
What Are “Lingzhi Wang” or “Zhu Lingzhi”? Notes on Ganoderma (Ganodermataceae, Polyporales) Species Characterized by Diminutive Pilei and Gracile Stipes from Hainan Island, Tropical China

Abstract
1. Introduction
2. Materials and Methods
2.1. Key Informant Interview Methodology
2.2. Sample Collection and Morphological Studies
2.3. Molecular Procedures
2.4. Dataset Assembly

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| GenBank Accession Nos. | ||||||
|---|---|---|---|---|---|---|
| Taxon | Voucher | Locality | ITS | rpb2 | tef1 | Reference |
| Cristataspora coffeata | FLOR 50933 | Brazil | KU315204 | — | — | [33] |
| C. coffeata | Robledo 3183 | Brazil | MN077526 | — | MN061695 | [34] |
| C. coffeata | Robledo 3182 | Brazil | MN077525 | — | — | [34] |
| C. flavipora | Robledo 3288 | Argentina | MN077521 | — | MN061694 | [34] |
| Foraminispora concentrica | Cui 16238 | Yunnan, SW China | MK119816 | MK121504 | MK121565 | [30] |
| F. concentrica | Cui 16239 | Yunnan, SW China | MK119817 | MK121506 | MK121566 | [30] |
| Ganoderma acaciicola | Cui 16815 | Australia | MZ354895 | MZ245384 | — | [2] |
| G. acaciicola | Cui 16814 | Australia | MZ354894 | MZ245383 | — | [2] |
| G. acontextum | JV 0611/21G | Guatemala | KF605667 | MG367489 | MG367538 | [2] |
| G. acontextum | JV 1208/11J | USA | KF605668 | MG367490 | MG367540 | [2] |
| G. adspersum | ITA 39 | Italy | EF060011 | — | — | [35] |
| G. adspersum | PF263 | Italy | JN176908 | — | — | Unpublished |
| G. alpinum | Cui 17467 | Yunnan, SW China | MZ354912 | — | — | [2] |
| G. alpinum | Cui 18402 | Xizhang, western China | MZ354910 | — | — | [2] |
| G. angustisporum | Cui 13817 | Fujian, SE China | MG279170 | MG367507 | MG367563 | [6] |
| G. angustisporum | Cui 14578 | Guangdong, southern China | MG279171 | — | MG367564 | [6] |
| G. applanatum | Cui 14062 | Jinlin, NE China | MZ354913 | MZ358846 | MZ221635 | [2] |
| G. applanatum | Cui 14070 | Jinlin, NE China | MZ354914 | MZ245387 | MZ221636 | [2] |
| G. aridicola | Dai 12588 | South Africa | KU572491 | — | KU572502 | [36] |
| G. aridicola | GanoTK25 | Cameroon | JN105707 | — | — | [2] |
| G. artocarpicola | HL173 | Yunnan, SW China | ON994239 | OP508428 | OP508442 | [37] |
| G. artocarpicola | HL188 | Yunnan, SW China | ON994240 | OP508427 | OP508441 | [37] |
| G. australe | DHCR411 | Australia | MF436675 | — | MF436677 | [38] |
| G. australe | DHCR417 | Australia | MF436676 | — | MF436678 | [38] |
| G. austroafricanum | CBS138724 | South Africa | KM507324 | — | — | [39] |
| G. bambusicola | Wu 1207-151 | Taiwan, SE China | MN957781 | LC517944 | LC517941 | [8] |
| G. baisuzhenii | N.K. Zeng2080 (FHMU2334) | Hainan, southern China | — | PP785032 | PV066218 | This study |
| G. baisuzhenii | N.K. Zeng2519 (FHMU7350) | Hainan, southern China | PP663110 | PP785031 | PV066219 | This study |
| G. bambusicola | N.K. Zeng1892 (FHMU1217) R71 | Hainan, southern China | — | — | PP922172 | This study |
| G. bambusicola | N.K. Zeng1892-1 (FHM7610) | Hainan, southern China | — | — | PP922173 | This study |
| G. bambusicola | N.K. Zeng10386 (FHMU8798) | Hainan, southern China | PV052368 | — | PV066216 | This study |
| G. bambusicola | N.K. Zeng10387 (FHMU8803) | Hainan, southern China | PV052369 | — | — | This study |
| G. bambusicola | N.K. Zeng10388 (FHMU8791) | Hainan, southern China | PV052370 | PV066220 | — | This study |
| G. bambusicola | N.K. Zeng10340 (FHMU7930) | Hainan, southern China | PV052371 | PV066221 | PV066217 | This study |
| G. boninense | WD 2028 | Japan | KJ143905 | KJ143964 | KJ143924 | [40] |
| G. boninense | WD 2085 | Japan | KJ143906 | KJ143965 | KJ143925 | [40] |
| G. brownii | JV 1105/9J | Australia | MG279159 | MG367494 | MG367547 | [6] |
| G. brownii | JV 0709/109 | — | KF605662 | MG367495 | MG367548 | [2,6] |
| G. bubalinomarginatum | Dai 20074 | Guangxi, southern China | MZ354926 | MZ245388 | MZ221637 | [2] |
| G. bubalinomarginatum | Dai 20075 | Guangxi, southern China | MZ354927 | MZ245389 | MZ221638 | [2] |
| G. calidophilum | MFLU 19-2174 | Yunnan, SW China | MN398337 | — | — | [3] |
| G. calidophilum | H36 | Yunnan, SW China | MW750241 | MW839003 | MW838997 | [1] |
| G. carnosum | JV 8709/8 | Czech Republic | KU572493 | — | — | [36] |
| G. carnosum | MJ 21/08 | Czech Republic | KU572492 | — | — | [36] |
| G. carocalcareus | DMC 322 | Cameroon | EU089969 | — | — | [41] |
| G. carocalcareus | DMC 513 | Cameroon | EU089970 | — | — | [41] |
| G. castaneum | Dai 16500 | Hainan, southern China | MZ354918 | MZ245390 | MZ221639 | [2] |
| G. castaneum | Cui 13893 | Hainan, southern China | MZ221640 | MZ245391 | MZ354919 | [2] |
| G. casuarinicola | Dai 16336 | Guangdong, southern China | MG279173 | MG367508 | MG367565 | [6] |
| G. casuarinicola | Dai 16337 | Guangdong, southern China | MG279174 | MG367509 | MG367566 | [6] |
| G. chalceum | URM80457 | Brazil | JX310812 | — | — | [42] |
| G. chocoense | QCAM 3123 | Ecuador | MH890527 | — | — | [43] |
| G. chuxiongense | Cui 17262 | Yunnan, SW China | MZ354907 | — | — | [2] |
| G. cocoicola | Cui 16791 | Australia | MZ354984 | MZ245393 | MZ221643 | [2] |
