
The Growth, Pathogenesis, and Secondary Metabolism of Fusarium verticillioides Are Epigenetically Modulated by Putative Heterochromatin Protein 1 (FvHP1)

 , , , and
, , , and
Abstract
1. Introduction
2. Materials and Methods
3. Results
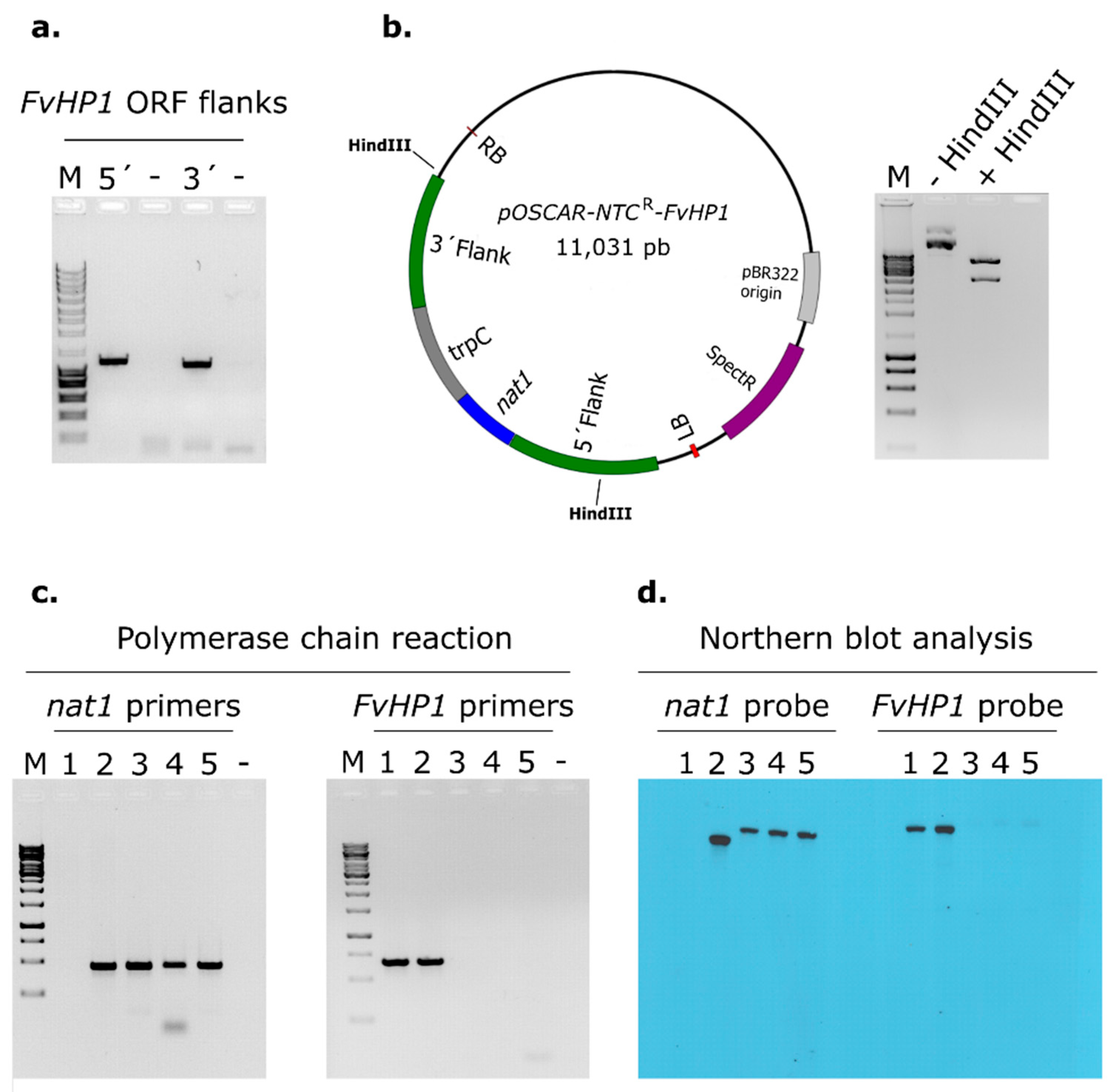
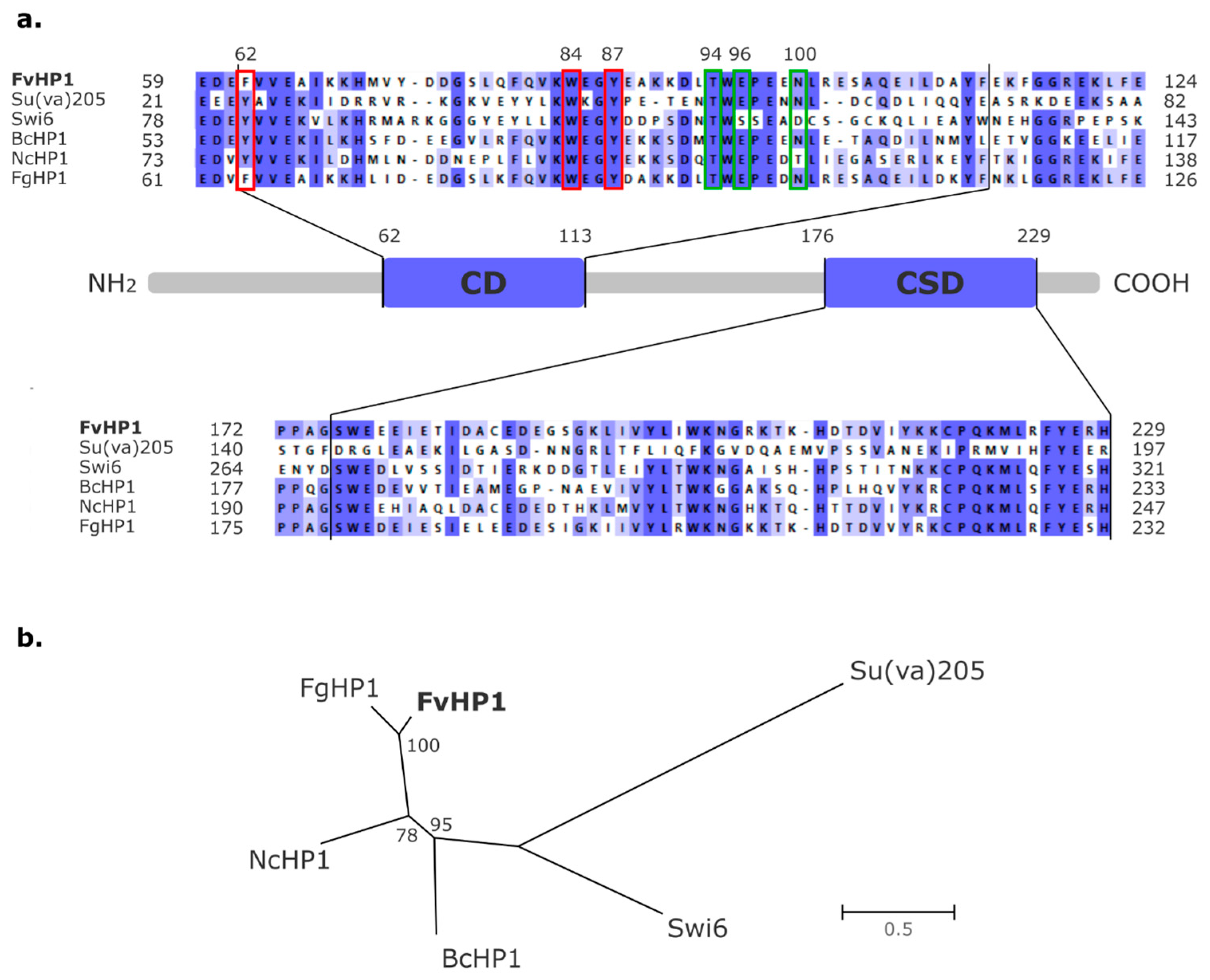
3.1. Identification of a Putative HP1 Protein in Fusarium verticillioides
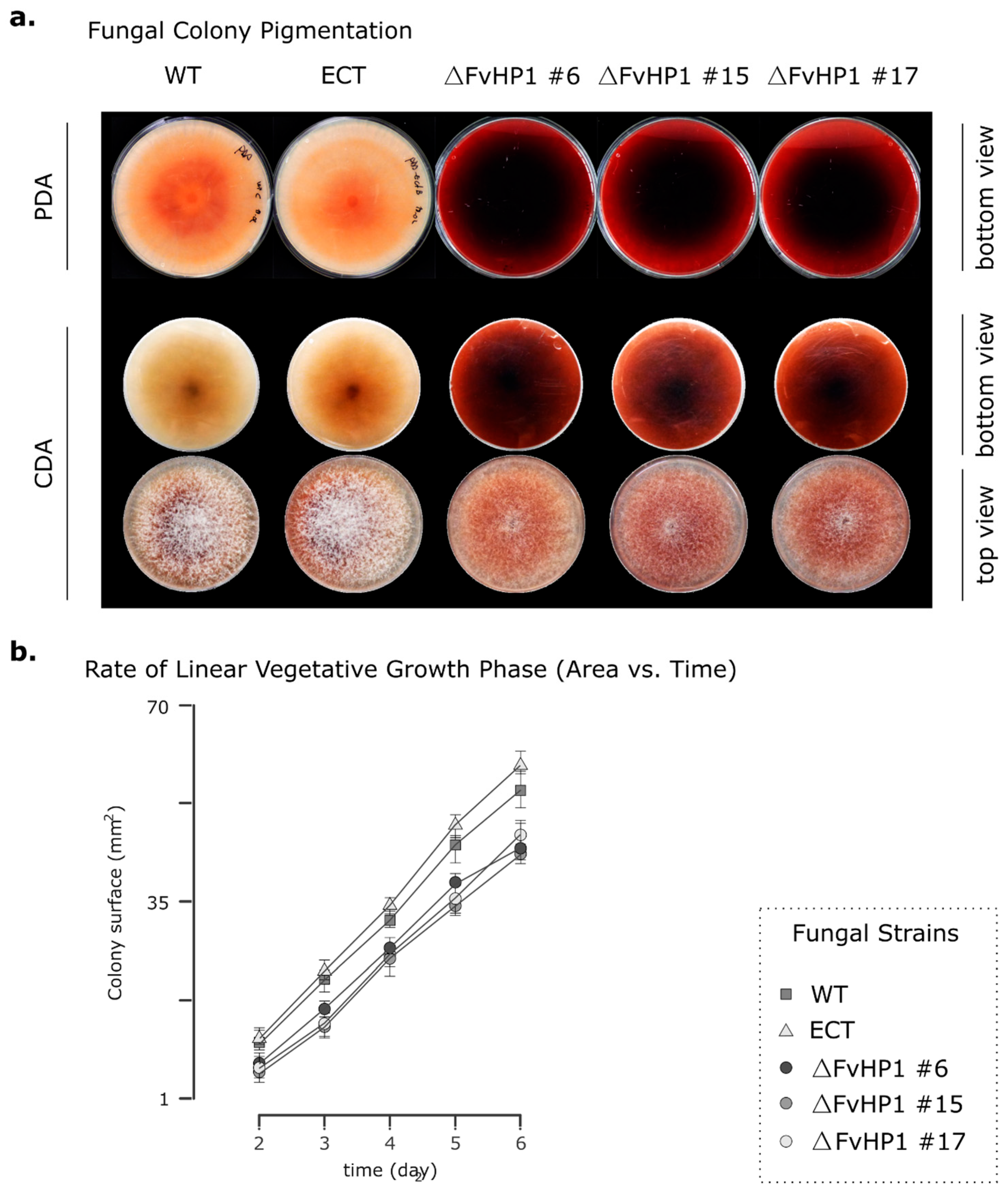
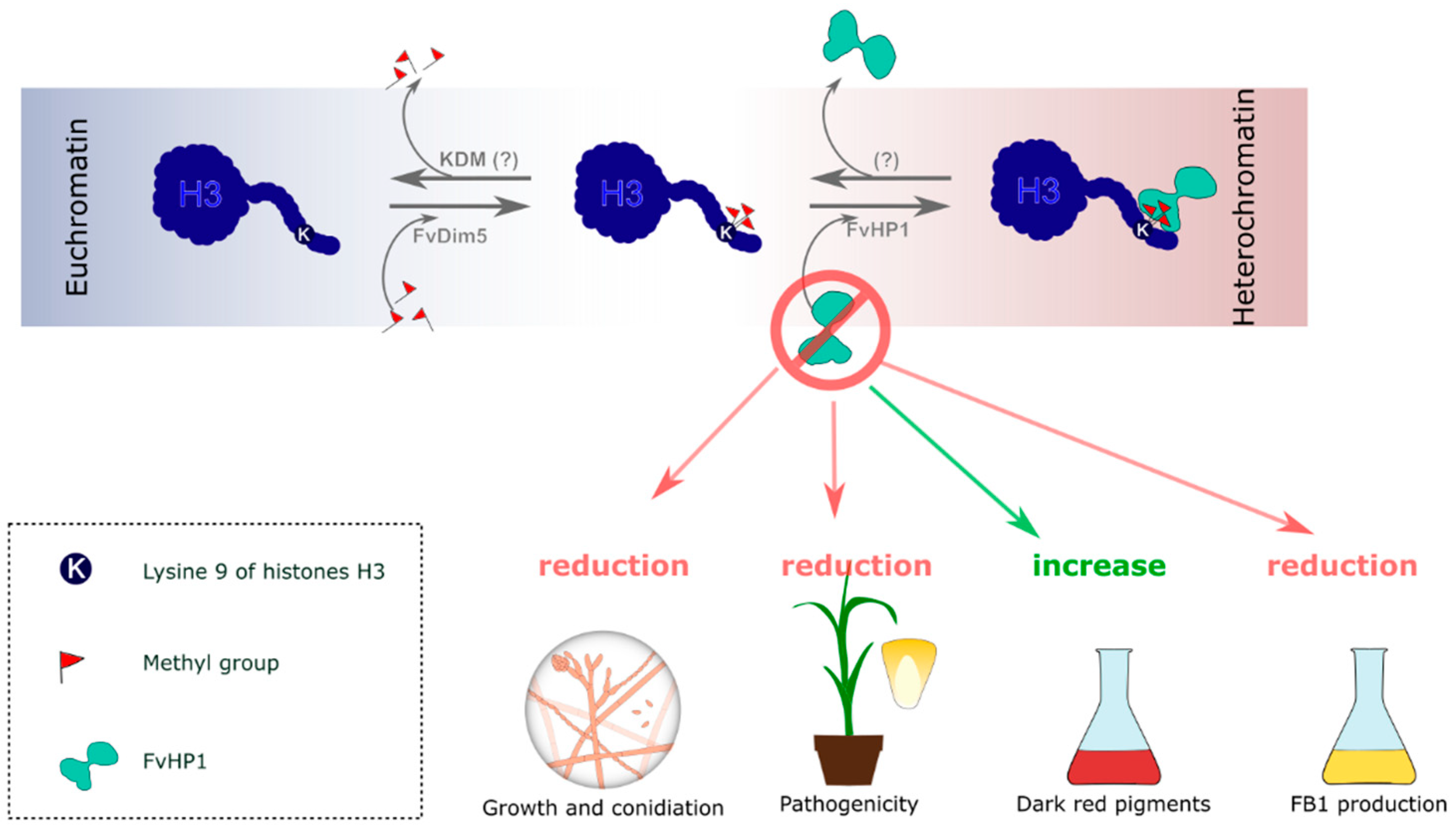
3.2. Deletion of FvHP1 Gene Affects the Growth, Asexual Reproduction, and Pigment Production
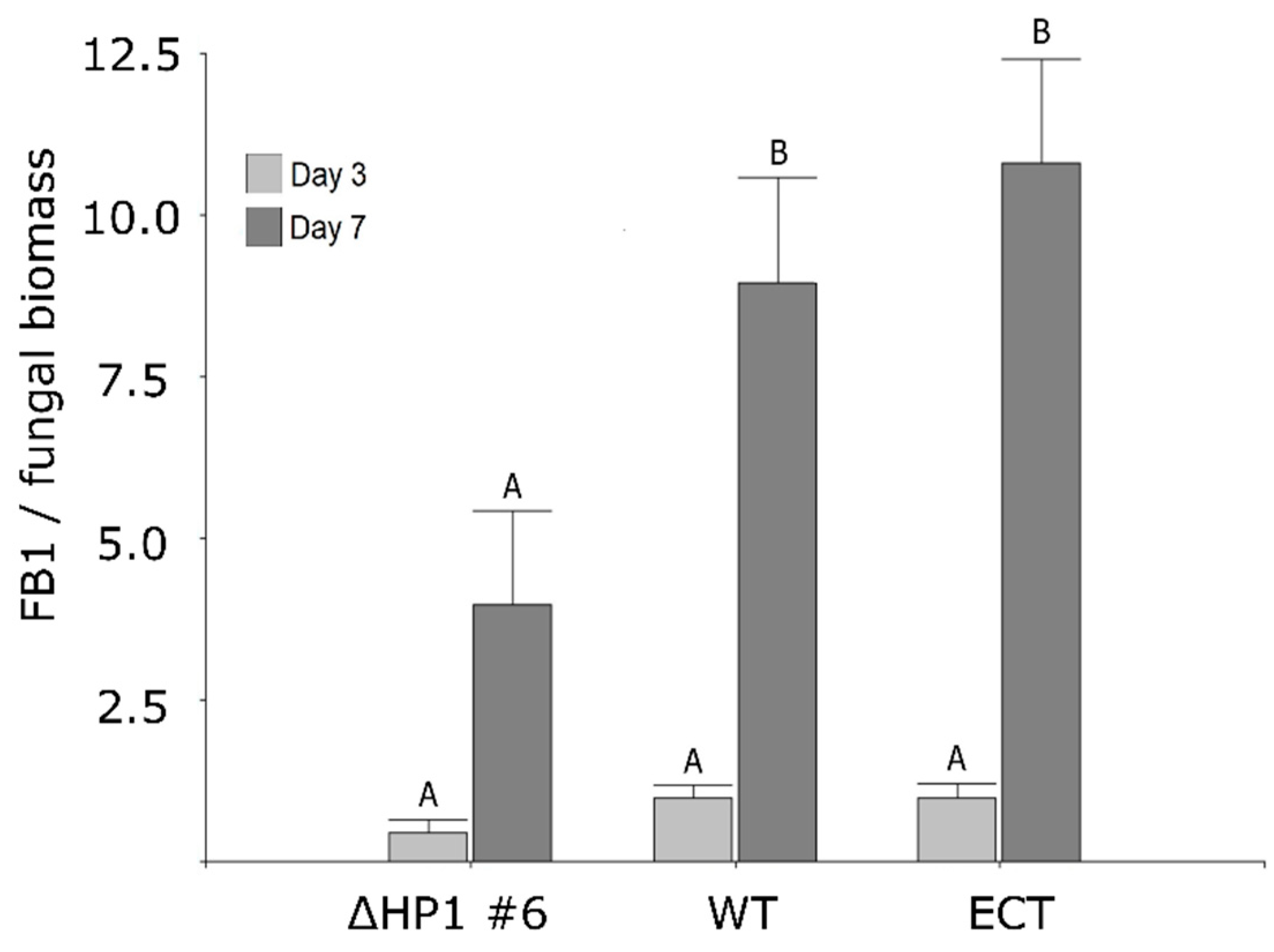
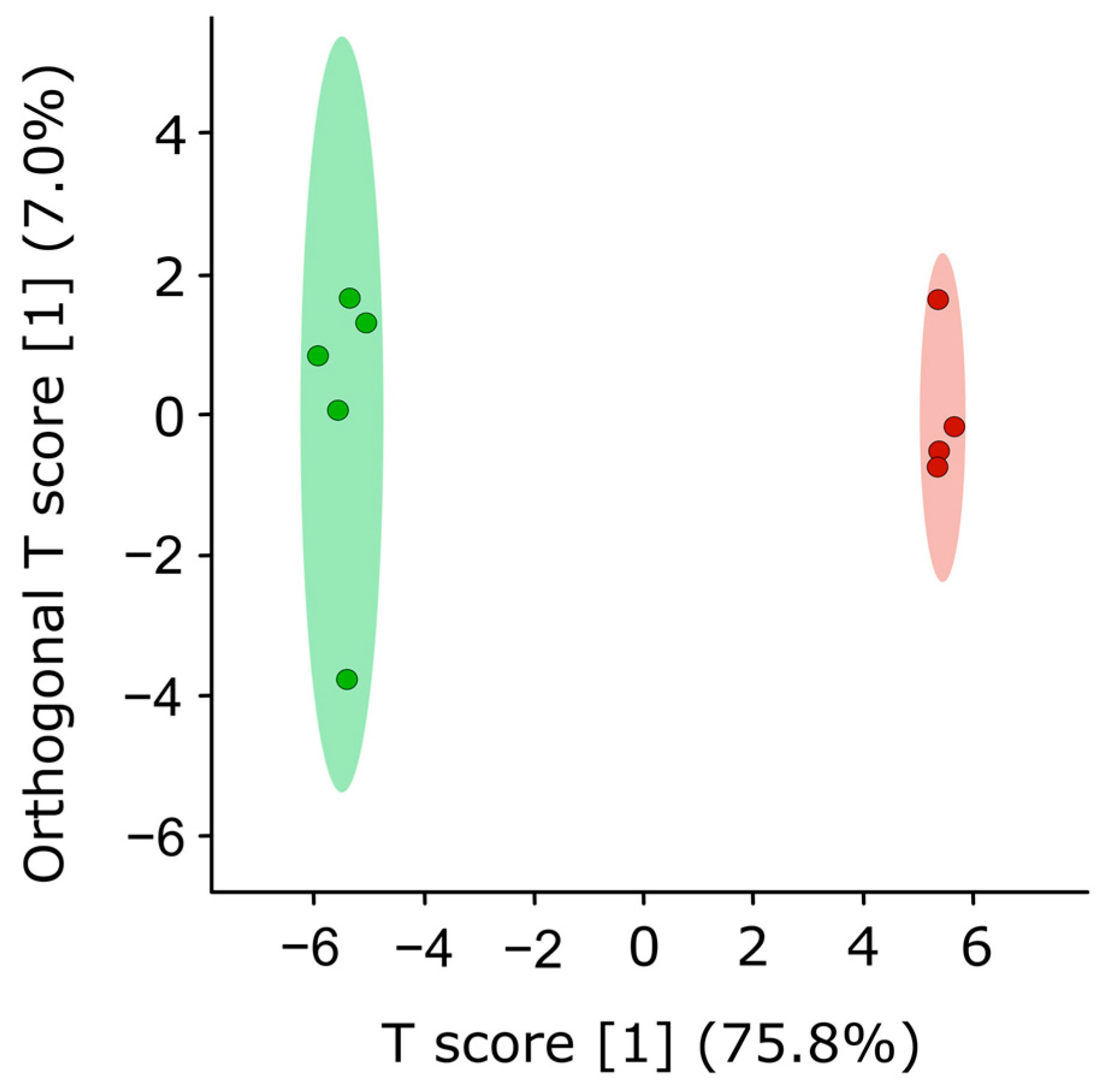
3.3. FvHP1 Protein Is Required for Normal Fumonisin B1 Production
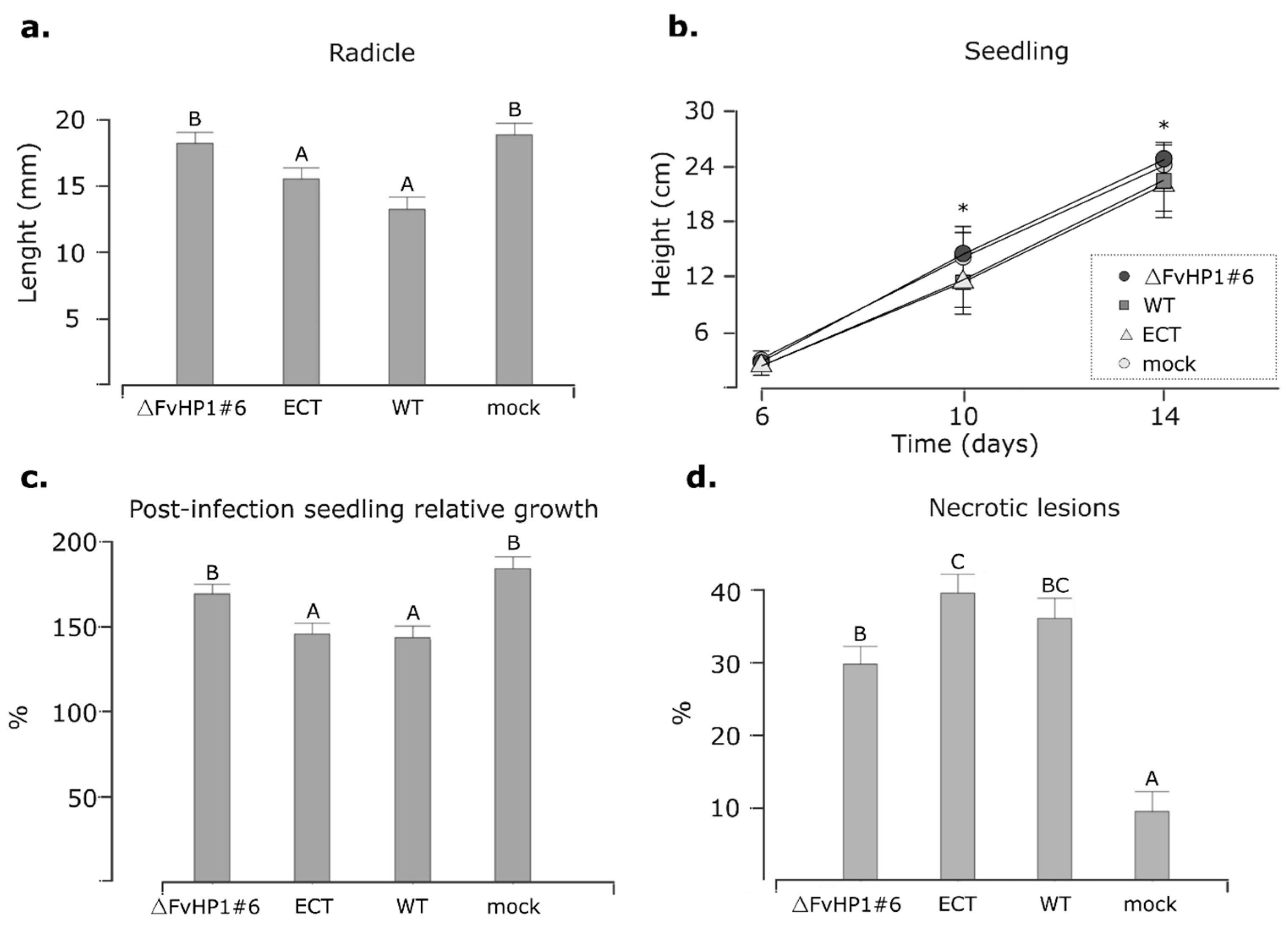
3.4. FvHP1 Plays a Role in the Pathogenicity of F. verticillioides
3.5. FvHP1 Is a Negative Regulator of Naphthoquinone Pigment Synthesis in F. verticillioides
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Blacutt, A.A.; Gold, S.E.; Voss, K.A.; Gao, M.; Glenn, A.E. Fusarium verticillioides: Advancements in Understanding the Toxicity, Virulence, and Niche Adaptations of a Model Mycotoxigenic Pathogen of Maize. Phytopathology 2018, 108, 312–326. [Google Scholar] [CrossRef] [PubMed]
- Bacon, C.W.; Hinton, D.M. Symptomless Endophytic Colonization of Maize by Fusarium moniliforme. Can. J. Bot. 1996, 74, 1195–1202. [Google Scholar] [CrossRef]
- Oren, L.; Ezrati, S.; Cohen, D.; Sharon, A. Early Events in the Fusarium verticillioides—Maize Interaction Characterized by Using a Green Fluorescent Protein-Expressing Transgenic Isolate. Appl. Environ. Microbiol. 2003, 69, 1695–1701. [Google Scholar] [CrossRef]
- Oldenburg, E.; Höppner, F.; Ellner, F.; Weinert, J. Fusarium Diseases of Maize Associated with Mycotoxin Contamination of Agricultural Products Intended to Be Used for Food and Feed. Mycotoxin Res. 2017, 33, 167–182. [Google Scholar] [CrossRef]
