Antiplasmodial Properties and Cytotoxicity of Endophytic Fungi from Symphonia globulifera (Clusiaceae)

 , and
, and
Abstract
1. Introduction
2. Materials and Methods
2.1. Collection of Plant Material
2.2. Isolation of Endophytic Fungi
2.3. Identification of Endophytic Fungi
2.4. Fermentation and Extraction
2.5. Biological Assays
2.5.1. In Vitro Cultivation of Plasmodium falciparum
2.5.2. Preparation of Stock Solution of Chloroquine and Fungal Extracts
2.5.3. In Vitro Antiplasmodial Assay
2.5.4. Cytotoxicity Assay
3. Results
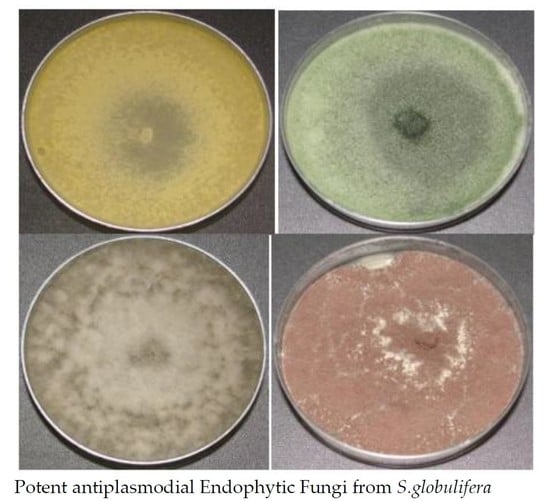
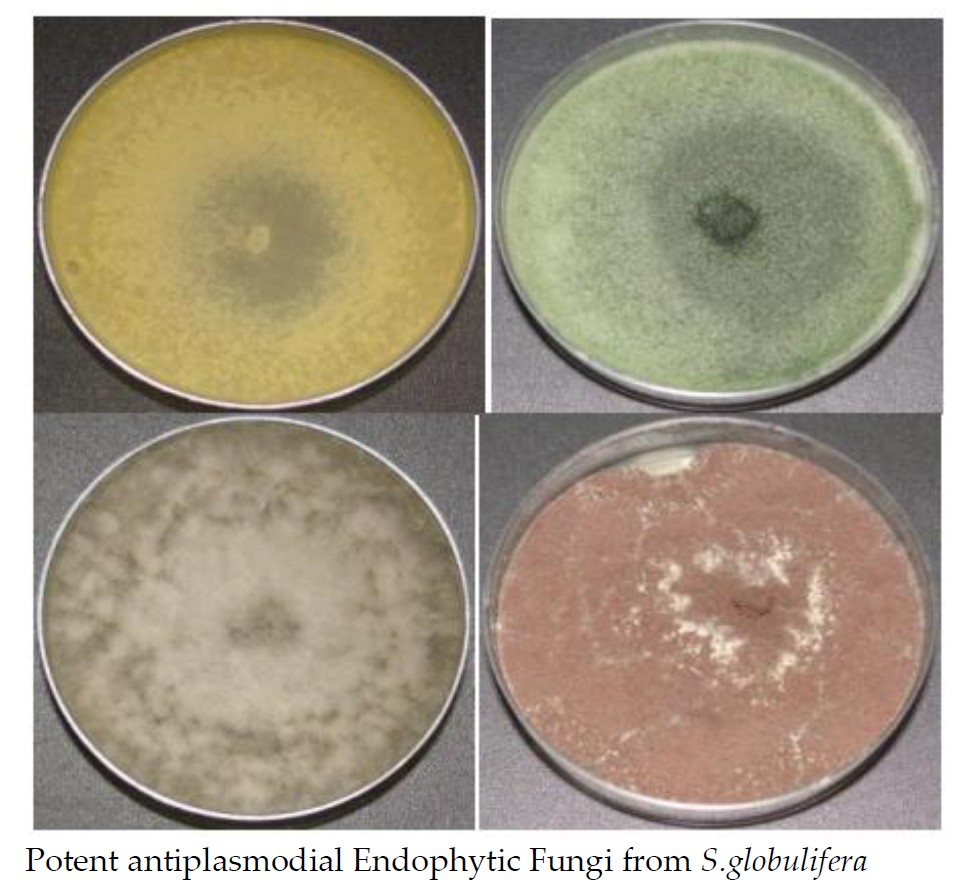
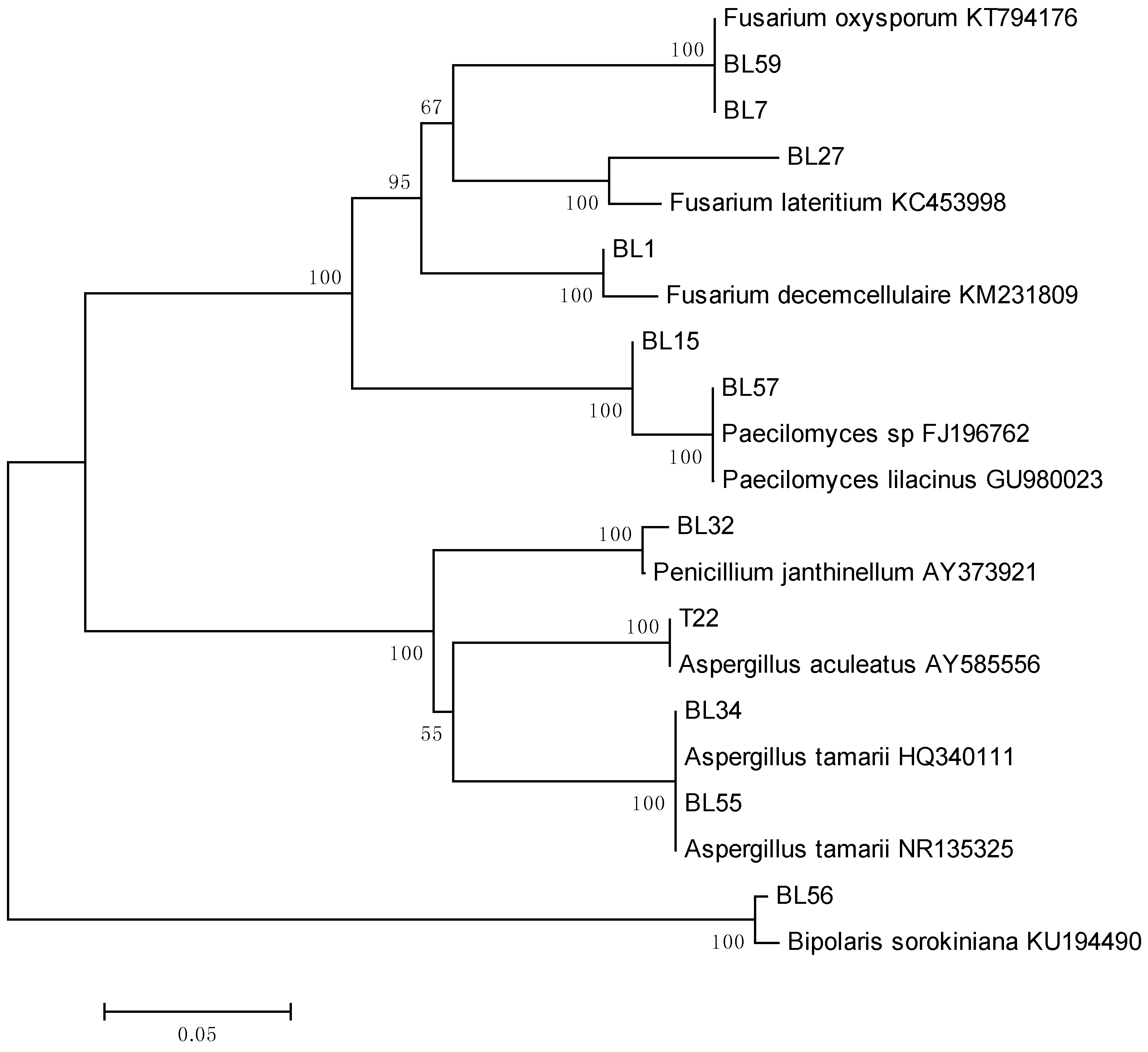
3.1. Isolation and Identification of Fungi
3.2. In Vitro Antiplasmodial Activity of Endophytic Extracts
3.3. Cytotoxicity of Extracts
4. Discussion
5. Conclusions
Author Contributions
Acknowledgments
Conflicts of Interest
References
- World Malaria Report 2017; Licence: CC BY-NC-SA 3.0 IGO; World Health Organization: Geneva, Switzerland, 2017.
