
Biocontrol Effects of Paecilomyces variotii against Fungal Plant Diseases


Abstract
1. Introduction
2. Materials and Methods
2.1. Fungal Isolates
2.2. Dual Culture Bioassays
2.3. Effects of Non-Volatile Metabolites of P. variotii on Fungal Pathogens
2.4. Antifungal Volatile Organic Compounds Bioassay
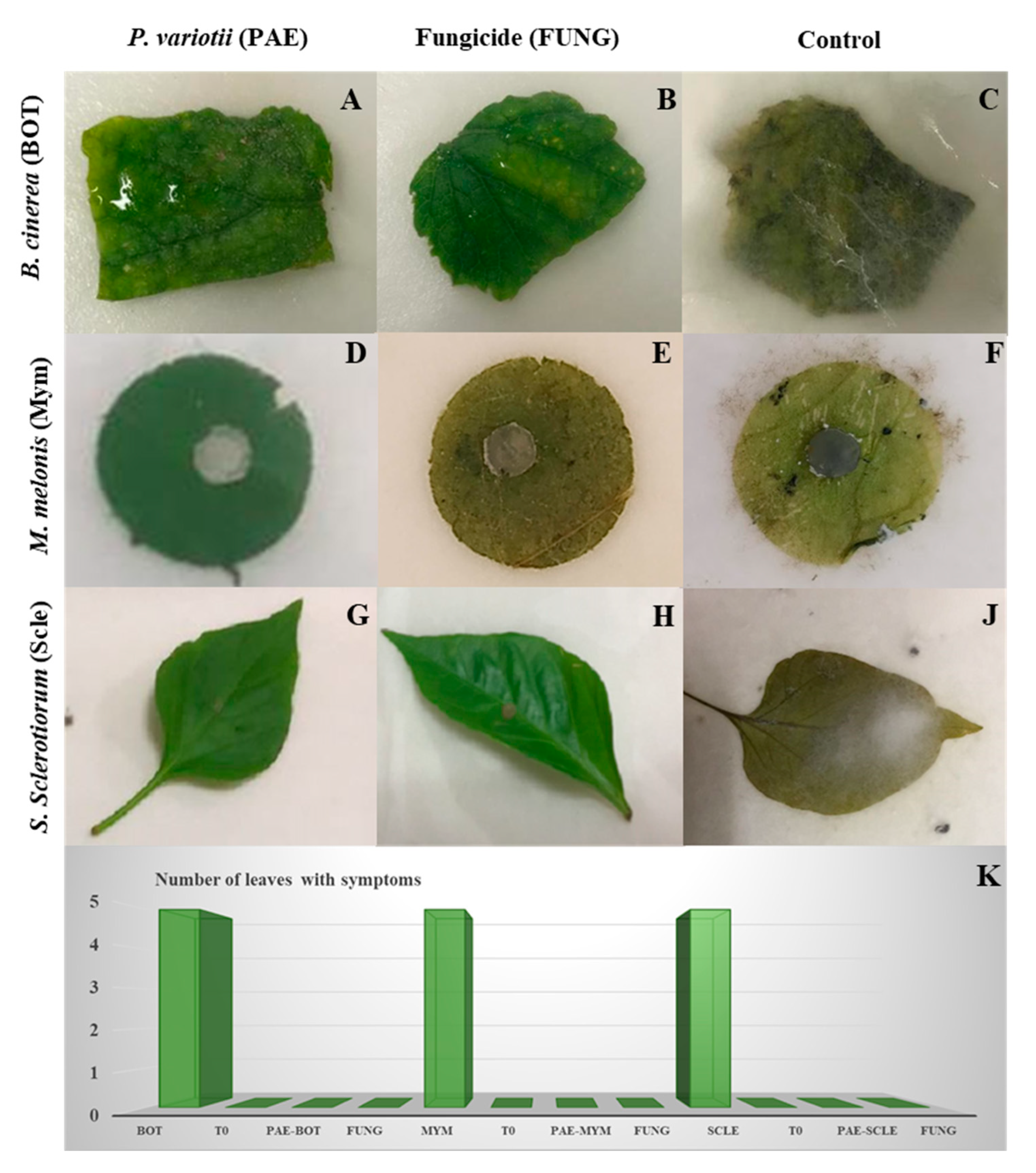
2.5. Detached Leaf Assay
2.6. Compatibility of P. variotii with Fungicides
2.7. Greenhouse Evaluation of P. variotii on Disease Severity of Seven Phytopathogens
2.8. Statistical Analyses
3. Results
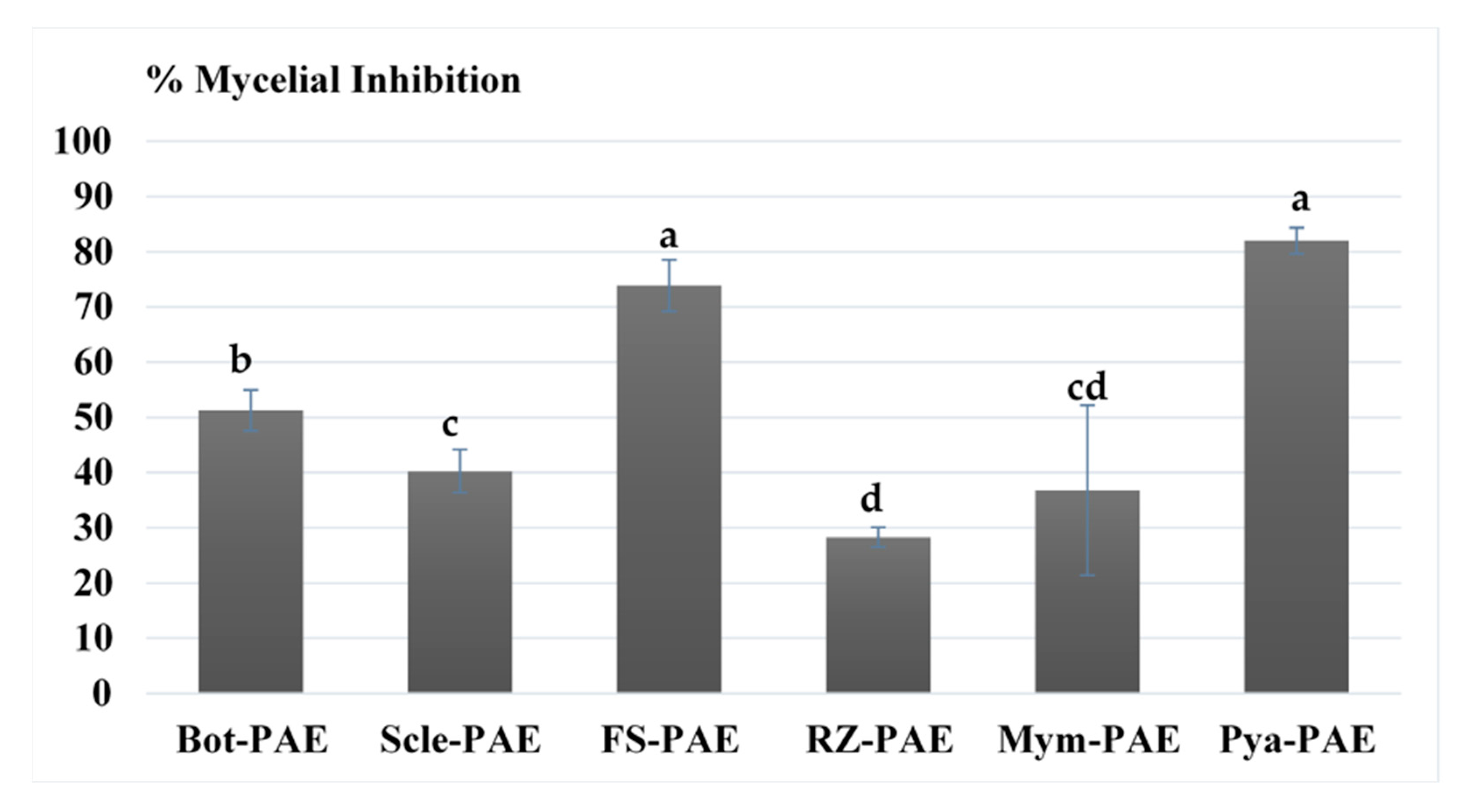
3.1. In Vitro Antagonism of P. variotii
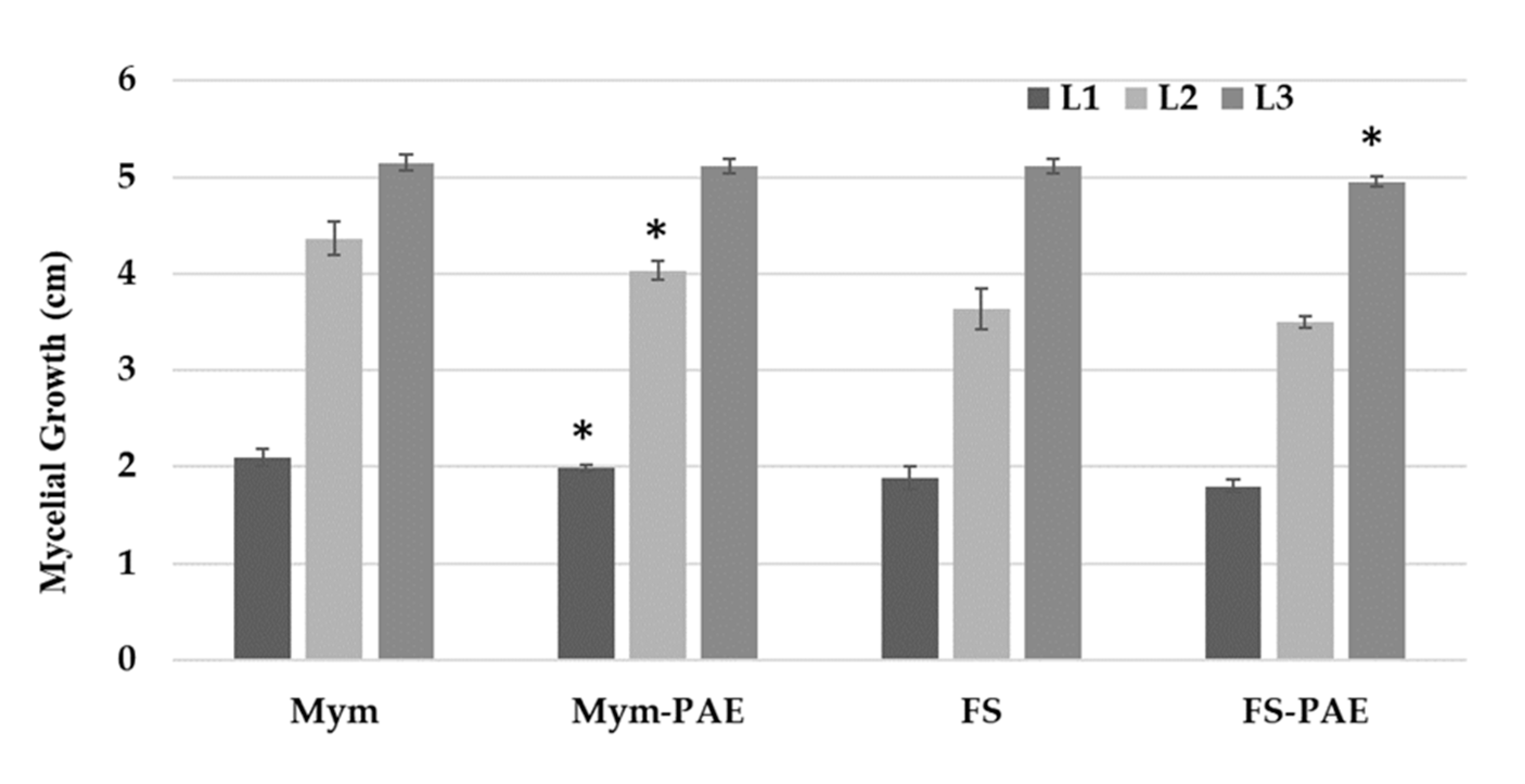
3.2. In Vitro Effects of Cell-Free Filtrate and Antifungal Volatile Compounds of P. variotii on Mycelial Growth of Phytopathogens
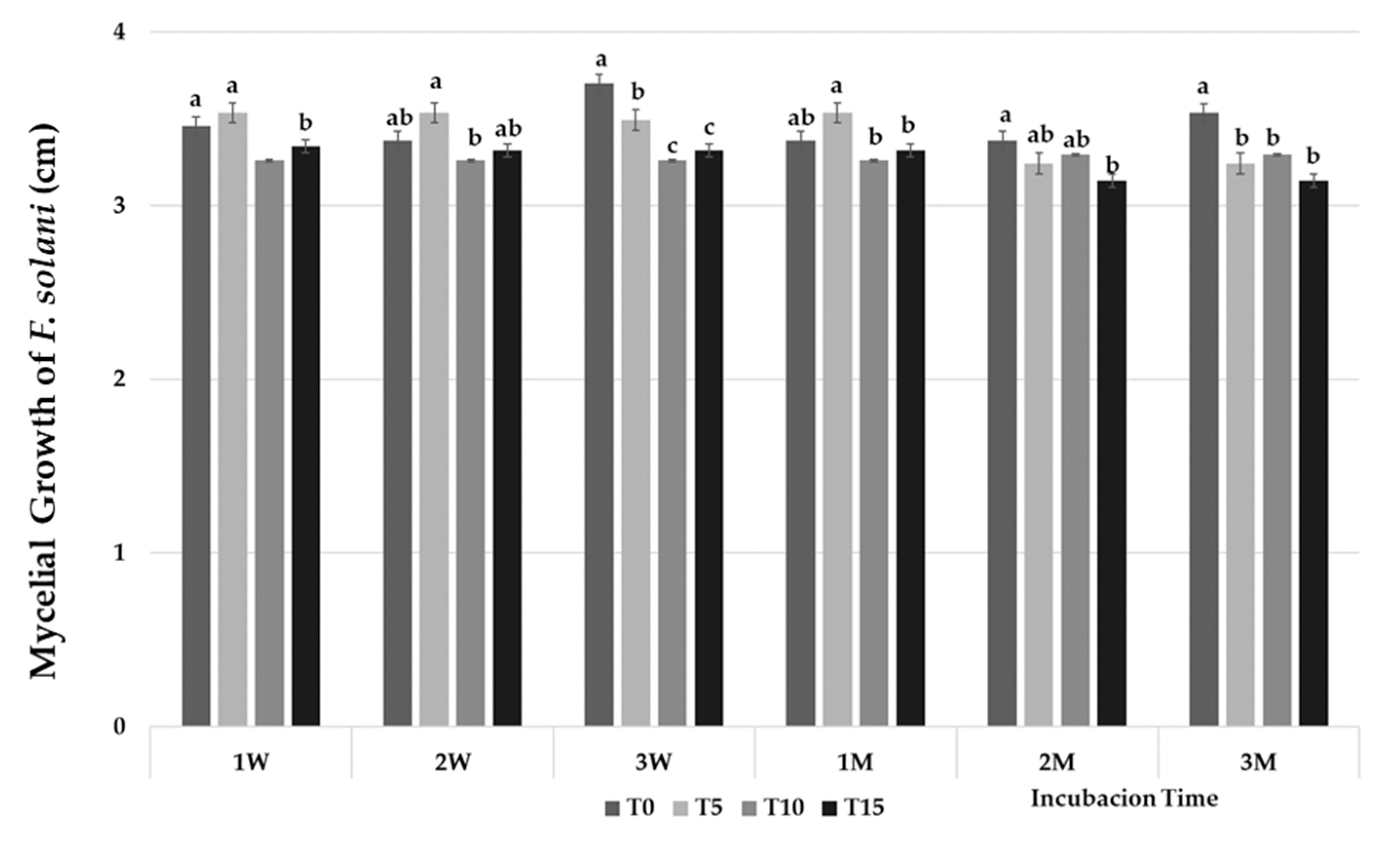
3.3. Compatibility for Use of P. variotii with Fungicides
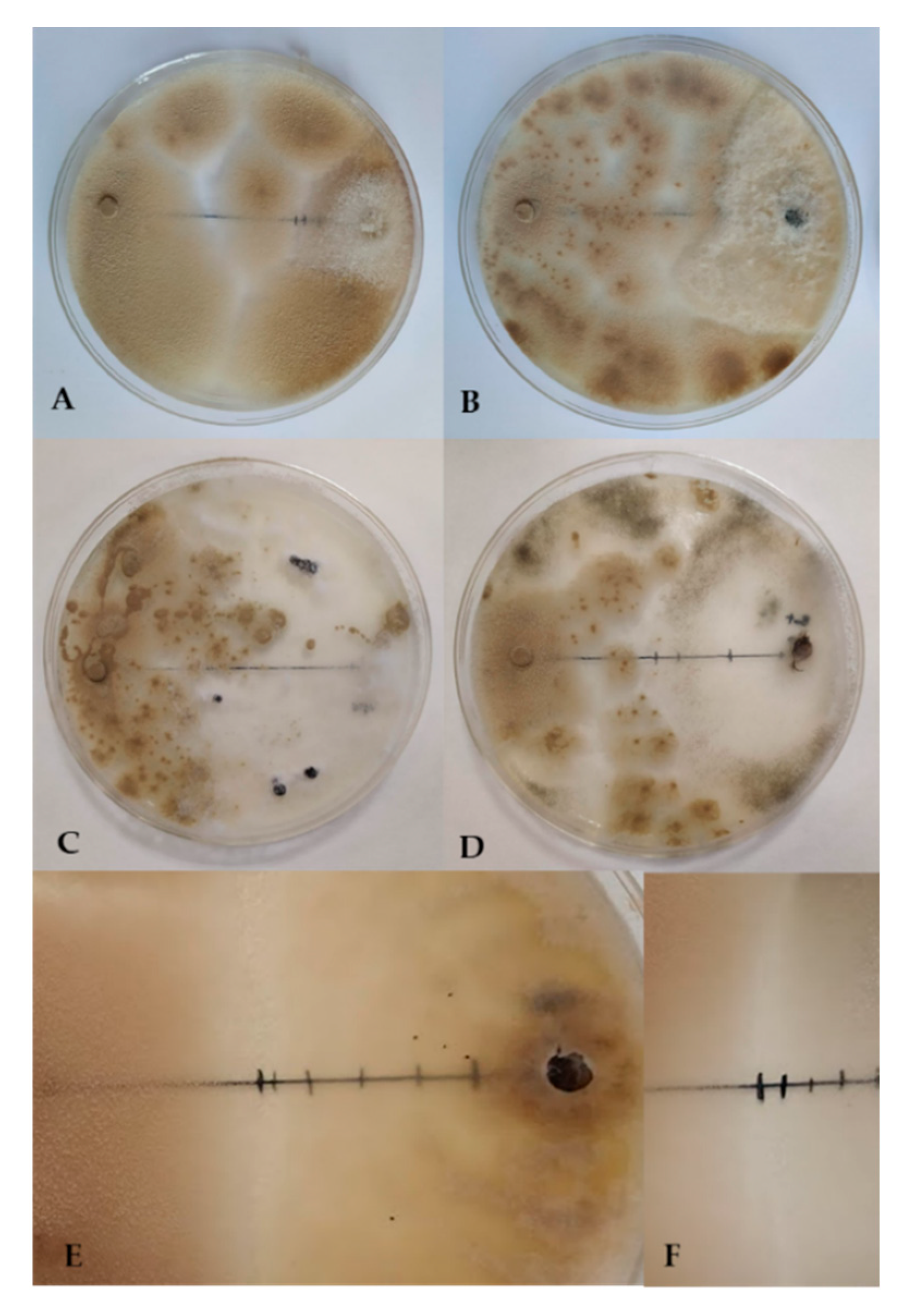
3.4. Detached Leaves
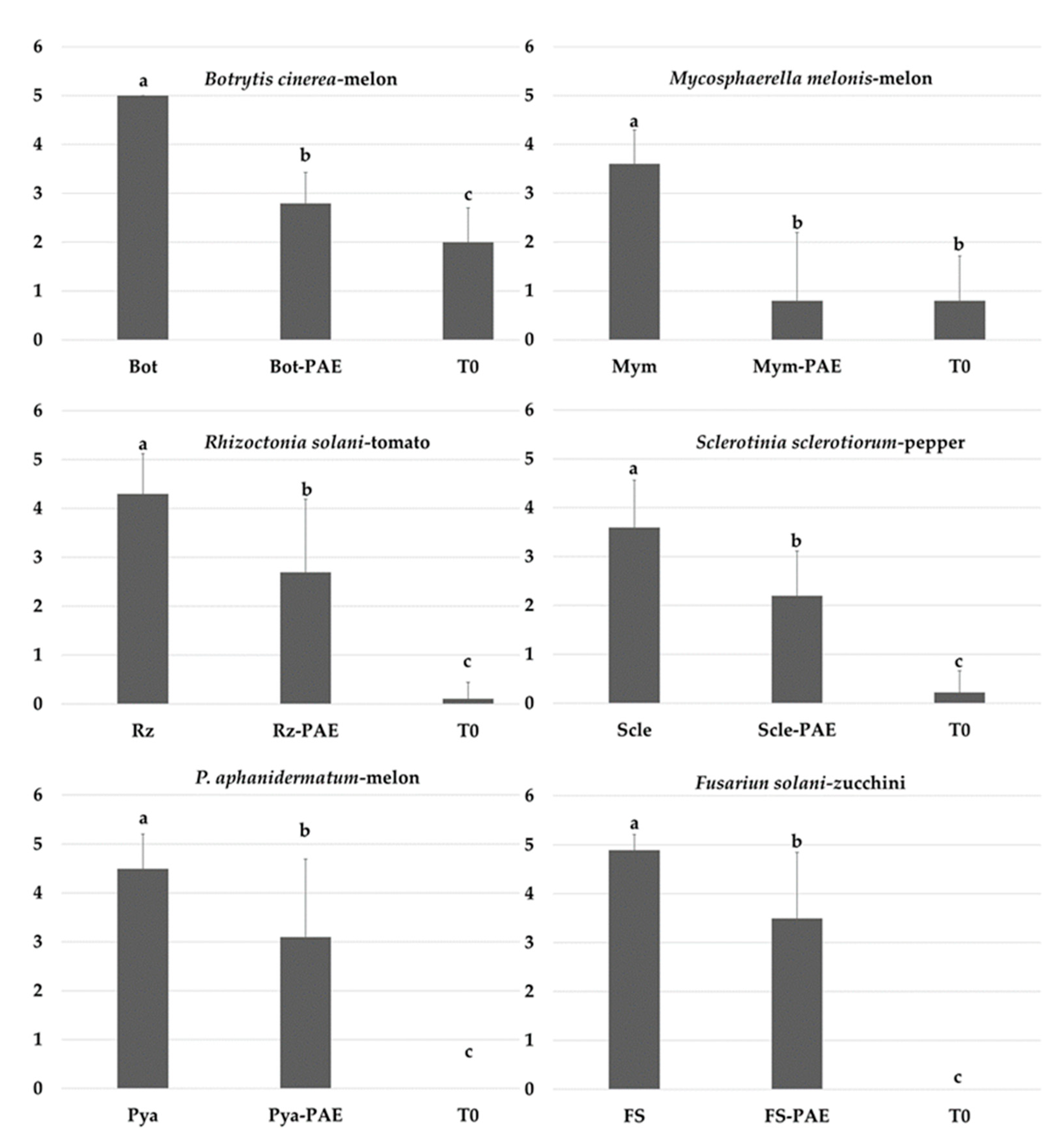
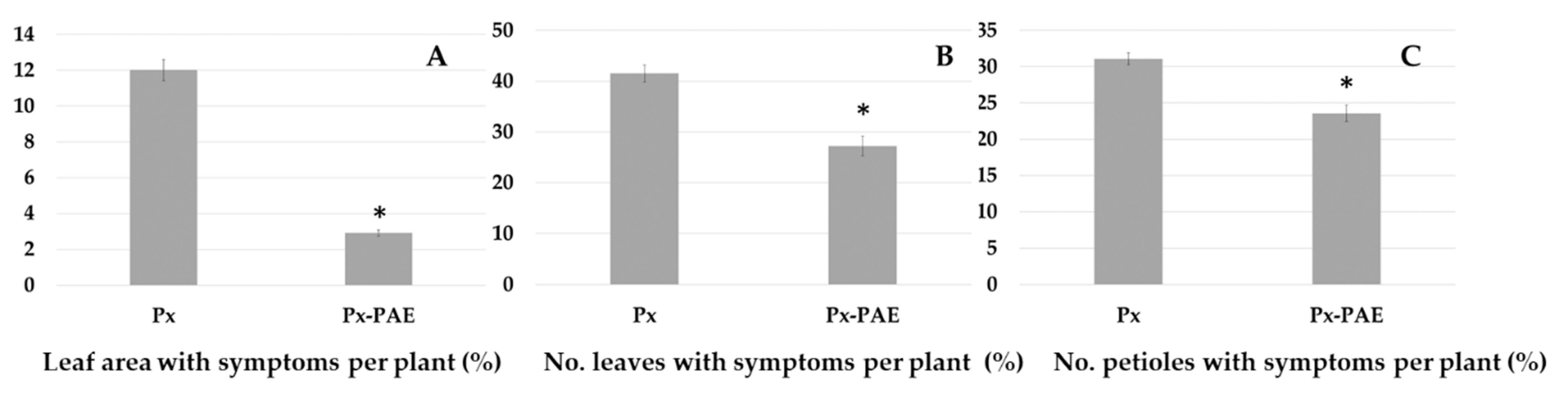
3.5. In Vivo Suppressive Effects of P. variotii on Diseases Caused by Phytopathogenic Fungi
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Romanazzi, G.; Smilanick, J.L.; Feliziani, E.; Droby, S. Integrated management of postharvest gray mold on fruit crops. Postharvest Biol. Technol. 2016, 113, 69–76. [Google Scholar] [CrossRef]
- Romeralo, C.; Santamaría, O.; Pando, V.; Diez, J.J. Fungal endophytes reduce necrosis length produced by Gremmeniella abietina in Pinus halepensis seedlings. Biol. Control 2015, 80, 30–39. [Google Scholar] [CrossRef]
- Xiong, W.; Li, R.; Ren, Y.; Liu, C.; Zhao, Q.; Wu, H.; Alexandre, J.; Qirong, S. Distinct roles for soil fungal and bacterial communities associated with the suppression of vanilla Fusarium wilt disease. Soil Biol. Biochem. 2017, 107, 198–207. [Google Scholar] [CrossRef]
- Gastavsson, J.; Cederberg, C.; Sonesson, U. Global Food Losses and Food Waste; Food and Agriculture Organization (FAO) of the United Nations: Rome, Italy, 2011; p. 37. [Google Scholar]
- Savary, S.; Ficke, A.; Aubertot, J.N.; Hollier, C. Crop losses due to diseases and their implications for global food production losses and food security. Food Secur. 2012, 4, 519–537. [Google Scholar] [CrossRef]
- Sellem, I.; Triki, M.A.; Elleuch, L.; Cheffi, M.; Chakchouk, A.; Smaoui, S.; Mellouli, L. The use of newly isolated Streptomyces strain TN258 as potential biocontrol agent of potato tubers leak caused by Pythium ultimum. J. Basic Microbiol. 2017, 57, 393–401. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Guan, Y.; Dong, Y.; Zhao, L.; Rong, S.; Chen, W.; Lv, M.; Xu, H.; Gao, X.; Chen, R.; et al. Isolation and evaluation of endophytic Bacillus tequilensis GYLH001 with potential application for biological control of Magnaporthe oryzae. PLoS ONE 2018, 13, e0203505. [Google Scholar] [CrossRef] [PubMed]