| G. cocoicola | Cui 16792 | Australia | MZ354985 | MZ245394 | MZ221644 | [2] |
| G. concinnum | Robledo 3192 (FCOS) | — | MN077522 | — | — | [34] |
| G. concinnum | Robledo 3235 (FCOS) | — | MN077523 | — | — | [34] |
| G. cupreum | GanoTK4 | Cameroon | JN105701 | — | — | Unpublished |
| G. cupreum | GanoTK7 | Cameroon | JN105702 | — | — | Unpublished |
| G. curtisii | CBS 100131 | NC, USA | JQ781848 | KJ143966 | KJ143926 | [11,40] |
| G. curtisii | CBS 100132 | NC, USA | JQ781849 | KJ143967 | KJ143927 | [11,40] |
| G. destructans | CMW43670 | South Africa | KR183856 | — | — | [39] |
| G. dianzhongense | L4331 | Yunnan, SW China | MW750237 | MZ467043 | MW838993 | [1] |
| G. dianzhongense | L4969 | Yunnan, SW China | MW750240 | MZ467044 | MW838996 | [1] |
| G. dorsale | MVHC 5701 | Uruguay | MN191581 | — | — | [44] |
| G. dorsale | MVHC 5653 | Uruguay | MN191578 | — | — | [44] |
| G. dunense | CMW42149 | South Africa | MG020248 | — | MG020226 | [45] |
| G. dunense | CMW42157 | South Africa | MG020255 | — | — | [45] |
| G. ecuadorense | ASL799 | Ecuador | KU128524 | — | — | [46] |
| G. ecuadorense | PMC126 | Ecuador | KU128525 | — | — | [46] |
| G. eickeri | CMW 49692 | South Africa | MH571690 | — | MH567287 | [47] |
| G. eickeri | CMW 50325 | South Africa | MH571689 | — | MH567290 | [47] |
| G. ellipsoideum | GACP14080966 | Hainan, southern China | MH106867 | — | — | [5] |
| G. ellipsoideum | GACP14080968 | Hainan, southern China | MH106868 | — | — | [5] |
| G. ellipsoideum | GACP14081228 | Hainan, southern China | MH106886 | — | — | [5] |
| G. enigmaticum | CMW43669 | South Africa | KR183855 | — | — | [39] |
| G. enigmaticum | CBS 139792 | South Africa | NR132918 | — | — | [39] |
| G. esculentum | L4935 | Yunnan, SW China | MW750242 | MW839004 | MW838998 | [1] |
| G. esculentum | HL107 | Yunnan, SW China | ON994243 | OP508424 | OP508437 | [48] |
| G. fallax | JV 1009/27 | USA | KF605655 | — | — | [2] |
| G. fallax | JV 0709/39 | USA | KF605658 | — | — | [2] |
| G. flexipes | Cui 13841 | Hainan, southern China | MZ354923 | MZ245401 | MZ221655 | [2] |
| G. flexipes | Cui 13863 | Hainan, southern China | MZ354924 | MZ245402 | MZ221656 | [2] |
| G. flexipes | GACP14045450 | Hainan, southern China | MH106873 | — | — | [5] |
| G. flexipes | Wei5200 | China | JN383978 | — | — | [49] |
| G. flexipes | N.K. Zeng2607 (FHMU3352) | Hainan, southern China | PP663099 | — | — | This study |
| G. flexipes | N.K. Zeng2042 (FHMU2329) | Hainan, southern China | PP663094 | PP785027 | PP911339 | This study |
| G. flexipes | N.K. Zeng2616 (FHMU5678) | Hainan, southern China | PP663101 | — | — | This study |
| G. flexipes | N.K. Zeng2617 (FHMU5663) | Hainan, southern China | PP663096 | — | — | This study |
| G. flexipes | N.K. Zeng2624 (FHMU5681) | Hainan, southern China | PP663103 | — | — | This study |
| G. flexipes | N.K. Zeng2627 (FHMU5659) | Hainan, southern China | PP663098 | — | — | This study |
| G. flexipes | N.K. Zeng2606 (FHMU3360) | Hainan, southern China | PP663100 | — | — | This study |
| G. flexipes | N.K. Zeng2614 (FHMU5672) | Hainan, southern China | PP663104 | — | — | This study |
| G. flexipes | N.K. Zeng210 (FHMU2292) | Hainan, southern China | PP663095 | — | — | This study |
| G. flexipes | N.K. Zeng2618 (FHMU5661) | Hainan, southern China | PP663097 | — | — | This study |
| G. flexipes | N.K. Zeng4561 (FHMU4863) | Hainan, southern China | PP663106 | — | PP922159 | This study |
| G. flexipes | N.K. Zeng4595 (FHMU4898) | Hainan, southern China | PP663102 | — | PP922158 | This study |
| G. flexipes | N.K. Zeng2087 (FHMU2337) | Hainan, southern China | PP663107 | PP785029 | PP922162 | This study |
| G. flexipes | N.K. Zeng2085 (FHMU2336) | Hainan, southern China | PP663108 | PP785028 | PP922161 | This study |
| “G. flexipes” | Wei5491 | Hainan, southern China | JQ781850 | KJ143968 | — | [11,40] |
| “G. flexipes” | Wei5494 | Hainan, southern China | JN383979 | — | — | [11] |
| G. gibbosum | Cui 13940 | China | MZ354972 | MZ245404 | MZ221658 | [2] |
| G. gibbosum | Cui 14338 | China | MZ354969 | MZ245405 | MZ221659 | [2] |
| G. guangxiense | Cui 14453 | Guangxi, southern China | MZ354939 | MZ245407 | MZ221661 | [2] |
| G. guangxiense | Cui 14454 | Guangxi, southern China | MZ354941 | MZ245408 | MZ221662 | [2] |
| G. guixiense | GXU3457 | Guangxi, southern China | OQ788244 | PP187389 | — | [50] |
| G. guixiense | GXU3709 | Guangxi, southern China | OR271986 | — | — | [50] |
| G. hochiminhense | MFLU 19-2224 | Vietnam | MN398324 | — | MN423176 | [3] |
| G. hochiminhense | MFLU 19-2225 | Vietnam | MN396662 | — | MN423177 | [3] |
| G. hoehnelianum | GACP14080913 | Hainan, southern China | MH106881 | — | — | [5] |
| G. hoehnelianum | MFLU 19-2168 | Myanmar | MN396316 | MN423123 | MN423158 | [3] |
| G. knysnamense | CMW 47755 | South Africa | MH571681 | — | MH567261 | [47] |
| G. knysnamense | CMW 47756 | South Africa | MH571684 | — | MH567274 | [47] |
| G. leucocontextum | GDGM 40200 | Xizang, western China | KF011548 | — | — | [51] |
| G. lingzhi | Dai 12479 | Anhui, central China | JQ781864 | JX029979 | JX029975 | [11] |
