A Re-Evaluation of the Relationship between Morphology and Pathogenicity in Candida Species


{kind=link}
Abstract
1. Introduction
2. Morphogenesis in Candida Species
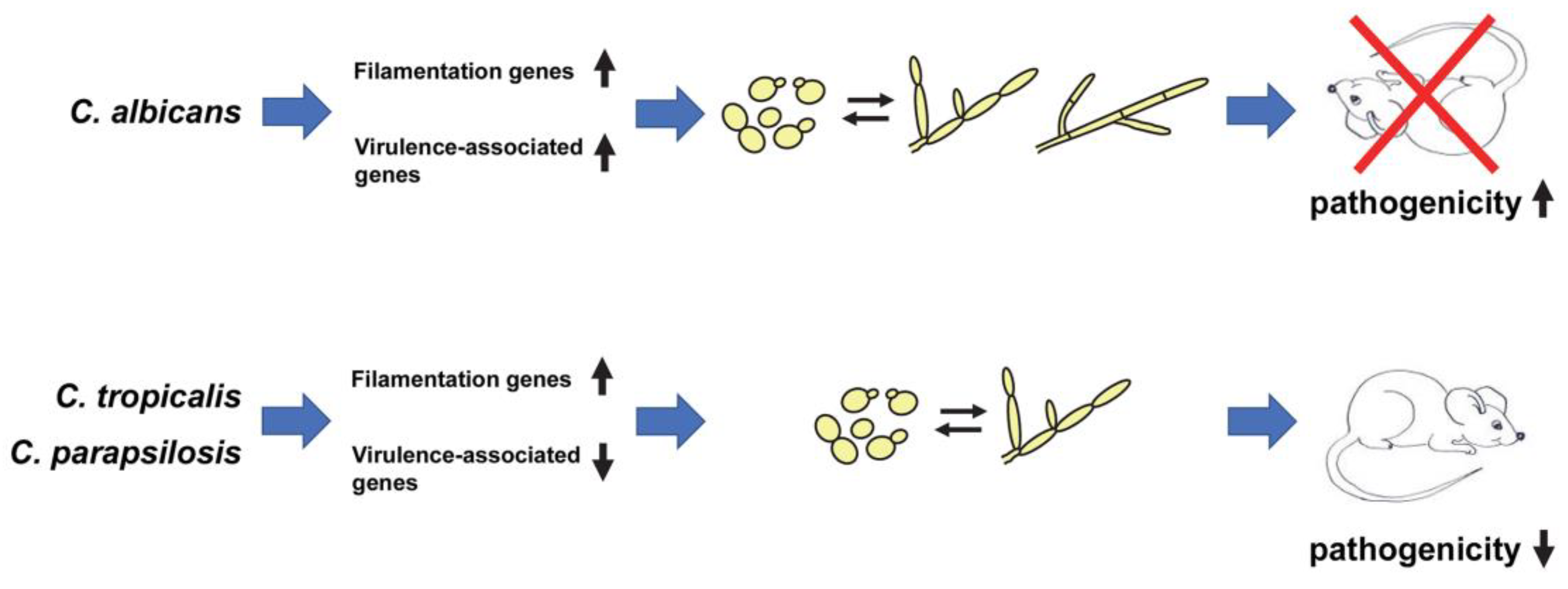
3. Relationship between Morphology and Pathogenicity in C. albicans
4. Relationship between Morphology and Pathogenicity in Non-albicans Candida Species
5. Evolutionary Differences in the Relationship between Morphology and Pathogenicity in C. albicans vs. Non-albicans Candida Species
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Edmond, M.B.; Wallace, S.E.; McClish, D.K.; Pfaller, M.A.; Jones, R.N.; Wenzel, R.P. Nosocomial bloodstream infections in United States hospitals: A three-year analysis. Clin. Infect. Dis. 1999, 29, 239–244. [Google Scholar] [CrossRef] [PubMed]
- Wisplinghoff, H.; Bischoff, T.; Tallent, S.M.; Seifert, H.; Wenzel, R.P.; Edmond, M.B. Nosocomial bloodstream infections in US hospitals: Analysis of 24,179 cases from a prospective nationwide surveillance study. Clin. Infect. Dis. 2004, 39, 309–317. [Google Scholar] [CrossRef] [PubMed]
- Odds, F.C. Candida species and virulence. ASM News 1994, 60, 313–318. [Google Scholar]
- Odds, F.C. Candida and Candidosis, 2nd ed.; Baillière Tindall: London, UK, 1988; p. 468. [Google Scholar]
- Odds, F.C. Pathogenesis of Candida infections. J. Am. Acad. Dermatol. 1994, 31, S2–S5. [Google Scholar] [CrossRef]
- Dupont, P.F. Candida albicans, the opportunist. A cellular and molecular perspective. J. Am. Podiatr. Med. Assoc. 1995, 85, 104–115. [Google Scholar] [CrossRef]
- Weig, M.; Gross, U.; Muhlschlegel, F. Clinical aspects and pathogenesis of Candida infection. Trends Microbiol. 1998, 6, 468–470. [Google Scholar] [CrossRef]
- Perlroth, J.; Choi, B.; Spellberg, B. Nosocomial fungal infections: Epidemiology, diagnosis, and treatment. Med. Mycol. 2007, 45, 321–346. [Google Scholar] [CrossRef]
- Gudlaugsson, O.; Gillespie, S.; Lee, K.; Vande Berg, J.; Hu, J.; Messer, S.; Herwaldt, L.; Pfaller, M.; Diekema, D. Attributable mortality of nosocomial candidemia, revisited. Clin. Infect. Dis. 2003, 37, 1172–1177. [Google Scholar] [CrossRef]
- Picazo, J.J.; Gonzalez-Romo, F.; Candel, F.J. Candidemia in the critically ill patient. Int. J. Antimicrob. Agents 2008, 32 (Suppl. S2), S83–S85. [Google Scholar] [CrossRef]
- Miller, L.G.; Hajjeh, R.A.; Edwards, J.E., Jr. Estimating the cost of nosocomial candidemia in the United States. Clin. Infect. Dis. 2001, 32, 1110. [Google Scholar] [CrossRef]
- McCarty, T.P.; Pappas, P.G. Invasive Candidiasis. Infect. Dis. Clin. N. Am. 2016, 30, 103–124. [Google Scholar] [CrossRef] [PubMed]
