

Fungal Species Associated with Tuber Rot of Foshou Yam (Dioscorea esculenta) in China

 ,
,  , and
, and
Abstract
1. Introduction
2. Materials and Methods
2.1. Fungal Isolation
2.2. DNA Extraction and PCR Amplification
2.3. Phylogenetic Analysis
2.4. Morphology
2.5. Pathogenicity Test
2.6. Statistical Analysis
3. Results
3.1. Fungal Strains
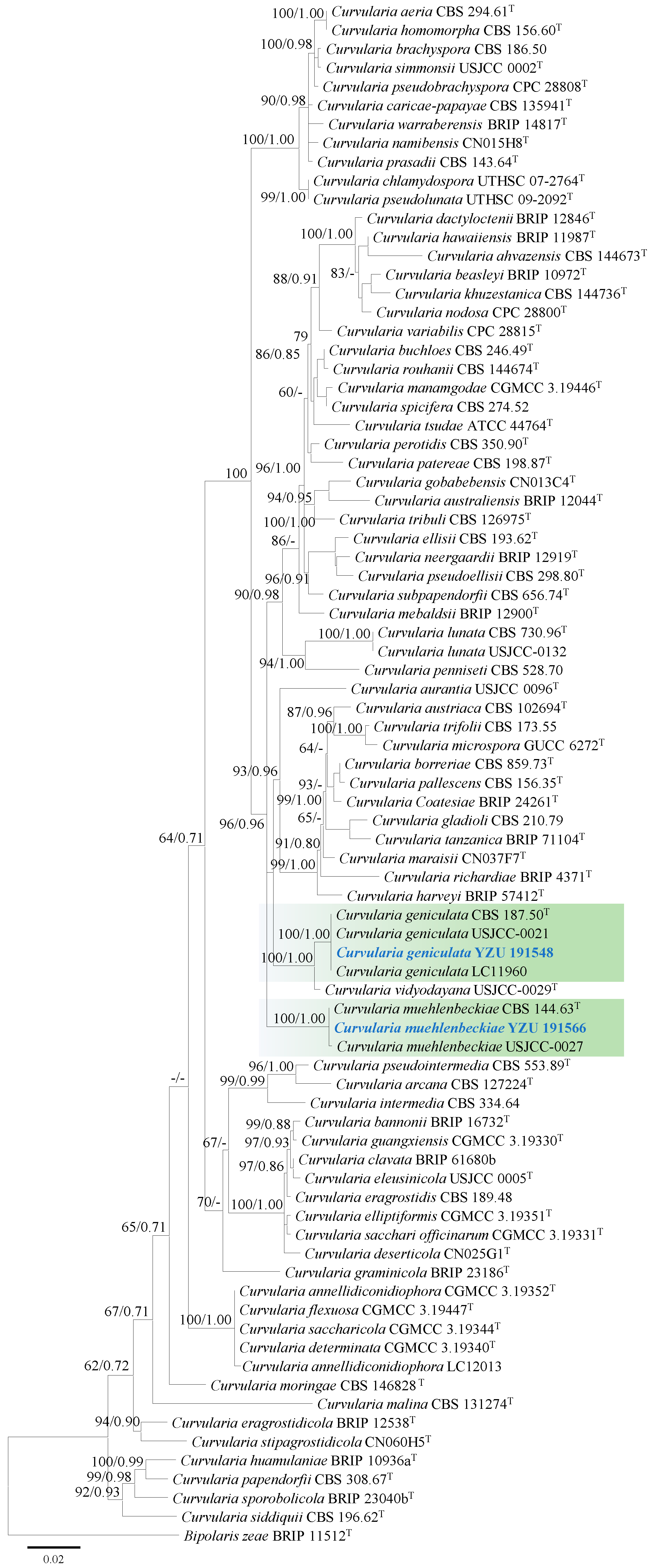
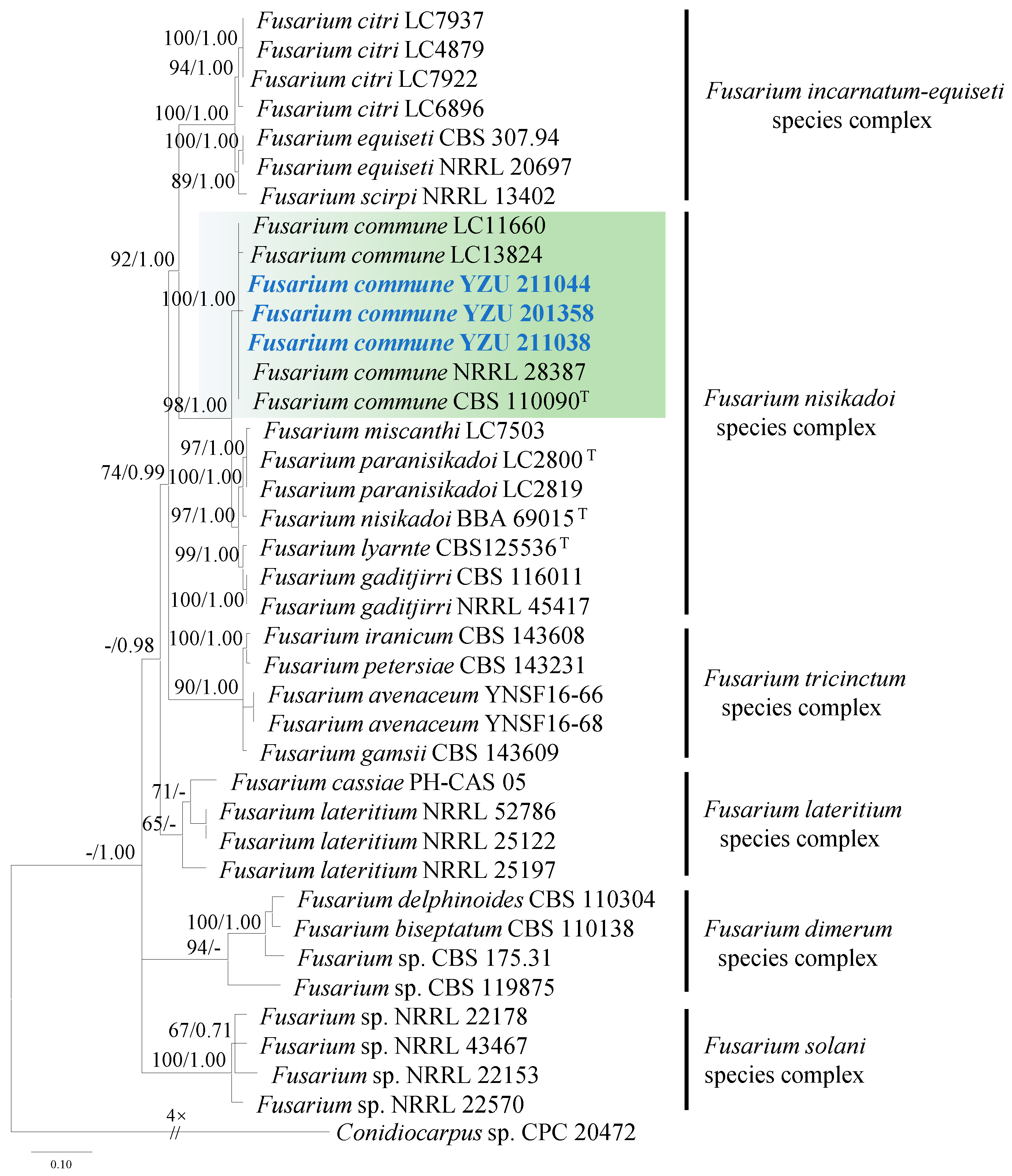
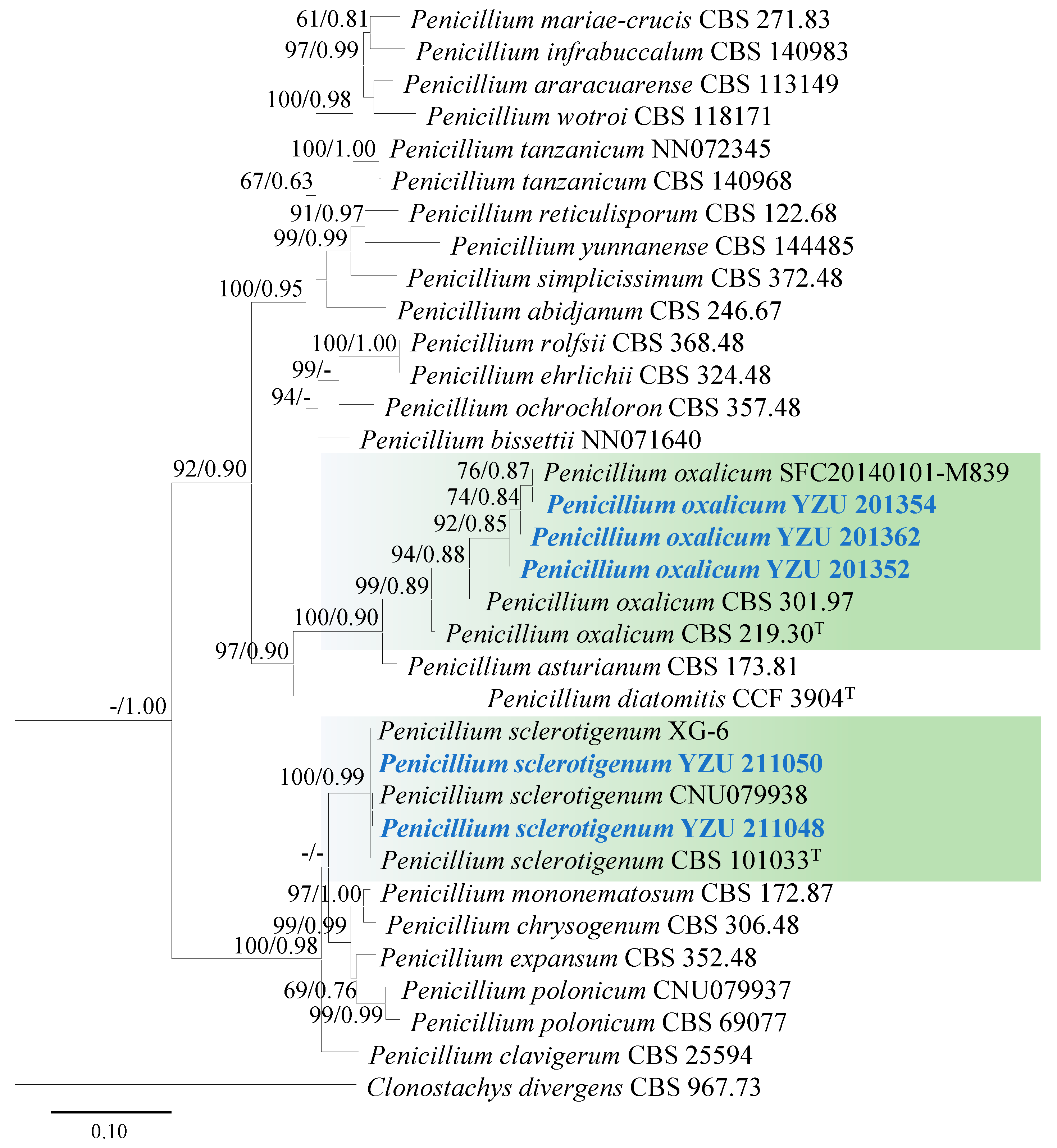
3.2. Molecular Phylogeny
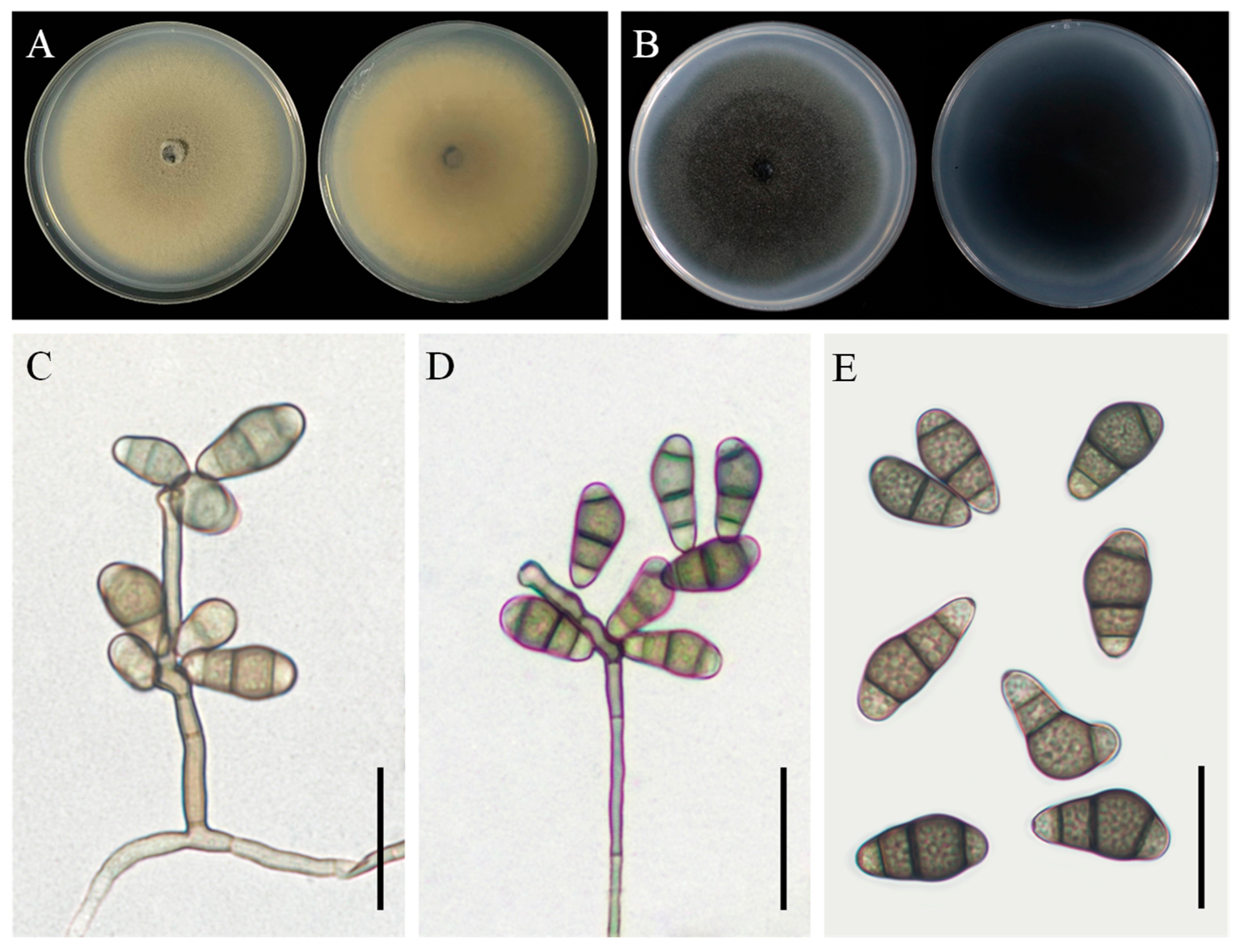
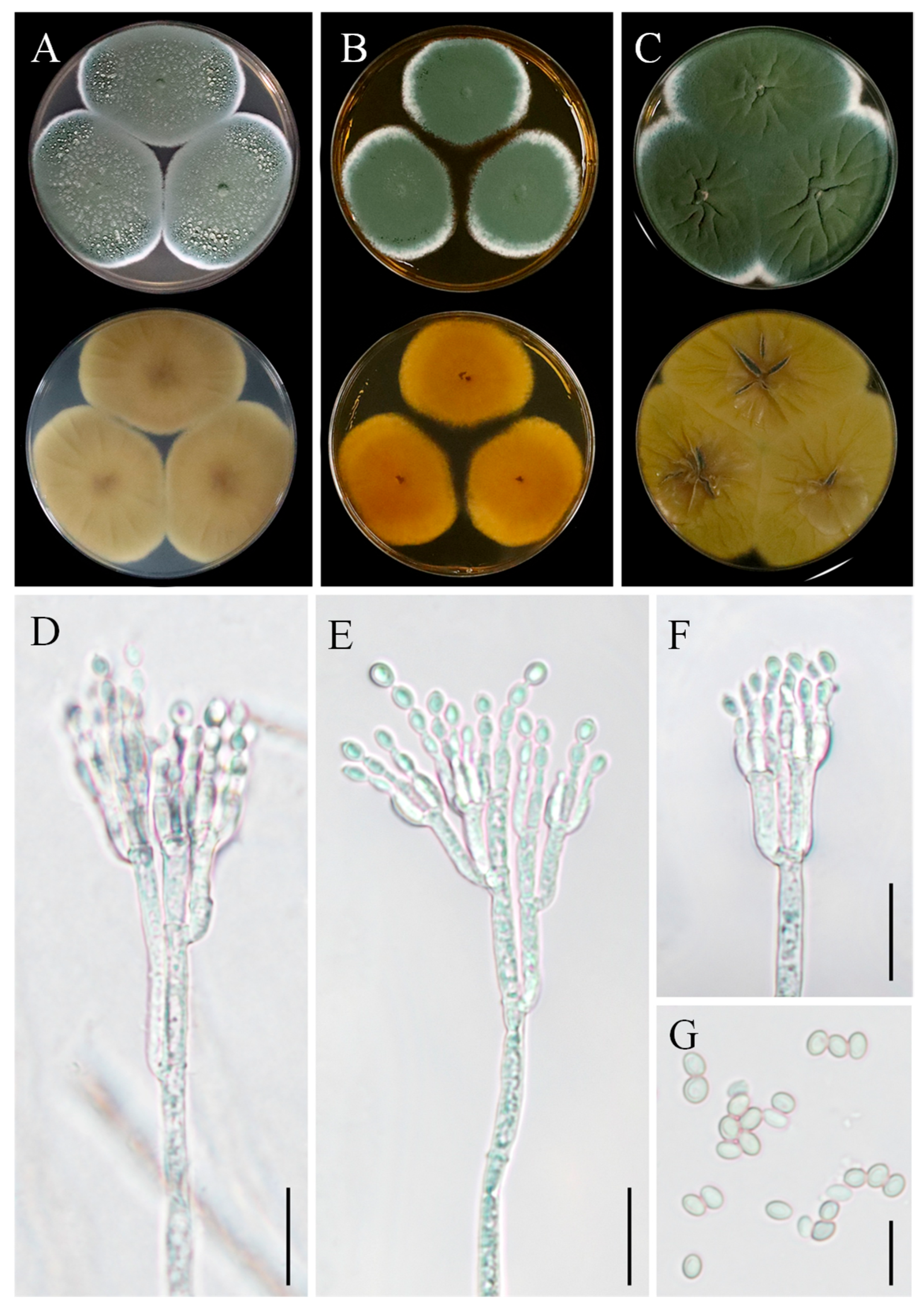
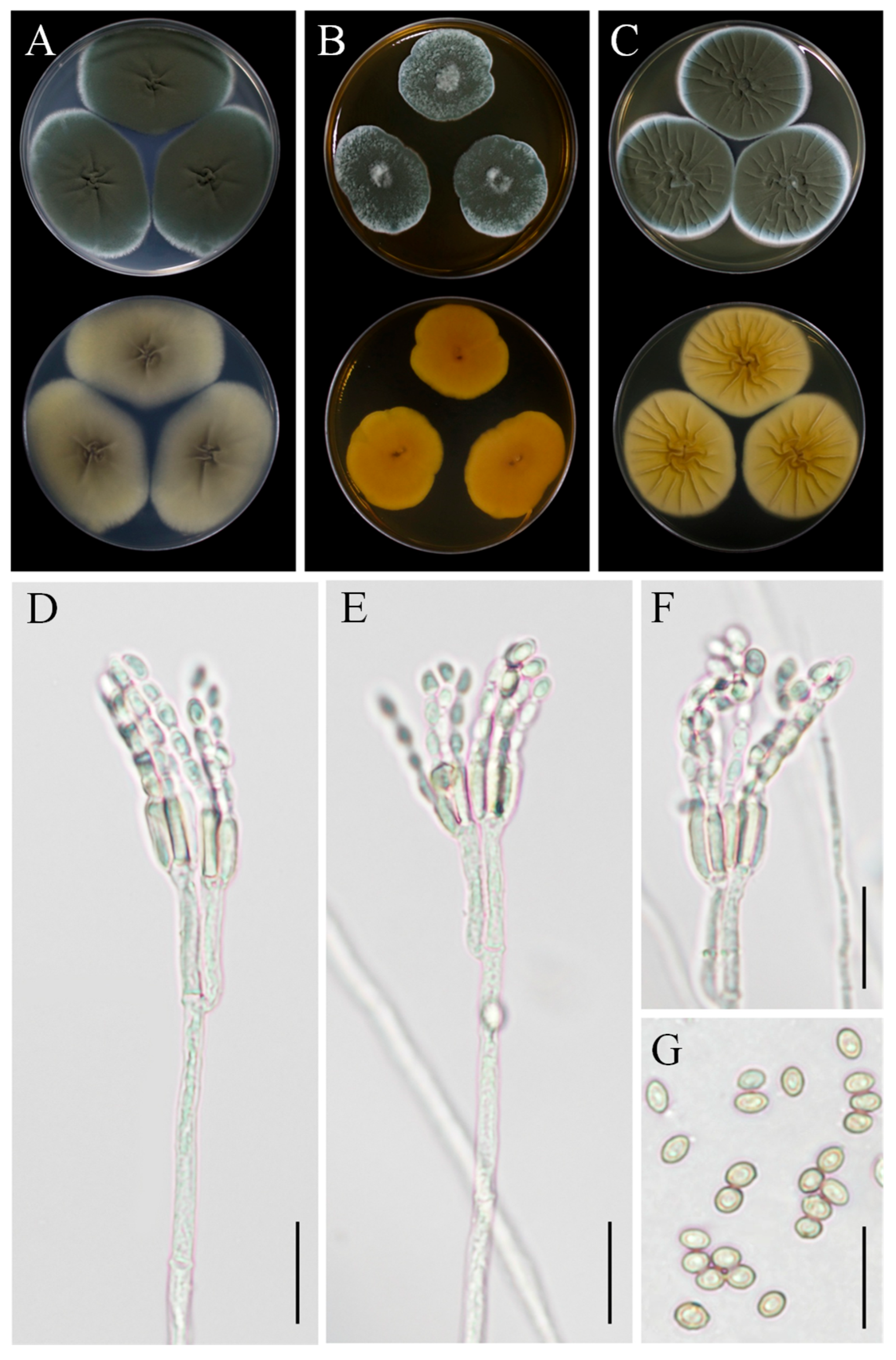
3.3. Taxonomy: Curvularia geniculata YZU 191548
3.4. Taxonomy: Curvularia muehlenbeckiae YZU 191566
3.5. Taxonomy: Fusarium commune YZU 201358
3.6. Taxonomy: Penicillium sclerotigenum YZU 211050
3.7. Taxonomy: Penicillium oxalicum YZU 201352
3.8. Pathogenicity
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Aidoo, K.A. Identification of Yam Tuber Rot Fungi from Storage Systems at the Kumasi Central Market. Ph.D. Thesis, Kwame Nkrumah University of Science and Technology, Kumasi, Ghana, 2007. [Google Scholar]
- Achar, K.; Vasanthakumari, M.; Mahishi, P.; Mallikarjunaswamy, G.; Shivanna, M. Prevalence and Severity of Anthracnose of Yam (Dioscorea alata and D. bulbifera) Caused by Colletotrichum gloeosporioides in Bhadra Wildlife Sanctuary in Karnataka. J. Mycol. Plant Pathol. 2013, 43, 282–290. [Google Scholar]
- Guan, Y.; Pearsall, D.M.; Gao, X.; Chen, F.; Pei, S.; Zhou, Z. Plant Use Activities during the Upper Paleolithic in East Eurasia: Evidence from the Shuidonggou Site, Northwest China. Quat. Int. 2014, 347, 74–83. [Google Scholar] [CrossRef]