- Gai, X.; Dong, H.; Wang, S.; Liu, B.; Zhang, Z.; Li, X.; Gao, Z. Infection Cycle of Maize Stalk Rot and Ear Rot Caused by Fusarium verticillioides. PLoS ONE 2018, 13, e0201588. [Google Scholar] [CrossRef]
- Glenn, A.E. Mycotoxigenic Fusarium Species in Animal Feed. Anim. Feed Sci. Technol. 2007, 137, 213–240. [Google Scholar] [CrossRef]
- Presello, D.A.; Botta, G.; Iglesias, J.; Eyhérabide, G.H. Effect of Disease Severity on Yield and Grain Fumonisin Concentration of Maize Hybrids Inoculated with Fusarium verticillioides. Crop Prot. 2008, 27, 572–576. [Google Scholar] [CrossRef]
- Braun, M.S.; Wink, M. Exposure, Occurrence, and Chemistry of Fumonisins and Their Cryptic Derivatives. Comp. Rev. Food Sci. Food Safe 2018, 17, 769–791. [Google Scholar] [CrossRef] [PubMed]
- Kamle, M.; Mahato, D.K.; Devi, S.; Lee, K.E.; Kang, S.G.; Kumar, P. Fumonisins: Impact on Agriculture, Food, and Human Health and Their Management Strategies. Toxins 2019, 11, 328. [Google Scholar] [CrossRef]
- Nelson, P.E.; Plattner, R.D.; Shackelford, D.D.; Desjardins, A.E. Production of Fumonisins by Fusarium moniliforme Strains from Various Substrates and Geographic Areas. Appl. Environ. Microbiol. 1991, 57, 2410–2412. [Google Scholar] [CrossRef]
- Brown, D.W.; Butchko, R.A.E.; Busman, M.; Proctor, R.H. Identification of Gene Clusters Associated with Fusaric Acid, Fusarin, and Perithecial Pigment Production in Fusarium verticillioides. Fungal Genet. Biol. 2012, 49, 521–532. [Google Scholar] [CrossRef] [PubMed]
- Achimón, F.; Krapacher, C.R.; Jacquat, A.G.; Pizzolitto, R.P.; Zygadlo, J.A. Carbon Sources to Enhance the Biosynthesis of Useful Secondary Metabolites in Fusarium verticillioides Submerged Cultures. World J. Microbiol. Biotechnol. 2021, 37, 78. [Google Scholar] [CrossRef] [PubMed]