- Samson, R.A.; Houbraken, J.; Varga, J.; Frisvad, J.C. Polyphasic taxonomy of the heat resistant ascomycete genus Byssochlamys and its Paecilomyces anamorphs. Persoonia 2009, 22, 14. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, T.T.T.; Paul, N.C.; Lee, H.B. Characterization of Paecilomyces variotii and Talaromyces amestolkiae in Korea based on the morphological characteristics and multigene phylogenetic analyses. Mycobiology 2016, 44, 248–259. [Google Scholar] [CrossRef]
- Houbraken, J.; Varga, J.; Rico-Munoz, E.; Johnson, S.; Samson, R.A. Sexual reproduction as the cause of heat resistance in the food spoilage fungus Byssochlamys spectabilis (anamorph Paecilomyces variotii). Appl. Environ. Microbiol. 2008, 74, 1613–1619. [Google Scholar] [CrossRef] [PubMed]
- Houbraken, J.; Samson, R.A.; Frisvad, J.C. Byssochlamys: Significance of Heat Resistance and Mycotoxin Production. In Advances in Food Mycology; Springer: Boston, MA, USA, 2006; pp. 211–224. [Google Scholar]
- Samson, R.A.; Houbraken, J.; Summerbell, R.C.; Flannigan, B.; Miller, J.D. Common and important species of fungi and actinomycetes in indoor environments. In Microorganisms in Home and Indoor Work Environments: Diversity, Health Impacts, Investigation and Control; Wiley: Boca Raton, FL, USA, 2002; pp. 285–473. [Google Scholar]
- He, J.; Kang, J.; Lei, B.; Wen, T. Paecilomyces wawuensis, a new species isolated from soil in China. Mycotaxon 2011, 115, 303–310. [Google Scholar] [CrossRef]
- Hussain, A.; Shrivastav, A.; Jain, S.K.; Baghel, R.K.; Rani, S.; Agrawal, M.K. Cellulolytic enzymatic activity of soft rot filamentous fungi Paecilomyces variotii. Adv. Biores. 2012, 3, 10–17. [Google Scholar]
- Marante, F.J.T.; Mioso, R.; Barrera, J.B.; González, J.E.G.; Rodríguez, J.J.S.; De Laguna, I.H.B. Structural characterization and metabolite profiling of the facultative marine fungus Paecilomyces variotii. Ann. Microbiol. 2012, 62, 1601–1607. [Google Scholar] [CrossRef]
- Silva, M.O.; Kawai, K.; Hosoe, T.; Takaki, G.C.; Gusmão, N.B.; Fukushima, K. Viriditoxin, An Antibacterial Substance Produced by Mangrove Endophytic Fungus Paecilomyces variotii. In Microbial Pathogens and Strategies for Combating Them: Science, Technology and Education; Formatex: Badajoz, Spain, 2013; Volume 2, pp. 1406–1411. [Google Scholar]
- Steiner, B.; Aquino, V.R.; Paz, A.A.; Silla, L.M.D.R.; Zavascki, A.; Goldani, L.Z. Paecilomyces variotii as an emergent pathogenic agent of pneumonia. Case Rep. Infect. Dis. 2013, 273848. [Google Scholar] [CrossRef]
- Li, X.Q.; Xu, K.; Liu, X.M.; Zhang, P.A. Systematic review on secondary metabolites of Paecilomyces species: Chemical diversity and biological activity. Planta Med. 2020, 86, 805–821. [Google Scholar] [CrossRef]
- Moreno-Gavíra, A.; Huertas, V.; Diánez, F.; Sánchez-Montesinos, B.; Santos, M. Paecilomyces and its importance in the biological control of agricultural pests and diseases. Plants 2020, 9, 1746. [Google Scholar] [CrossRef] [PubMed]
- Ui, H.; Shiomi, K.; Suzuki, H.; Hatano, H.; Morimoto, H.; Yamaguchi, Y.; Masuma, R.; Sakamoto, K.; Kita, K.; Miyoshi, H.; et al. Paecilaminol, a new NADH-fumarate reductase inhibitor, produced by Paecilomyces sp. FKI-0550. J. Antibiot. 2006, 59, 591–596. [Google Scholar] [CrossRef]