- Peng, B.; Zhang, Y.; Sun, X.; Li, M.; Xue, J.; Hang, Y. Genetic Relationship and Identification of Dioscorea polystachya Cultivars Accessed by ISAP and SCAR Markers. Arch. Biol. Sci. 2017, 69, 277–284. [Google Scholar] [CrossRef]
- Fan, D.; Zhong, H.; Hu, B.; Tian, Z.; Sun, L.; Fischer, G.; Wang, X.; Jiang, Z. Agro-Ecological Suitability Assessment of Chinese Medicinal Yam under Future Climate Change. Environ. Geochem. Health 2020, 42, 987–1000. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Shan, N.; Ali, A.; Sun, J.; Luo, S.; Xiao, Y.; Wang, S.; Hu, R.; Huang, Y.; Zhou, Q. Comprehensive Evaluation of Functional Components, Biological Activities, and Minerals of Yam Species (Dioscorea polystachya and D. alata) from China. LWT 2022, 168, 113964. [Google Scholar] [CrossRef]
- Li, Y.; Ji, S.; Xu, T.; Zhong, Y.; Xu, M.; Liu, Y.; Li, M.; Fan, B.; Wang, F.; Xiao, J.; et al. Chinese Yam (Dioscorea): Nutritional Value, Beneficial Effects, and Food and Pharmaceutical Applications. Trends Food Sci. Technol. 2023, 134, 29–40. [Google Scholar] [CrossRef]
- Hu, T.; Wang, F.; Liu, B.; Zheng, Y.; Wu, P.; Li, S. Novel Alkali Extraction, Optimisation, Characterisation, and Antioxidant Activity of Polysaccharides from Foshou Yam. Int. Food Res. J. 2024, 31, 1134–1143. [Google Scholar]
- Chen, S.; Yu, S.; Deng, X.; Hu, T. Research and Optimization of Foshou Yam and Honey Nutritional Can by Response Surface Methodology. J. Agric. Biotechnol. 2020, 9, 156–161. [Google Scholar]
- Tariq, H.; Xiao, C.; Wang, L.; Ge, H.; Wang, G.; Shen, D.; Dou, D. Current Status of Yam Diseases and Advances of Their Control Strategies. Agronomy 2024, 14, 1575. [Google Scholar] [CrossRef]