| G. lingzhi | Wu 1006-38 | Hubei, eastern China | JQ781858 | JX029980 | JX029976 | [11] |
| G. lobatum | JV 0402/24 | — | KF605677 | — | — | Unpublished |
| G. lobatum | JV 1212/10J | — | KF605676 | — | — | Unpublished |
| G. lucidum | Rivoire 4195 | France | KJ143909 | KJ143969 | — | [40] |
| G. lucidum | K 175217 | UK | KJ143911 | KJ143971 | KJ143929 | [40] |
| G. magniporum | Dai 19966 | Yunnan, SW China | — | MZ345728 | MZ221670 | [2] |
| G. martinicense | SWMart08-55 | Martinique | KF963256 | — | — | [52] |
| G. mbrekobenum | UMN7-3 GHA | Ghana | KX000896 | — | — | [46] |
| G. mbrekobenum | UMN7-4 GHA | Ghana | KX000898 | — | — | [46] |
| G. meredithiae | UMNFL50 | USA | MG654103 | — | — | [53] |
| G. meredithiae | UMNFL64 | USA | MG654106 | MG754863 | — | [53] |
| G. mexicanum | MUCL 49453 SW17 | Martinique | MK531811 | MK531836 | MK531825 | [54] |
| G. mexicanum | MUCL 55832 | Martinique | MK531815 | MK531839 | MK531829 | [54] |
| G. mirabile | Cui 18271 | Malaysia | MZ354958 | MZ345729 | MZ221672 | [2] |
| G. mirabile | Cui 18283 | Malaysia | MZ354959 | MZ345730 | MZ221673 | [2] |
| G. mizoramense | UMN-MZ4 | India | KY643750 | — | — | [55] |
| G. mizoramense | UMN-MZ5 | India | KY643751 | — | — | [55] |
| G. multipileum | CWN 04670 (TNM) | Taiwan, SE China | KJ143913 | KJ143972 | KJ143931 | [40] |
| G. multipileum | Dai 9447 (IFP) | Hainan, southern China | KJ143914 | KJ143973 | KJ143932 | [40] |
| G. multipileum | MFLU 19-2166 | Thailand | MN401406 | MN423142 | MN423172 | [3] |
| G. mutabile | Yuan 2289 | Yunnan, SW China | JN383977 | — | — | [50] |
| G. mutabile | CLZhao 982 | Yunnan, SW China | MG231527 | — | — | Unpublished |
| G. myanmarense | MFLU 19-2167 | Myanmar | MN396330 | — | — | [3] |
| G. myanmarense | MFLU 19-2169 | Myanmar | MN396330 | — | — | [3] |
| G. nasalanense | GACP17060211 | Laos | MK345441 | — | — | [3] |
| G. nasalanense | GACP17060212 | Laos | MK345442 | — | — | [49] |
| G. neojaponicum | FFPRI WD-1285 | Japan | MN957784 | — | — | [8] |
| G. neojaponicum | FFPRI WD-1532 | Japan | MN957785 | — | — | [8] |
| G. nitidum | JV 1504/73 | Hainan, southern China | MZ354933 | — | MZ221681 | [2] |
| G. obscuratum | Lsh88 | Yunnan, SW China | ON994237 | — | OP508450 | [48] |
| G. obscuratum | Lsh89 | Yunnan, SW China | ON994238 | — | OP508451 | [48] |
| G. orbiforme | Cui 13880 | Hainan, southern China | MG279187 | MG367523 | MG367577 | [6] |
| G. orbiforme | Cui 13891 | China | MZ354953 | MZ345736 | MZ221682 | [2] |
| G. oregonense | CBS 265.88 | USA | JQ781875 | KJ143974 | KJ143933 | [11,40] |
| G. oregonense | CBS 266.88 | USA | JQ781876 | KJ143975 | — | [11,40] |
| G. ovisporum | HKAS123193 | Guizhou, SW China | MZ519547 | MZ547661 | — | [9] |
| G. ovisporum | GACP20071602 | Guizhou, SW China | MZ519548 | MZ547662 | — | [9] |
| G. parvulum | URM83343 | Brazil | JQ618246 | — | — | [45] |
| G. parvulum | URM80765 | Brazil | JX310822 | — | — | [45] |
| G. pfeifferi | K(M)120818 | UK | AY884185 | — | — | Unpublished |
| G. philippii | MFLU 19-2222 | Thailand | MN401410 | — | MN423174 | [3] |
| G. philippii | MFLU 19-2223 | Thailand | MN401411 | — | MN423175 | [3] |
| G. phyllanthicola | L4948 | Yunnan, SW China | PP869245 | — | — | [37] |
| G. phyllanthicola | HL308 | Yunnan, SW China | PP869246 | — | [37] | |
| G. platense | BAFC384 | Argentina | AH008109 | — | — | [56] |
| G. platense | BAFC2374 | Argentina | AH008110 | — | — | [56] |
| G. podocarpense | QCAM 6422 | Ecuador | MF796661 | — | — | [55] |
| G. podocarpense | JV 1504/126 | — | MZ354942 | MZ345737 | MZ221687 | [2] |
| G. polychromum | UMNOR3 | USA | MG654204 | — | MG754744 | [53] |
| G. polychromum | MS343OR | USA | MG654197 | — | MG754743 | [53] |
| G. puerense | Dai 20427 | Yunnan, SW China | MZ345738 | MZ221688 | [2] | |
| G. ramosissium | xsd08085 | — | FJ478127 | — | — | [2] |
| G. ramosissium | xsd08032 | — | EU918700 | — | — | [2] |
| G. ravenelii | MS187FL | USA | MG654211 | MG754865 | MG754745 | [53] |
| G. ravenelii | 151FL | USA | MG654208 | — | — | [53] |
| G. resinaceum | BCRC 36147 | Netherlands | KJ143916 | — | KJ143934 | [40] |
| G. resinaceum | BR 4150 | France | KJ143915 | — | — | [40] |
| G. ryvardenii | HKAS58053 | Cameroon | HM138671 | — | — | [57] |
| G. ryvardenii | HKAS 58054 | Cameroon | HM138672 | — | — | [57] |
| G. sanduense | SA18012501 | Guizhou, SW China | MK345450 | — | — | [58] |
| G. sanduense | L4906 | Yunnan, SW China | ON994251 | OP508430 | OP508444 | [48] |
| G. sessile | JV 1209/9 | USA | KF605629 | — | KJ143936 | [40] |
| G. sessile | V 1209/27 | USA | KF605630 | KJ143976 | KJ143937 | [40] |
| G. shanxiense | BJTC FM423 | Shanxi, northern China | MK764268 | MK783940 | MK783937 | [59] |
| G. shanxiense | HSA 539 | Shanxi, northern China | MK764269 | MK789681 | — | [59] |
| G. shennongii | N.K. Zeng203 (FHMU2290) | Hainan, southern China | PP663109 | — | — | This study |
| G. sichuanense | HMAS 42798 | Sichuan, SW China | JQ781877 | — | — | [11] |
| G. sichuanense | Cui 7691 | Guangdong, southern China | JQ781878 | — | — | [11] |
| G. sinense | Cui 14526 | Guangxi, southern China | MZ354961 | MZ345743 | MZ221694 | [2] |