- Pratella, G.C.; Mari, M. Effectiveness of Trichoderma, Gliocladium and Paecilomyces in postharvest fruit protection. Postharvest Biol. Technol. 1993, 3, 49–56. [Google Scholar] [CrossRef]
- Zhang, P.; Li, X.M.; Wang, J.N.; Wang, B.G. Oxepine-containing diketopiperazine alkaloids from the algal-derived endophytic fungus Paecilomyces variotii EN-291. Helv. Chim. Acta 2015, 98, 800–804. [Google Scholar] [CrossRef]
- Rodrigo, S.; Santamaria, O.; Halecker, S.; Lledó, S.; Stadler, M. Antagonism between Byssochlamys spectabilis (anamorph Paecilomyces variotii) and plant pathogens: Involvement of the bioactive compounds produced by the endophyte. Ann. Appl. Biol. 2017, 171, 464–476. [Google Scholar] [CrossRef]
- Suárez-Estrella, F.; Arcos-Nievas, M.A.; López, M.J.; Vargas-García, M.C.; Moreno, J. Biological control of plant pathogens by microorganisms isolated from agro-industrial composts. Biol. Control 2013, 67, 509–515. [Google Scholar] [CrossRef]
- Arora, K.; Sharma, S.; Krishna, S.B.; Adam, J.K.; Kumar, A. Non-edible Oil cakes as a novel substrate for DPA production and augmenting biocontrol activity of Paecilomyces variotii. Front. Microbiol. 2017, 8, 753. [Google Scholar] [CrossRef] [PubMed]
- Maitlo, S.A.; Rajput, N.A.; Syed, R.N.; Khanzada, M.A.; Rajput, A.Q.; Lodhi, A.M. Microbial control of Fusarium wilt of chickpea caused by Fusarium oxysporum f. sp. ciceris. Pak. J. Bot. 2019, 51, 2261–2268. [Google Scholar] [CrossRef]
- Ramzan, N.; Shahzad, S. Inhibition of in vitro growth of soil-borne pathogens by compost-inhabiting indigenous bacteria and fungi. Pak. J. Bot. 2014, 46, 1093–1099. [Google Scholar]
- Anis, M.; Abbasi, M.W.; Zaki, M.J. Bioefficacy of microbial antagonists against Macrophomina phaseolina on sunflower. Pak. J. Bot. 2010, 42, 2935–2940. [Google Scholar]
- Khanzada, M.; Shah, G.S. In-Vitro evaluation of fungicides, plant extracts and bio-control agents against rice blast pathogen Magnaporthe oryzae couch. Pak. J. Bot. 2012, 44, 1775–1778. [Google Scholar]
- Mohammadi, S.; Soltani, J.; Piri, K. Soilborne and invertebrate pathogenic Paecilomyces species show activity against pathogenic fungi and bacteria. J. Crop Prot. 2016, 5, 377–387. [Google Scholar] [CrossRef]
- Al-Sheikh, H.; Abdelzaher, H.M.A. Isolation of Aspergillus sulphureus, Penicillium islandicum and Paecilomyces variotii from agricultural soil and their biological activity against Pythium spinosum, the damping-off organism of soybean. J. Biol. Sci. 2010, 10, 178–189. [Google Scholar] [CrossRef]
- Amrina, H.; Shahzad, S.; Siddiqui, Z.S. Photochemistry of Luffa cylindrica (L.) Roem under fungal biocontrol interaction. Photosynthetica 2018, 56, 743–749. [Google Scholar] [CrossRef]
- Perveen, Z.; Ramzan, N.; Noreen, N.; Rajpuit, A.Q.; Shahzad, S. In vitro evaluation of biocontrol potential of Paecilomyces species against Sclerotium rolfsii and Pythium aphanidermatum. Int. J. Biol. Biotechnol. 2015, 12, 407–411. [Google Scholar]
- Barakat, K.; Saleh, M. Bioactive betulin produced by marine Paecilomyces WE3-F. J. Appl. Pharm. Sci. 2016, 6, 34–40. [Google Scholar] [CrossRef]