- Amusa, A.N.; Adegbite, A.A.; Muhammed, S.; Baiyewu, R.A. Yam Diseases and Its Management in Nigeria. Afr. J. Biotechnol. 2003, 2, 497–502. [Google Scholar] [CrossRef]
- Akinbo, O. Pre and Post Harvest Studies of Yam Diseases and Their Control Measure in South Eastern Nigeria. Niger. Agric. J. 2019, 50, 193–197. [Google Scholar]
- Paparu, P.; Acur, A.; Kato, F.; Acam, C.; Nakibuule, J.; Nkuboye, A.; Musoke, S.; Mukankusi, C. Morphological and Pathogenic Characterization of Sclerotium rolfsii, the Causal Agent of Southern Blight Disease on Common Bean in Uganda. Plant Dis. 2020, 104, 2130–2137. [Google Scholar] [CrossRef] [PubMed]
- Lu, X.; Xiao, C.; Zhou, K.; Fu, L.; Shen, D.; Dou, D. First Report of Nigrospora oryzae Causing Leaf Spot on Yam in China. Plant Dis. 2023, 107, 2256. [Google Scholar] [CrossRef] [PubMed]
- Hong, S.-K.; Kim, W.-G.; Lee, Y.-K.; Choi, H.-W.; Choi, K.-J.; Lee, S.-Y. Leaf Spot of Yam Caused by Pseudophloeosporella dioscoreae in Korea. Mycobiology 2010, 38, 78–80. [Google Scholar] [CrossRef] [PubMed]
- Aboagye-Nuamah, F.; Offei, S.K.; Cornelius, E.W.; Bancroft, R.D. Severity of Spoilage Storage Rots of White Yam (Dioscorea rotundata Poir.). Ann. Appl. Biol. 2005, 147, 183–190. [Google Scholar] [CrossRef]
- Di Martino, C.; Torino, V.; Minotti, P.; Pietrantonio, L.; Del Grosso, C.; Palmieri, D.; Palumbo, G.; Crawford, T.W.; Carfagna, S. Mycorrhized Wheat Plants and Nitrogen Assimilation in Coexistence and Antagonism with Spontaneous Colonization of Pathogenic and Saprophytic Fungi in a Soil of Low Fertility. Plants 2022, 11, 924. [Google Scholar] [CrossRef] [PubMed]
