | ||||
| PAL | −0.801 | 0.705 | −0.761 | −0.902 | ||
| 0.046 | 0.101 | 0.070 | 0.005 | |||
| SOD | −0.869 | 0.716 | −0.826 | 0.927 | 0.971 | |
| 0.015 | 0.091 | 0.029 | <0.001 | <0.001 | ||
| POD | −0.874 | 0.695 | −0.724 | −0.919 | 0.941 | 0.969 |
| 0.013 | 0.111 | 0.085 | <0.001 | <0.001 | <0.001 | |
| Salicylic Acid (SA) | ||||||
| FF | LC | TSPC | TAC | PAL | SOD | |
| LC | 0.622 | |||||
| 0.186 | ||||||
| TSPC | 0.849 | −0.614 | ||||
| 0.009 | 0.202 | |||||
| TAC | 0.897 | −0.671 | 0.928 | |||
| 0.002 | 0.132 | <0.001 | ||||
| PAL | −0.798 | 0.863 | −0.776 | −0.841 | ||
| 0.051 | 0.015 | 0.066 | 0.022 | |||
| SOD | −0.702 | 0.878 | −0.621 | −0.725 | 0.944 | |
| 0.102 | 0.012 | 0.188 | 0.085 | <0.001 | ||
| POD | −0.766 | 0.931 | −0.689 | −0.784 | 0.962 | 0.975 |
| 0.071 | 0.002 | 0.116 | 0.061 | <0.001 | <0.001 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ezzat, A.; Szabó, S.; Szabó, Z.; Hegedűs, A.; Berényi, D.; Holb, I.J. Temporal Patterns and Inter-Correlations among Physical and Antioxidant Attributes and Enzyme Activities of Apricot Fruit Inoculated with Monilinia laxa under Salicylic Acid and Methyl Jasmonate Treatments under Shelf-Life Conditions. J. Fungi 2021, 7, 341. https://doi.org/10.3390/jof7050341
Ezzat A, Szabó S, Szabó Z, Hegedűs A, Berényi D, Holb IJ. Temporal Patterns and Inter-Correlations among Physical and Antioxidant Attributes and Enzyme Activities of Apricot Fruit Inoculated with Monilinia laxa under Salicylic Acid and Methyl Jasmonate Treatments under Shelf-Life Conditions. Journal of Fungi. 2021; 7(5):341. https://doi.org/10.3390/jof7050341
Chicago/Turabian StyleEzzat, Ahmed, Szilárd Szabó, Zoltán Szabó, Attila Hegedűs, Dorina Berényi, and Imre J. Holb. 2021. "Temporal Patterns and Inter-Correlations among Physical and Antioxidant Attributes and Enzyme Activities of Apricot Fruit Inoculated with Monilinia laxa under Salicylic Acid and Methyl Jasmonate Treatments under Shelf-Life Conditions" Journal of Fungi 7, no. 5: 341. https://doi.org/10.3390/jof7050341
APA StyleEzzat, A., Szabó, S., Szabó, Z., Hegedűs, A., Berényi, D., & Holb, I. J. (2021). Temporal Patterns and Inter-Correlations among Physical and Antioxidant Attributes and Enzyme Activities of Apricot Fruit Inoculated with Monilinia laxa under Salicylic Acid and Methyl Jasmonate Treatments under Shelf-Life Conditions. Journal of Fungi, 7(5), 341. https://doi.org/10.3390/jof7050341