- Liu, J.; Li, F.; Lee, Y.M.; Li, J.L.; Hong, J.K.; Yoon, W.D.; Kim, E.K.; Jung, J.H. An anacardic acid analog from the jellyfish-derived fungus Paecilomyces variotii. Nat. Prod. Sci. 2012, 18, 8–12. [Google Scholar]
- Varma, P.K.; Gandhi, S.K.; Surender, S. Biological control of Alternaria solani, the causal agent of early blight of tomato. J. Biol. Control 2008, 22, 67–72. [Google Scholar]
- Sarfaraz, H.; Tariq, M.; Dawar, S. Utilization of microbial antagonists with Thuja orientalis L. for the control of pathognic fungi. Int. J. Biol. Res. 2017, 5, 15–20. [Google Scholar]
- Lu, C.; Liu, H.; Jiang, D.; Wang, L.; Jiang, Y.; Tang, S.; Hou, X.; Han, X.; Liu, Z.; Zhang, M. Paecilomyces variotii extracts (ZNC) enhance plant immunity and promote plant growth. Plant Soil 2019, 441, 383–397. [Google Scholar] [CrossRef]
- Herrera, I.; Toledo, F.J.; Mioso, R. Enzymes and bioproducts produced by the ascomycete fungus Paecilomyces variotii. J. Appl. Microbiol. 2015, 119, 1455–1466. [Google Scholar] [CrossRef]
- Liu, J.; Li, F.; Kim, E.L.; Li, J.L.; Hong, J.; Bae, K.S.; Chung, H.Y.; Hyung, S.K.; Jung, J.H. Antibacterial polyketides from the jellyfish-derived fungus Paecilomyces variotii. J. Nat. Prod. 2011, 74, 1826–1829. [Google Scholar] [CrossRef]
- Horn, W.S.; Kurtz, M.B.; Liesch, J.M.; Smith, J.L.; Martin, I.; Vicente, F. Antibiotic Eicosenoic Acids and Their Manufacture with Paecilomyces variotii. U.S. Patent US523306, 1993. [Google Scholar]
- Terekhova, L.P.; Trenin, A.S.; Ozerskaya, S.M.; Rudenskaya, Y.A.; Maksimova, T.S.; Katrukha, G.S.; Potapova, N.P.; Kosykh, V.A. B synthesis of ascofuranone by the fungus Paecilomyces variotii Bainier. Микрoбиoлoгия 1997, 66, 611–615. [Google Scholar]
- Martinková, M.; Gonda, J.; Raschmanová, J.Š.; Slaninková, M.; Kuchár, J. Total synthesis of a protected form of sphingofungin E using the (3, 3)-sigmatropic rearrangement of an allylic thiocyanate as the key reaction. Carbohydr. Res. 2010, 345, 2427–2437. [Google Scholar] [CrossRef] [PubMed]
- Degenkolb, T.; Vilcinskas, A. Metabolites from nematophagous fungi and nematicidal natural products from fungi as an alternative for biological control. Part I: Metabolites from nematophagous ascomycetes. Appl. Microbiol. Biotechnol. 2016, 100, 3799–3812. [Google Scholar] [CrossRef] [PubMed]
- Abbas, Z.; Siddiqui, B.S.; Shahzad, S.; Sattar, S.; Begum, S.; Batool, A.; Choudhary, M.I. Lawsozaheer, a new chromone produced by an endophytic fungus Paecilomyces variotii isolated from Lawsonia Alba Lam. inhibits the growth of Staphylococcus aureus. Nat. Prod. Res. 2020, 1–6. [Google Scholar] [CrossRef]
- Kanai, Y.; Fujimaki, T.; Kochi, S.I.; Konno, H.; Kanazawa, S.; Tokumasu, S. Paeciloxazine, a novel nematicidal antibiotic from Paecilomyces sp. J. Antibiot. 2004, 57, 24–28. [Google Scholar] [CrossRef]