| G. sinense | Cui 14461 | China | MZ354963 | MZ345744 | MZ221695 | [2] |
| G. steyaertanum | II-121-1 | Indonesia | KJ654427 | — | — | [60] |
| G. steyaertanum | 6-WN-15(M)-A | Indonesia | KJ654459 | — | — | [60] |
| G. suae | L4651 | Yunnan, SW China | PP869243 | PP894784 | PP894782 | [37] |
| G. suae | L4817 | Yunnan, SW China | PP869244 | — | PP894783 | [37] |
| G. subangustisporum | Cui 18592 | Yunnan, SW China | MZ354981 | — | MZ221697 | [2] |
| G. subangustisporum | Cui 18593 | Yunnan, SW China | MZ354982 | — | MZ221698 | [2] |
| G. subellipsoideum | Cui 18241 | Malaysia | — | — | MZ221701 | [2] |
| G. subellipsoideum | Cui 18325 | Malaysia | — | — | MZ221702 | [2] |
| G. subflexipes | HKAS81926-3 | Fujian, SE China | PP465553 | — | PP922169 | [61], this study |
| G. subflexipes | N.K. Zeng1893-2 (FHMU7611) | Hainan, southern China | — | — | PP922171 | This study |
| G. subflexipes | HKAS80249 | Fujian, SE China | — | — | PP922168 | This study |
| G. subflexipes | N.K. Zeng4086 (FHMU3731) | Guangdong, southern China | — | — | PP922165 | This study |
| G. subflexipes | N.K. Zeng1455 (FHMU2320) | Fujian, SE China | PP465552 | PP785030 | PP922167 | [61], this study |
| G. subflexipes | N.K. Zeng4114 (FHMU5725) | Guangdong, southern China | PP465551 | — | PP922164 | [61], this study |
| G. subflexipes | HKAS79603 | Guangdong, southern China | PP465550 | — | PP922166 | [61], this study |
| G. subflexipes | HKAS81926-1 | Hainan, southern China | PP465549 | — | PP922163 | [61], this study |
| G. subflexipes | Cui 17257 | Guangdong, southern China | MZ354922 | MZ245396 | MZ221646 | [2] |
| G. sublobatum | Cui 16804 | Australia | MZ354973 | MZ345747 | MZ221704 | [2] |
| G. sublobatum | Cui 16806 | Australia | MZ354974 | — | MZ221705 | [2] |
| G. thailandicum | HKAS 104640 | Thailand | MK848681 | MK875831 | MK875829 | [62] |
| G. thailandicum | HKAS 104641 | Thailand | MK848682 | MK875832 | MK875830 | [62] |
| G. tongshanense | Cui 17168 | Hubei, central China | MZ354975 | — | MZ221706 | [2] |
| G. tropicum | BCRC 37122 | Taiwan, SE China | EU021457 | — | — | [63] |
| G. tropicum | GACP1408 1518 | Hainan, southern China | MH106884 | — | — | [5] |
| G. tsugae | UMNMI20 | USA | MG654324 | MG754764 | [64] | |
| G. tsugae | UMNMI30 | USA | MG654326 | MG754871 | MH025362 | [64] |
| G. tuberculosum | GVL-21 | Mexico | MT232639 | — | — | [65] |
| G. tuberculosum | GVL-40 | Mexico | MT232634 | — | — | [65] |
| G. weberianum | GanoTK17 | Cameroon | JN105705 | — | — | [66] |
| G. weixiensis | YL02 | Yunnan, SW China | MK302445 | — | MK302443 | [7] |
| G. weixiensis | YL01 | Yunnan, SW China | MK302444 | — | MK302442 | [7] |
| G. wiiroense | UMN-21-GHA | Ghana | KT952363 | — | — | [67] |
| G. wiiroense | UMN-20-GHA | Ghana | KT952361 | — | — | [67] |
| G. williamsianum | Dai 17790 | Singapore | MZ354947 | — | — | [2] |
| G. williamsianum | Dai 16809 | Thailand | MG279183 | MG367535 | MG367588 | [6] |
| G. yunlingense | Cui 16288 | Yunnan, SW China | MZ354915 | — | MZ221718 | [2] |
| G. yunlingense | Cui 17043 | Yunnan, SW China | MZ354916 | — | MZ221719 | [2] |
| G. yunnanense | HL45 | Yunnan, SW China | ON994235 | OP508422 | OP508436 | [48] |
| G. yunnanense | L4812 | Yunnan, SW China | ON994236 | OP508429 | OP508443 | [48] |
| G. zonatum | FL-02 | USA | KJ143921 | KJ143979 | KJ143941 | [40] |
| G. zonatum | FL-03 | USA | KJ143922 | KJ143980 | KJ143942 | [40] |
2.5. Phylogenetic Analyses
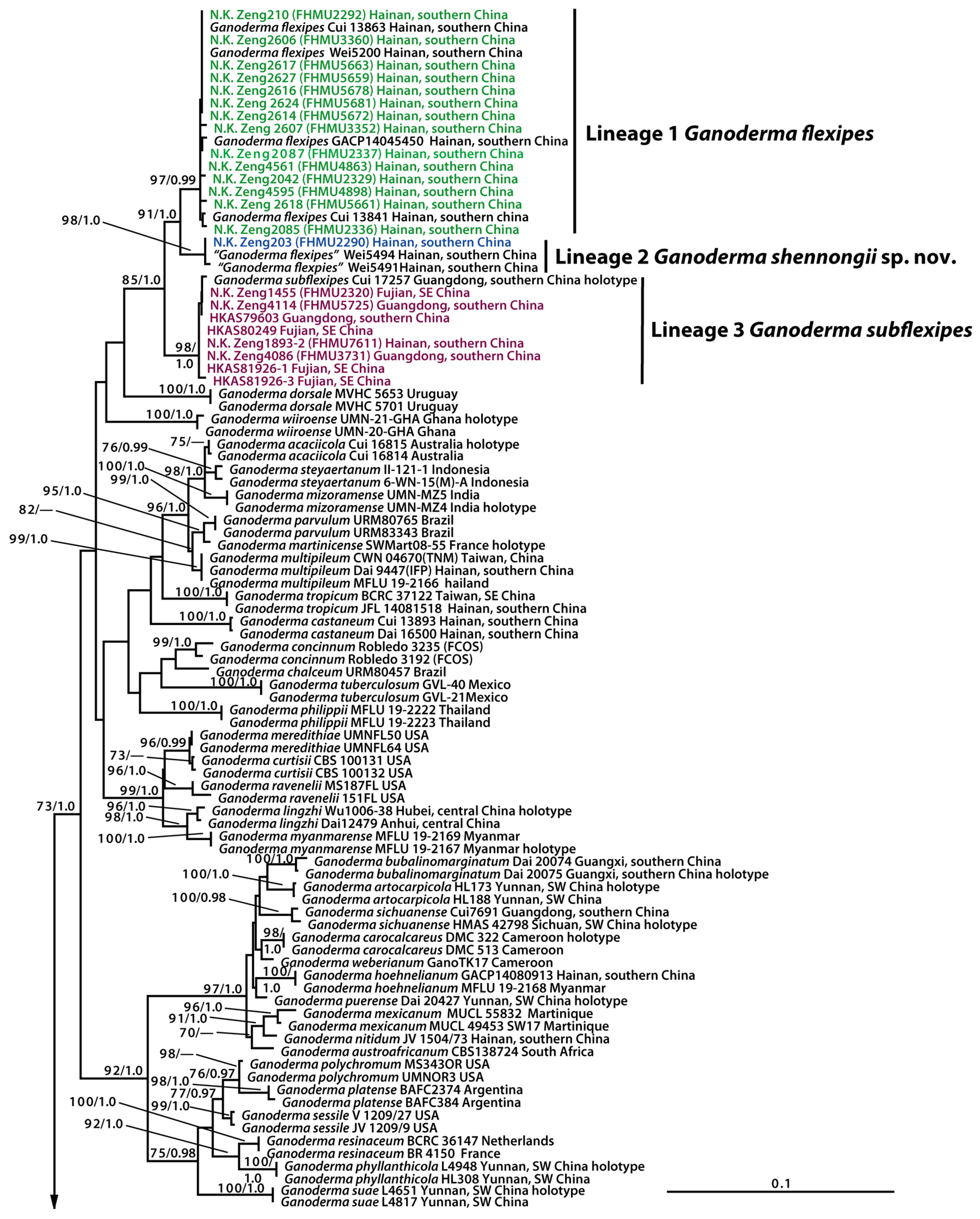
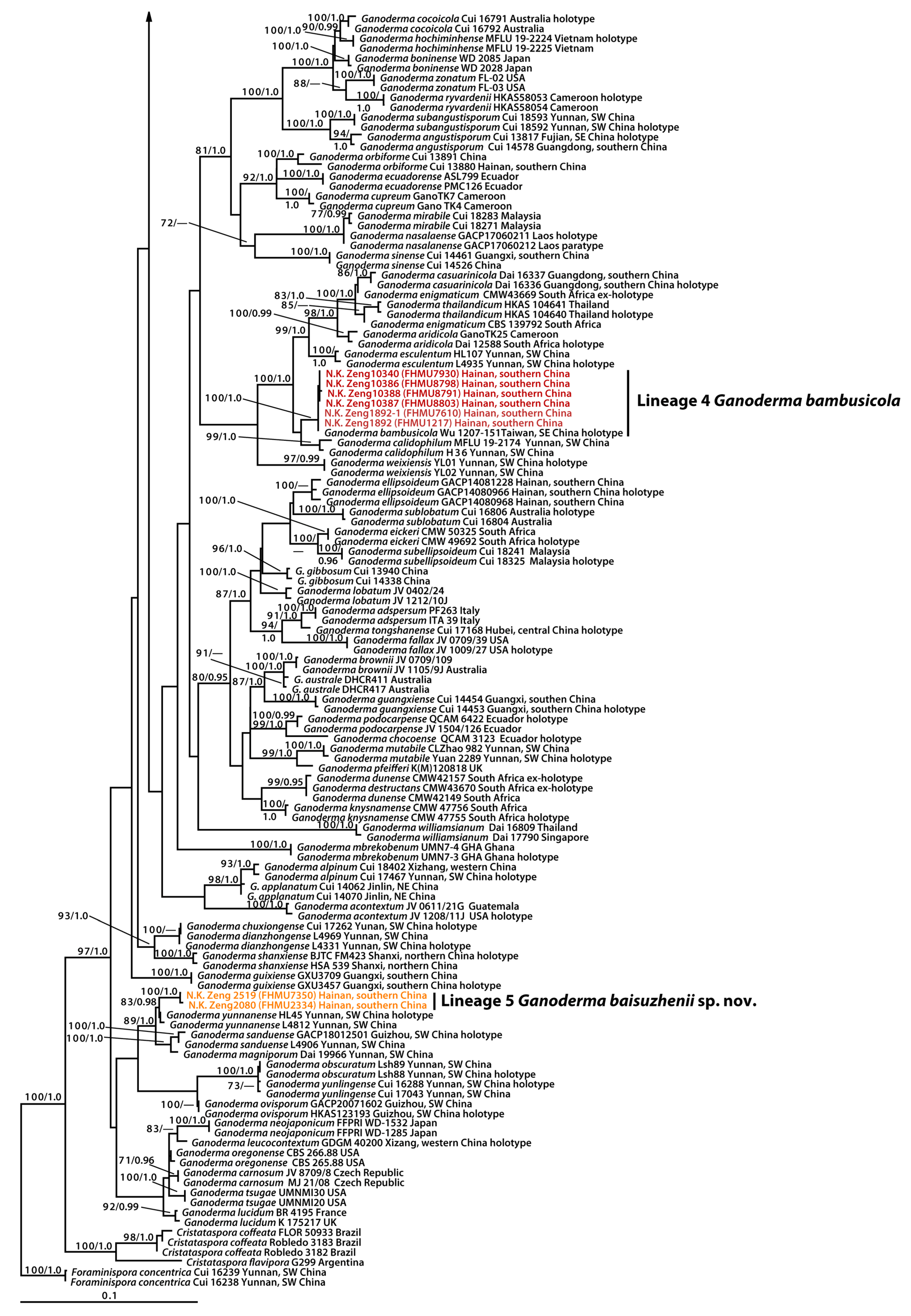
3. Results
3.1. Molecular Data
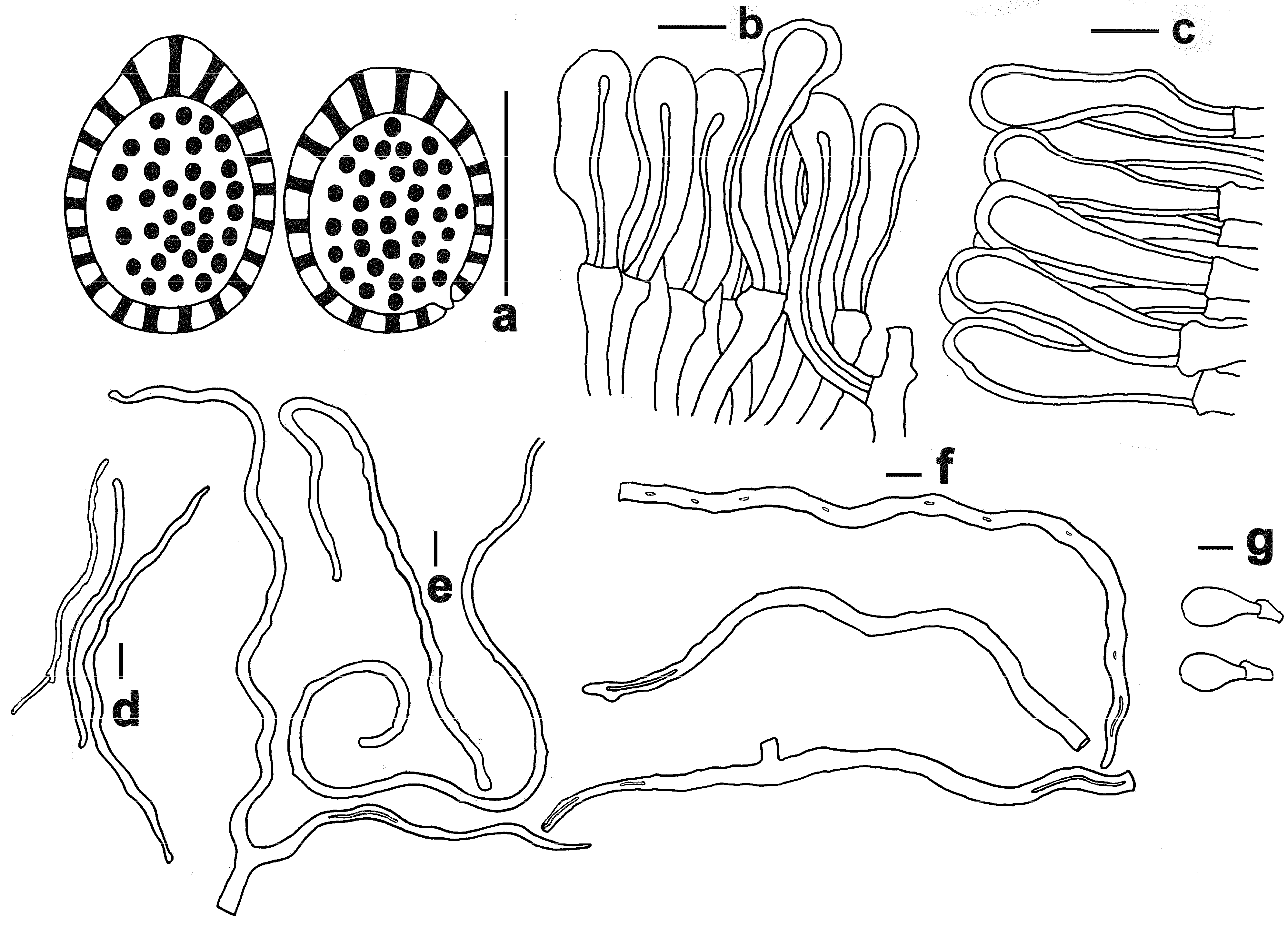
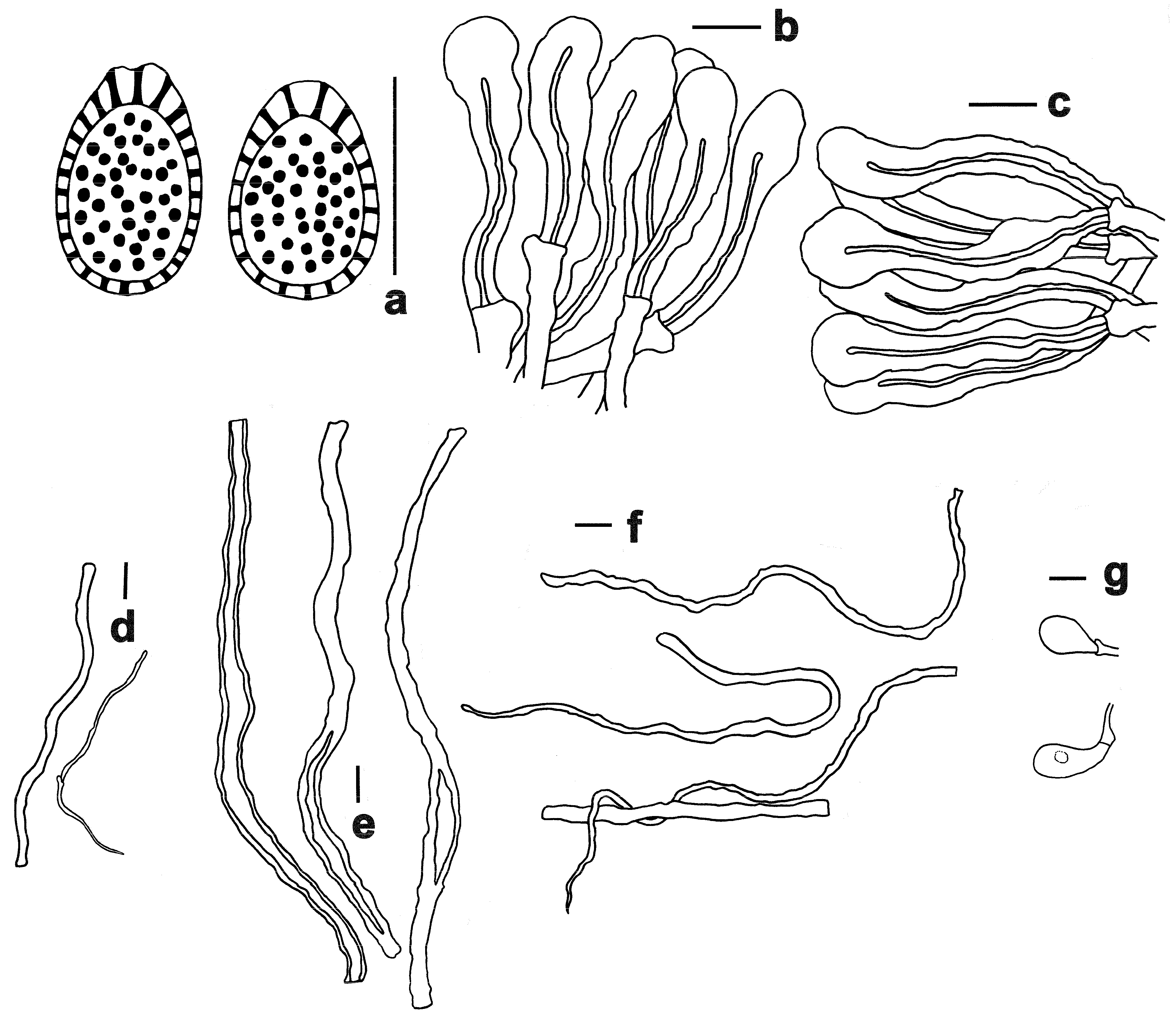
3.2. Taxonomy
4. Discussion
5. Conclusions
| Key to five Ganoderma species called “Lingzhi Wang” or “Zhu Lingzhi” in Hainan Island, tropical China | |
| 1. Host is bamboo | 2 |
| 1. Host is hardwood (especially Fagaceae trees) | 3 |
| 2. Pileus color is brownish-black, reddish-black to purplish-brown; basidiospore size 9.5–12.5 × 5–7 μm | G. bambusicola |
| 2. Pileus color is reddish-brown to orangish-brown; basidiospore size 8–11.5 × 5–7.5 μm | G. subflexipes |
| 3. Pileus context is nearly white; pore density up to 3 per mm; basidiospore up to 14 μm in length and 10 μm in width | G. baisuzhenii |
| 3. Pileus context is brown, pore density up to 5 per mm; basidiospore up to 12 μm in length and 8.5 μm in width | 4 |
| 4. Pileus color is brownish-red to reddish; pore density 3–4 per mm; basidiospore size 8.5–12 × 5.5–7.5 μm | G. flexipes |
| 4. Pileus color is dark reddish; pore density 4–5 per mm; basidiospore size 10–11.5 × 7–8.5 μm | G. shennongii |
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- He, J.; Luo, Z.L.; Tang, S.M.; Li, Y.J.; Li, S.H.; Su, H.Y. Phylogenetic analyses and morphological characters reveal two new species of Ganoderma from Yunnan province, China. MycoKeys 2021, 84, 141. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.F.; Xing, J.H.; He, X.L.; Wu, D.M.; Song, C.G.; Liu, S.; Vlasák, J.; Gates, G.; Gibertoni, T.B.; Cui, B.K. Species diversity, systematic revision and molecular phylogeny of Ganodermataceae (Polyporales, Basidiomycota) with an emphasis on Chinese collections. Stud. Mycol. 2022, 101, 287–415. [Google Scholar] [CrossRef]