- Cubero, O.F.; Crespo, A.; Fatehi, J.; Bridge, P.D. DNA Extraction and PCR Amplification Method Suitable for Fresh, Herbarium-Stored, Lichenized, and Other Fungi. Plant Syst. Evol. 1999, 216, 243–249. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and Direct Sequencing of Fungal Ribosomal RNA Genes for Phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: San Diego, CA, USA, 1990; Volume 18, pp. 315–322. [Google Scholar]
- Glass, N.L.; Donaldson, G.C. Development of Primer Sets Designed for Use with the PCR to Amplify Conserved Genes from Filamentous Ascomycetes. Appl. Environ. Microbiol. 1995, 61, 1323–1330. [Google Scholar] [CrossRef]
- Hong, S.-B.; Cho, H.-S.; Shin, H.-D.; Frisvad, J.C.; Samson, R.A. Novel Neosartorya Species Isolated from Soil in Korea. Int. J. Syst. Evol. Microbiol. 2006, 56, 477–486. [Google Scholar] [CrossRef]
- Carbone, I.; Kohn, L.M. A Method for Designing Primer Sets for Speciation Studies in Filamentous Ascomycetes. Mycologia 1999, 91, 553–556. [Google Scholar] [CrossRef]
- Berbee, M.L.; Pirseyedi, M.; Hubbard, S. Cochliobolus Phylogenetics and the Origin of Known, Highly Virulent Pathogens, Inferred from ITS and Glyceraldehyde-3-Phosphate Dehydrogenase Gene Sequences. Mycologia 1999, 91, 964–977. [Google Scholar] [CrossRef]
- Liu, Y.J.; Whelen, S.; Hall, B.D. Phylogenetic Relationships among Ascomycetes: Evidence from an RNA Polymerse II Subunit. Mol. Biol. Evol. 1999, 16, 1799–1808. [Google Scholar] [CrossRef] [PubMed]
- Hall, T.A. BioEdit: A User-Friendly Biological Sequence Alignment Editor and Analysis Program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Ferdinandez, H.S.; Manamgoda, D.S.; Udayanga, D.; Munasinghe, M.S.; Castlebury, L.A. Molecular Phylogeny and Morphology Reveal Two New Graminicolous Species, Curvularia aurantia sp. nov. and C. vidyodayana sp. nov. with New Records of Curvularia spp. from Sri Lanka. Fungal Syst. Evol. 2023, 12, 219–246. [Google Scholar] [CrossRef] [PubMed]