- D’Alessandro, U. Existing antimalarial agents and malaria-treatment strategies. Expert Opin. Pharmacother. 2009, 10, 1291–1306. [Google Scholar] [CrossRef] [PubMed]
- Dhar, R.; Zhang, K.; Talwar, G.P.; Garg, S.; Kumar, N. Inhibition of the growth and development of asexual and sexual stages of drug-sensitive and resistant strains of the human malaria parasite Plasmodium falciparum by Neem (Azadirachta indica) fractions. J. Ethnopharmacol. 1998, 61, 31–39. [Google Scholar] [CrossRef]
- Strobel, G.A.; Daisy, B. Bioprospecting for microbial endophytes and their natural products. Microbiol. Mol. Biol. Rev. 2003, 67, 491–502. [Google Scholar] [CrossRef] [PubMed]
- Porras-Alfaro, A.; Bayman, P. Hidden fungi, emergent properties: Endophytes and microbiomes. Ann. Rev. Phytopathol. 2011, 49, 291–315. [Google Scholar] [CrossRef] [PubMed]
- Stierle, A.; Strobel, G.A.; Stierle, D.B. Taxol and taxane production by Taxomyces andreanae, an endophytic fungus of Pacific yew. Science 1993, 260, 214–216. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.W.; Zhang, Y.L.; Lin, F.C.; Hu, Y.Z.; Zhang, C.L. Natural products with antitumor activity from endophytic fungi. Mini Rev. Med. Chem. 2011, 11, 1056–1074. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Han, T.; Ming, Q.; Wu, L.; Rahman, K.; Qin, L. Alkaloids produced by endophytic fungi: A review. Nat. Prod. Commun. 2012, 7, 963–968. [Google Scholar] [PubMed]
- Strobel, G. Harnessing endophytes for industrial microbiology. Curr. Opin. Microbiol. 2006, 9, 240–244. [Google Scholar] [CrossRef] [PubMed]
- Riyaz-Ul-Hassan, S.; Strobel, G.A.; Booth, E.; Knighton, B.; Floerchinger, C.; Sears, J. Modulation of volatile organic compound formation in the Mycodiesel producing endophyte- Hypoxylon sp. C1-4. Microbiology 2012, 158, 464–473. [Google Scholar] [CrossRef]
- Ajibesin, K.; Ekpo, B.; Bala, D.; Essien, E.; Adesanya, S. Ethnobotanical survey of AkwaIbom State of Nigeria. J. Ethnopharmacol. 2008, 115, 387–408. [Google Scholar] [CrossRef] [PubMed]
- Ssegawa, P.; Kasenene, J.M. Medicinal plant diversity and uses in the Sango bay area, Southern Uganda. J. Ethnopharmacol. 2007, 113, 521–540. [Google Scholar] [CrossRef] [PubMed]
- Fromentin, Y.; Cottet, K.; Kritsanida, M.; Miche, S.; Gaboriaud-Kolar, N.; Lallemand, M.-C. Symphoniaglobulifera, a widespread Source of Complex Metabolites with Potent Biological Activities. Planta Med. 2015, 81, 95–107. [Google Scholar] [CrossRef] [PubMed]
- Toghueo, R.M.K.; Zabalgogeazcoa, I.; Vázquez de Aldana, B.R.; Boyom, F.F. Enzymatic activity of endophytic fungi from the medicinal plants Terminalia catappa, Terminalia mantaly and Cananga odorata. S. Afr. J. Bot. 2017, 109, 146–153. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and Direct Sequencing of Fungalribosomal RNA Genes for Phylogenetics. In PCR Protocols. A Guide to Methods and Applications; Innis, M.A., Gelfland, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: Cambridge, MA, USA, 1990; pp. 315–322. [Google Scholar]
- Trager, W.; Jensen, J.B. Human malaria parasites in continuous culture. Science 1976, 193, 673–675. [Google Scholar] [CrossRef] [PubMed]
- Lambros, C.; Vanderberg, J.P. Synchronization of Plasmodium falciparum erythrocytic stages in culture. J. Parasitol. 1979, 65, 418–420. [Google Scholar] [CrossRef] [PubMed]