- Studt, L.; Wiemann, P.; Kleigrewe, K.; Humpf, H.-U.; Tudzynski, B. Biosynthesis of Fusarubins Accounts for Pigmentation of Fusarium fujikuroi Perithecia. Appl. Environ. Microbiol. 2012, 78, 4468–4480. [Google Scholar] [CrossRef]
- Cam, H.P.; Sugiyama, T.; Chen, E.S.; Chen, X.; FitzGerald, P.C.; Grewal, S.I.S. Comprehensive Analysis of Heterochromatin- and RNAi-Mediated Epigenetic Control of the Fission Yeast Genome. Nat. Genet. 2005, 37, 809–819. [Google Scholar] [CrossRef]
- Palmer, J.M.; Keller, N.P. Secondary Metabolism in Fungi: Does Chromosomal Location Matter? Curr. Opin. Microbiol. 2010, 13, 431–436. [Google Scholar] [CrossRef] [PubMed]
- Strauss, J.; Reyes-Dominguez, Y. Regulation of Secondary Metabolism by Chromatin Structure and Epigenetic Codes. Fungal Genet. Biol. 2011, 48, 62–69. [Google Scholar] [CrossRef]
- Khaldi, N.; Collemare, J.; Lebrun, M.-H.; Wolfe, K.H. Evidence for Horizontal Transfer of a Secondary Metabolite Gene Cluster between Fungi. Genome Biol. 2008, 9, R18. [Google Scholar] [CrossRef]
- Tralamazza, S.M.; Rocha, L.O.; Oggenfuss, U.; Corrêa, B.; Croll, D. Complex Evolutionary Origins of Specialized Metabolite Gene Cluster Diversity among the Plant Pathogenic Fungi of the Fusarium graminearum Species Complex. Genome Biol. Evol. 2019, 11, 3106–3122. [Google Scholar] [CrossRef]
- Proctor, R.H.; Brown, D.W.; Plattner, R.D.; Desjardins, A.E. Co-Expression of 15 Contiguous Genes Delineates a Fumonisin Biosynthetic Gene Cluster in Gibberella Moniliformis. Fungal Genet. Biol. 2003, 38, 237–249. [Google Scholar] [CrossRef]
- Yu, J.-H.; Keller, N. Regulation of Secondary Metabolism in Filamentous Fungi. Annu. Rev. Phytopathol. 2005, 43, 437–458. [Google Scholar] [CrossRef]
- Grewal, S.I.S.; Jia, S. Heterochromatin Revisited. Nat. Rev. Genet. 2007, 8, 35–46. [Google Scholar] [CrossRef] [PubMed]