- Van Vuuren, N.; Yilmaz, N.; Wingfield, M.J.; Visagie, C.M. Five Novel Curvularia Species (Pleosporaceae, Pleosporales) Isolated from Fairy Circles in the Namib Desert. Mycol. Prog. 2024, 23, 39. [Google Scholar] [CrossRef]
- Crous, P.W.; Costa, M.M.; Kandemir, H.; Vermaas, M.; Vu, D.; Zhao, L.; Arumugam, E.; Flakus, A.; Jurjević, Ž.; Kaliyaperumal, M.; et al. Fungal Planet Description Sheets: 1550–1613. Persoonia 2023, 51, 280–417. [Google Scholar] [CrossRef]
- Skovgaard, K.; Rosendahl, S.; O’Donnell, K.; Nirenberg, H.I. Fusarium commune Is a New Species Identified by Morphological and Molecular Phylogenetic Data. Mycologia 2003, 95, 630–636. [Google Scholar] [CrossRef]
- Wang, M.M.; Crous, P.W.; Sandoval-Denis, M.; Han, S.L.; Liu, F.; Liang, J.M.; Duan, W.J.; Cai, L. Fusarium and Allied Genera from China: Species Diversity and Distribution. Persoonia 2022, 48, 1–53. [Google Scholar] [CrossRef]
- Han, S.L.; Wang, M.M.; Ma, Z.Y.; Raza, M.; Zhao, P.; Liang, J.M.; Gao, M.; Li, Y.J.; Wang, J.W.; Hu, D.M.; et al. Fusarium Diversity Associated with Diseased Cereals in China, with an Updated Phylogenomic Assessment of the Genus. Stud. Mycol. 2023, 104, 87–148. [Google Scholar] [CrossRef]
- Park, M.S.; Fong, J.J.; Oh, S.-Y.; Kwon, K.K.; Sohn, J.H.; Lim, Y.W. Marine-Derived Penicillium in Korea: Diversity, Enzyme Activity, and Antifungal Properties. Antonie Van Leeuwenhoek 2014, 106, 331–345. [Google Scholar] [CrossRef]
- Zeng, X.; Tan, T.; Tian, F.; Wang, Y.; Wen, T. OFPT: A One-Stop Software for Fungal Phylogeny. Mycosphere 2023, 14, 1730–1741. [Google Scholar] [CrossRef]