- Hegde, V.R.; Silver, J.; Patel, M.; Gullo, V.P.; Puar, M.S.; Das, P.R.; Loebenberg, D. Novel fungal metabolites as cell wall active antifungals. J. Antibiot. 2003, 56, 437–447. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Li, X.M.; Mao, X.X.; Mándi, A.; Kurtán, T.; Wang, B.G. Varioloid A, a new indolyl-6, 10b-dihydro-5aH-[1] benzofuro [2, 3-b] indole derivative from the marine alga-derived endophytic fungus Paecilomyces variotii EN-291. Beilstein J. Org. Chem. 2016, 12, 2012–2018. [Google Scholar] [CrossRef] [PubMed]
- López-Fernández, S.; Campisano, A.; Schulz, B.J.; Steinert, M.; Stadler, M.; Surup, F. Viriditins from Byssochlamys spectabilis, their stereochemistry and biosynthesis. Tetrahedron Lett. 2020, 61, 151446. [Google Scholar] [CrossRef]
- Hyung, J.J.; Kang, H.; Joong, J.J.; Soo, K.Y. Paecilomyces Variotii Extracts for Preventing and Treating Infections Caused by Fish Pathogenic Microorganisms. KR Patent 2013051523, 2013. [Google Scholar]
- Moreno-Gavíra, A.; Diánez, F.; Sánchez-Montesinos, B.; Santos, M. Paecilomyces variotii as a plant-growth promoter in horticulture. Agronomy 2020, 10, 597. [Google Scholar] [CrossRef]
- Santos, M.; Dianez, F.; Gonzalez del Valle, M.; Tello, J.C. Grape marc compost: Microbial studies and suppression of soil-borne mycosis in vegetable seedlings. World J. Microbiol. Biotechnol. 2008, 24, 1493–1505. [Google Scholar] [CrossRef]
- Novak, N.G.; Perez, F.G.; Jones, R.W.; Lawrence, S.D. Detached leaf assays to simplify gene expression studies in potato during infestation by chewing insect Manduca sexta. J. Vis. Exp. 2019, 147. [Google Scholar] [CrossRef]
- Patial, M.; Kumar, J.; Pal, D. Detached leaf assay for evaluating resistance to leaf rust Pst. 104-2 in wheat (Triticum aestivum L.). Indian J. Exp. Biol. 2017, 55, 789–794. [Google Scholar]
- Diánez, F.; Santos, M.; Blanco, R.; Tello, J.C. Fungicide resistance in Botrytis cinerea isolates from strawberry crops in huelva (Southwestern Spain). Phytoparasitica 2002, 30, 529–534. [Google Scholar] [CrossRef]
- Nene, Y.L.; Thapliyal, R.N. Evaluation of Fungicides for Plant Disease Control, 3rd ed.; IBH Publishing Co.: New Dehli, India, 1993; p. 33. [Google Scholar]
- Viñuela, E.; Jacas, J.A.; Marco, V.; Adan, A.; Budia, F. Los efectos de los plaguicidas sobre los organismos beneficiosos en agricultura. Grupo de trabajo de OILB plaguicidas y organismos beneficiosos. I. Insecticidas y acaricidas. Phytoma 1993, 45, 18–25. [Google Scholar]
- Marín, F.; Diánez, F.; Santos, M.; Carretero, F.; Gea, F.J.; Castañeda, C.; Navarro, M.J.; Yau, J.A. Control of Phytophthora capsici and Phytophthora parasitica on pepper (Capsicum annuum L.) with compost teas from different sources, and their effects on plant growth promotion. Phytopathol. Mediterr. 2014, 53, 216–228. [Google Scholar]
- Chen, Y.; Wang, D. Two convenient methods to evaluate soybean for resistance to Sclerotinia sclerotiorum. Plant Dis. 2005, 89, 1268–1272. [Google Scholar] [CrossRef] [PubMed]
- Adebola, M.O.; Amadi, J.E. Antagonistic activities of Paecilomyces and Rhizopus species against the cocoa black pod pathogen (Phytophthora palmivora). Afr. Sci. 2010, 11, 235–239. [Google Scholar]