- Strahl, B.D.; Allis, C.D. The Language of Covalent Histone Modifications. Nature 2000, 403, 41–45. [Google Scholar] [CrossRef] [PubMed]
- O’Kane, C.J.; Hyland, E.M. Yeast Epigenetics: The Inheritance of Histone Modification States. Biosci. Rep. 2019, 39, BSR20182006. [Google Scholar] [CrossRef]
- Eissenberg, J.C.; Elgin, S.C.R. HP1a: A Structural Chromosomal Protein Regulating Transcription. Trends Genet. 2014, 30, 103–110. [Google Scholar] [CrossRef] [PubMed]
- Machida, S.; Takizawa, Y.; Ishimaru, M.; Sugita, Y.; Sekine, S.; Nakayama, J.; Wolf, M.; Kurumizaka, H. Structural Basis of Heterochromatin Formation by Human HP1. Mol. Cell 2018, 69, 385–397.e8. [Google Scholar] [CrossRef]
- Eissenberg, J.C.; Elgin, S.C. The HP1 Protein Family: Getting a Grip on Chromatin. Curr. Opin. Genet. Dev. 2000, 10, 204–210. [Google Scholar] [CrossRef]
- Lomberk, G.; Wallrath, L.; Urrutia, R. The Heterochromatin Protein 1 family. Genome Biol. 2006, 7, 228. [Google Scholar] [CrossRef]
- Kumar, A.; Kono, H. Heterochromatin Protein 1 (HP1): Interactions with Itself and Chromatin Components. Biophys. Rev. 2020, 12, 387–400. [Google Scholar] [CrossRef]
- Eissenberg, J.C.; Morris, G.D.; Reuter, G.; Hartnett, T. The Heterochromatin-Associated Protein HP-1 Is an Essential Protein in Drosophila with Dosage-Dependent Effects on Position-Effect Variegation. Genetics 1992, 131, 345–352. [Google Scholar] [CrossRef]
- Fanti, L.; Berloco, M.; Piacentini, L.; Pimpinelli, S. Chromosomal Distribution of Heterochromatin Protein 1 (HP1) in Drosophila: A Cytological Map of Euchromatic HP1 Binding Sites. Genetica 2003, 117, 135–147. [Google Scholar] [CrossRef]
- Tortora, M.M.C.; Brennan, L.D.; Karpen, G.; Jost, D. HP1-Driven Phase Separation Recapitulates the Thermodynamics and Kinetics of Heterochromatin Condensate Formation. Proc. Natl. Acad. Sci. USA 2023, 120, e2211855120. [Google Scholar] [CrossRef] [PubMed]