- Luangharn, T.; Karunarathna, S.C.; Dutta, A.K.; Paloi, S.; Promputtha, I.; Hyde, K.D.; Xu, J.; Mortimer, P.E. Ganoderma (Ganodermataceae, Basidiomycota) species from the greater Mekong subregion. J. Fungi 2021, 7, 819. [Google Scholar] [CrossRef] [PubMed]
- Kirk, P.M.; Cannon, P.F.; Minter, D.; Stalpers, J.A. Dictionary of the Fungi, 10th ed.; CAB International: Wallingford, UK, 2008. [Google Scholar]
- Hapuarachchi, K.K.; Karunarathna, S.C.; Raspé, O.; De Silva, K.H.W.L.; Thawthong, A.; Wu, X.L.; Kakumyan, P.; Hyde, K.D.; Wen, T.C. High diversity of Ganoderma and Amauroderma (Ganodermataceae, Polyporales) in Hainan Island, China. Mycosphere 2018, 9, 931–982. [Google Scholar] [CrossRef]
- Xing, J.H.; Sun, Y.F.; Han, Y.L.; Cui, B.K.; Dai, Y.C. Morphological and molecular identification of two new Ganoderma species on Casuarina equisetifolia from China. MycoKeys 2018, 34, 93–108. [Google Scholar] [CrossRef]
- Ye, L.; Karunarathna, S.C.; Mortimer, P.E.; Li, H.; Qiu, M.; Peng, X.; Luangharn, T.; Li, Y.; Promputtha, I.; Hyde, K.; et al. Ganoderma weixiensis (Polyporaceae, Basidiomycota), a new member of the G. lucidum complex from Yunnan Province, China. Phytotaxa 2019, 423, 75–86. [Google Scholar] [CrossRef]
- Wu, S.H.; Chern, C.L.; Wei, C.L.; Chen, Y.P.; Akiba, M.; Hattori, T. Ganoderma bambusicola sp. nov. (Polyporales, Basidiomycota) from southern Asia. Phytotaxa 2020, 456, 75–85. [Google Scholar] [CrossRef]
- Yang, H.D.; Ding, Y.; Wen, T.C.; Hupuarachchi, K.K.; Wei, D.P. Ganoderma ovisporum sp. nov. (Polyporales, Polyporaceae) from Southwest China. Biodivers. Data J. 2022, 197, e80034. [Google Scholar] [CrossRef]
- Cui, B.K.; Pan, X.H.; Pan, F.; Sun, Y.F.; Xing, J.H.; Dai, Y.C. Species diversity and resources of Ganoderma in China. Mycosystema 2023, 42, 170–178. [Google Scholar] [CrossRef]
- Cao, Y.; Wu, S.H.; Dai, Y.C. Species clarification of the prize medicinal Ganoderma mushroom “Lingzhi. Fungal Divers. 2012, 56, 49–62. [Google Scholar] [CrossRef]
- Yao, Y.J.; Li, Y.; Du, Z.; Wang, K.; Wang, X.C.; Kirk, P.M.; Spooner, B.M. On the typification of Ganoderma sichuanense (Agaricomycetes)−the widely cultivated lingzhi medicinal mushroom. Int. J. Med. Mushrooms 2020, 22, 45–54. [Google Scholar] [CrossRef]
- Du, Z.; Li, Y.; Wang, X.C.; Wang, K.; Yao, Y.J. Re-examination of the holotype of Ganoderma sichuanense (Ganodermataceae, Polyporales) and a clarification of the identity of Chinese cultivated lingzhi. J. Fungi 2023, 9, 323. [Google Scholar] [CrossRef] [PubMed]
- Wagner, R.; Mitchell, D.A.; Sassaki, G.L.; Amazonas, M.A.L.A.; Berovič, M. Current techniques for the cultivation of Ganoderma lucidum for the production of biomass, ganoderic acid and polysaccharides. Food Technol. Biotech. 2003, 41, 371–382. Available online: https://hrcak.srce.hr/125310 (accessed on 13 March 2025).
- Wu, X.L.; Mao, X.L.; Tuli, G.E.; Song, B.; Li, T.H. Medicinal Fungi of China; Science Press: Beijing, China, 2013. [Google Scholar]
- Yuan, Y.; Wang, Y.J.; Sun, G.P.; Wang, Y.R.; Cao, L.J.; Shen, Y.M.; Yuan, B.; Han, D.; Huang, L. Archaeological evidence suggests earlier use of Ganoderma in Neolithic China. Chin. Sci. Bull. 2018, 63, 1180–1188. [Google Scholar] [CrossRef]
- Lin, Z.; Sun, L. Antitumor effect of Ganoderma (Lingzhi) mediated by immunological mechanism and its clinical application. In Ganoderma and Health; Springer: Berlin/Heidelberg, Germany, 2019; pp. 39–77. [Google Scholar] [CrossRef]
- Ren, L.; Zhang, J.; Zhang, T. Immunomodulatory activities of polysaccharides from Ganoderma on immune effector cells. Food Chem. 2021, 340, 127933. [Google Scholar] [CrossRef]
- Chen, S.; Guan, X.; Yong, T.; Gao, X.; Xiao, C.; Xie, Y.; Chen, D.; Hu, H.; Wu, Q. Structural characterization and hepatoprotective activity of an acidic polysaccharide from Ganoderma lucidum. Food Chem. 2022, 13, 100204. [Google Scholar] [CrossRef]
- Li, B.Y.; Wu, X.L.; Song, B.; Li, T.H.; Zhong, J.X.; Chen, H.Q. Distribution of the medicinal Ganoderma ssp. in Hainan. Guangdong Agric. Sci. 2013, 3, 171–172. [Google Scholar] [CrossRef]
- Zhao, J.D. The Ganodermataceae in China. Bibl. Mycol. 1989, 132, 1–176. [Google Scholar]
- Zhong, J.X.; Guo, J.R.; Xiao, M.; Chen, H.Q.; He, S.Z.; Zou, F.L.; Wu, X.L. Study on the medicinal ganoderma resources in Hainan Island. Chin. Pharm. J. 1998, 33, 652–655. [Google Scholar]
- Zeng, N.K.; Jiang, S. Atlas of Macrofungi from Yinggeling of Hainan, China; South Sea Publishing Company: Haikou, China, 2020. [Google Scholar]
- Kornerup, A.; Wanscher, J.H. Taschenlexikon der Farben. 3. Aufl.; Muster-Schmidt Verlag: Göttingen, Germany, 1981. [Google Scholar]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelflfland, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: New York, NY, USA, 1990; pp. 315–322. [Google Scholar] [CrossRef]
- Matheny, P.B. Improving phylogenetic inference of mushrooms with RPB1 and RPB2 nucleotide sequences (Inocybe; Agaricales). Mol. Phylogenet. Evol. 2005, 35, 1–20. [Google Scholar] [CrossRef]