- Comby, M.; Gacoin, M.; Robineau, M.; Rabenoelina, F.; Ptas, S.; Dupont, J.; Bailliieul, F. Screening of wheat endophytes as biological control agents against Fusarium head blight using two different in vitro tests. Microbiol. Res. 2017, 202, 11–20. [Google Scholar] [CrossRef] [PubMed]
- Besset-Manzoni, Y.; Joly, P.; Brutel, A.; Gerin, F.; Soudière, O.; Langin, T.; Prigent-Combaret, C. Does in vitro selection of biocontrol agents guarantee success in planta? A study case of wheat protection against Fusarium seedling blight by soil bacteria. PLoS ONE 2019, 5, 2–18. [Google Scholar] [CrossRef] [PubMed]
- Mioso, R.; Marante, F.J.T.; De Laguna, I.H.B. The chemical diversity of the ascomycete fungus Paecilomyces variotii. Appl. Biochem. Biotechnol. 2015, 177, 781–791. [Google Scholar] [CrossRef]
- Dai, Z.B.; Wang, X.; Li, G.H. Secondary Metabolites and their bioactivities produced by Paecilomyces. Molecules 2020, 25, 5077. [Google Scholar] [CrossRef] [PubMed]
- Aronson, J.; Pereira, J.S.; Pausas, J.G. Cork Oak Woodlands on the Edge. Ecology, Adaptative Management and Restoration; Island Press: Washington, DC, USA, 2009. [Google Scholar]
- Houbraken, J.; Verweij, P.E.; Rijs, A.J.M.M.; Borman, A.M.; Samson, R.A. Identification of Paecilomyces variotii in clinical samples and settings. J. Clin. Microbiol. 2010, 48, 2754–2761. [Google Scholar] [CrossRef]
- Chamilos, G.; Kontoyiannis, D.P. Voriconazole-resistant disseminated Paecilomyces variotii infection in a neutropenic patient with leukaemia on voriconazole prophylaxis. Clin. Case Rep. 2005, 51, 225–228. [Google Scholar] [CrossRef]
- Aguilar, C.; Pujol, I.; Sala, J.; Guarro, J. Antifungal susceptibilities of Paecilomyces species. Antimicrob. Agents Chemother. 1998, 42, 1601–1604. [Google Scholar] [CrossRef]
- Biango-Daniels, M.N.; Ayer, K.M.; Cox, K.D.; Hodge, K.T. Paecilomyces niveus: Pathogenicity in the orchard and sensitivity to three fungicides. Plant Dis. 2019, 103, 125–131. [Google Scholar] [CrossRef]
- Cantone, F.A.; Vandenberg, J.D. Intraspecific diversity in Paecilomyces fumosoroseus. Mycol. Res. 1998, 102, 209–215. [Google Scholar] [CrossRef]
- Tigano-Milani, M.S.; Honeycutt, R.J.; Lacey, L.A.; Assis, R.; McClelland, M.; Sobral, B.W.S. Genetic variability of Paecilomyces fumosoroseus isolates revealed by molecular markers. J. Invertebr. Pathol. 1995, 65, 274–282. [Google Scholar] [CrossRef]
- Er, M.K.; Gökçe, A. Effects of selected pesticides used against glasshouse tomato pests on colony growth and conidial germination of Paecilomyces fumosoroseus. Biol. Control 2004, 31, 398–404. [Google Scholar] [CrossRef]
- Loureiro, E.; Moino, A.; Arnosti, A.; De Souza, G. Efeito de productos fitossanitarios químicos utilizados em alfase e crisantemo sobre fungus enomopatogenicos. Neotrop. Entomol. 2002, 31, 263–269. [Google Scholar] [CrossRef]
- Gallego-Velásquez, J.; Cardona-Bustos, N.L.; Restrepo-Betancur, F. Compatibility of the entomopathogenic fungus Purpureocillium sp. cepa UdeA0106 with biocontrollers fungi and plant protection products, used on chrysanthemum crops. Act. Biol. 2014, 36, 173–187. [Google Scholar]