- Gu, Q.; Ji, T.; Sun, X.; Huang, H.; Zhang, H.; Lu, X.; Wu, L.; Huo, R.; Wu, H.; Gao, X. Histone H3 Lysine 9 Methyltransferase FvDim5 Regulates Fungal Development, Pathogenicity and Osmotic Stress Responses in Fusarium verticillioides. FEMS Microbiol. Lett. 2017, 364, fnx184. [Google Scholar] [CrossRef] [PubMed]
- Bok, J.W.; Noordermeer, D.; Kale, S.P.; Keller, N.P. Secondary Metabolic Gene Cluster Silencing in Aspergillus nidulans. Mol. Microbiol. 2006, 61, 1636–1645. [Google Scholar] [CrossRef]
- Reyes-Dominguez, Y.; Bok, J.W.; Berger, H.; Shwab, E.K.; Basheer, A.; Gallmetzer, A.; Scazzocchio, C.; Keller, N.; Strauss, J. Heterochromatic Marks Are Associated with the Repression of Secondary Metabolism Clusters in Aspergillus nidulans: Heterochromatin Regulation of Secondary Metabolism. Mol. Microbiol. 2010, 76, 1376–1386. [Google Scholar] [CrossRef] [PubMed]
- Reyes-Dominguez, Y.; Boedi, S.; Sulyok, M.; Wiesenberger, G.; Stoppacher, N.; Krska, R.; Strauss, J. Heterochromatin Influences the Secondary Metabolite Profile in the Plant Pathogen Fusarium graminearum. Fungal Genet. Biol. 2012, 49, 39–47. [Google Scholar] [CrossRef]
- Nishitani, A.; Hiramatsu, K.; Kadooka, C.; Mori, K.; Okutsu, K.; Yoshizaki, Y.; Takamine, K.; Tashiro, K.; Goto, M.; Tamaki, H.; et al. Expression of Heterochromatin Protein 1 Affects Citric Acid Production in Aspergillus luchuensis Mut. kawachii. J. Biosci. Bioeng. 2023, 136, 443–451. [Google Scholar] [CrossRef]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic Local Alignment Search Tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Marchler-Bauer, A.; Bryant, S.H. CD-Search: Protein Domain Annotations on the Fly. Nucleic Acids Res. 2004, 32, W327–W331. [Google Scholar] [CrossRef]
- Pei, J.; Grishin, N.V. PROMALS: Towards Accurate Multiple Sequence Alignments of Distantly Related Proteins. Bioinformatics 2007, 23, 802–808. [Google Scholar] [CrossRef]