- Zeng, N.K.; Tang, L.P.; Li, Y.C.; Tolgor, B.; Zhu, X.T.; Zhao, Q.; Yang, Z.L. The genus Phylloporus (Boletaceae, Boletales) from China: Morphological and multilocus DNA sequence analyses. Fungal Divers. 2013, 58, 73–101. [Google Scholar] [CrossRef]
- Zeng, N.K.; Chai, H.; Jiang, S.; Xue, R.; Wang, Y.; Hong, D.; Liang, Z.Q. Retiboletus nigrogriseus and Tengioboletus fujianensis, two new boletes from the south of China. Phytotaxa 2018, 367, 45–54. [Google Scholar] [CrossRef]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analyses program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar] [CrossRef]
- Sun, Y.F.; Costa-Rezende, D.H.; Xing, J.H.; Zhou, J.L.; Zhang, B.; Gibertoni, T.B.; Gates, G.; Dai, Y.C.; Cui, B.K. Multi-gene phylogeny and taxonomy of Amauroderma s. lat. (Ganodermataceae). Persoonia 2020, 44, 206–239. [Google Scholar] [CrossRef]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef]
- Smith, S.A.; Dunn, C.W. Phyutility: A phyloinformatics tool for trees, alignments and molecular data. Bioinformation 2008, 24, 715–716. [Google Scholar] [CrossRef]
- Costa-Rezende, D.H.; Gugliotta, A.M.; Góes-Neto, A.; Reck, M.A.; Robledo, G.L.; Drechsler-Santos, E.R. Amauroderma calcitum sp. nov. and notes on taxonomy and distribution of Amauroderma species (Ganodermataceae). Phytotaxa 2016, 244, 101–124. [Google Scholar] [CrossRef]
- Costa-Rezende, D.H.; Robledo, G.L.; Drechsler-Santos, E.R.; Glen, M.; Gates, G.; de Madrignac Bonzi, B.R.; Popoff, O.F.; Crespo, E.; Góes-Neto, A. Taxonomy and phylogeny of polypores with ganodermatoid basidiospores (Ganodermataceae). Mycol. Prog. 2020, 19, 725–741. [Google Scholar] [CrossRef]
- Terho, M.; Hantula, J.; Hallaksela, A.M. Occurrence and decay patterns of common wood-decay fungi in hazardous trees felled in the Helsinki City. Forest Pathol. 2007, 37, 420–432. [Google Scholar] [CrossRef]
- Xing, J.; Song, J.; Decock, C.; Cui, B.K. Morphological characters and phylogenetic analysis reveal a new species within the Ganoderma lucidum complex from South Africa. Phytotaxa 2016, 266, 115–124. [Google Scholar] [CrossRef]
- He, J.; Li, X.J.; Tan, W.Z.; Wu, X.Q.; Wu, D.; Luo, Z.L.; Zhou, Q.W.; Li, E.X.; Li, S.H. Two new species of Ganoderma (Ganodermataceae, Basidiomycota) from Southwest China. MycoKeys 2024, 106, 97–116. [Google Scholar] [CrossRef]
- Costa-Rezende, D.H.; Robledo, G.L.; Góes-Neto, A.; Reck, M.A.; Crespo, E.; Drechsler-Santos, E.R. Morphological reassessment and molecular phylogenetic analyses of Amauroderma s. lat. raised new perspectives in the generic classification of the Ganodermataceae family. Persoonia 2017, 39, 254–269. [Google Scholar] [CrossRef] [PubMed]
- Coetzee, M.P.; Marincowitz, S.; Muthelo, V.G.; Wingfield, M.J. Ganoderma species, including new taxa associated with root rot of the iconic Jacaranda mimosifolia in Pretoria, South Africa. IMA Fungus 2015, 6, 249–256. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.W.; Cao, Y.; Wu, S.H.; Vlasák, J.; Li, D.W.; Li, M.J.; Dai, Y.C. Global diversity of the Ganoderma lucidum complex (Ganodermataceae, Polyporales) inferred from morphology and multilocus phylogeny. Phytochemistry 2015, 114, 7–15. [Google Scholar] [CrossRef]
- Douanla-Meli, C.; Langer, E. Ganoderma carocalcareus sp. nov., with crumbly-friable context parasite to saprobe on Anthocleista nobilis and its phylogenetic relationship in G. resinaceum group. Mycol. Prog. 2009, 8, 145–155. [Google Scholar] [CrossRef]
- de Lima Júnior, N.C.; Gibertone, T.B.; Malosso, E. Delimitation of some neotropical laccate Ganoderma (Ganodermataceae): Molecular phylogeny and morphology. Rev. Biol. Trop. 2014, 62, 1197–1208. [Google Scholar] [CrossRef]
- Crous, P.W.; Luangsa-ard, J.J.; Wingfield, M.J.; Carnegie, A.J.; Hernández-Restrepo, M.; Lombard, L.; Roux, J.; Barreto, R.W.; Baseia, I.G.; Cano-Lira, J.F.; et al. Fungal planet description sheets: 785–867. Persoonia 2018, 41, 238–417. [Google Scholar] [CrossRef] [PubMed]
- Morera, G.; Lupo, S.; Alaniz, S.; Robledo, G. Diversity of the Ganoderma species in Uruguay. Neotrop. Biodivers. 2021, 7, 570–585. [Google Scholar] [CrossRef]
- Tchoumi, J.M.T.; Coetzee, M.P.A.; Rajchenberg, M.; Wingfield, M.J.; Roux, J. Three Ganoderma species, including Ganoderma dunense sp. nov., associated with dying Acacia cyclops trees in South Africa. Australas. Plant Pathol. 2018, 47, 431–447. [Google Scholar] [CrossRef]
- Crous, P.W.; Wingfield, M.J.; Richardson, D.M.; Leroux, J.J.; Strasberg, D.; Edwards, J.; Roets, F.; Hubka, V.; Taylor, P.W.J.; Heykoop, M.; et al. Fungal planet description sheets: 400–468. Persoonia 2016, 36, 316–458. [Google Scholar] [CrossRef]
- Tchoumi, J.M.T.; Coetzee, M.P.A.; Rajchenberg, M.; Roux, J. Taxonomy and species diversity of Ganoderma species in the Garden Route National Park of South Africa inferred from morphology and multilocus phylogenies. Mycologia 2019, 111, 730–747. [Google Scholar] [CrossRef]