- Avery, P.B.; Pick, D.A.; Aristizábal, L.F.; Kerrigan, J.; Powell, C.A.; Rogers, M.E.; Arthurs, S.P. Compatibility of Isaria fumosorosea (Hypocreales: Cordycipitaceae) blastospores with agricultural chemicals used for management of the Asian citrus psyllid, Diaphorina citri (Hemiptera: Liviidae). Insects 2013, 4, 694–711. [Google Scholar] [CrossRef] [PubMed]
- Ondráèková, E.; Seidenglanz, M.; Šafáo, J. Effect of seventeen pesticides on mycelial growth of Akanthomyces, Beauveria, Cordyceps and Purpureocillium strains. Czech Mycol. 2019, 71, 123–135. [Google Scholar] [CrossRef]
- Jacobs, H.; Gray, S.N.; Crump, D.H. Interactions between nematophagous fungi and consequences for their potential as biological agents for the control of potato cyst nematodes. Mycol. Res. 2003, 107, 47–56. [Google Scholar] [CrossRef] [PubMed]
- Diánez, F.; Santos, M.; Carretero, F.; Marín, F. Trichoderma saturnisporum: A new biological control agent. J. Sci. Food Agric. 2016, 96, 1934–1944. [Google Scholar] [CrossRef] [PubMed]
- Taufik, M.; Yusuf, D.N.; Boer, D.; Botek, M. Evaluating the ability of endophyte fungus to control VSD diseases in cocoa seeding. IOP Conf. Ser. Earth Environ. Sci. 2019, 382, 012011. [Google Scholar] [CrossRef]
- Sornakili, A.; Thankappan, S.; Sridharan, A.P.; Nithya, P.; Uthandi, S. Antagonistic fungal endophytes and their metabolite-mediated interactions against phytopathogens in rice. Physiol. Mol. Plant Pathol. 2020, 112, 101525. [Google Scholar] [CrossRef]
- Marín-Chacón, M.A.; Rivera-Coto, G.; Villalobos-Moya, K.; Orozco-Rodríguez, R.; Orozco-Cayasso, S. Evaluación de hongos antagonistas de Botrytis cinerea Pers., en plantaciones de mora, Costa Rica. Agron. Costarric. 2017, 41, 7–18. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Doses (ppm) | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| D1 (0.5 × D2) | D2 | D3 | D4 (1.5 × D3) | |||||||
| FUNGICIDE-ACTION MECHANISMS | D2 | D3 | Growth | Inhib. | Growth | Inhib. | Growth | Inhib. | Growth | Inhib. |
| Thiophanate-methyl 70% (WP) w/w- (1) | 500 | 1000 | 27.8 ± 2.8 a | 60.1% | 13.6 ± 1.7 b | 80.5% | 11.8 ± 1.9 b | 83.0% | 12.6 ± 1.5 b | 81.9% |
| Diethofencarb 25% (WP) w/w- (1) | 1000 | 1500 | 42.4 ± 2.3 a | 41.4% | 44.6 ± 1.3 a | 38.4% | 32.8 ± 2.9 b | 54.7% | 18.2 ± 1.3 c | 74.9% |
| Pencycuron 25% (SC) w/v- (1) | 5000 | 8000 | 58.2 ± 5.8 a | 19.6% | 60.8 ± 4.8 a | 16.0% | 59.2 ± 3.3 a | 18.2% | 63.2 ± 1.6 a | 12.7% |
| Iprodione 50% (SC) w/v- (2) | 1000 | 1500 | 0 ± 0 a | 100.0% | 0 ± 0 a | 100.0% | 0 ± 0 a | 100.0% | 0 ± 0 a | 100.0% |
| Flutriafol 12.5% (SC) w/v- (3) | 2000 | 2500 | 13.5 ± 0.4 a | 78.1% | 12.6 ± 2.1 a | 79.5% | 12.2 ± 1.1 a | 80.2% | 11.8 ± 1.6 a | 80.8% |
| Triadimenol 25% (EC) w/v- (3) | 250 | 500 | 61 ± 8.6 a | 1.0% | 36.8 ± 6 b | 40.3% | 26.8 ± 3 c | 56.5% | 29 ± 1.9 c | 52.9% |
| Myclobutanil 24% (EC) w/v- (3) | 200 | 400 | 17.6 ± 2.4 a | 71.4% | 16 ± 1 a | 74.0% | 15.4 ± 1.1 a,b | 75.0% | 13.2 ± 1.6 b | 78.6% |
| Tetraconazole 12.5% (ME) w/v- (3) | 200 | 400 | 21 ± 1.6 a | 68.7% | 4.4 ± 1.3 b | 93.4% | 0 ± 0 c | 100.0% | 0 ± 0 c | 100.0% |
| Fenhexamid 50% (WG) w/w- (3) | 1500 | 2000 | 14 ± 2.1 a | 77.3% | 9.2 ± 0.4 a | 85.1% | 3 ± 0.7 b | 95.1% | 3.7 ± 0.7 c | 94.0% |