- Pei, J.; Kim, B.-H.; Grishin, N.V. PROMALS3D: A Tool for Multiple Protein Sequence and Structure Alignments. Nucleic Acids Res. 2008, 36, 2295–2300. [Google Scholar] [CrossRef]
- Nguyen, L.-T.; Schmidt, H.A.; Von Haeseler, A.; Minh, B.Q. IQ-TREE: A Fast and Effective Stochastic Algorithm for Estimating Maximum-Likelihood Phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.K.F.; Von Haeseler, A.; Jermiin, L.S. ModelFinder: Fast Model Selection for Accurate Phylogenetic Estimates. Nat. Methods 2017, 14, 587–589. [Google Scholar] [CrossRef]
- Okonechnikov, K.; Golosova, O.; Fursov, M.; the UGENE team. Unipro UGENE: A Unified Bioinformatics Toolkit. Bioinformatics 2012, 28, 1166–1167. [Google Scholar] [CrossRef]
- Paz, Z.; García-Pedrajas, M.D.; Andrews, D.L.; Klosterman, S.J.; Baeza-Montañez, L.; Gold, S.E. One Step Construction of Agrobacterium-Recombination-Ready-Plasmids (OSCAR), an Efficient and Robust Tool for ATMT Based Gene Deletion Construction in Fungi. Fungal Genet. Biol. 2011, 48, 677–684. [Google Scholar] [CrossRef]
- Mullins, E.D.; Chen, X.; Romaine, P.; Raina, R.; Geiser, D.M.; Kang, S. Agrobacterium-Mediated Transformation of Fusarium oxysporum: An Efficient Tool for Insertional Mutagenesis and Gene Transfer. Phytopathology 2001, 91, 173–180. [Google Scholar] [CrossRef]
- Ochoa-Velasco, C.E.; Navarro-Cruz, A.R.; Vera-López, O.; Palou, E.; Avila-Sosa, R. Growth Modeling to Control (in Vitro) Fusarium verticillioides and Rhizopus stolonifer with Thymol and Carvacrol. Rev. Argent. Microbiol. 2018, 50, 70–74. [Google Scholar] [CrossRef] [PubMed]
- Di Rienzo, J.A.; Casanoves, F.; Balzarini, M.G.; Gonzalez, L.; Tablada, M.; Robledo, C.W. InfoStat Versión 2020. Centro de Transferencia InfoStat, FCA, Universidad Nacional de Córdoba, Argentina. 2020. Available online: http://www.infostat.com.ar (accessed on 1 July 2020).