- He, J.; Han, X.; Luo, Z.L.; Li, E.X.; Tang, S.M.; Luo, H.M.; Niu, K.Y.; Su, X.J.; Li, S.H. Species diversity of Ganoderma (Ganodermataceae, Polyporales) with three new species and a key to Ganoderma in Yunnan Province, China. Front. Microbiol. 2022, 13, 1035434. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.; Yuan, H.S. Ganoderma mutabile sp. nov. from southwestern China based on morphological and molecular data. Mycol. Prog. 2013, 12, 121–126. [Google Scholar] [CrossRef]
- Wei, Q.L.; Zheng, H.F.; Shao, Y.Y.; Rasheed, U.; Lin, J.T.; Huang, F.C.; Liu, B. A new species of Ganoderma (Ganodermataceae, Polyporales) from southern China and optimum condition for mycelia production. Mycobiology 2024, 52, 58–67. [Google Scholar] [CrossRef] [PubMed]
- Li, T.H.; Hu, H.P.; Deng, W.Q.; Wu, S.H.; Wang, D.M. Ganoderma leucocontextum, a new member of the G. lucidum complex from southwestern China. Mycoscience 2015, 56, 81–85. [Google Scholar] [CrossRef]
- Welti, S.; Courtecuisse, R. The Ganodermataceae in the French West Indies (Guadeloupe and Martinique). Fungal Divers. 2010, 43, 103–126. [Google Scholar] [CrossRef]
- Loyd, A.L.; Held, B.W.; Linder, E.R.; Smith, J.A.; Blanchette, R.A. Elucidating wood decomposition by four species of Ganoderma from the United States. Fungal Biol. 2018, 122, 254–263. [Google Scholar] [CrossRef] [PubMed]
- Cabarroi-Hernández, M.; Villalobos-Arámbula, A.R.; Torres-Torres, M.G.; Decock, C.; Guzmán-Dávalos, L. The Ganoderma weberianum-resinaceum lineage: Multilocus phylogenetic analysis and morphology confirm G. mexicanum and G. parvulum in the Neotropics. MycoKeys 2019, 59, 95–131. [Google Scholar] [CrossRef]
- Crous, P.W.; Wingfield, M.J.; Burgess, T.I.; Hardy, G.E.S.T.J.; Barber, P.A.; Alvarado, P.; Barnes, C.W.; Buchanan, P.K.; Heykoop, M.; Moreno, G.; et al. Fungal planet description sheets: 558–624. Persoonia 2017, 38, 240. [Google Scholar] [CrossRef]
- Gottlieb, A.M.; Ferrer, E.; Wright, J.E. rDNA analyses as an aid to the taxonomy of species of Ganoderma. Mycol. Res. 2000, 104, 1033–1045. [Google Scholar] [CrossRef]
- Kinge, T.R.; Mih, A.M. Ganoderma ryvardense sp. nov. associated with basal stem rot (BSR) disease of oil palm in Cameroon. Mycosphere 2011, 2, 179–188. [Google Scholar]
- Hapuarachchi, K.K.; Karunarathna, S.C.; Phengsintham, P.; Yang, H.D.; Kakumyan, P.; Hyde, K.D.; Wen, T.C. Ganodermataceae (Polyporales): Diversity in Greater Mekong Subregion countries (China, Laos, Myanmar, Thailand and Vietnam). Mycosphere 2019, 10, 221–309. [Google Scholar] [CrossRef]
- Liu, H.; Guo, L.J.; Li, S.L.; Fan, L. Ganoderma shanxiense, a new species from northern China based on morphological and molecular evidence. Phytotaxa 2019, 406, 129–136. [Google Scholar] [CrossRef]
- Glen, M.; Bougher, N.L.; Francis, A.A.; Nigg, S.Q.; Lee, S.S.; Irianto, R.; Barry, K.M.; Beadle, C.L.; Mohammed, C.L. Ganoderma and Amauroderma species associated with root-rot disease of Acacia mangium plantation trees in Indonesia and Malaysia. Australas. Plant Pathol. 2009, 38, 345–356. [Google Scholar] [CrossRef]
- Tian, R.; Chen, L.F.; Zhao, R.X.; Zeng, N.K. Biological characteristics and cultivation of Ganoderma subflexipes. Mycosystema 2024, 1–22. [Google Scholar] [CrossRef]
- Luangharn, T.; Karunarathna, S.C.; Mortimer, P.E.; Hyde, K.D.; Xu, J. Additions to the knowledge of Ganoderma in Thailand: Ganoderma casuarinicola, a new record; and Ganoderma thailandicum sp. nov. MycoKeys 2019, 59, 47. [Google Scholar] [CrossRef]
- Wang, D.M.; Wu, S.H.; Su, C.H.; Peng, J.T.; Shih, Y.H.; Chen, L.C. Ganoderma multipileum, the correct name for ‘G. lucidum’ in tropical Asia. Bot. Stud. 2009, 50, 451–458. [Google Scholar]
- Loyd, A.L.; Barnes, C.W.; Held, B.W.; Schink, M.J.; Smith, M.E.; Smith, J.A.; Blanchette, R.A. Elucidating “lucidum”: Distinguishing the diverse laccate Ganoderma species of the United States. PLoS ONE 2018, 13, e0199738. [Google Scholar] [CrossRef]
- Espinosa-García, V.; Mendoza, G.; Shnyreva, V.A.; Padrón, J.M.; Trigos, Á. Biological activities of different strains of the genus Ganoderma spp. (Agaricomycetes) from Mexico. Int. J. Med. Mushrooms 2021, 23, 67–77. Available online: https://pubmed.ncbi.nlm.nih.gov/33639082/ (accessed on 13 March 2025). [CrossRef]
- Park, Y.J.; Kwon, O.C.; Son, E.S.; Yoon, D.E.; Han, W.; Nam, J.Y.; Yoo, Y.B.; Lee, C.S. Genetic diversity analysis of Ganoderma species and development of a specific marker for identification of medicinal mushroom Ganoderma lucidum. Afr. J. Microbiol. Res. 2012, 25, 5417–5425. [Google Scholar] [CrossRef]
- Crous, P.W.; Wingfield, M.J.; Le Roux, J.J.; Richardson, D.M.; Strasberg, D.; Shivas, R.G.; Alvarado, P.; Edwards, J.; Moreno, G.; Sharma, R.; et al. Fungal planet description sheets: 371–399. Persoonia 2015, 35, 264–327. [Google Scholar] [CrossRef]
- Stamatakis, A. RAxML-VI-HPC: Maximum likelihood-based phylogenetic analyses with thousands of taxa and mixed models. Bioinformatics 2006, 22, 2688–2690. [Google Scholar] [CrossRef] [PubMed]
- Nylander, J.A.A. MrModeltest 2.3. Program Distributed by the Author; Evolutionary Biology Center, Uppsala University: Uppsala, Sweden, 2004. [Google Scholar]
- Huelsenbeck, J.P.; Ronquist, F. Bayesian Analysis of Molecular Evolution Using MrBayes; Springer: New York, NY, USA, 2005; pp. 183–226. [Google Scholar] [CrossRef]