- Katoh, K.; Standley, D.M. MAFFT Multiple Sequence Alignment Software Version 7: Improvements in Performance and Usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [PubMed]
- Capella-Gutiérrez, S.; Silla-Martínez, J.M.; Gabaldón, T. trimAl: A Tool for Automated Alignment Trimming in Large-Scale Phylogenetic Analyses. Bioinformatics 2009, 25, 1972–1973. [Google Scholar] [CrossRef] [PubMed]
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.K.F.; Von Haeseler, A.; Jermiin, L.S. ModelFinder: Fast Model Selection for Accurate Phylogenetic Estimates. Nat. Methods 2017, 14, 587–589. [Google Scholar] [CrossRef]
- Nguyen, L.-T.; Schmidt, H.A.; Von Haeseler, A.; Minh, B.Q. IQ-TREE: A Fast and Effective Stochastic Algorithm for Estimating Maximum-Likelihood Phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef]
- Ronquist, F.; Teslenko, M.; Van Der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian Phylogenetic Inference and Model Choice across a Large Model Space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef]
- Kim, W.-K.; Hwang, Y.-S.; Yu, S.-H. Two Species of Penicillium Associated with Blue Mold of Yam in Korea. Mycobiology 2008, 36, 217–221. [Google Scholar] [CrossRef]
- Lin, C.-H.; Chen, Y.-X.; Liu, W.-B.; Wu, W.-Q.; Miao, W.-G.; Zheng, F.-C. First Report of Dioscorea esculenta Dieback Caused by Neoscytalidium dimidiatum in China. Plant Dis. 2017, 101, 1320. [Google Scholar] [CrossRef]
- Madrid, H.; Da Cunha, K.C.; Gené, J.; Dijksterhuis, J.; Cano, J.; Sutton, D.A.; Guarro, J.; Crous, P.W. Novel Curvularia Species from Clinical Specimens. Persoonia 2014, 33, 48–60. [Google Scholar] [CrossRef]