- Caceres, D.H.; Forsberg, K.; Welsh, R.M.; Sexton, D.J.; Lockhart, S.R.; Jackson, B.R.; Chiller, T. Candida auris: A review of recommendations for detection and control in healthcare settings. J. Fungi 2019, 5, 111. [Google Scholar] [CrossRef] [PubMed]
- Pfaller, M.A.; Diekema, D.J. Epidemiology of invasive candidiasis: A persistent public health problem. Clin. Microbiol. Rev. 2007, 20, 133–163. [Google Scholar] [CrossRef]
- Patel, P.K.; Erlandsen, J.E.; Kirkpatrick, W.R.; Berg, D.K.; Westbrook, S.D.; Louden, C.; Cornell, J.E.; Thompson, G.R.; Vallor, A.C.; Wickes, B.L.; et al. The changing epidemiology of oropharyngeal candidiasis in patients with HIV/AIDS in the era of antiretroviral therapy. AIDS Res. Treat. 2012, 2012, 262471. [Google Scholar] [CrossRef] [PubMed]
- Moran, G.P.; Sullivan, D.J.; Coleman, D.C. Emergence of non-Candida albicans Candida species as pathogens. In Candida and Candidiasis; Calderone, R.A., Ed.; ASM Press: Washinton, DC, USA, 2002; pp. 37–54. [Google Scholar]
- Calderone, R.A.; Fonzi, W.A. Virulence factors of Candida albicans. Trends Microbiol. 2001, 9, 327–335. [Google Scholar] [CrossRef]
- Kumamoto, C.A.; Vinces, M.D. Contributions of hyphae and hypha-co-regulated genes to Candida albicans virulence. Cell. Microbiol. 2005, 7, 1546–1554. [Google Scholar] [CrossRef]
- Sudbery, P.; Gow, N.; Berman, J. The distinct morphogenic states of Candida albicans. Trends Microbiol. 2004, 12, 317–324. [Google Scholar] [CrossRef]
- Thompson, D.S.; Carlisle, P.L.; Kadosh, D. Coevolution of morphology and virulence in Candida species. Eukaryot. Cell 2011, 10, 1173–1182. [Google Scholar] [CrossRef]
- Gow, N.A.; Henderson, G.; Gooday, G.W. Cytological interrelationships between the cell cycle and duplication cycle of Candida albicans. Microbios 1986, 47, 97–105. [Google Scholar]
- Sudbery, P.E. The germ tubes of Candida albicans hyphae and pseudohyphae show different patterns of septin ring localization. Mol. Microbiol. 2001, 41, 19–31. [Google Scholar] [CrossRef]
- Warenda, A.J.; Konopka, J.B. Septin function in Candida albicans morphogenesis. Mol. Biol. Cell 2002, 13, 2732–2746. [Google Scholar] [CrossRef]
- Sutton, D.A.; Fothergill, A.W.; Rinaldi, M.G. Guide to Clinically Significant Fungi, 1st ed.; Williams and Wilkins: Baltimore, MD, USA, 1998. [Google Scholar]
- Staib, P.; Morschhauser, J. Chlamydospore formation in Candida albicans and Candida dubliniensis—An enigmatic developmental programme. Mycoses 2007, 50, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Chabasse, D.; Bouchara, J.P.; de Gentile, L.; Chennebault, J.M. Candida albicans chlamydospores observed in vivo in a patient with AIDS. Ann. Biol. Clin. 1988, 46, 817–818. [Google Scholar]