- Zhao, J.D.; Xu, L.W.; Zhang, X.Q. Taxonomic studies on the Ganodermataceae of China III. Acta Mycol. Sin. 1984, 3, 15–23. [Google Scholar]
- Wang, D.M.; Wu, S.H. Two species of Ganoderma new to Taiwan. Mycotaxon 2007, 102, 373–378. [Google Scholar]
- Patouillard, N. Champignons du Kouytcheou. Monde Plants 1907, 2, 31–32. [Google Scholar]
- Steyaert, R.L. Species of Ganoderma and related genera mainly of the Bogor and Leiden Herbaria. Persoonia 1972, 7, 55–118. [Google Scholar]
- Zhao, J.D.; Zhang, X.Q. Flora Fungorum Sinicorum Vol. 18: Ganodermataceae; Science Press: Beijing, China, 2000. [Google Scholar]
- Wang, J.C.; Liang, W.; Yan, Y.H.; Zeng, N.K. A Field Guide to the Priority Protected Species in Hainan Tropical Rainforest National Park; China Environment Publishing Group: Beijing, China, 2023. [Google Scholar]
- Xiao, F.R.; Liang, W.; Yan, Y.H.; Zeng, N.K.; Hao, X.; Wang, J.C. The checklist of priority protected species in Hainan Tropical Rainforest National Park and its application in national park managemen. Natl. Park 2024, 2, 43–55. [Google Scholar] [CrossRef]
- Chen, S.; Pang, X.; Song, J.; Shi, L.; Yao, H.; Han, J.; Leon, C.A. Renaissance in herbal medicine identification: From morphology to DNA. Biotechnol. Adv. 2014, 32, 1237–1244. [Google Scholar] [CrossRef]
- Zhao, J.D.; Xu, L.W.; Zhang, X.Q. Taxonomic studies on the subfamily Ganodermoideae of China. Acta Mycol. Sin. 1979, 19, 265–279. [Google Scholar]
- Zhao, J.D.; Zhang, X.Q.; Xu, L.W. Studies on the taxonomy of Ganodermataceae in China IV. Acta Mycol. Sin. 1986, 5, 86–93. [Google Scholar]
- Hapuarachchi, K.K.; Karunarathna, S.C.; McKenzie, E.H.C.; Wu, X.L.; Kakumyan, P.; Hyde, K.D.; Wen, T.C. High phenotypic plasticity of Ganoderma sinense (Ganodermataceae, Polyporales) in China. Asian J. Mycol. 2019, 2, 1–47. [Google Scholar] [CrossRef]
- Gilbertson, R.L.; Ryvarden, L. North American Polypores, Volume 1, Abortiporus–Lindtneria. J. Basic Microbiol. 1986, 27, 282. [Google Scholar]



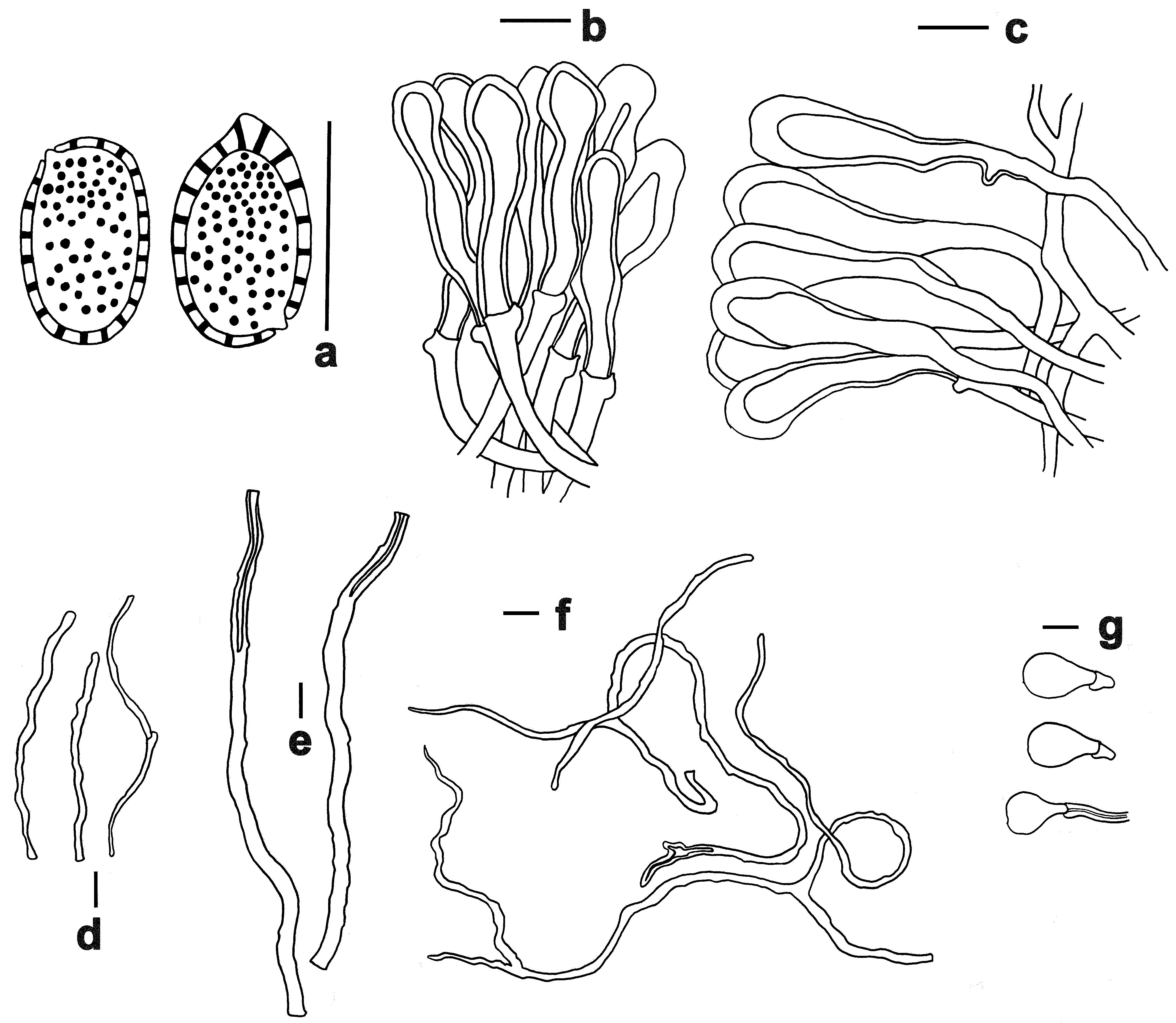

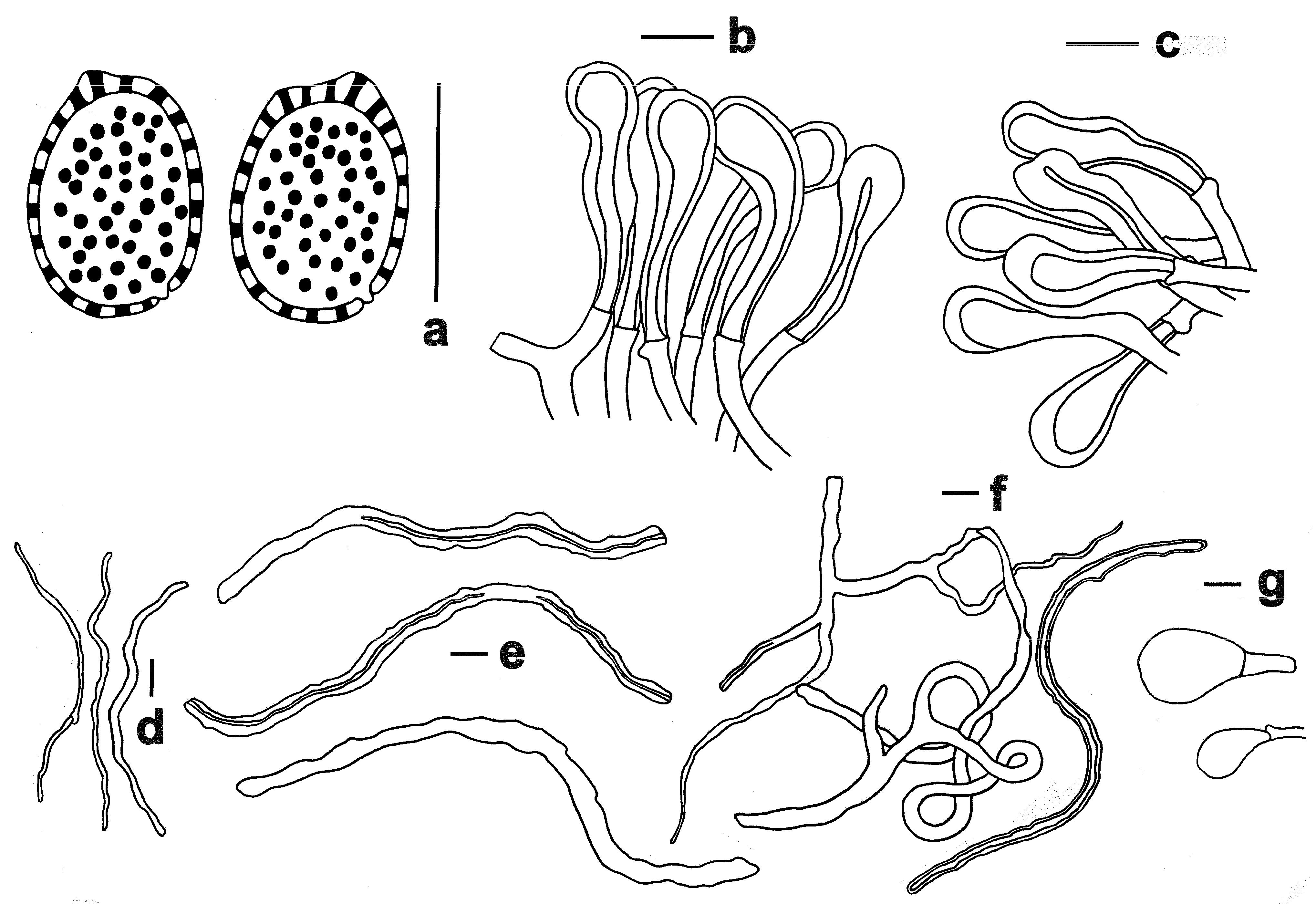

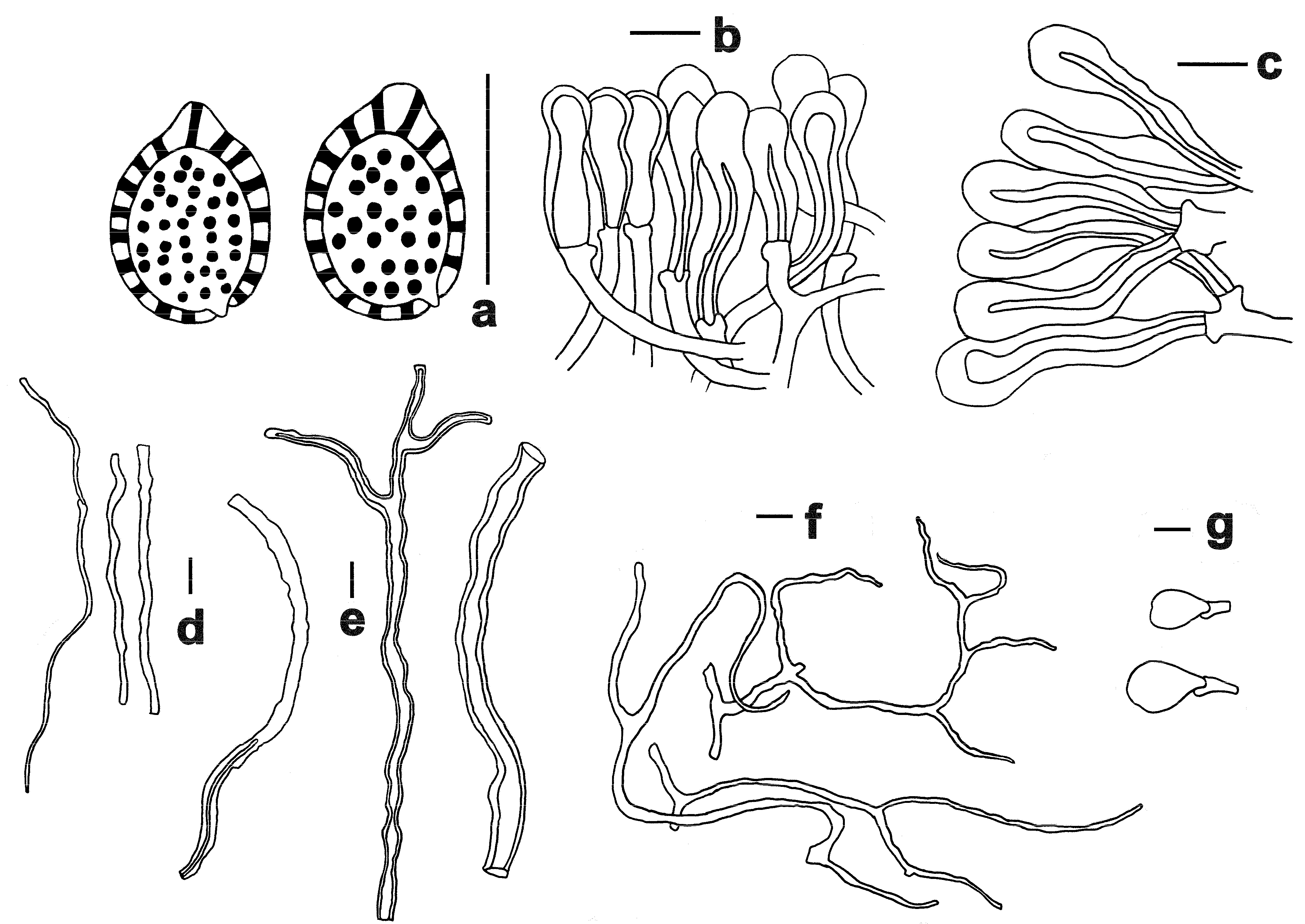

| Species | Local Name | Pilei Color | Pore | Context | Basidiospore (μm) | Host |
|---|---|---|---|---|---|---|
| Ganoderma baisuzhenii | Lingzhi Wang | Brownish-red to dark brownish-red | 2–3 per mm; surface yellowish when young, then white | Nearly white | 11.5–14 × 8–10 | On decaying hardwood (often underground) of fagaceous trees, particularly those of the genus Cyclobalanopsis |
| G. bambusicola | Zhu Lingzhi | Brownish-black, reddish-black to purplish-brown | 4–6 per mm; surface white | Dark brown | 9.5–12.5 × 5–7 | On dead roots of bamboo, particularly those of Dendrocalamus latiflorus |
| G. flexipes | Lingzhi Wang | Brownish-red to reddish | 3–4 per mm; surface white | Upper layer yellowish-brown, lower layer brown to dark brown | 8.5–12 × 5.5–7.5 | On decaying hardwood (often underground) of fagaceous trees, particularly those of Quercus patelliformis |
| G. shennongii | Lingzhi Wang | Dark reddish | 4–5 per mm; surface white | Yellowish-brown to brown | 10–11.5 × 7–8.5 | On decaying hardwood (often underground) in forests predominantly composed of fagaceous trees |
| G. subflexipes | Zhu Lingzhi | Reddish-brown to orangish-brown | 5–6 per mm; surface white | Yellowish-brown to brown | 8–11.5 × 5–7.5 | On dead roots of bamboo, particularly those of Bambusa chungii, sometimes on decaying hardwood (often underground) in forests dominated by fagaceous trees, or mixed forests dominated by fagaceous trees and Pinus massoniana |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tian, R.; Qin, H.-Z.; Zhong, Q.; Liang, Z.-Q.; Zhang, X.-Y.; Mu, X.-D.; Xu, L.; Wen, T.-C.; Chen, X.-D.; Zeng, N.-K. What Are “Lingzhi Wang” or “Zhu Lingzhi”? Notes on Ganoderma (Ganodermataceae, Polyporales) Species Characterized by Diminutive Pilei and Gracile Stipes from Hainan Island, Tropical China. J. Fungi 2025, 11, 237. https://doi.org/10.3390/jof11030237
Tian R, Qin H-Z, Zhong Q, Liang Z-Q, Zhang X-Y, Mu X-D, Xu L, Wen T-C, Chen X-D, Zeng N-K. What Are “Lingzhi Wang” or “Zhu Lingzhi”? Notes on Ganoderma (Ganodermataceae, Polyporales) Species Characterized by Diminutive Pilei and Gracile Stipes from Hainan Island, Tropical China. Journal of Fungi. 2025; 11(3):237. https://doi.org/10.3390/jof11030237
Chicago/Turabian StyleTian, Run, Hua-Zhi Qin, Qing Zhong, Zhi-Qun Liang, Xue-Yan Zhang, Xiao-Dong Mu, Lin Xu, Ting-Chi Wen, Xiang-Dong Chen, and Nian-Kai Zeng. 2025. "What Are “Lingzhi Wang” or “Zhu Lingzhi”? Notes on Ganoderma (Ganodermataceae, Polyporales) Species Characterized by Diminutive Pilei and Gracile Stipes from Hainan Island, Tropical China" Journal of Fungi 11, no. 3: 237. https://doi.org/10.3390/jof11030237
APA StyleTian, R., Qin, H.-Z., Zhong, Q., Liang, Z.-Q., Zhang, X.-Y., Mu, X.-D., Xu, L., Wen, T.-C., Chen, X.-D., & Zeng, N.-K. (2025). What Are “Lingzhi Wang” or “Zhu Lingzhi”? Notes on Ganoderma (Ganodermataceae, Polyporales) Species Characterized by Diminutive Pilei and Gracile Stipes from Hainan Island, Tropical China. Journal of Fungi, 11(3), 237. https://doi.org/10.3390/jof11030237