- Smilkstein, M.; Sriwilaijaroen, N.; Kelly, J.X.; Wilairat, P.; Riscoe, M. Simple and inexpensive fluorescence-based techniquefor high-throughput antimalarial drug screening. Antimicrob. Agents Chemother. 2004, 48, 1803–1806. [Google Scholar] [CrossRef] [PubMed]
- Bagavan, A.; Rahuman, A.A.; Kumaraj, C.; Kaushik, N.K.; Mohanahrishnan, D.; Sahal, D. Antiplasmodial activity of botanical extracts against Plasmodium falciparum. Parasitol. Res. 2011, 108, 1099–1109. [Google Scholar] [CrossRef] [PubMed]
- Mosmann, T. Rapid colorimetric assay for cellular growth and survival: Application to proliferation and cytotoxicity assays. J. Immunol. Methods 1983, 65, 55–63. [Google Scholar] [CrossRef]
- Zou, W.X.; Meng, J.C.; Lu, H.; Chen, G.X.; Shi, G.X.; Zhang, T.Y.; Tan, R.X. Metabolites of Colletotrichum gloeosporioides, an endophytic fungus in Artemisia mongolica. J. Nat. Prod. 2000, 63, 1529–1530. [Google Scholar] [CrossRef] [PubMed]
- Arnold, A.E.; Lutzoni, F. Diversity and host range of foliar fungal endophytes: Are tropical leaves biodiversity hotspots? Ecology 2007, 88, 541–549. [Google Scholar] [CrossRef] [PubMed]
- Sánchez, M.S.; Bills, G.F.; Zabalgogeazcoa, I. The endophytic mycobiota of the grass Dactylis glomerata. Fungal Divers. 2007, 27, 171–195. [Google Scholar]
- Sun, X.; Ding, Q.; Hyde, H.D.; Guo, L.D. Community structure and preference of endophytic fungi of three woody plants in a mixed forest. Fungal Ecol. 2012, 5, 624–632. [Google Scholar] [CrossRef]
- Felber, C.A.; Orlandelli, C.R.; Rhoden, S.A.; Garcia, A.; Costa, A.T.; Azevedo, J.L.; Pamphile, J.A. Bioprospecting foliar endophytic fungi of Vitisla brusca L., cv. Bordô and Concord. Ann. Microbiol. 2015, 26, 765–775. [Google Scholar] [CrossRef]
- Arnold, A.E.; Maynard, Z.; Gilbert, G.S.; Coley, P.D.; Kursar, T.A. Are tropical fungal endophytes hyperdiverse? Ecol. Lett. 2000, 3, 267–274. [Google Scholar] [CrossRef]
- Iwatsuki, M.; Takada, S.; Mori, M.; Ishiyama, A.; Namatame, M.; Nishihara-Tsukashima, A.; Nonaka, K.; Masuma, R.; Otoguro, K.; Shiomi, K.; et al. In vitro and in vivo antimalarial activity of puberulic acid and its new analogs, viticolins A-C, produced by Penicillium sp. FKI-4410. J. Antibiot. 2011, 64, 183–188. [Google Scholar] [CrossRef] [PubMed]
- Kaushik, N.K.; Murali, T.S.; Sahal, D.; Suryanarayanan, T.S. A search for antiplasmodial metabolites among fungal endophytes of terrestrial and marine plants of southern India. Acta Parasitol. 2014, 59, 745–757. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; He, Z.; Xue, J.; Chen, X.; Wei, X. B-Resorcyclic acid lactones from a Paecilomyces fungus. J. Nat. Prod. 2010, 73, 885–889. [Google Scholar] [CrossRef] [PubMed]
- Isaka, M.; Palasarn, S.; Lapanun, S.; Sriklung, K. Paecilodepsipeptide A, an antimalarial and antitumor cyclohexadepsipeptide from the insect pathogenic fungus Paecilomyces cinnamomeus BCC 9616. J. Nat. Prod. 2007, 70, 675–678. [Google Scholar] [CrossRef] [PubMed]
- Nilanonta, C.; Isaka, M.; Kittakoop, P.; Palittapongarnpim, P.; Kamchonwongpaisan, S.; Pittayakhajonwut, D.; Tanticharoen, M.; Thebtaranonth, Y. Antimycobacterial and antiplasmodial cyclodepsipetides fron the insect pathogenic fungus Paecilomyces tenuipes BCC 1614. Planta Med. 2000, 66, 756–758. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