- Bacon, C.W. A Corn Seedling Assay for Resistance to Fusarium moniliforme. Plant Dis. 1994, 78, 302. [Google Scholar] [CrossRef]
- Zörb, C.; Geilfus, C.-M.; Mühling, K.H.; Ludwig-Müller, J. The Influence of Salt Stress on ABA and Auxin Concentrations in Two Maize Cultivars Differing in Salt Resistance. J. Plant Physiol. 2013, 170, 220–224. [Google Scholar] [CrossRef]
- Shephard, G.S.; Sydenham, E.W.; Thiel, P.G.; Gelderblom, W.C.A. Quantitative determination of fumonisins B1 and B2 by high-performance liquid chromatography with fluorescence detection. J. Liq. Chromatogr. 1990, 13, 2077–2087. [Google Scholar] [CrossRef]
- Eissenberg, J.C.; James, T.C.; Foster-Hartnett, D.M.; Hartnett, T.; Ngan, V.; Elgin, S.C. Mutation in a Heterochromatin-Specific Chromosomal Protein Is Associated with Suppression of Position-Effect Variegation in Drosophila Melanogaster. Proc. Natl. Acad. Sci. USA 1990, 87, 9923–9927. [Google Scholar] [CrossRef] [PubMed]
- Elgin, S.C.R.; Reuter, G. Position-Effect Variegation, Heterochromatin Formation, and Gene Silencing in Drosophila. Cold Spring Harb. Perspect. Biol. 2013, 5, a017780. [Google Scholar] [CrossRef] [PubMed]
- Lorentz, A.; Ostermann, K.; Fleck, O.; Schmidt, H. Switching Gene Swi6, Involved in Repression of Silent Mating-Type Loci in Fission Yeast, Encodes a Homologue of Chromatin-Associated Proteins from Drosophila and Mammals. Gene 1994, 143, 139–143. [Google Scholar] [CrossRef]
- Zhang, X.; Liu, X.; Zhao, Y.; Cheng, J.; Xie, J.; Fu, Y.; Jiang, D.; Chen, T. Histone H3 Lysine 9 Methyltransferase DIM5 Is Required for the Development and Virulence of Botrytis cinerea. Front. Microbiol. 2016, 7. [Google Scholar] [CrossRef]
- Freitag, M.; Hickey, P.C.; Khlafallah, T.K.; Read, N.D.; Selker, E.U. HP1 Is Essential for DNA Methylation in Neurospora. Mol. Cell 2004, 13, 427–434. [Google Scholar] [CrossRef]
- Nielsen, P.R.; Nietlispach, D.; Mott, H.R.; Callaghan, J.; Bannister, A.; Kouzarides, T.; Murzin, A.G.; Murzina, N.V.; Laue, E.D. Structure of the HP1 Chromodomain Bound to Histone H3 Methylated at Lysine 9. Nature 2002, 416, 103–107. [Google Scholar] [CrossRef]
- Bacon, C.W.; Yates, I.E.; Hinton, D.M.; Meredith, F. Biological Control of Fusarium moniliforme in Maize. Environ. Health Perspect. 2001, 109, 325–332. [Google Scholar] [CrossRef]
- Ramos, M.; Ghosson, H.; Raviglione, D.; Bertrand, C.; Salvia, M.-V. Untargeted Metabolomics as a Tool to Monitor Biocontrol Product Residues’ Fate on Field-Treated Prunus Persica. Sci. Total Environ. 2022, 807, 150717. [Google Scholar] [CrossRef] [PubMed]
- Sumner, L.W.; Amberg, A.; Barrett, D.; Beale, M.H.; Beger, R.; Daykin, C.A.; Fan, T.W.-M.; Fiehn, O.; Goodacre, R.; Griffin, J.L.; et al. Proposed Minimum Reporting Standards for Chemical Analysis: Chemical Analysis Working Group (CAWG) Metabolomics Standards Initiative (MSI). Metabolomics 2007, 3, 211–221. [Google Scholar] [CrossRef]
- Ma, L.-J.; van der Does, H.C.; Borkovich, K.A.; Coleman, J.J.; Daboussi, M.-J.; Di Pietro, A.; Dufresne, M.; Freitag, M.; Grabherr, M.; Henrissat, B.; et al. Comparative Genomics Reveals Mobile Pathogenicity Chromosomes in Fusarium. Nature 2010, 464, 367–373. [Google Scholar] [CrossRef]
- Watanabe, M.; Yonezawa, T.; Lee, K.; Kumagai, S.; Sugita-Konishi, Y.; Goto, K.; Hara-Kudo, Y. Molecular Phylogeny of the Higher and Lower Taxonomy of the Fusariumgenus and Differences in the Evolutionary Histories of Multiple Genes. BMC Evol. Biol. 2011, 11, 322. [Google Scholar] [CrossRef] [PubMed]
- Casale, A.M.; Cappucci, U.; Piacentini, L. Unravelling HP1 Functions: Post-Transcriptional Regulation of Stem Cell Fate. Chromosoma 2021, 130, 103–111. [Google Scholar] [CrossRef] [PubMed]
- Chujo, T.; Lukito, Y.; Eaton, C.J.; Dupont, P.-Y.; Johnson, L.J.; Winter, D.; Cox, M.P.; Scott, B. Complex Epigenetic Regulation of Alkaloid Biosynthesis and Host Interaction by Heterochromatin Protein I in a Fungal Endophyte-Plant Symbiosis. Fungal Genet. Biol. 2019, 125, 71–83. [Google Scholar] [CrossRef]
- Lanubile, A.; Ferrarini, A.; Maschietto, V.; Delledonne, M.; Marocco, A.; Bellin, D. Functional Genomic Analysis of Constitutive and Inducible Defense Responses to Fusarium verticillioides Infection in Maize Genotypes with Contrasting Ear Rot Resistance. BMC Genom. 2014, 15, 710. [Google Scholar] [CrossRef]
- Glenn, A.E.; Zitomer, N.C.; Zimeri, A.M.; Williams, L.D.; Riley, R.T.; Proctor, R.H. Transformation-Mediated Complementation of a FUM Gene Cluster Deletion in Fusarium verticillioides Restores Both Fumonisin Production and Pathogenicity on Maize Seedlings. Mol. Plant-Microbe Interact. 2008, 21, 87–97. [Google Scholar] [CrossRef] [PubMed]
- Brown, D.W.; Busman, M.; Proctor, R.H. Fusarium verticillioides SGE1 Is Required for Full Virulence and Regulates Expression of Protein Effector and Secondary Metabolite Biosynthetic Genes. Mol. Plant-Microbe Interact. 2014, 27, 809–823. [Google Scholar] [CrossRef]
- Arias, S.L.; Mary, V.S.; Otaiza, S.N.; Wunderlin, D.A.; Rubinstein, H.R.; Theumer, M.G. Toxin Distribution and Sphingoid Base Imbalances in Fusarium verticillioides-Infected and Fumonisin B1-Watered Maize Seedlings. Phytochemistry 2016, 125, 54–64. [Google Scholar] [CrossRef]
- Otaiza-González, S.N.; Mary, V.S.; Arias, S.L.; Bertrand, L.; Velez, P.A.; Rodriguez, M.G.; Rubinstein, H.R.; Theumer, M.G. Cell Death Induced by Fumonisin B1 in Two Maize Hybrids: Correlation with Oxidative Status Biomarkers and Salicylic and Jasmonic Acids Imbalances. Eur. J. Plant Pathol. 2022, 163, 203–221. [Google Scholar] [CrossRef]
- Visentin, I.; Montis, V.; Döll, K.; Alabouvette, C.; Tamietti, G.; Karlovsky, P.; Cardinale, F. Transcription of Genes in the Biosynthetic Pathway for Fumonisin Mycotoxins Is Epigenetically and Differentially Regulated in the Fungal Maize Pathogen Fusarium verticillioides. Eukaryot. Cell 2012, 11, 252–259. [Google Scholar] [CrossRef]