- Bartie, K.L.; Williams, D.W.; Wilson, M.J.; Potts, A.J.; Lewis, M.A. Differential invasion of Candida albicans isolates in an in vitro model of oral candidosis. Oral Microbiol. Immunol. 2004, 19, 293–296. [Google Scholar] [CrossRef] [PubMed]
- Lo, H.J.; Kohler, J.R.; DiDomenico, B.; Loebenberg, D.; Cacciapuoti, A.; Fink, G.R. Nonfilamentous C. albicans mutants are avirulent. Cell 1997, 90, 939–949. [Google Scholar] [CrossRef]
- Braun, B.R.; Head, W.S.; Wang, M.X.; Johnson, A.D. Identification and characterization of TUP1-regulated genes in Candida albicans. Genetics 2000, 156, 31–44. [Google Scholar]
- Braun, B.R.; Johnson, A.D. Control of filament formation in Candida albicans by the transcriptional repressor TUP1. Science 1997, 277, 105–109. [Google Scholar] [CrossRef]
- Saville, S.P.; Lazzell, A.L.; Monteagudo, C.; Lopez-Ribot, J.L. Engineered control of cell morphology in vivo reveals distinct roles for yeast and filamentous forms of Candida albicans during infection. Eukaryot. Cell 2003, 2, 1053–1060. [Google Scholar] [CrossRef]
- Carlisle, P.L.; Banerjee, M.; Lazzell, A.; Monteagudo, C.; Lopez-Ribot, J.L.; Kadosh, D. Expression levels of a filament-specific transcriptional regulator are sufficient to determine Candida albicans morphology and virulence. Proc. Natl. Acad. Sci. USA 2009, 106, 599–604. [Google Scholar] [CrossRef]
- Banerjee, M.; Uppuluri, P.; Zhao, X.R.; Carlisle, P.L.; Vipulanandan, G.; Villar, C.C.; Lopez-Ribot, J.L.; Kadosh, D. Expression of UME6, a key regulator of Candida albicans hyphal development, enhances biofilm formation via Hgc1- and Sun41-dependent mechanisms. Eukaryot. Cell 2013, 12, 224–232. [Google Scholar] [CrossRef]
- Zheng, X.; Wang, Y.; Wang, Y. Hgc1, a novel hypha-specific G1 cyclin-related protein regulates Candida albicans hyphal morphogenesis. EMBO J. 2004, 23, 1845–1856. [Google Scholar] [CrossRef] [PubMed]
- O’Meara, T.R.; Veri, A.O.; Ketela, T.; Jiang, B.; Roemer, T.; Cowen, L.E. Global analysis of fungal morphology exposes mechanisms of host cell escape. Nat. Commun. 2015, 6, 6741. [Google Scholar] [CrossRef] [PubMed]
- Noble, S.M.; French, S.; Kohn, L.A.; Chen, V.; Johnson, A.D. Systematic screens of a Candida albicans homozygous deletion library decouple morphogenetic switching and pathogenicity. Nat. Genet. 2010, 42, 590–598. [Google Scholar] [CrossRef]
- Priest, S.J.; Lorenz, M.C. Characterization of virulence-related phenotypes in Candida species of the CUG clade. Eukaryot. Cell 2015, 14, 931–940. [Google Scholar] [CrossRef]