| Pyrimethanil 40% (SC) P/V- (4) | 1500 | 2000 | 0 ± 0 a | 100.0% | 0 ± 0 a | 100.0% | 0 ± 0 a | 100.0% | 0 ± 0 a | 100.0% |
| Azoxystrobin 25% (SC) w/v- (5) | 800 | 1000 | 22.2 ± 3.2 a | 64.0% | 18 ± 2.2 a | 70.8% | 19.4 ± 1.5 a | 68.5% | 19.2 ± 2.8 a | 68.8% |
| Kresoxim-methyl 50% (WG) w/w- (5) | 200 | 500 | 31 ± 1.6 a | 53.7% | 31.4 ± 0.5 a | 53.1% | 26.8 ± 4.9 b | 60.0% | 28 ± 2.1 a,b | 58.2% |
| Etridiazole 48% (EC) w/v- (6) | 2000 | 3000 | 5.6 ± 0.9 a | 92.3% | 1.6 ± 0.5 b | 97.8% | 0 ± 0 c | 100.0% | 0 ± 0 c | 100.0% |
| Propamocarb 60.5% (SL) w/v- (6) | 2500 | 5000 | 56.4 ± 4.9 a | 22.1% | 62.8 ± 6.6 a | 13.3% | 59.2 ± 3.3 a | 18.2% | 64.4 ± 2.5 a | 11.0% |
| Copper hydroxide 35% (WG) w/w- (7) | 2000 | 3000 | 29.2 ± 2.2 a | 52.6% | 20.4 ± 0.9 b | 66.9% | 0 ± 0 c | 100.0% | 0 ± 0 c | 100.0% |
| Copper oxychloride 38% (SC) w/v- (7) | 2000 | 3000 | 0 ± 0 a | 100.0% | 0 ± 0 a | 100.0% | 0 ± 0 a | 100.0% | 0 ± 0 a | 100.0% |
| Mancozeb 80% (WG) w/w- (7) | 2000 | 3000 | 6 ± 1.2 a | 90.3% | 4 ± 0.7 b | 93.5% | 2.6 ± 0.9 c | 95.8% | 2.2 ± 0.4 c | 96.4% |
| Chlorothalonil 50% (SC) w/v- (7) | 2500 | 3000 | 13.6 ± 1.8 a | 77.9% | 13.4 ± 1.8 a | 78.2% | 15.2 ± 1.8 a | 75.3% | 15.4 ± 1.5 a | 75.0% |
| Fosetyl-AL 80% (WG) w/w- (8) | 2500 | 3000 | 52.4 ± 1.8 a | 14.9% | 41.8 ± 1.3 b | 32.1% | 39.2 ± 2.5 c | 36.4% | 35.4 ± 1.8 d | 42.5% |
| Cymoxanil 60% (WG) w/w- (9) | 200 | 300 | 36.2 ± 4 a | 46.0% | 16.2 ± 0.8 b,c | 75.8% | 17 ± 1.6 b | 74.6% | 13.2 ± 1.6 c | 80.3% |
| Cyprodinil 37.5% + Fludioxonil 25% (WG) w/w- (4 + 2) | 600 | 1000 | 0 ± 0 a | 100.0% | 0 ± 0 a | 100.0% | 0 ± 0 a | 100.0% | 0 ± 0 a | 100.0% |
| Folpet 40% + Metalaxyl-M 10% (WP) w/w- (7 + 10) | 2000 | 2500 | 0 ± 0 a | 100.0% | 0 ± 0 a | 100.0% | 0 ± 0 a | 100.0% | 0 ± 0 a | 100.0% |
| Dimethomorph 7.5% + Mancozeb 66.7% (WG) w/w- (11 + 7) | 2000 | 3000 | 0 ± 0 a | 100.0% | 0 ± 0 a | 100.0% | 0 ± 0 a | 100.0% | 0 ± 0 a | 100.0% |
| Benalaxil 6% + Cymoxanil 3.2% + Mancozeb 40% (WP) w/w- (10 + 9 + 7) | 2500 | 3500 | 0 ± 0 a | 100.0% | 0 ± 0 a | 100.0% | 0 ± 0 a | 100.0% | 0 ± 0 a | 100.0% |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moreno-Gavíra, A.; Diánez, F.; Sánchez-Montesinos, B.; Santos, M. Biocontrol Effects of Paecilomyces variotii against Fungal Plant Diseases. J. Fungi 2021, 7, 415. https://doi.org/10.3390/jof7060415
Moreno-Gavíra A, Diánez F, Sánchez-Montesinos B, Santos M. Biocontrol Effects of Paecilomyces variotii against Fungal Plant Diseases. Journal of Fungi. 2021; 7(6):415. https://doi.org/10.3390/jof7060415
Chicago/Turabian StyleMoreno-Gavíra, Alejandro, Fernando Diánez, Brenda Sánchez-Montesinos, and Mila Santos. 2021. "Biocontrol Effects of Paecilomyces variotii against Fungal Plant Diseases" Journal of Fungi 7, no. 6: 415. https://doi.org/10.3390/jof7060415
APA StyleMoreno-Gavíra, A., Diánez, F., Sánchez-Montesinos, B., & Santos, M. (2021). Biocontrol Effects of Paecilomyces variotii against Fungal Plant Diseases. Journal of Fungi, 7(6), 415. https://doi.org/10.3390/jof7060415