- Khemmuk, W.; Shivas, R.G.; Henry, R.J.; Geering, A.D.W. Fungi Associated with Foliar Diseases of Wild and Cultivated Rice (Oryza Spp.) in Northern Queensland. Australas. Plant Pathol. 2016, 45, 297–308. [Google Scholar] [CrossRef]
- Raza, M.; Zhang, Z.-F.; Hyde, K.D.; Diao, Y.-Z.; Cai, L. Culturable Plant Pathogenic Fungi Associated with Sugarcane in Southern China. Fungal Divers. 2019, 99, 1–104. [Google Scholar] [CrossRef]
- Olivas-Peraza, D.D.; Leyva-Madrigal, K.Y.; Maldonado-Mendoza, I.E.; Félix-Gastélum, R. Curvularia muehlenbeckiae Causing Leaf Spot on Johnson Grass in Mexico. Mycol. Progress 2022, 21, 50. [Google Scholar] [CrossRef]
- Tan, Y.P.; Crous, P.W.; Shivas, R.G. Cryptic Species of Curvularia in the Culture Collection of the Queensland Plant Pathology Herbarium. Mycokeys 2018, 35, 1–25. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Tang, T.; Chen, C.; Wei, L.; Zhou, D. First Report of Curvularia Leaf Spot Caused by Curvularia muehlenbeckiae on Zizania latifolia in China. J. Plant Pathol. 2021, 103, 1073. [Google Scholar] [CrossRef]
- Wang, J.; Chai, Z.; Bao, Y.; Wang, H.; Li, Y.; Rao, G.P.; Zhang, M. First Report of Fusarium commune Causing Root Rot Disease of Sugarcane (Var. Badila) in China. Plant Dis. 2018, 102, 1660. [Google Scholar] [CrossRef]
- Ellis, M.L.; Arias, M.M.D.; Jimenez, D.R.C.; Munkvold, G.P.; Leandro, L.F. First Report of Fusarium commune Causing Damping-off, Seed Rot, and Seedling Root Rot on Soybean (Glycine max) in the United States. Plant Dis. 2013, 97, 284. [Google Scholar] [CrossRef]
- Hamini-Kadar, N.; Edel-Hermann, V.; Gautheron, N.; Steinberg, C. First Report of Fusarium commune and Fusarium redolens Causing Crown and Root Rot on Tomato in Algeria. New Dis. Rep. 2010, 22, 3. [Google Scholar] [CrossRef]