- Picot, A.; Barreau, C.; Pinson-Gadais, L.; Caron, D.; Lannou, C.; Richard-Forget, F. Factors of the Fusarium verticillioides -Maize Environment Modulating Fumonisin Production. Crit. Rev. Microbiol. 2010, 36, 221–231. [Google Scholar] [CrossRef]
- Flaherty, J.E.; Pirttilä, A.M.; Bluhm, B.H.; Woloshuk, C.P. PAC1, a pH-Regulatory Gene from Fusarium verticillioides. Appl. Environ. Microbiol. 2003, 69, 5222–5227. [Google Scholar] [CrossRef] [PubMed]
- Wiemann, P.; Willmann, A.; Straeten, M.; Kleigrewe, K.; Beyer, M.; Humpf, H.; Tudzynski, B. Biosynthesis of the Red Pigment Bikaverin in Fusarium fujikuroi: Genes, Their Function and Regulation. Mol. Microbiol. 2009, 72, 931–946. [Google Scholar] [CrossRef] [PubMed]
- Frandsen, R.J.N.; Rasmussen, S.A.; Knudsen, P.B.; Uhlig, S.; Petersen, D.; Lysøe, E.; Gotfredsen, C.H.; Giese, H.; Larsen, T.O. Black Perithecial Pigmentation in Fusarium Species Is Due to the Accumulation of 5-Deoxybostrycoidin-Based Melanin. Sci. Rep. 2016, 6, 26206. [Google Scholar] [CrossRef] [PubMed]
- Proctor, R.H.; Butchko, R.A.E.; Brown, D.W.; Moretti, A. Functional Characterization, Sequence Comparisons and Distribution of a Polyketide Synthase Gene Required for Perithecial Pigmentation in Some Fusarium Species. Food Addit. Contam. 2007, 24, 1076–1087. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Yu, W.; Lu, Y.; Wu, Y.; Ouyang, Z.; Tu, Y.; He, B. Epigenetic Regulation of Fungal Secondary Metabolism. J. Fungi 2024, 10, 648. [Google Scholar] [CrossRef]
- Lamka, G.F.; Harder, A.M.; Sundaram, M.; Schwartz, T.S.; Christie, M.R.; DeWoody, J.A.; Willoughby, J.R. Epigenetics in Ecology, Evolution, and Conservation. Front. Ecol. Evol. 2022, 10, 871791. [Google Scholar] [CrossRef]
- Herman, J.J.; Sultan, S.E.; Horgan-Kobelski, T.; Riggs, C. Adaptive Transgenerational Plasticity in an Annual Plant: Grandparental and Parental Drought Stress Enhance Performance of Seedlings in Dry Soil. Integr. Comp. Biol. 2012, 52, 77–88. [Google Scholar] [CrossRef]
- Sakashita, A.; Ooga, M.; Otsuka, K.; Maezawa, S.; Takeuchi, C.; Wakayama, S.; Wakayama, T.; Namekawa, S.H. Polycomb Protein SCML2 Mediates Paternal Epigenetic Inheritance through Sperm Chromatin. Nucleic Acids Res. 2023, 51, 6668–6683. [Google Scholar] [CrossRef]
- Reardon, R.M.; Walsh, A.K.; Larsen, C.I.; Schmidberger, L.H.; Morrow, L.A.; Thompson, A.E.; Wellik, I.M.; Thompson, J.S. An Epigenetically Inherited UV Hyper-Resistance Phenotype in Saccharomyces Cerevisiae. Epigenetics Chromatin 2022, 15, 31. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Lag Phase (95% CI) 1 | Growth Rate 2 | Colony Size at 5 Days of Incubation (mm2) 3 | Conidiation (Conidia/mL) 4 |
|---|---|---|---|---|
| ∆FvHP1 #6 | 38 (30–41) | 412 ± 7 a | 1841 ± 110 a | 4 ± 2 × 107 a |
| ECT | 24 (19–27) | 466 ± 7 b | 2695 ± 104 b | 9 ± 1 × 107 b |
| WT | 23 (15–27) | 490 ± 9 b | 2477 ± 104 b | 12 ± 1 ×107 c |
| Metabolite | RT 1 (min) | Experimental 2 [M + H]+ m/z | Calculated 3 [M + H]+ m/z | Tentative M. Formula | Δm 4 (ppm) | # Sigma 5 | MS Fragmentation (Intensity) 6 | MS2 Fragmentation (Intensity) 7 | l (nm) 8 | Putative Compound | MSI 9 |
|---|---|---|---|---|---|---|---|---|---|---|---|
| V3 | 8.18 | 421.1085 | 421.1071 | C20H24N2O4Zn | 3.7 | 5.5 | 421 (16); 377 (25); 351 (100); 260 (20); 242 (6) | 224; 271 | IV | ||
| V8 | 9.80 | 319.0797 | 319.0812 | C16H14O7 | 4.7 | 4.6 | 319 (42); 304 (41); 301 (100); 289 (36); 286 (81); 273 (35); 272 (53); 261 (19); 258 (25); 257 (35); 249 (14); 243 (40); 229 (10) | 301 → 303 (52); 289 (18); 288 (70); 273 (100); 270 (13); 261 (48); 260 (26); 259 (41); 245 (56); 246 (33); 243 (7); 233 (37); 229 (11) 303 → 303 (62); 289 (15); 288 (80); 273 (100); 261 (38); 260 (35); 259 (61); 246 (25); 245 (42); 233 (46) | 224; 258sh; 304; 444 | 8-O-Methylnectriafurone | II |
| V16 | 11.08 | 301.0699 | 301.0707 | C16H12O6 | 2.7 | 66.4 | 303 (7); 301 (5); 289 (14); 288 (100); 287 (17); 273 (72); 257 (11); 245 (23); 243 (9); 229 (6) | 288 -> 289 (35); 288 (13); 274 (89); 271 (51); 256 (33); 247 (93); 246 (91); 245 (32); 244 (44); 243 (100); 233 (34); 231 (66); 229 (70) | 268; 344; 506 | Naphthoquinone | III |
| V24 | 12.00 | 436.1012 | 436.1013 | C23H17NO8 | 3.6 | 2.6 | 436 (100); 421 (49); 420 (33); 408 (12); 403 (10); 393 (39); 379 (13); 375 (48); 358 (6); 347 (11); 330 (11); 304 (19); 286 (41); 258 (8) | 223 | IV | ||
| V29 | 13.57 | 301.0705 | 301.0707 | C16H12O6 | 0.4 | 16.3 | 301 (22); 303 (10); 288 (100); 286 (23); 273 (71); 272 (40); 258 (36); 257 (93); 245 (21); 243 (96); 242 (32); 229 (45) | 265; 344; 514 | Naphthoquinone | III |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jacquat, A.G.; Podio, N.S.; Cañizares, M.C.; Velez, P.A.; Theumer, M.G.; Areco, V.A.; Garcia-Pedrajas, M.D.; Dambolena, J.S. The Growth, Pathogenesis, and Secondary Metabolism of Fusarium verticillioides Are Epigenetically Modulated by Putative Heterochromatin Protein 1 (FvHP1). J. Fungi 2025, 11, 424. https://doi.org/10.3390/jof11060424
Jacquat AG, Podio NS, Cañizares MC, Velez PA, Theumer MG, Areco VA, Garcia-Pedrajas MD, Dambolena JS. The Growth, Pathogenesis, and Secondary Metabolism of Fusarium verticillioides Are Epigenetically Modulated by Putative Heterochromatin Protein 1 (FvHP1). Journal of Fungi. 2025; 11(6):424. https://doi.org/10.3390/jof11060424
Chicago/Turabian StyleJacquat, Andrés G., Natalia S. Podio, María Carmen Cañizares, Pilar A. Velez, Martín G. Theumer, Vanessa A. Areco, María Dolores Garcia-Pedrajas, and José S. Dambolena. 2025. "The Growth, Pathogenesis, and Secondary Metabolism of Fusarium verticillioides Are Epigenetically Modulated by Putative Heterochromatin Protein 1 (FvHP1)" Journal of Fungi 11, no. 6: 424. https://doi.org/10.3390/jof11060424
APA StyleJacquat, A. G., Podio, N. S., Cañizares, M. C., Velez, P. A., Theumer, M. G., Areco, V. A., Garcia-Pedrajas, M. D., & Dambolena, J. S. (2025). The Growth, Pathogenesis, and Secondary Metabolism of Fusarium verticillioides Are Epigenetically Modulated by Putative Heterochromatin Protein 1 (FvHP1). Journal of Fungi, 11(6), 424. https://doi.org/10.3390/jof11060424