| Endophyte Isolates | % Sequence Homology | Organism with the Highest Sequence Identity, GenBank Acc. No. |
|---|---|---|
| Fusarium decemcellulaire | 98 | Fusarium decemcellulaire KM231809 |
| Paecilomyces sp. | 100 | Paecilomyces sp. FJ196762 |
| Fusarium oxysporum | 100 | Fusarium oxysporum KT794176 |
| Paecilomyces lilacinus | 100 | Paecilomyces lilacinus GU980023 |
| Penicillium janthinellum | 99 | Penicillium janthinellum AY373921 |
| Aspergillus tamarri | 100 | Aspergillus tamarri HQ340111 |
| Penicilium sp. (1) | 99 | Penicillium sp. JN021538 |
| Bipolaris sorokiniana | 94 | Bipolaris sorokiniana KU194490 |
| Fusarium lateritium | 94 | Fusarium lateritium KC453998 |
| Penicillium sp. (2) | 98 | Penicillium sp. JN021538 |
| Aspergillus aculeatus | 100 | Aspergillus aculeatus AY585556 |
| Mucor falcatus | 89 | Mucor falcatus NR103647 |
| Sample | Dry Mass of Extract (g) | IC50 against PfINDO Strain (μg·mL−1 ± SD) | CC50 against HEK239T Cells (μg·mL−1 ± SD) | SI (CC50/IC50) |
|---|---|---|---|---|
| Fusarium decemcellulaire | 10.81 | 2.19 ± 0.07 | 70.91 ± 0.18 | 32.37 |
| Paecilomyces sp. | 14.66 | 0.55 ± 0.01 | 38.81 ± 0.19 | 70.56 |
| Fusarium oxysporum | 8.23 | 1.70 ± 0.22 | 18.24 ± 0.11 | 10.729 |
| Paecilomyces lilacinus | 6.20 | 0.44 ± 0.03 | 10.79 ± 0.2 | 24.52 |
| Penicillium janthinellum | 6.30 | 0.20 ± 0.01 | 9.14 ± 0.12 | 45.7 |
| Aspergillus tamarri | 21.70 | >100 | *** | NA |
| Penicilium sp. | 14.57 | 3.63 ± 0.02 | 3.06 ± 0.36 | 0.842 |
| Bipolaris sorokiniana | 9.89 | 6.10 ± 0.37 | 3.08 ± 0.19 | 0.504 |
| Fusarium lateritium | 11.67 | 6.61± 0.01 | 2.82± 0.23 | 0.426 |
| Penicillium sp.(2) | 16.32 | 9.08 ± 0.13 | 6.28 ± 1.23 | 0.69 |
| Aspergillus aculeatus | 8.74 | 22.08 ± 0.43 | *** | NA |
| Mucor falcatus | 9.22 | 17.51 ± 0.19 | *** | NA |
| Chloroquine | NA | *400 | *** | NA |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ateba, J.E.T.; Toghueo, R.M.K.; Awantu, A.F.; Mba’ning, B.M.; Gohlke, S.; Sahal, D.; Rodrigues-Filho, E.; Tsamo, E.; Boyom, F.F.; Sewald, N.; et al. Antiplasmodial Properties and Cytotoxicity of Endophytic Fungi from Symphonia globulifera (Clusiaceae). J. Fungi 2018, 4, 70. https://doi.org/10.3390/jof4020070
Ateba JET, Toghueo RMK, Awantu AF, Mba’ning BM, Gohlke S, Sahal D, Rodrigues-Filho E, Tsamo E, Boyom FF, Sewald N, et al. Antiplasmodial Properties and Cytotoxicity of Endophytic Fungi from Symphonia globulifera (Clusiaceae). Journal of Fungi. 2018; 4(2):70. https://doi.org/10.3390/jof4020070
Chicago/Turabian StyleAteba, Joël E. T., Rufin M. K. Toghueo, Angelbert F. Awantu, Brice M. Mba’ning, Sebastian Gohlke, Dinkar Sahal, Edson Rodrigues-Filho, Etienne Tsamo, Fabrice F. Boyom, Norbert Sewald, and et al. 2018. "Antiplasmodial Properties and Cytotoxicity of Endophytic Fungi from Symphonia globulifera (Clusiaceae)" Journal of Fungi 4, no. 2: 70. https://doi.org/10.3390/jof4020070
APA StyleAteba, J. E. T., Toghueo, R. M. K., Awantu, A. F., Mba’ning, B. M., Gohlke, S., Sahal, D., Rodrigues-Filho, E., Tsamo, E., Boyom, F. F., Sewald, N., & Lenta, B. N. (2018). Antiplasmodial Properties and Cytotoxicity of Endophytic Fungi from Symphonia globulifera (Clusiaceae). Journal of Fungi, 4(2), 70. https://doi.org/10.3390/jof4020070