- Bramley, T.A.; Menzies, G.S.; Williams, R.J.; Kinsman, O.S.; Adams, D.J. Binding sites for LH in Candida albicans: Comparison with the mammalian corpus luteum LH receptor. J. Endocrinol. 1991, 130, 177–190. [Google Scholar] [CrossRef]
- Caticha, O.; Grover, S.; Winge, D.; Odell, W.D. Stimulation of Candida albicans transition by human chorionic gonadotrophin and a bacterial protein. Endocr. Res. 1992, 18, 133–143. [Google Scholar] [CrossRef]
- Kinsman, O.S.; Pitblado, K.; Coulson, C.J. Effect of mammalian steroid hormones and luteinizing hormone on the germination of Candida albicans and implications for vaginal candidosis. Mycoses 1988, 31, 617–626. [Google Scholar] [CrossRef]
- Merson-Davies, L.A.; Odds, F.C. A morphology index for characterization of cell shape in Candida albicans. J. Gen. Microbiol. 1989, 135, 3143–3152. [Google Scholar] [CrossRef]
- Brown, A.J. Expression of growth form-specific factors during morphogenesis in Candida albicans. In Candida and Candidiasis; Calderone, R.A., Ed.; ASM Press: Washington, DC, USA, 2002; pp. 87–93. [Google Scholar]
- Lackey, E.; Vipulanandan, G.; Childers, D.S.; Kadosh, D. Comparative evolution of morphological regulatory functions in Candida species. Eukaryot. Cell 2013, 12, 1356–1368. [Google Scholar] [CrossRef]
- Yu, S.; Li, W.; Liu, X.; Che, J.; Wu, Y.; Lu, J. Distinct expression levels of ALS, LIP, and SAP genes in Candida tropicalis with diverse virulent activities. Front. Microbiol. 2016, 7, 1175. [Google Scholar] [CrossRef]
- Silva, S.; Hooper, S.J.; Henriques, M.; Oliveira, R.; Azeredo, J.; Williams, D.W. The role of secreted aspartyl proteinases in Candida tropicalis invasion and damage of oral mucosa. Clin. Microbiol. Infect. 2011, 17, 264–272. [Google Scholar] [CrossRef]
- Jiang, C.; Li, Z.; Zhang, L.; Tian, Y.; Dong, D.; Peng, Y. Significance of hyphae formation in virulence of Candida tropicalis and transcriptomic analysis of hyphal cells. Microbiol. Res. 2016, 192, 65–72. [Google Scholar] [CrossRef]
- Silva, S.; Henriques, M.; Oliveira, R.; Azeredo, J.; Malic, S.; Hooper, S.J.; Williams, D.W. Characterization of Candida parapsilosis infection of an in vitro reconstituted human oral epithelium. Eur. J. Oral Sci. 2009, 117, 669–675. [Google Scholar] [CrossRef]
- Toth, R.; Cabral, V.; Thuer, E.; Bohner, F.; Nemeth, T.; Papp, C.; Nimrichter, L.; Molnar, G.; Vagvolgyi, C.; Gabaldon, T.; et al. Investigation of Candida parapsilosis virulence regulatory factors during host-pathogen interaction. Sci. Rep. 2018, 8, 1346. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, M.; Lazzell, A.L.; Romo, J.A.; Lopez-Ribot, J.L.; Kadosh, D. Filamentation is associated with reduced pathogenicity of multiple non-albicans Candida species. mSphere 2019, 4. [Google Scholar] [CrossRef]