- Yu, J.M.; Babadoost, M. Occurrence of Fusarium commune and F. oxysporum in Horseradish Roots. Plant Dis. 2013, 97, 453–460. [Google Scholar] [CrossRef]
- Zhu, Z.; Zheng, L.; Pan, L.; Hsiang, T.; Huang, J. Identification and Characterization of Fusarium Species Associated with Wilt of Eleocharis dulcis (Chinese Water Chestnut) in China. Plant Dis. 2014, 98, 977–987. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fungal Strains | Measurement of Lesion (mm) * |
|---|---|
| Curvularia geniculata YZU 191548 | 10.65 ± 0.69 b |
| Curvularia muehlenbeckiae YZU 191566 | 13.31 ± 0.41 b |
| Fusarium commune YZU 201358 | 13.01 ± 0.36 b |
| Penicillium oxalicum YZU 201352 | 22.17 ± 2.01 a |
| Penicillium sclerotigenum YZU 211050 | 11.50 ± 0.68 b |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, H.; Htun, A.A.; Aung, S.L.L.; Sang, H.; Deng, J.; Tao, Y. Fungal Species Associated with Tuber Rot of Foshou Yam (Dioscorea esculenta) in China. J. Fungi 2025, 11, 380. https://doi.org/10.3390/jof11050380
Liu H, Htun AA, Aung SLL, Sang H, Deng J, Tao Y. Fungal Species Associated with Tuber Rot of Foshou Yam (Dioscorea esculenta) in China. Journal of Fungi. 2025; 11(5):380. https://doi.org/10.3390/jof11050380
Chicago/Turabian StyleLiu, Haifeng, Aye Aye Htun, Sein Lai Lai Aung, Hyunkyu Sang, Jianxin Deng, and Yaqun Tao. 2025. "Fungal Species Associated with Tuber Rot of Foshou Yam (Dioscorea esculenta) in China" Journal of Fungi 11, no. 5: 380. https://doi.org/10.3390/jof11050380
APA StyleLiu, H., Htun, A. A., Aung, S. L. L., Sang, H., Deng, J., & Tao, Y. (2025). Fungal Species Associated with Tuber Rot of Foshou Yam (Dioscorea esculenta) in China. Journal of Fungi, 11(5), 380. https://doi.org/10.3390/jof11050380