- Kadosh, D. Control of Candida albicans morphology and pathogenicity by post-transcriptional mechanisms. Cell. Mol. Life Sci. 2016, 73, 4265–4278. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.H.; Iyer, K.R.; Pardeshi, L.; Munoz, J.F.; Robbins, N.; Cuomo, C.A.; Wong, K.H.; Cowen, L.E. Genetic analysis of Candida auris implicates Hsp90 in morphogenesis and azole tolerance and Cdr1 in azole resistance. mBio 2019, 10. [Google Scholar] [CrossRef] [PubMed]
- Yue, H.; Bing, J.; Zheng, Q.; Zhang, Y.; Hu, T.; Du, H.; Wang, H.; Huang, G. Filamentation in Candida auris, an emerging fungal pathogen of humans: Passage through the mammalian body induces a heritable phenotypic switch. Emerg. Microbes Infect. 2018, 7, 188. [Google Scholar] [CrossRef]
- Kadosh, D.; Johnson, A.D. Induction of the Candida albicans filamentous growth program by relief of transcriptional repression: A genome-wide analysis. Mol. Biol. Cell 2005, 16, 2903–2912. [Google Scholar] [CrossRef]
- Nantel, A.; Dignard, D.; Bachewich, C.; Harcus, D.; Marcil, A.; Bouin, A.P.; Sensen, C.W.; Hogues, H.; van het Hoog, M.; Gordon, P.; et al. Transcription profiling of Candida albicans cells undergoing the yeast-to-hyphal transition. Mol. Biol. Cell 2002, 13, 3452–3465. [Google Scholar] [CrossRef]
- Chen, Y.V.; Rosli, R.; Fong, S.H.; Sidik, S.M.; Pei, C.P. Histopathological characteristics of experimental Candida tropicalis induced acute systemic candidiasis in BALB/c mice. Int. J. Zool. Res. 2012, 8, 12–22. [Google Scholar]

© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kadosh, D.; Mundodi, V. A Re-Evaluation of the Relationship between Morphology and Pathogenicity in Candida Species. J. Fungi 2020, 6, 13. https://doi.org/10.3390/jof6010013
Kadosh D, Mundodi V. A Re-Evaluation of the Relationship between Morphology and Pathogenicity in Candida Species. Journal of Fungi. 2020; 6(1):13. https://doi.org/10.3390/jof6010013
Chicago/Turabian StyleKadosh, David, and Vasanthakrishna Mundodi. 2020. "A Re-Evaluation of the Relationship between Morphology and Pathogenicity in Candida Species" Journal of Fungi 6, no. 1: 13. https://doi.org/10.3390/jof6010013
APA StyleKadosh, D., & Mundodi, V. (2020). A Re-Evaluation of the Relationship between Morphology and Pathogenicity in Candida Species. Journal of Fungi, 6(1), 13. https://doi.org/10.3390/jof6010013